
Early Consumption of Cannabinoids: From Adult Neurogenesis to Behavior

 , , and
, , and
Abstract
:1. A Worldwide View of Cannabinoid Consumption
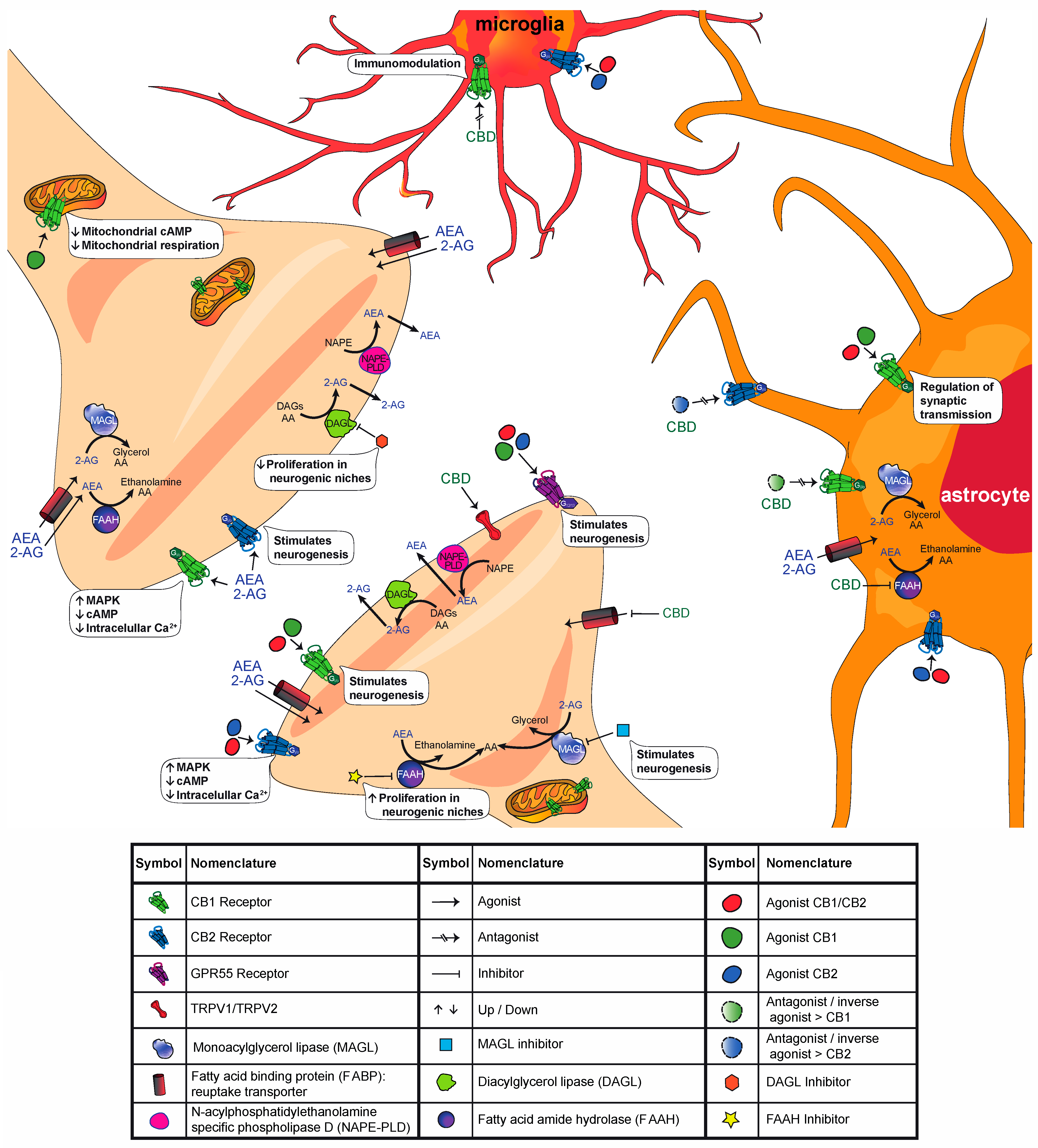
2. The Endocannabinoid System (ECS)
2.1. ECS in the Central Nervous System
2.2. ECS during Development
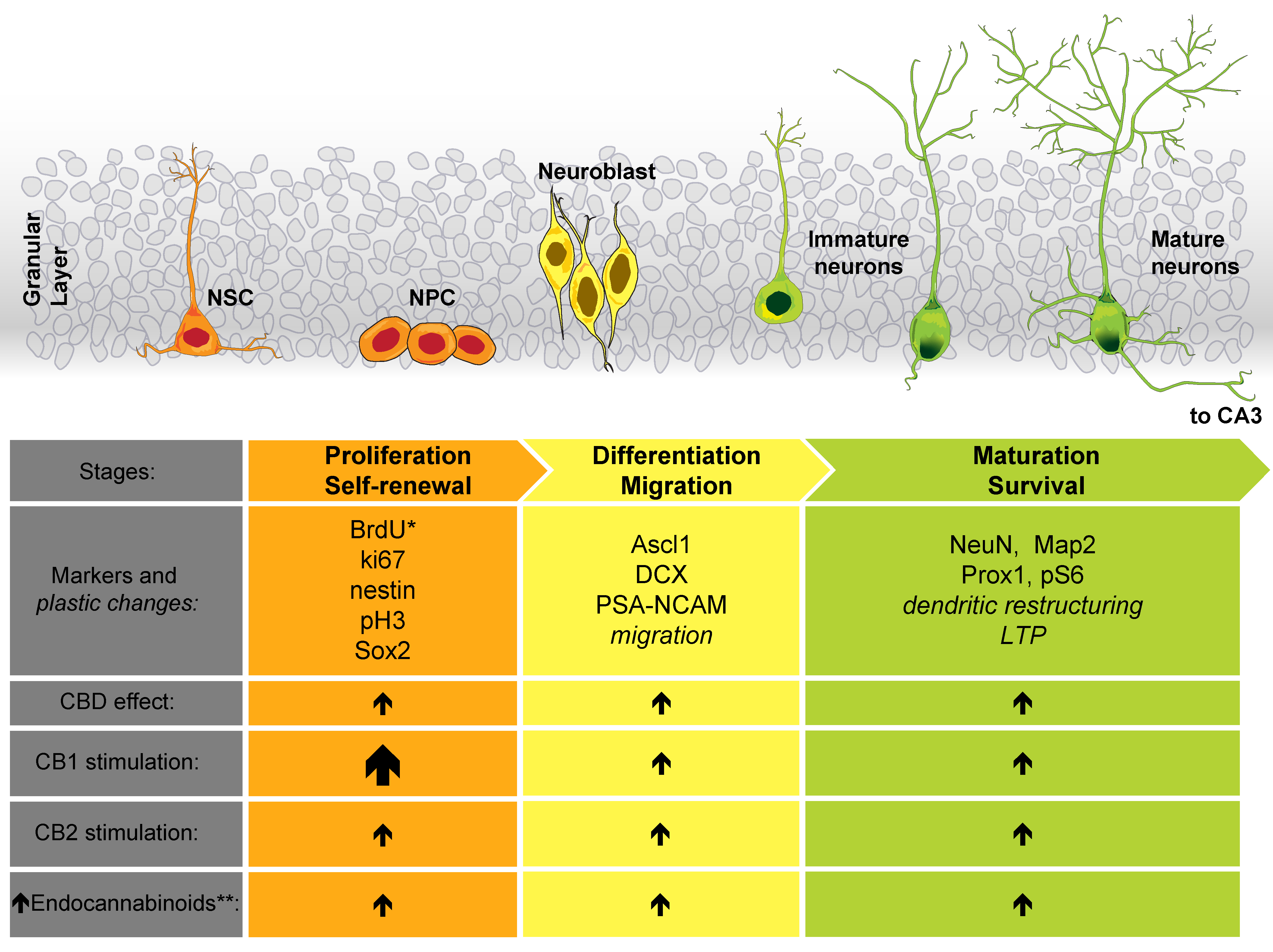
3. Cannabinoid Effect on Neurogenesis
4. Effect of Cannabinoids on Behavioral Processes: Stress, Anxiety, Learning and Memory
4.1. Anxiety
4.2. Stress
4.3. Learning and Memory
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Status Report on Alcohol and Health 2018; Poznyak, V., Rekve, D., Eds.; World Health Organization: Geneva, Switzerland, 2018; ISBN 978-92-4-156563-9. [Google Scholar]
- World Health Organization. WHO Report on the Global Tobacco Epidemic, 2019; WHO: Geneva, Switzerland, 2019; ISBN 9789241516204. [Google Scholar]
- Nations, U. World Drug Report 2020; Sales No. E.20.XI.6; United Nations Publication: New York, NY, USA, 2020; ISBN 9789211483451. [Google Scholar]
- Zarei, S.; Salimi, Y.; Repo, E.; Daglioglu, N.; Safaei, Z.; Güzel, E.; Asadi, A. A global systematic review and meta-analysis on illicit drug consumption rate through wastewater-based epidemiology. Environ. Sci. Pollut. Res. 2020, 27, 1–15. [Google Scholar] [CrossRef]
- Hartman, M. Cannabis Overview. Available online: https://www.ncsl.org/research/civil-and-criminal-justice/marijuana-overview.aspx (accessed on 6 June 2021).
- República de Uruguay Decreto no. 120/014. Reglamentacion de la Ley 19.172 Relativo a la Regulación y Control del Cannabis—Ley De Marihuana. Available online: https://www.impo.com.uy/bases/decretos/120-2014 (accessed on 25 May 2021).
- Observatorio Uruguayo de Drogas VII Encuesta Nacional Sobre Consumo de Drogas en Población General. Informe de Investigación; Junta Nacional de Drogas: Montevideo, Uruguay, 2019.
- Laqueur, H.; Rivera-Aguirre, A.; Shev, A.; Castillo-Carniglia, A.; Rudolph, K.E.; Ramirez, J.; Martins, S.S.; Cerdá, M. The impact of cannabis legalization in Uruguay on adolescent cannabis use. Int. J. Drug Policy 2020, 80, 102748. [Google Scholar] [CrossRef] [PubMed]
- Rotermann, M. Analysis of trends in the prevalence of cannabis use and related metrics in Canada. Public Health Rep. 2019, 30, 3–13. [Google Scholar]
- Rusby, J.C.; Westling, E.; Crowley, R.; Light, J.M. Legalization of recreational marijuana and community sales policy in Oregon: Impact on adolescent willingness and intent to use, parent use, and adolescent use. Psychol. Addict. Behav. 2018, 32, 84–92. [Google Scholar] [CrossRef]
- De Luca, M.A.; Di Chiara, G.; Cadoni, C.; Lecca, D.; Orsolini, L.; Papanti, D.; Corkery, J.M.; Schifano, F. Cannabis; Epidemiological, Neurobiological and Psychopathological Issues: An Update. CNS & Neurol. Disord. Drug Targets 2017, 16, 598–609. [Google Scholar] [CrossRef] [Green Version]
- Kingston, C.R.; Kirk, P.L. Separation of Components of Marijuana by Gas-Liquid Chromatography. Anal. Chem. 1961, 33, 1794–1795. [Google Scholar] [CrossRef]
- Mechoulam, R.; Parker, L.A. The Endocannabinoid System and the Brain. Annu. Rev. Psychol. 2013, 64, 21–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaoni, Y.; Mechoulam, R. Isolation, Structure, and Partial Synthesis of an Active Constituent of Hashish. J. Am. Chem. Soc. 1964, 86, 1646–1647. [Google Scholar] [CrossRef]
- ElSohly, M.A.; Radwan, M.M.; Gul, W.; Chandra, S.; Galal, A. Phytochemistry of Cannabis sativa L. Prog. Chem. Org. Nat. Prod. 2017, 103, 1–36. [Google Scholar] [CrossRef]
- Turner, S.E.; Williams, C.M.; Iversen, L.; Whalley, B.J. Molecular Pharmacology of Phytocannabinoids. In Phytocannabinoids. Progress in the Chemistry of Organic Natural Products; Springer: Cham, Switzerland, 2017; Volume 103, pp. 61–101. [Google Scholar] [CrossRef]
- Kilaru, A.; Chapman, K.D. The endocannabinoid system. Essays Biochem. 2020, 64, 485–499. [Google Scholar] [CrossRef]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Kano, M.; Ohno-Shosaku, T.; Hashimotodani, Y.; Uchigashima, M.; Watanabe, M. Endocannabinoid-Mediated Control of Synaptic Transmission. Physiol. Rev. 2009, 89, 309–380. [Google Scholar] [CrossRef] [PubMed]
- A Devane, W.; A Dysarz, F.; Johnson, M.R.; Melvin, L.S.; Howlett, A. Determination and characterization of a cannabinoid receptor in rat brain. Mol. Pharm. 1988, 34, 605–613. [Google Scholar]
- Munro, S.; Thomas, K.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nat. Cell Biol. 1993, 365, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kim, J. Neuronal expression of CB2 cannabinoid receptor mRNAs in the mouse hippocampus. Neuroscience 2015, 311, 253–267. [Google Scholar] [CrossRef] [Green Version]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.; Patel, S.; Perchuk, A.; Meozzi, P.A.; Myers, L.; Mora, Z.; Tagliaferro, P.; Gardner, E.; et al. Discovery of the Presence and Functional Expression of Cannabinoid CB2 Receptors in Brain. Ann. N. Y. Acad. Sci. 2006, 1074, 514–536. [Google Scholar] [CrossRef] [PubMed]
- Onaivi, E.S.; Leonard, C.M.; Ishiguro, H.; Zhang, P.W.; Lin, Z.; Akinshola, B.E.; Uhl, G.R. Endocannabinoids and cannabinoid receptor genetics. Prog. Neurobiol. 2002, 66, 307–344. [Google Scholar] [CrossRef]
- Sánchez-Zavaleta, R.; Cortés, H.; Avalos-Fuentes, J.A.; García, U.; Vila, J.S.; Erlij, D.; Florán, B. Presynaptic cannabinoid CB2 receptors modulate [3H]-Glutamate release at subthalamo-nigral terminals of the rat. Synapse 2018, 72, e22061. [Google Scholar] [CrossRef]
- Araque, A.; Castillo, P.E.; Manzoni, O.J.; Tonini, R. Synaptic functions of endocannabinoid signaling in health and disease. Neuropharmacology 2017, 124, 13–24. [Google Scholar] [CrossRef]
- Zou, S.; Kumar, U. Cannabinoid Receptors and the Endocannabinoid System: Signaling and Function in the Central Nervous System. Int. J. Mol. Sci. 2018, 19, 833. [Google Scholar] [CrossRef] [Green Version]
- Bénard, G.; Massa, F.; Puente, N.; Lourenço, J.; Bellocchio, L.; Soria-Gómez, E.; Matias, I.; Delamarre, A.; Metna-Laurent, M.; Cannich, A.; et al. Mitochondrial CB1 receptors regulate neuronal energy metabolism. Nat. Neurosci. 2012, 15, 558–564. [Google Scholar] [CrossRef]
- Hebert-Chatelain, E.; Desprez, T.; Serrat, R.; Bellocchio, L.; Soria-Gomez, E.; Busquets-Garcia, A.; Zottola, A.C.P.; Delamarre, A.; Cannich, A.; Vincent, P.; et al. A cannabinoid link between mitochondria and memory. Nature 2016, 539, 555–559. [Google Scholar] [CrossRef]
- Robin, L.M.; da Cruz, J.F.O.; Langlais, V.C.; Martin-Fernandez, M.; Metna-Laurent, M.; Busquets-Garcia, A.; Bellocchio, L.; Soria-Gomez, E.; Papouin, T.; Varilh, M.; et al. Astroglial CB1 Receptors Determine Synaptic D-Serine Availability to Enable Recognition Memory. Neuron 2018, 98, 935–944.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, C.; Xi, Z.-X. Progress in brain cannabinoid CB2 receptor research: From genes to behavior. Neurosci. Biobehav. Rev. 2019, 98, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Bystrowska, B.; Frankowska, M.; Smaga, I.; Pomierny-Chamiolo, L.; Filip, M. Effects of Cocaine Self-Administration and Its Extinction on the Rat Brain Cannabinoid CB1 and CB2 Receptors. Neurotox. Res. 2018, 34, 547–558. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.-Y.; Gao, M.; Liu, Q.-R.; Bi, G.-H.; Li, X.; Yang, H.-J.; Gardner, E.L.; Wu, J.; Xi, Z.-X. Cannabinoid CB2 receptors modulate midbrain dopamine neuronal activity and dopamine-related behavior in mice. Proc. Natl. Acad. Sci. USA 2014, 111, E5007–E5015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atwood, B.K.; Mackie, K. CB2: A cannabinoid receptor with an identity crisis. Br. J. Pharm. 2010, 160, 467–479. [Google Scholar] [CrossRef] [Green Version]
- A Pumroy, R.; Samanta, A.; Liu, Y.; Hughes, T.E.; Zhao, S.; Yudin, Y.; Rohacs, T.; Han, S.; Moiseenkova-Bell, V.Y. Author response: Molecular mechanism of TRPV2 channel modulation by cannabidiol. bioRxiv 2019, 4, 1–17. [Google Scholar] [CrossRef]
- Muller, C.; Morales, P.; Reggio, P.H. Cannabinoid Ligands Targeting TRP Channels. Front. Mol. Neurosci. 2019, 11, 487. [Google Scholar] [CrossRef]
- Bainbridge, J.S.; Davies, S.H. CCXXXI.—The essential oil of cocoa. J. Chem. Soc. Trans. 1912, 101, 2209–2221. [Google Scholar] [CrossRef] [Green Version]
- Mechoulam, R.; Ben-Shabat, S.; Hanus, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R.; et al. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharm. 1995, 50, 83–90. [Google Scholar] [CrossRef]
- Sugiura, T.; Kondo, S.; Sukagawa, A.; Nakane, S.; Shinoda, A.; Itoh, K.; Yamashita, A.; Waku, K. 2-arachidonoylglycerol: A possible endogenous cannabinoid receptor ligand in brain. Biochem. Biophys. Res. Commun. 1995, 215, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Ferber, S.G.; Trezza, V.; Weller, A. Early life stress and development of the endocannabinoid system: A bidirectional process in programming future coping. Dev. Psychobiol. 2021, 63, 143–152. [Google Scholar] [CrossRef]
- Rodrigues, R.S.; Ribeiro, F.; Ferreira, F.; Vaz, S.; Sebastião, A.M.; Xapelli, S. Interaction between Cannabinoid Type 1 and Type 2 Receptors in the Modulation of Subventricular Zone and Dentate Gyrus Neurogenesis. Front. Pharm. 2017, 8, 516. [Google Scholar] [CrossRef] [Green Version]
- Fowler, C.J. Anandamide uptake explained? Trends Pharm. Sci. 2012, 33, 181–185. [Google Scholar] [CrossRef]
- Baggelaar, M.P.; Maccarrone, M.; van der Stelt, M. 2-Arachidonoylglycerol: A signaling lipid with manifold actions in the brain. Prog. Lipid Res. 2018, 71, 1–17. [Google Scholar] [CrossRef]
- Chicca, A.; Marazzi, J.; Nicolussi, S.; Gertsch, J. Evidence for Bidirectional Endocannabinoid Transport across Cell Membranes. J. Biol. Chem. 2012, 287, 34660–34682. [Google Scholar] [CrossRef] [Green Version]
- Maccarrone, M. Missing Pieces to the Endocannabinoid Puzzle. Trends Mol. Med. 2020, 26, 263–272. [Google Scholar] [CrossRef]
- Bisby, M.; Richardson, R.; Baker, K.D. Developmental differences in the effects of CB1/2R agonist WIN55212-2 on extinction of learned fear. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 99, 109834. [Google Scholar] [CrossRef]
- Cirelli, C.; Tononi, G. Cortical Development, Electroencephalogram Rhythms, and the Sleep/Wake Cycle. Biol. Psychiatry 2015, 77, 1071–1078. [Google Scholar] [CrossRef]
- Hill, M.N.; Eiland, L.; Lee, T.T.; Hillard, C.J.; McEwen, B.S. Early life stress alters the developmental trajectory of corticolimbic endocannabinoid signaling in male rats. Neuropharmacology 2019, 146, 154–162. [Google Scholar] [CrossRef]
- Schneider, M.; Kasanetz, F.; Lynch, D.L.; Friemel, C.M.; Lassalle, O.; Hurst, D.P.; Steindel, F.; Monory, K.; Schäfer, C.; Miederer, I.; et al. Enhanced Functional Activity of the Cannabinoid Type-1 Receptor Mediates Adolescent Behavior. J. Neurosci. 2015, 35, 13975–13988. [Google Scholar] [CrossRef]
- Chesworth, R.; Long, L.E.; Weickert, C.S.; Karl, T. The Endocannabinoid System across Postnatal Development in Transmembrane Domain Neuregulin 1 Mutant Mice. Front. Psychiatry 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Vangopoulou, C.; Bourmpoula, M.T.; Koupourtidou, C.; Giompres, P.; Stamatakis, A.; Kouvelas, E.D.; Mitsacos, A. Effects of an early life experience on rat brain cannabinoid receptors in adolescence and adulthood. Ibro Rep. 2018, 5, 1–9. [Google Scholar] [CrossRef]
- Gomes, F.V.; Edelson, J.R.; Volk, D.W.; Grace, A.A. Altered brain cannabinoid 1 receptor mRNA expression across postnatal development in the MAM model of schizophrenia. Schizophr. Res. 2018, 201, 254–260. [Google Scholar] [CrossRef]
- Bernabeu, A.; Bara, A.; Manduca, A.; Borsoi, M.; Lassalle, O.; Pelissier-Alicot, A.-L.; Manzoni, O.J.; Manzoni, O. Title Sex-specific maturational trajectory of endocannabinoid plasticity in the rat prefrontal cortex. bioRxiv 2020, 2020. [Google Scholar] [CrossRef]
- Borsoi, M.; Manduca, A.; Bara, A.; Lassalle, O.; Pelissier-Alicot, A.-L.; Manzoni, O. Sex Differences in the Behavioral and Synaptic Consequences of a Single in vivo Exposure to the Synthetic Cannabimimetic WIN55,212-2 at Puberty and Adulthood. Front. Behav. Neurosci. 2019, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Mira, H.; Morante, J. Neurogenesis from Embryo to Adult Lessons From Flies and Mice. Front. Cell Dev. Biol. 2020, 8, 533. [Google Scholar] [CrossRef]
- Paridaen, J.T.M.L.; Huttner, W.B. Neurogenesis during development of the vertebrate central nervous system. Embo Rep. 2014, 15, 351–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altman, J.; Das, G.D. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J. Comp. Neurol. 1965, 124, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, P.S.; Perfilieva, E.; Bjork-Eriksson, T.; Alborn, A.-M.; Nordborg, C.; Peterson, D.A.; Gage, F.H. Neurogenesis in the adult human hippocampus. Nat. Med. 1998, 4, 1313–1317. [Google Scholar] [CrossRef] [PubMed]
- Spalding, K.; Bergmann, O.; Alkass, K.; Bernard, S.; Salehpour, M.; Huttner, H.B.; Boström, E.; Westerlund, I.; Vial, C.; Buchholz, B.; et al. Dynamics of Hippocampal Neurogenesis in Adult Humans. Cell 2013, 153, 1219–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobin, M.K.; Musaraca, K.; Disouky, A.; Shetti, A.; Bheri, A.; Honer, W.G.; Kim, N.; Dawe, R.J.; Bennett, D.A.; Arfanakis, K.; et al. Human Hippocampal Neurogenesis Persists in Aged Adults and Alzheimer’s Disease Patients. Cell Stem Cell 2019, 24, 974–982.e3. [Google Scholar] [CrossRef]
- Seki, T.; Arai, Y. Temporal and spacial relationships between PSA-NCAM-expressing, newly generated granule cells, and radial glia-like cells in the adult dentate gyrus. J. Comp. Neurol. 1999, 410, 503–513. [Google Scholar] [CrossRef]
- Boldrini, M.; Fulmore, C.A.; Tartt, A.; Simeon, L.R.; Pavlova, I.; Poposka, V.; Rosoklija, G.B.; Stankov, A.; Arango, V.; Dwork, A.J.; et al. Human Hippocampal Neurogenesis Persists throughout Aging. Cell Stem Cell 2018, 22, 589–599.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niklison-Chirou, M.V.; Agostini, M.; Amelio, I.; Melino, G. Regulation of Adult Neurogenesis in Mammalian Brain. Int. J. Mol. Sci. 2020, 21, 4869. [Google Scholar] [CrossRef] [PubMed]
- Faigle, R.; Song, H. Signaling mechanisms regulating adult neural stem cells and neurogenesis. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 2435–2448. [Google Scholar] [CrossRef] [Green Version]
- Gage, F.H. Adult neurogenesis in mammals. Science 2019, 364, 827–828. [Google Scholar] [CrossRef]
- Knoth, R.; Singec, I.; Ditter, M.; Pantazis, G.; Capetian, P.; Meyer, R.P.; Horvat, V.; Volk, B.; Kempermann, G. Murine Features of Neurogenesis in the Human Hippocampus across the Lifespan from 0 to 100 Years. PLoS ONE 2010, 5, e8809. [Google Scholar] [CrossRef]
- Seki, T. Understanding the Real State of Human Adult Hippocampal Neurogenesis from Studies of Rodents and Non-human Primates. Front. Neurosci. 2020, 14, 839. [Google Scholar] [CrossRef]
- Kempermann, G.; Gage, F.H.; Aigner, L.; Song, H.; Curtis, M.A.; Thuret, S.; Kuhn, H.-G.; Jessberger, S.; Frankland, P.W.; Cameron, H.A.; et al. Human Adult Neurogenesis: Evidence and Remaining Questions. Cell Stem Cell 2018, 23, 25–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurkowski, M.P.; Bettio, L.; Woo, E.K.; Patten, A.; Yau, S.-Y.; Gil-Mohapel, J. Beyond the Hippocampus and the SVZ: Adult Neurogenesis Throughout the Brain. Front. Cell. Neurosci. 2020, 14, 576444. [Google Scholar] [CrossRef] [PubMed]
- Buenrostro-Jáuregui, M.; Rodríguez-Serrano, L.M.; Chávez-Hernández, M.E.; Tapia-De-Jesús, A.; Mata-Luevanos, J.; Mata, F.; Galicia-Castillo, O.; Tirado-Martínez, D.; Ortega-Martinez, S.; Bojorges-Valdez, E. Simultaneous Monitoring of Wireless Electrophysiology and Memory Behavioral Test as a Tool to Study Hippocampal Neurogenesis. J. Vis. Exp. 2020, e61494. [Google Scholar] [CrossRef] [PubMed]
- Toda, T.; Gage, F.H. Review: Adult neurogenesis contributes to hippocampal plasticity. Cell Tissue Res. 2018, 373, 693–709. [Google Scholar] [CrossRef]
- Gonçalves, J.T.; Schafer, S.; Gage, F.H. Adult Neurogenesis in the Hippocampus: From Stem Cells to Behavior. Cell 2016, 167, 897–914. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.; Wen, Z.; Song, H.; Christian, K.M.; Ming, G.-L. Adult Neurogenesis and Psychiatric Disorders. Cold Spring Harb. Perspect. Biol. 2016, 8, a019026. [Google Scholar] [CrossRef] [Green Version]
- Apple, D.M.; Fonseca, R.S.; Kokovay, E. The role of adult neurogenesis in psychiatric and cognitive disorders. Brain Res. 2017, 1655, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Jessberger, S.; Clark, R.E.; Broadbent, N.J.; Clemenson, J.G.D.; Consiglio, A.; Lie, D.C.; Squire, L.R.; Gage, F.H. Dentate gyrus-specific knockdown of adult neurogenesis impairs spatial and object recognition memory in adult rats. Learn. Mem. 2009, 16, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Netzahualcoyotzi, C.; Pellerin, L. Neuronal and astroglial monocarboxylate transporters play key but distinct roles in hippocampus-dependent learning and memory formation. Prog. Neurobiol. 2020, 194, 101888. [Google Scholar] [CrossRef]
- Martinez-Canabal, A.; Akers, K.; Josselyn, S.A.; Frankland, P.W. Age-dependent effects of hippocampal neurogenesis suppression on spatial learning. Hippocampus 2013, 23, 66–74. [Google Scholar] [CrossRef]
- Saez, T.M.M.; Aronne, M.P.; Caltana, L.; Brusco, A.H. Prenatal exposure to the CB1 and CB2 cannabinoid receptor agonist WIN 55,212-2 alters migration of early-born glutamatergic neurons and GABAergic interneurons in the rat cerebral cortex. J. Neurochem. 2014, 129, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Prenderville, J.A.; Kelly, Á.M.; Downer, E.J. The role of cannabinoids in adult neurogenesis. Br. J. Pharm. 2015, 172, 3950–3963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusznák, K.; Csekő, K.; Varga, Z.; Csabai, D.; Bóna, Á.; Mayer, M.; Kozma, Z.; Helyes, Z.; Czéh, B. Long-Term Stress and Concomitant Marijuana Smoke Exposure Affect Physiology, Behavior and Adult Hippocampal Neurogenesis. Front. Pharm. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Aguareles, J.; Paraíso-Luna, J.; Palomares, B.; Bajo-Grañeras, R.; Navarrete, C.M.; Ruiz-Calvo, A.; García-Rincón, D.; García-Taboada, E.; Guzmán, M.; Muñoz, E.; et al. Oral administration of the cannabigerol derivative VCE-003.2 promotes subventricular zone neurogenesis and protects against mutant huntingtin-induced neurodegeneration. Transl. Neurodegener. 2019, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Buenrostro-Jauregui, M.; Tapia-De-Jesús, A.; Mata, J.; Rodríguez-Serrano, L.M.; Chávez-Hernández, M.E.; Mata, F.; Monroy-Plasencia, M.; Alonso-Flores, A.C.; Leal-Galicia, P. Immunohistochemistry Techniques to Analyze Cellular Proliferation and Neurogenesis in Rats using the Thymidine Analog BrdU. J. Vis. Exp. 2020, e61483. [Google Scholar] [CrossRef]
- Schiavon, A.P.; Bonato, J.M.; Milani, H.; Guimarães, F.S.; de Oliveira, R.M.W. Influence of single and repeated cannabidiol administration on emotional behavior and markers of cell proliferation and neurogenesis in non-stressed mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 64, 27–34. [Google Scholar] [CrossRef]
- Campos, A.C.; Ortega, Z.; Palazuelos, J.; Fogaça, M.V.; De Aguiar, D.C.; Díaz-Alonso, J.; Ortega-Gutiérrez, S.; Vázquez-Villa, H.; Moreira, F.A.; Guzmán, M.; et al. The anxiolytic effect of cannabidiol on chronically stressed mice depends on hippocampal neurogenesis: Involvement of the endocannabinoid system. Int. J. Neuropsychopharmacol. 2013, 16, 1407–1419. [Google Scholar] [CrossRef] [Green Version]
- Luján, M.Á.; Castro-Zavala, A.; Alegre-Zurano, L.; Valverde, O. Repeated Cannabidiol treatment reduces cocaine intake and modulates neural proliferation and CB1R expression in the mouse hippocampus. Neuropharmacology 2018, 143, 163–175. [Google Scholar] [CrossRef]
- Fogaça, M.V.; Campos, A.C.; Coelho, L.D.; Duman, R.S.; Guimarães, F.S. The anxiolytic effects of cannabidiol in chronically stressed mice are mediated by the endocannabinoid system: Role of neurogenesis and dendritic remodeling. Neuropharmacology 2018, 135, 22–33. [Google Scholar] [CrossRef]
- Mori, M.A.; Meyer, E.; Soares, L.M.; Milani, H.; Guimarães, F.S.; de Oliveira, R.M.W. Cannabidiol reduces neuroinflammation and promotes neuroplasticity and functional recovery after brain ischemia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 75, 94–105. [Google Scholar] [CrossRef]
- Rivera, P.; Blanco, E.; Bindila, L.; Alen, F.; Vargas, A.; Rubio, L.; Pavon, F.J.; Serrano, A.; Lutz, B.; De Fonseca, F.R.; et al. Pharmacological activation of CB2 receptors counteracts the deleterious effect of ethanol on cell proliferation in the main neurogenic zones of the adult rat brain. Front. Cell. Neurosci. 2015, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Hutch, C.R.; Hegg, C.C. Cannabinoid receptor signaling induces proliferation but not neurogenesis in the mouse olfactory epithelium. Neurogenesis 2016, 3, e1118177. [Google Scholar] [CrossRef]
- Zimmermann, T.; Maroso, M.; Beer, A.; Baddenhausen, S.; Ludewig, S.; Fan, W.; Vennin, C.; Loch, S.; Berninger, B.; Hofmann, C.; et al. Neural stem cell lineage-specific cannabinoid type-1 receptor regulates neurogenesis and plasticity in the adult mouse hippocampus. Cereb. Cortex 2018, 28, 4454–4471. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Calvo, E.; Rivera, P.; Arrabal, S.; Vargas, A.; Pavon, F.J.; Serrano, A.; Castilla-Ortega, E.; Galeano, P.; Rubio, L.; Suarez, J.; et al. Pharmacological blockade of either cannabinoid CB1 or CB2 receptors prevents both cocaine-induced conditioned locomotion and cocaine-induced reduction of cell proliferation in the hippocampus of adult male rat. Front. Integr. Neurosci. 2014, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolato, M.; Bini, V.; Frau, R.; Devoto, P.; Pardu, A.; Fan, Y.; Solbrig, M.V. Juvenile cannabinoid treatment induces frontostriatal gliogenesis in Lewis rats. Eur. Neuropsychopharmacol. 2014, 24, 974–985. [Google Scholar] [CrossRef] [PubMed]
- Abboussi, O.; Tazi, A.; Paizanis, E.; El Ganouni, S. Chronic exposure to WIN55,212-2 affects more potently spatial learning and memory in adolescents than in adult rats via a negative action on dorsal hippocampal neurogenesis. Pharm. Biochem. Behav. 2014, 120, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Palazuelos, J.; Ortega, Z.; Díaz-Alonso, J.; Guzmán, M.; Galve-Roperh, I. CB2 Cannabinoid Receptors Promote Neural Progenitor Cell Proliferation via mTORC1 Signaling. J. Biol. Chem. 2012, 287, 1198–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeffer, C.A.; Klann, E. mTOR signaling: At the crossroads of plasticity, memory and disease. Trends Neurosci. 2010, 33, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, M.B.; Suetterlin, P.; Yip, P.; Molina-Holgado, F.; Walker, D.J.; Oudin, M.; Zentar, M.P.; Pollard, S.; Yáñez-Muñoz, R.J.; Williams, G.; et al. A diacylglycerol lipase-CB2 cannabinoid pathway regulates adult subventricular zone neurogenesis in an age-dependent manner. Mol. Cell. Neurosci. 2008, 38, 526–536. [Google Scholar] [CrossRef]
- Hill, J.D.; Zuluaga-Ramirez, V.; Gajghate, S.; Winfield, M.; Persidsky, Y. Activation of GPR55 increases neural stem cell proliferation and promotes early adult hippocampal neurogenesis. Br. J. Pharm. 2018, 175, 3407–3421. [Google Scholar] [CrossRef] [Green Version]
- Jenniches, I.; Ternes, S.; Albayram, O.; Otte, D.M.; Bach, K.; Bindila, L.; Michel, K.; Lutz, B.; Bilkei-Gorzo, A.; Zimmer, A. Anxiety, Stress, and Fear Response in Mice With Reduced Endocannabinoid Levels. Biol. Psychiatry 2016, 79, 858–868. [Google Scholar] [CrossRef] [Green Version]
- Rivera, P.; Bindila, L.; Pastor, A.; Perez-Martin, M.; Pavon, F.J.; Serrano, A.; De La Torre, R.; Lutz, B.; De Fonseca, F.R.; Suã¡rez, J.; et al. Pharmacological blockade of the fatty acid amide hydrolase (FAAH) alters neural proliferation, apoptosis and gliosis in the rat hippocampus, hypothalamus and striatum in a negative energy context. Front. Cell. Neurosci. 2015, 9, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.-X.; Ke, B.-W.; Liu, J.; Ma, G.; Hai, K.-R.; Gong, D.-Y.; Yang, Z.; Zhou, C. Inhibition of Fatty Acid Amide Hydrolase Improves Depressive-Like Behaviors Independent of Its Peripheral Antinociceptive Effects in a Rat Model of Neuropathic Pain. Anesth. Analg. 2019, 129, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, W.; Zhong, P.; Liu, S.J.; Long, J.Z.; Zhao, L.; Gao, H.-Q.; Cravatt, B.F.; Liu, Q.-S. Blockade of 2-arachidonoylglycerol hydrolysis produces antidepressant-like effects and enhances adult hippocampal neurogenesis and synaptic plasticity. Hippocampus 2015, 25, 16–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, T.; Lee, H.; Young, A.H.; Aarsland, D.; Thuret, S. Adult Hippocampal Neurogenesis in Major Depressive Disorder and Alzheimer’s Disease. Trends Mol. Med. 2020, 26, 803–818. [Google Scholar] [CrossRef]
- Winner, B.; Winkler, J. Adult Neurogenesis in Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2015, 7, a021287. [Google Scholar] [CrossRef] [Green Version]
- Leal-Galicia, P.; Romo-Parra, H.; Rodríguez-Serrano, L.; Buenrostro-Jáuregui, M. Regulation of adult hippocampal neurogenesis exerted by sexual, cognitive and physical activity: An update. J. Chem. Neuroanat. 2019, 101, 101667. [Google Scholar] [CrossRef]
- Spanagel, R. Cannabinoids and the endocannabinoid system in reward processing and addiction: From mechanisms to interventions. Dialog Clin. Neurosci. 2020, 22, 241–250. [Google Scholar] [CrossRef]
- Rodríguez-Manzo, G.; González-Morales, E. Endocannabinoids mediate long-lasting behavioural and physiological changes in male rats induced by the repeated activation of the mesolimbic system by copulation to satiety. Behav. Brain Res. 2020, 383, 112510. [Google Scholar] [CrossRef]
- Tarragon, E.; Moreno, J.J. Role of Endocannabinoids on Sweet Taste Perception, Food Preference, and Obesity-related Disorders. Chem. Senses 2017, 43, 3–16. [Google Scholar] [CrossRef]
- Robinson, O.J.; Pike, A.C.; Cornwell, B.; Grillon, C. The translational neural circuitry of anxiety. J. Neurol. Neurosurg. Psychiatry 2019, 90, 1353–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simone, J.J.; Baumbach, J.L.; McCormick, C.M. Effects of CB1 receptor antagonism and stress exposures in adolescence on socioemotional behaviours, neuroendocrine stress responses, and expression of relevant proteins in the hippocampus and prefrontal cortex in rats. Neuropharmacology 2018, 128, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Ellgren, M.; Artmann, A.; Tkalych, O.; Gupta, A.; Hansen, H.S.; Hansen, S.; Devi, L.; Hurd, Y. Dynamic changes of the endogenous cannabinoid and opioid mesocorticolimbic systems during adolescence: THC effects. Eur. Neuropsychopharmacol. 2008, 18, 826–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, H.C.; Lee, F.S.; Gee, D.G. The Role of the Endocannabinoid System and Genetic Variation in Adolescent Brain Development. Neuropsychopharmacology 2018, 43, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heng, L.; Beverley, J.A.; Steiner, H.; Tseng, K.Y. Differential developmental trajectories for CB1 cannabinoid receptor expression in limbic/associative and sensorimotor cortical areas. Synapse 2011, 65, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Rey, A.A.; Purrio, M.; Viveros, M.-P.; Lutz, B. Biphasic Effects of Cannabinoids in Anxiety Responses: CB1 and GABAB Receptors in the Balance of GABAergic and Glutamatergic Neurotransmission. Neuropsychopharmacology 2012, 37, 2624–2634. [Google Scholar] [CrossRef] [Green Version]
- Ruehle, S.; Remmers, F.; Romo-Parra, H.; Massa, F.; Wickert, M.; Wörtge, S.; Häring, M.; Kaiser, N.; Marsicano, G.; Pape, H.-C.; et al. Cannabinoid CB1 Receptor in Dorsal Telencephalic Glutamatergic Neurons: Distinctive Sufficiency for Hippocampus-Dependent and Amygdala-Dependent Synaptic and Behavioral Functions. J. Neurosci. 2013, 33, 10264–10277. [Google Scholar] [CrossRef] [Green Version]
- Keeley, R.; Bye, C.; Trow, J.; McDonald, R. Strain and sex differences in brain and behaviour of adult rats: Learning and memory, anxiety and volumetric estimates. Behav. Brain Res. 2015, 288, 118–131. [Google Scholar] [CrossRef]
- Keeley, R.; Trow, J.; Bye, C.; McDonald, R. Part II: Strain- and sex-specific effects of adolescent exposure to THC on adult brain and behaviour: Variants of learning, anxiety and volumetric estimates. Behav. Brain Res. 2015, 288, 132–152. [Google Scholar] [CrossRef]
- Renard, J.; Rosen, L.G.; Loureiro, M.; De Oliveira, C.; Schmid, S.; Rushlow, W.J.; LaViolette, S.R. Adolescent Cannabinoid Exposure Induces a Persistent Sub-Cortical Hyper-Dopaminergic State and Associated Molecular Adaptations in the Prefrontal Cortex. Cereb. Cortex 2016, 27, 1297–1310. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.; Mills, S.; Winstone, J.; Leishman, E.; Wager-Miller, J.; Bradshaw, H.; Mackie, K. Chronic Adolescent Δ9-Tetrahydrocannabinol Treatment of Male Mice Leads to Long-Term Cognitive and Behavioral Dysfunction, Which Are Prevented by Concurrent Cannabidiol Treatment. Cannabis Cannabinoid Res. 2017, 2, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Rubino, T.; Vigano’, D.; Realini, N.; Guidali, C.; Braida, D.; Capurro, V.; Castiglioni, C.; Cherubino, F.; Romualdi, P.; Candeletti, S.; et al. Chronic Δ9-Tetrahydrocannabinol During Adolescence Provokes Sex-Dependent Changes in the Emotional Profile in Adult Rats: Behavioral and Biochemical Correlates. Neuropsychopharmacology 2008, 33, 2760–2771. [Google Scholar] [CrossRef]
- De Gregorio, D.; Conway, J.D.; Canul, M.-L.; Posa, L.; Bambico, F.R.; Gobbi, G. Effects of Chronic Exposure to Low-Dose delta-9-Tetrahydrocannabinol in Adolescence and Adulthood on Serotonin/Norepinephrine Neurotransmission and Emotional Behavior. Int. J. Neuropsychopharmacol. 2020, 23, 751–761. [Google Scholar] [CrossRef]
- Stopponi, S.; Soverchia, L.; Ubaldi, M.; Cippitelli, A.; Serpelloni, G.; Ciccocioppo, R. Chronic THC during adolescence increases the vulnerability to stress-induced relapse to heroin seeking in adult rats. Eur. Neuropsychopharmacol. 2014, 24, 1037–1045. [Google Scholar] [CrossRef]
- Silva, L.; Black, R.; Michaelides, M.; Hurd, Y.L.; Dow-Edwards, D. Sex and age specific effects of delta-9-tetrahydrocannabinol during the periadolescent period in the rat: The unique susceptibility of the prepubescent animal. Neurotoxicol. Teratol. 2016, 58, 88–100. [Google Scholar] [CrossRef]
- Renard, J.; Vitalis, T.; Rame, M.; Krebs, M.-O.; Lenkei, Z.; Le Pen, G.; Jay, T.M. Chronic cannabinoid exposure during adolescence leads to long-term structural and functional changes in the prefrontal cortex. Eur. Neuropsychopharmacol. 2016, 26, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Bambico, F.R.; Nguyen, N.-T.; Katz, N.; Gobbi, G. Chronic exposure to cannabinoids during adolescence but not during adulthood impairs emotional behaviour and monoaminergic neurotransmission. Neurobiol. Dis. 2010, 37, 641–655. [Google Scholar] [CrossRef] [PubMed]
- Morena, M.; Leitl, K.D.; Vecchiarelli, H.A.; Gray, J.M.; Campolongo, P.; Hill, M.N. Emotional arousal state influences the ability of amygdalar endocannabinoid signaling to modulate anxiety. Neuropharmacology 2016, 111, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Simone, J.J.; Baumbach, J.L.; McCormick, C.M. Sex-specific effects of CB1 receptor antagonism and stress in adolescence on anxiety, corticosterone concentrations, and contextual fear in adulthood in rats. Int. J. Dev. Neurosci. 2018, 69, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Panahi, Y.; Sahraei, H.; Johnston, T.P.; Sahebkar, A. The impact of stress on body function: A review. EXCLI J. 2017, 16, 1057–1072. [Google Scholar] [CrossRef] [PubMed]
- Morena, M.; Patel, S.; Bains, J.; Hill, M.N. Neurobiological Interactions Between Stress and the Endocannabinoid System. Neuropsychopharmacology 2015, 41, 80–102. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.T.-Y.; Wainwright, S.R.; Hill, M.N.; Galea, L.A.; Gorzalka, B.B. Sex, drugs, and adult neurogenesis: Sex-dependent effects of escalating adolescent cannabinoid exposure on adult hippocampal neurogenesis, stress reactivity, and amphetamine sensitization. Hippocampus 2014, 24, 280–292. [Google Scholar] [CrossRef]
- Alteba, S.; Korem, N.; Akirav, I. Cannabinoids reverse the effects of early stress on neurocognitive performance in adulthood. Learn. Mem. 2016, 23, 349–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.T.-Y.; Hill, M.N.; Hillard, C.J.; Gorzalka, B.B. Disruption of peri-adolescent endocannabinoid signaling modulates adult neuroendocrine and behavioral responses to stress in male rats. Neuropharmacology 2015, 99, 89–97. [Google Scholar] [CrossRef]
- Surkin, P.N.; Gallino, S.L.; Luce, V.; Correa, F.; Fernandez-Solari, J.; De Laurentiis, A. Pharmacological augmentation of endocannabinoid signaling reduces the neuroendocrine response to stress. Psychoneuroendocrinology 2018, 87, 131–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Houwer, J.; Barnes-Holmes, D.; Moors, A. What is learning? On the nature and merits of a functional definition of learning. Psychon. Bull. Rev. 2013, 20, 631–642. [Google Scholar] [CrossRef] [PubMed]
- García-Lázaro, H.G.; Ramirez-Carmona, R.; Lara-Romero, R.; Roldan-Valadez, E. Neuroanatomy of episodic and semantic memory in humans: A brief review of neuroimaging studies. Neurol. India 2012, 60, 613. [Google Scholar] [CrossRef] [PubMed]
- Segev, A.; Korem, N.; Zer-Aviv, T.M.; Abush, H.; Lange, R.; Sauber, G.; Hillard, C.J.; Akirav, I. Role of endocannabinoids in the hippocampus and amygdala in emotional memory and plasticity. Neuropsychopharmacology 2018, 43, 2017–2027. [Google Scholar] [CrossRef] [Green Version]
- Scarante, F.F.; Vila-Verde, C.; Detoni, V.L.; Ferreira-Junior, N.C.; Guimarães, F.S.; Campos, A.C. Cannabinoid Modulation of the Stressed Hippocampus. Front. Mol. Neurosci. 2017, 10, 411. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-T.; Mackie, K. Adolescent Δ9-Tetrahydrocannabinol Exposure Selectively Impairs Working Memory but Not Several Other mPFC-Mediated Behaviors. Front. Psychiatry 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Zamberletti, E.; Beggiato, S.; Steardo, L.; Prini, P.; Antonelli, T.; Ferraro, L.; Rubino, T.; Parolaro, D. Alterations of prefrontal cortex GABAergic transmission in the complex psychotic-like phenotype induced by adolescent delta-9-tetrahydrocannabinol exposure in rats. Neurobiol. Dis. 2014, 63, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Gibula-Tarlowska, E.; Wydra, K.; Kotlinska, J.H. Deleterious Effects of Ethanol, Δ(9)-Tetrahydrocannabinol (THC), and Their Combination on the Spatial Memory and Cognitive Flexibility in Adolescent and Adult Male Rats in the Barnes Maze Task. Pharmaceutics 2020, 12, 654. [Google Scholar] [CrossRef] [PubMed]
- Renard, J.; Rushlow, W.J.; LaViolette, S.R. What Can Rats Tell Us about Adolescent Cannabis Exposure? Insights from Preclinical Research. Can. J. Psychiatry 2016, 61, 328–334. [Google Scholar] [CrossRef] [Green Version]
- Kruse, L.C.; Cao, J.K.; Viray, K.; Stella, N.; Clark, J.J. Voluntary oral consumption of Δ9-tetrahydrocannabinol by adolescent rats impairs reward-predictive cue behaviors in adulthood. Neuropsychopharmacology 2019, 44, 1406–1414. [Google Scholar] [CrossRef]
- Poulia, N.; Delis, F.; Brakatselos, C.; Lekkas, P.; Kokras, N.; Dalla, C.; Antoniou, K. Escalating low-dose Δ9-tetrahydrocannabinol exposure during adolescence induces differential behavioral and neurochemical effects in male and female adult rats. Eur. J. Neurosci. 2019, 52, 2681–2693. [Google Scholar] [CrossRef]
- Stringfield, S.J.; Torregrossa, M.M. Disentangling the lasting effects of adolescent cannabinoid exposure. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 104, 110067. [Google Scholar] [CrossRef]
- Kucewicz, M.T.; Tricklebank, M.D.; Bogacz, R.; Jones, M.W. Dysfunctional Prefrontal Cortical Network Activity and Interactions following Cannabinoid Receptor Activation. J. Neurosci. 2011, 31, 15560–15568. [Google Scholar] [CrossRef] [Green Version]
- Cass, D.K.; Flores-Barrera, E.; Thomases, D.R.; Vital, W.F.; Caballero, A.; Tseng, K.Y. CB1 cannabinoid receptor stimulation during adolescence impairs the maturation of GABA function in the adult rat prefrontal cortex. Mol. Psychiatry 2014, 19, 536–543. [Google Scholar] [CrossRef] [Green Version]
- Stringfield, S.J.; Torregrossa, M.M. Intravenous self-administration of delta-9-THC in adolescent rats produces long-lasting alterations in behavior and receptor protein expression. Psychopharmacology 2021, 238, 305–319. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Substance | Worldwide | Americas | Oceania | Africa | Asia | Europe |
|---|---|---|---|---|---|---|
| Alcohol [1] | 43.00 | 54.10 | 53.80 | 32.20 | 33.10 | 59.90 |
| Tobacco [2] | 23.40 | 15.00 | 33.50 | 17.30 | 43.70 | 31.20 |
| Cannabis [3] | 3.80 | 8.80 | 10.57 | 6.32 | 1.86 | 5.39 |
| Amphetamines [3] | 0.55 | 1.30 | 1.35 | 0.41 | 0.42 | 0.47 |
| Opioids [3] | 1.16 | 1.86 | 2.47 | 1.04 | 1.11 | 0.68 |
| MDMA (Ecstasy) [3] | 0.41 | 0.53 | 1.67 | 0.26 | 0.37 | 0.61 |
| Cocaine [3] | 0.38 | 1.49 | 1.56 | 0.27 | 0.06 | 0.89 |
| Mechanism of Action | Compounds |
|---|---|
| Agonist CB1/CB2 | 2-AG *, AEA *, Δ9-THC **, HU-210, WIN 55,212-2, CP55,940 |
| Agonist CB1 | ACEA |
| Agonist CB2 | AM1241, HU-308, JWH-133, JWH-015, JWH-056 |
| Antagonist/inverse agonist > CB1 | AM251, ibipinabant, LY320135, AM28, rimonabant (SR141716A), surinabant |
| Antagonist/inverse agonist > CB2 | AM630, JTE907, SR 144528 |
| DAGL Inhibitor | RHC-80267, tetrahydrolipstatin(THL) |
| FAAH Inhibitor | URB597, URB937 |
| MAGL inhibitor | JZL184, KLM29 |
| CB1/CB2 weak antagonist TRPV1 and TRPV2 agonist GPR55 antagonist FABP inhibitor FAAH inhibitor | CBD ** |
| Animal Model ♂ * (Age in PND) | Dose Duration | Niche Analyzed | Markers | Effects | References |
|---|---|---|---|---|---|
| Mouse B6 ** + chronic unpredictable stress (CUS) (PND: 84) | 30 mg/kg i.p. 14 days | dorsal HPC | BrdU NeuN | CBD alone: ↑ BrdU+ cells, DCX+ cells, BrdU+NeuN+ cells CBD restored CUS-induced effects (↑ BrdU+, DCX+ cells) | [84] |
| mouse B6 + CUS (PND: 58) | 30 mg/kg i.p. 14 days | dorsal HPC | BrdU DCX Dendritic spines | CBD restored CUS-induced effects (↑ DCX+ cells and their migration, BrdU+ cells, NeuN+BrdU+ cells, number of dendritic spines and their length/branch number) | [86] |
| mouse CD1 + cocaine (PND: 42) | 20 mg/kg i.p. 10 days | dorsal HPC | BrdU DCX NeuN | CBD alone: ↑ BrdU+NeuN+ cells CBD restored cocaine-induced effects (↑ BrdU+NeuN+ cells, DCX+ cells) | [85] |
| mouse B6 + bilateral common carotid artery occlusion (BCCAO) (PND: 70) | 10 mg/kg i.p. 3 days | dorsal HPC | DCX | CBD restored BCCAO-induced effects (↑ DCX+ cells, dendritic restructuring) | [87] |
| mouse Swiss albino (PND: 42) | 3, 30 mg/kg i.p. 15 days | dorsal HPC SVZ | BrdU DCX Ki67 | 3 mg/kg: ↑ BrdU+ cells, DCX+ cells, Ki67+ cells 30 mg/kg: ↓ BrdU+ cells, DCX+ cells, Ki67+ | [83] |
| mouse B6 + AAV-mediated expression of mutant huntingtin in striatum (PND: 70) | VCE-003.2 *** 10 mg/kg p.o. 18 days | SVZ | Ascl1 GFAP Ki67 | VCE restored the huntingtin-induced effects (↑ GFAP+Ki67+ cells, Ascl1+ cells mobilization) | [81] |
| striatum | BrdU DCX NeuN | (↑ DCX+ cells, NeuN+BrdU+ cells) |
| Animal Model ♂ * (Age in PND) | Drug | Drug Category | Dose Duration | Niche Analyzed | Markers | Effects | References |
|---|---|---|---|---|---|---|---|
| rat Wistar + forced consumption of ethanol or sucrose (PND: 77) | ACEA | CB1 agonist | 3 mg/kg i.p. 5 days | dorsal HPC HPT SVZ | BrdU pH3 | ACEA restored the forced consumption-induced effects HPC: ↑ pH3+ cells HPC, HPT, SVZ: ↑ BrdU+ cells | [88] |
| rat Wistar + cocaine (PND: 77) | rimonabant | CB1 antagonist/inverse agonist | 3 mg/kg i.p. 1 day | dorsal HPC SVZ | BrdU GFAP | Those effects are exerted by ribonabant alone or + cocaine SVZ: ↓ BrdU+ cells HPC: ↑ BrdU+ cells, GFAP+ cells | [91] |
| mouse B6 + CUS (PND: 58) | AM251 | CB1 antagonist/inverse agonist | 0.3 mg/kg i.p. 14 days | dorsal HPC | BrdU DCX Dendritic spines | Block the neurogenic effect of CBD (↓ DCX+ cell migration, BrdU+ cells, and spines) | [86] |
| mouse Swiss Webster or CB1/CB2 KO mouse (PND: 49) | AM251 | CB1 antagonist/inverse agonist | 50 μL, 10 μM/mouse i.n. 1 day | olfactory epithelium | BrdU | In WT: Blocked the neurogenic effect of WIN55,212-2 (↓ BrdU+ cells) In KO: No changes | [89] |
| mouse nestin-CB1 KO (neuronal stem cells CB1 KO) (PND: 56) | No pharmacological treatment | dorsal HPC | BrdU DCX Dendritic spines NeuN Nestin LTP | At 28 and 56 dptm (days post-tamoxifen): ↓ nestin+ cells, DCX+ cells, NeuN+ cells, BrdU+ cells At 28 dptm: ↓ dendritic length and dendritic protrusions. Altered LTP | [90] | ||
| Sex, Animal Model (Age in PND) | Drug | Drug Category | Dose Duration | Niche analyzed | Markers | Effects | References |
|---|---|---|---|---|---|---|---|
| (sex not specified) mouse B6 or CB2 KO mouse (PND: 56) | HU-308 | CB2 agonist | 15 mg/kg i.p. 5 days | dorsal HPC | BrdU Nestin pS6 | B6: ↑ BrdU+ cell, pS6+ cells, BrdU+pS6+cells, Nestin+pS6+ cells KO: No changes | [94] |
| ♂ rat Wistar + forced consumption of ethanol or sucrose (PND: 77) | JWH133 | CB2 agonist | 0.2 mg/kg i.p. 5 days | dorsal HPC HPT SVZ | BrdU pH3 | Restored the forced consumption-induced effects in HPC, HPT, SVZ: ↑ BrdU+ cells, pH3+ cells | [88] |
| ♂ rat Wistar (28 or 56) | WIN55,212-2 | CB2 > CB1 agonist | 1 mg/kg 20 days | dorsal HPC ventral HPC | DCX | 28 PND: ↓ dorsal DCX+ cells | [93] |
| ♂ rat Lewis (PND: 42) | WIN55,212-2 | CB2 > CB1 agonist | 2 mg/kg i.p. 14 days | PFC striatum SVZ | BrdU | PFC, striatum, SVZ: ↑ BrdU+ cells | [92] |
| ♀ mouse B6 (PND: 42 or 168) | JWH-133 | CB2 agonist | 0.6 mg/kg i.p. on days 1–3 0.9 mg/kg i.p. on days 4–7 1.2 mg/kg i.p. on days 8–10 | olfactory bulb (OB) SVZ | BrdU Ki67 Neuroblast migration to OB | JWH: ↑ Ki67+ cells, neuroblast migration to OB | [96] |
| AM630 JTE907 | CB2 antagonist/inverse agonist | 5 mg/kg i.p. 5 days | AM: ↓ Ki67+ cells, neuroblast migration to the OB JTE: ↓ Ki67+ cells JTE: Block the neurogenic effect of WIN55,212-2 and JWH (↓ Ki67+ cells, neuroblast migration) | ||||
| ♂ rat Wistar + cocaine (PND: 77) | AM630 | CB2 antagonist/inverse agonist | 3 mg/kg i.p. 1 day | SVZ dorsal HPC | BrdU GFAP | Those effects are exerted by AM630 alone or + cocaine SVZ: ↓ BrdU+ cells HPC: ↑ BrdU+ cells, GFAP+ cells | [91] |
| ♂ mouse B6 + CUS (PND: 58) | AM630 | CB2 antagonist/inverse agonist | 0.3 mg/kg i.p. 14 days | dorsal HPC | BrdU DCX Dendritic spines | Block the neurogenic effect of CBD (↓ DCX+ cells, DCX+ cells migration, NeuN+BrdU+ cells, spines) | [86] |
| Sex, Animal Model (Age in PND) | Drug | Drug Category | Dose Duration | Niche Analyzed | Markers | Effects | References |
|---|---|---|---|---|---|---|---|
| ♀ mouse B6 (PND: 42 or 168) | RHC-80267 THL | DAGL inhibitor | RH = 0.01 μg or 0.3 μg i.c.v. THL = 0.15 μg i.c.v q.a.d. 7 days | OB SVZ | BrdU Ki67 Neuroblast migration to OB | RHC: ↓ Ki67+ cells RHC + washout: Partial recovery of Ki67+ cells RHC or THL: ↓ neuroblast migration | [96] |
| DAGL KO mouse (sex and age not specified) | No pharmacological treatment | dorsal HPC | BrdU | ↓ BrdU+ cells | [98] | ||
| ♂ mouse Swiss Webster or CB1/CB2 KO mouse (PND: 49) | JZL184 + URB597 | MAGL inhibitor + FAAH inhibitor | JZ = 50 μL, 10 μM/mouse i.n. 1 d URB = 50 μL, 100 μM/mouse i.n. 1 day | Olfactory epithelium | BrdU | In WT: ↑ BrdU+ cells In KO: No change | [89] |
| ♂ mouse B6 + CUS (PND: 63) | JZL184 | MAGL inhibitor | 8 mg/kg i.p.q.a.d. 3 weeks | dorsal HPC | BrdU DCX LTP | JZL restored the CUS -induced effects (↑ BrdU+ cells, DCX+ cells and LTP) | [101] |
| ♂ rat Wistar + chronic constriction injury (CCI) (PND: 49) | URB597 | FAAH inhibitor (systemic) | 5.8 mg/kg i.p. 14 days | dorsal HPC | BrdU BDNFmRNA Ki67 | URB597 restored the CCI-induced effects (↑ BrdU+ cells, Ki67, BDNFmRNA) | [100] |
| URB937 | FAAH inhibitor (peripheral) | 1.6 mg/kg i.p. 14 days | No change | ||||
| ♂ rat Wistar (PND: 77) | URB597 | FAAH inhibitor | 0.3 mg/kg i.p. 1 day | dorsal HPC HPT SVZ | BrdU pH3 | SVZ: ↑ pH3+ cells HPT, SVZ: ↑ BrdU+ cells | [99] |
| 0.3 mg/kg i.p. 5 days | HPC, HPT: ↓ pH3+ cells HPC: ↓ BrdU+ cells and its survival | ||||||
| ♂ rat Wistar + forced consumption of ethanol or sucrosesucrose (PND: 77) | URB597 | FAAH inhibitor | 0.3 mg/kg i.p. 5 days | dorsal HPC HPT SVZ | BrdU pH3 | No effect | [88] |
| Sex, Animal Model (Age in PND) | Drug | Drug Category | Dose Duration | Behavioral Effects | References |
|---|---|---|---|---|---|
| ♂ rat Sprague-Dawley (30) | THC | CB1/CB2 agonist | 1 mg/kg i.p. 20 days | ↓ Spent time in open arms (EPM), latency to immobility (FST), sucrose preference (SPT) = Latency to feed (NSFT), distance traveled (OFT) | [120] |
| ♂♀ mouse B6 (28) | 3 mg/kg i.p. 21 days | ↓ Impaired performance (delayed alternating T-maze) = Social behavior, open arm entries (EPM) and decision making (T-maze) | [137] | ||
| ♂ rat Wistar (30) | 1 mg/kg i.p. 4 days | ↑ Primary latency (Barnes maze) = Horizontal locomotor activity test | [139] | ||
| ♂ mouse CD1 (28) | 3 mg/kg, i.p. 20 days | ↑ % shredded (nestlet shredding task), marbles buried (marble burying task) ↓ Discrimination index (NORT), open arm entries (EPM) = Total distance traveled (OFT) | [118] | ||
| ♂♀ rat Sprague-Dawley (35) | 0.3 mg/kg i.p. on day 1–3 1 mg/kg i.p. on day 4–7 3 mg/kg i.p. on day 8–11 | ↑ Ambulatory counts (OFT) ↓ Discrimination index (OLT) | [142] | ||
| ♂ rat Sprague-Dawley (35) | 2.5 mg/kg i.p. on day 1–3 5 mg/kg i.p. on day 4–7 10 mg/kg i.p. on day 8–11 | ↑ Exploration time (light–dark box test) ↓ Distance traveled (OF), exploration time (social motivation and social cognition test), % inhibitory prepulse (SR) | [117] | ||
| ♂♀ rat Sprague-Dawley (35) | 2.5 mg/kg i.p. on day 1–3 5 mg/kg i.p. on day 4–7 10 mg/kg i.p. on day 8–11 | ↑ Immobility time (FST) ↓ % of sucrose preference (SPT) = Open arm entries (EPM), time spent in center (OF), spontaneous locomotor activity | [119] | ||
| ♂ rat Wistar (35) | 2.5 mg/kg i.p. on day 1–3 5 mg/kg i.p. on day 4–7 10 mg/kg i.p. on day 8–11 | ↓ Time in open arms (EPM) | [121] | ||
| ♂♀ rat Sprague-Dawley (32) | Self-administer escalating doses intravenously 3 μg/kg on day 1–3 10 μg/kg on day 4–6 30 μg/kg on day 7–20 | ↑ Discrimination index (delayed-match-to-sample working memory task) | [143] | ||
| ♀ rat Sprague-Dawley (35) | 2.5 mg/kg i.p. on day 1–3 5 mg/kg i.p. on day 4–7 10 mg/kg i.p. on day 8–11 | ↑ Immobility time (FST) ↓ Discrimination index (NOR), time spent in active social behaviors (SIT) | [138] | ||
| ♂ mouse CD1 (41) | CBD | ECS stimulator | 20 mg/kg i.p. 10 days | ↑ Time spent in open arms (EPM), discrimination index (NORT) | [85] |
| ♂ mouse Swiss albino (35) | 3, 10, 30 mg/kg i.p. 15 days | ↑ % open arm entries (EPM), latency for the first immobility episode (tail suspension test) | [83] | ||
| ♂ rat Wistar (27) | WIN55,212-2 | CB2 > CB1 agonist | 1 mg/kg i.p. 20 days | ↑ Latency to find the platform (MWM) ↓ Time in target area (MWM) | [93] |
| ♂♀ rat (strain not specified) (45) | 1.2 mg/kg i.p. 15 days | ↓♂♀Anxiety index (OFT) ↓ ♂♀Impaired performance (OLT) ↓ ♂♀ Impaired performance on the social recognition test ↓ ♂ Impaired performance (NORT) | [130] | ||
| ♀ rat Sprague-Dawley (30) | 0.2 and 1.0 mg/kg i.p. 20 days | ↑ Latency to feed (NSFT) ↓ Swimming and climbing (FST) = Time in open/close arms (EPM), distance traveled (OFT) | [124] | ||
| ♂ rat Lewis (35) | 2 mg/kg i.p. 13 days | ↑ Latency to starle peak (SR), duration of exploration approaches (NORT) = Open arm entries, duration of open arm entries (EPM), social interaction | [92] | ||
| ♂ rat Sprague-Dawley (35) | AM251 | CB1 antagonist/inverse agonist | 5 mg/kg i.p. 10 days | ↓ Immobility duration (FST) = Time spent in open arm (EPM) | [131] |
| ♂ rat Long-Evans (30) | 1 mg/kg, i.p. 14 days | = Time in open arm (EPM), time of interaction (SIT) | [109] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Netzahualcoyotzi, C.; Rodríguez-Serrano, L.M.; Chávez-Hernández, M.E.; Buenrostro-Jáuregui, M.H. Early Consumption of Cannabinoids: From Adult Neurogenesis to Behavior. Int. J. Mol. Sci. 2021, 22, 7450. https://doi.org/10.3390/ijms22147450
Netzahualcoyotzi C, Rodríguez-Serrano LM, Chávez-Hernández ME, Buenrostro-Jáuregui MH. Early Consumption of Cannabinoids: From Adult Neurogenesis to Behavior. International Journal of Molecular Sciences. 2021; 22(14):7450. https://doi.org/10.3390/ijms22147450
Chicago/Turabian StyleNetzahualcoyotzi, Citlalli, Luis Miguel Rodríguez-Serrano, María Elena Chávez-Hernández, and Mario Humberto Buenrostro-Jáuregui. 2021. "Early Consumption of Cannabinoids: From Adult Neurogenesis to Behavior" International Journal of Molecular Sciences 22, no. 14: 7450. https://doi.org/10.3390/ijms22147450