
Rapid Internalization and Nuclear Translocation of CCL5 and CXCL4 in Endothelial Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
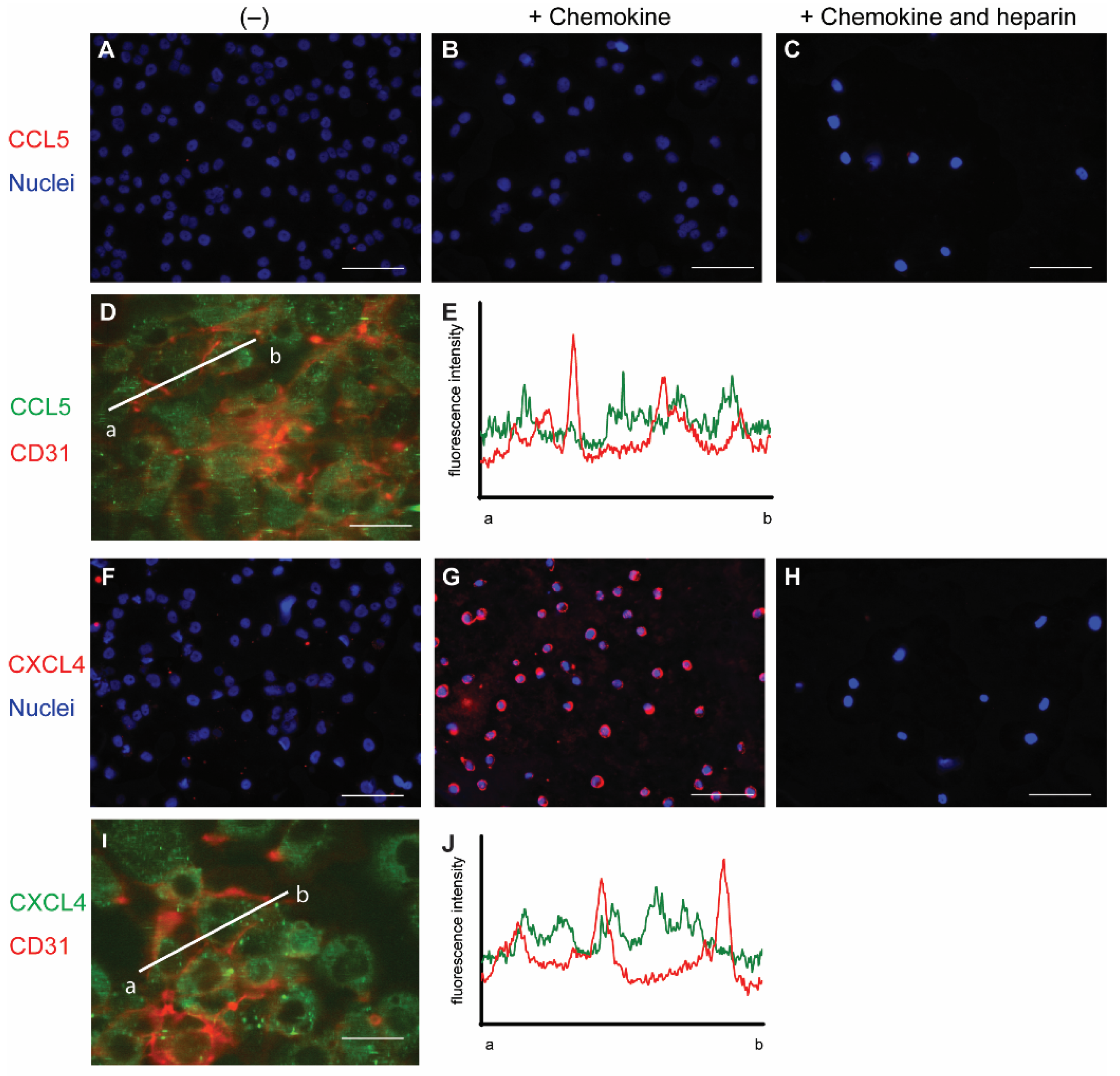
2.1. Surface Presentation of the Chemokines CCL5 and CXCL4 on EC
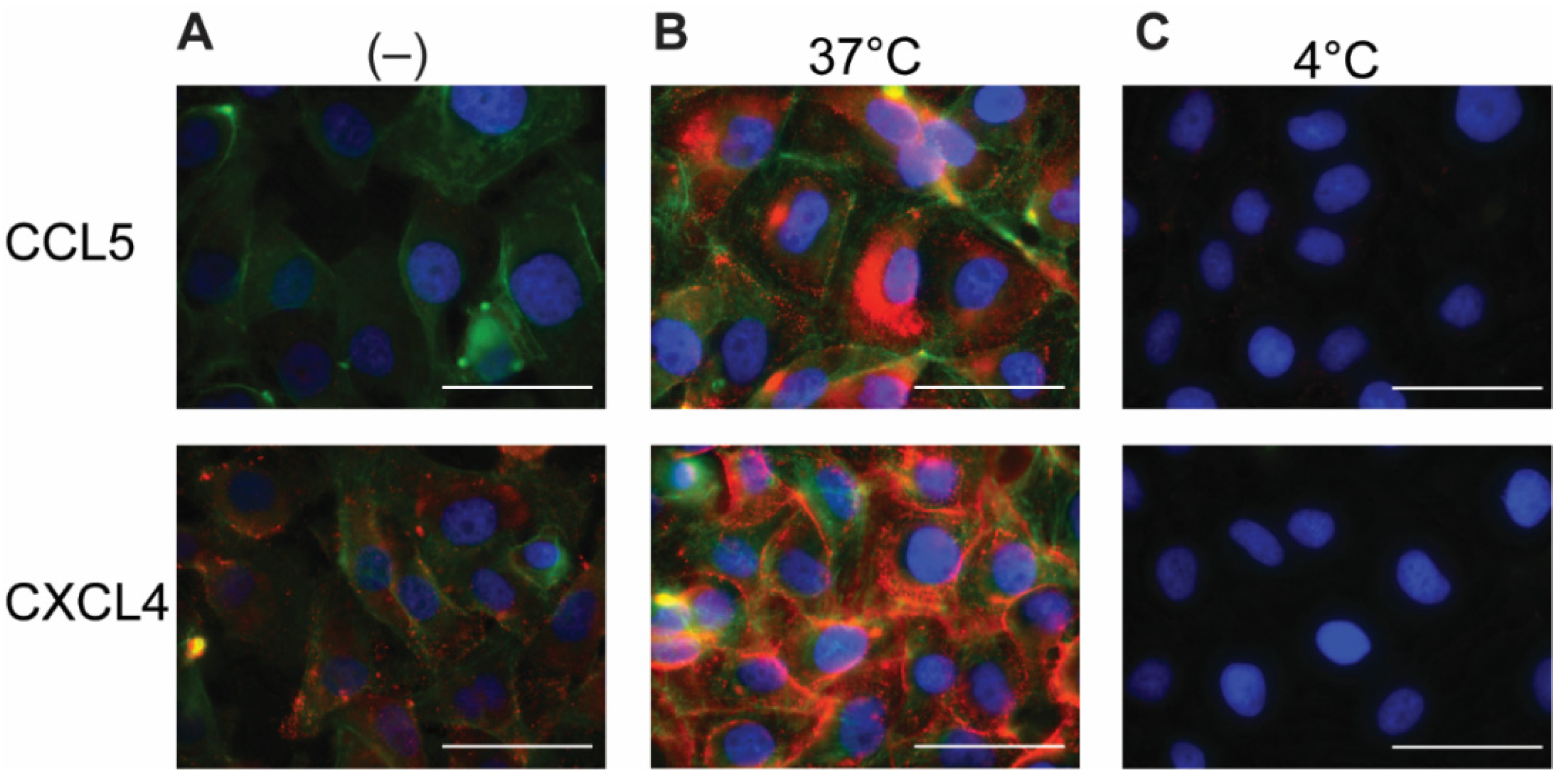
2.2. Permeabilization of EAHy Increases the CCL5 and CXCL4 Antigen Signal
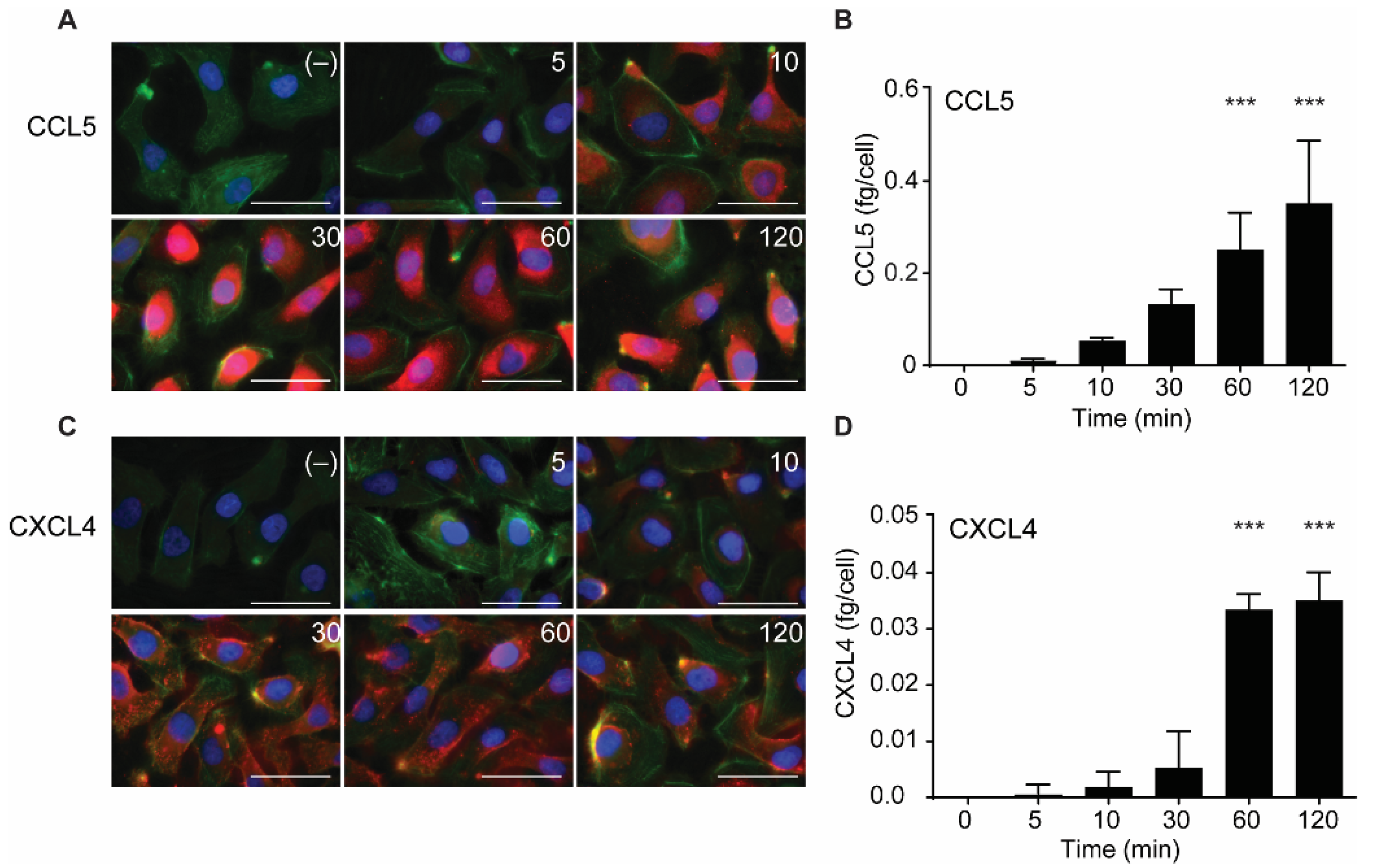
2.3. Intracellular Accumulation of CCL5 and CXCL4 Is Time-Dependent
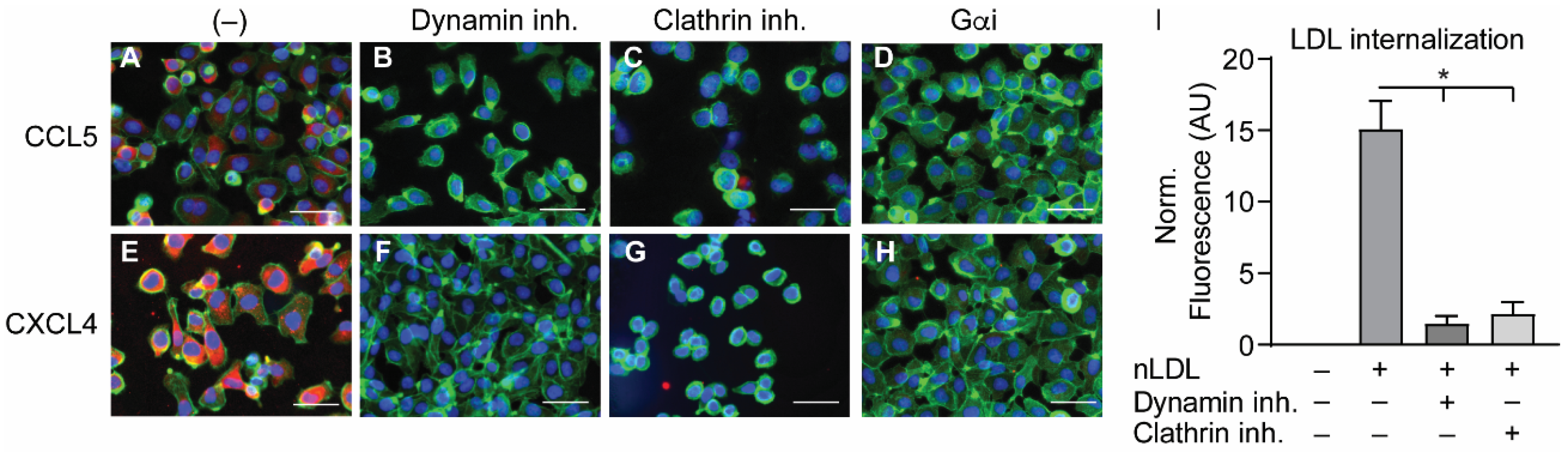
2.4. Internalization of CCL5 and CXCL4 Is Dependent on Dynamin- and Clathrin-Mediated Endocytosis
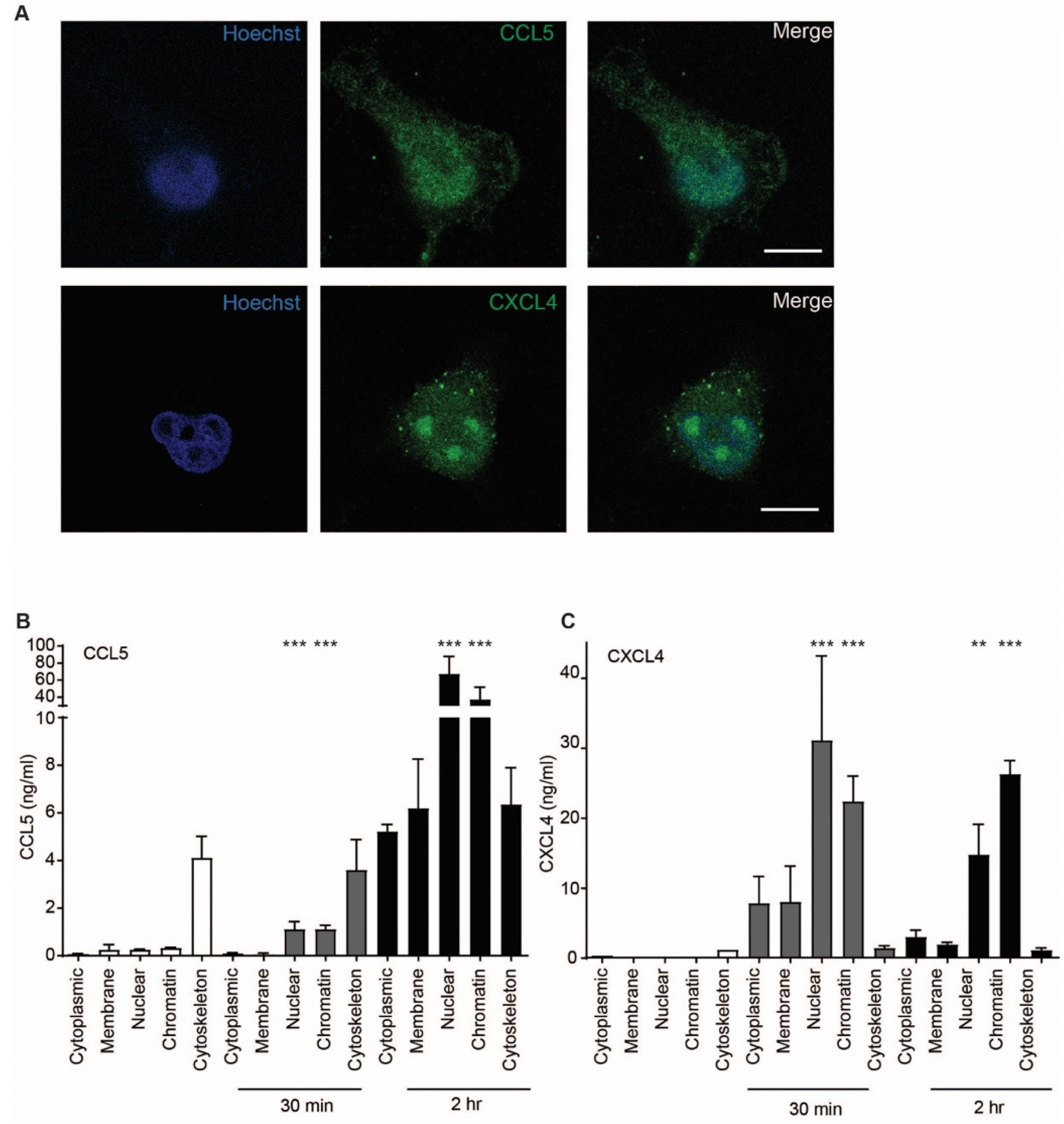
2.5. CCL5 and CXCL4 Are Targeted to the Nucleus after Endothelial Uptake
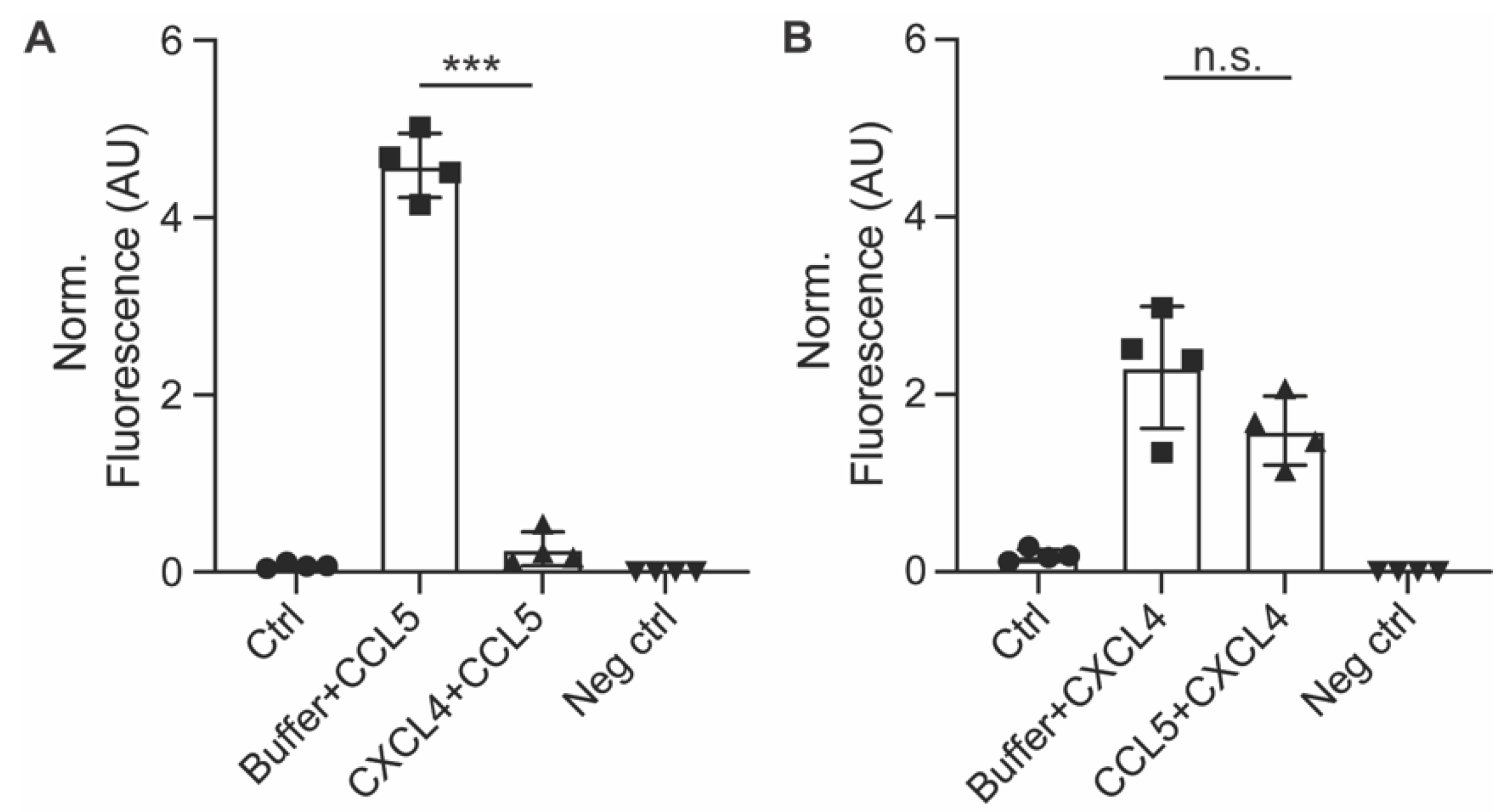
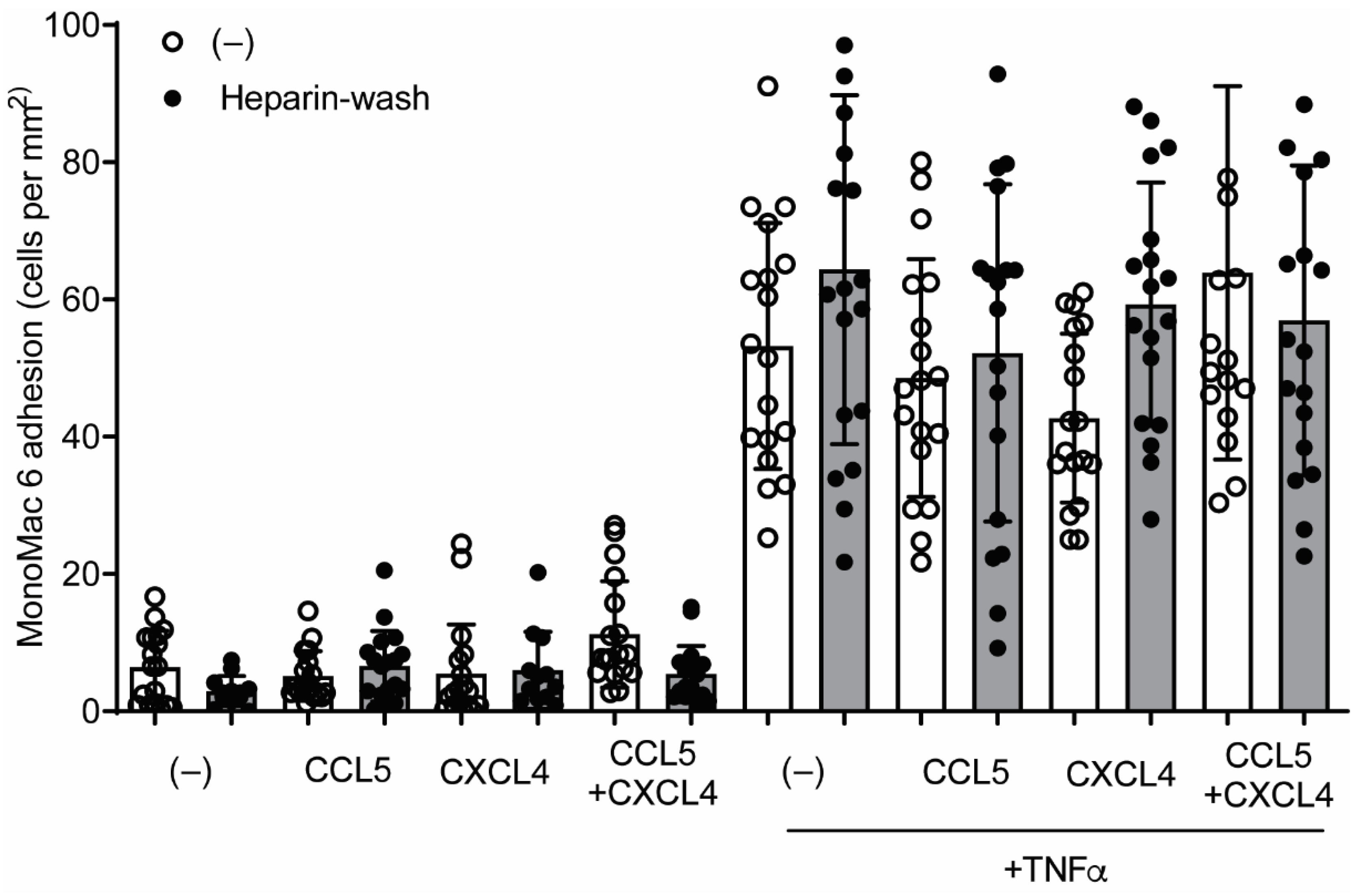
2.6. CCL5 and CXCL4 Internalization Does Not Affect Leukocyte Arrest
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Chemokine Internalization
4.3. Inhibitors
4.4. Localization
4.5. Immunocytochemistry
4.6. Live Cell Imaging
4.7. Confocal Imaging
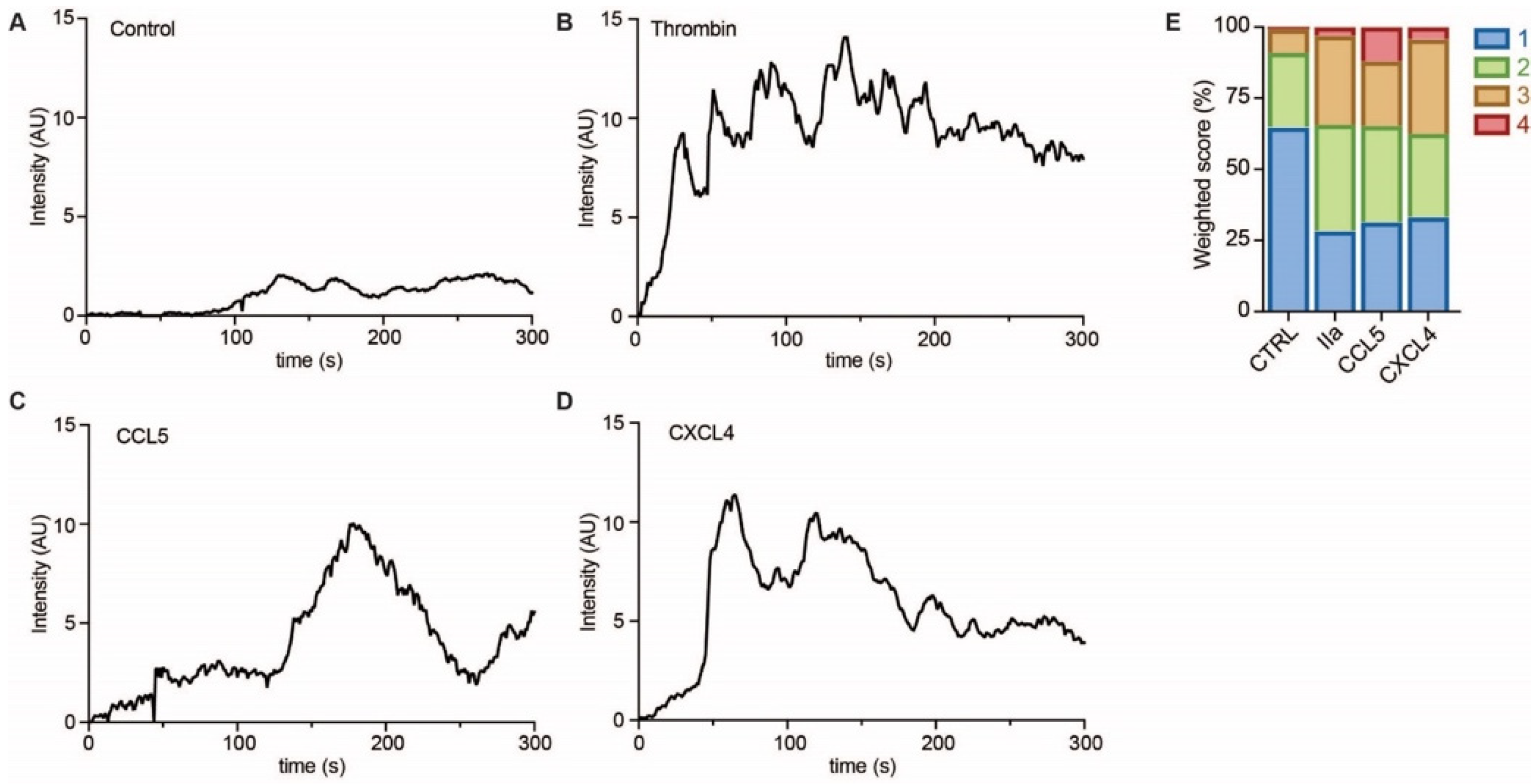
4.8. Calcium Influx
4.9. Quantification of Internalized Chemokines
4.10. Laminar Flow-Based Leukocyte Adhesion Assay
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noels, H.; Weber, C.; Koenen, R.R. Chemokines as therapeutic targets in cardiovascular disease. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 583–592. [Google Scholar] [CrossRef] [Green Version]
- Weber, C.; Noels, H. Atherosclerosis: Current pathogenesis and therapeutic options. Nat. Med. 2011, 17, 1410–1422. [Google Scholar] [CrossRef]
- Graham, G.J.; Handel, T.M.; Proudfoot, A.E.I. leukocyte adhesion: Reconceptualizing chemokine presentation by glycosaminoglycans. Trends Immunol. 2019, 40, 472–481. [Google Scholar] [CrossRef] [Green Version]
- Rot, A. Chemokine patterning by glycosaminoglycans and interceptors. Front. Biosci. 2010, 15, 645–660. [Google Scholar] [CrossRef]
- Shulman, Z.; Cohen, S.J.; Roediger, B.; Kalchenko, V.; Jain, R.; Grabovsky, V.; Klein, E.; Shinder, V.; Stoler-Barak, L.; Feigelson, S.W.; et al. Transendothelial migration of lymphocytes mediated by intraendothelial vesicle stores rather than by extracellular chemokine depots. Nat. Immunol. 2011, 13, 67–76. [Google Scholar] [CrossRef]
- Middleton, J.; Neil, S.; Wintle, J.; Clark-Lewis, I.; Moore, H.; Lam, C.; Auer, M.; Hub, E.; Rot, A. Transcytosis and surface presentation of IL-8 by venular endothelial cells. Cell 1997, 91, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Pruenster, M.; Mudde, L.; Bombosi, P.; Dimitrova, S.; Zsak, M.; Middleton, J.; Richmond, A.; Graham, G.J.; Segerer, S.; Nibbs, R.J.; et al. The Duffy antigen receptor for chemokines transports chemokines and supports their promigratory activity. Nat. Immunol. 2009, 10, 101–108. [Google Scholar] [CrossRef] [PubMed]
- von Hundelshausen, P.; Weber, K.S.; Huo, Y.; Proudfoot, A.E.; Nelson, P.J.; Ley, K.; Weber, C. RANTES deposition by platelets triggers monocyte arrest on inflamed and atherosclerotic endothelium. Circulation 2001, 103, 1772–1777. [Google Scholar] [CrossRef] [PubMed]
- Schober, A.; Manka, D.; von Hundelshausen, P.; Huo, Y.; Hanrath, P.; Sarembock, I.J.; Ley, K.; Weber, C. Deposition of platelet RANTES triggering monocyte recruitment requires P-selectin and is involved in neointima formation after arterial injury. Circulation 2002, 106, 1523–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, Y.; Schober, A.; Forlow, S.B.; Smith, D.F.; Hyman, M.C.; Jung, S.; Littman, D.R.; Weber, C.; Ley, K. Circulating activated platelets exacerbate atherosclerosis in mice deficient in apolipoprotein E. Nat. Med. 2003, 9, 61–67. [Google Scholar] [CrossRef] [PubMed]
- von Hundelshausen, P.; Koenen, R.R.; Sack, M.; Mause, S.F.; Adriaens, W.; Proudfoot, A.E.; Hackeng, T.M.; Weber, C. Heterophilic interactions of platelet factor 4 and RANTES promote monocyte arrest on endothelium. Blood 2005, 105, 924–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Xiong, X.; Zhang, Y.; Yan, D.; Jian, Z.; Xu, B.; Zhao, H. MKEY, a peptide inhibitor of CXCL4-CCL5 heterodimer formation, protects against stroke in mice. J. Am. Heart Assoc. 2016, 5, e003615. [Google Scholar] [CrossRef] [PubMed]
- Iida, Y.; Xu, B.; Xuan, H.; Glover, K.J.; Tanaka, H.; Hu, X.; Fujimura, N.; Wang, W.; Schultz, J.R.; Turner, C.R.; et al. Peptide inhibitor of CXCL4-CCL5 heterodimer formation, MKEY, inhibits experimental aortic aneurysm initiation and progression. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 718–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koenen, R.R.; von Hundelshausen, P.; Nesmelova, I.V.; Zernecke, A.; Liehn, E.A.; Sarabi, A.; Kramp, B.K.; Piccinini, A.M.; Paludan, S.R.; Kowalska, M.A.; et al. Disrupting functional interactions between platelet chemokines inhibits atherosclerosis in hyperlipidemic mice. Nat. Med. 2009, 15, 97–103. [Google Scholar] [CrossRef] [PubMed]
- von Hundelshausen, P.; Agten, S.M.; Eckardt, V.; Blanchet, X.; Schmitt, M.M.; Ippel, H.; Neideck, C.; Bidzhekov, K.; Leberzammer, J.; Wichapong, K.; et al. Chemokine interactome mapping enables tailored intervention in acute and chronic inflammation. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef]
- Vajen, T.; Koenen, R.R.; Werner, I.; Staudt, M.; Projahn, D.; Curaj, A.; Sonmez, T.T.; Simsekyilmaz, S.; Schumacher, D.; Mollmann, J.; et al. Blocking CCL5-CXCL4 heteromerization preserves heart function after myocardial infarction by attenuating leukocyte recruitment and NETosis. Sci. Rep. 2018, 8, 10647. [Google Scholar] [CrossRef] [PubMed]
- Proudfoot, A.E.; Handel, T.M.; Johnson, Z.; Lau, E.K.; LiWang, P.; Clark-Lewis, I.; Borlat, F.; Wells, T.N.; Kosco-Vilbois, M.H. Glycosaminoglycan binding and oligomerization are essential for the in vivo activity of certain chemokines. Proc. Natl. Acad. Sci. USA 2003, 100, 1885–1890. [Google Scholar] [CrossRef] [Green Version]
- Oynebraten, I.; Barois, N.; Bergeland, T.; Kuchler, A.M.; Bakke, O.; Haraldsen, G. Oligomerized, filamentous surface presentation of RANTES/CCL5 on vascular endothelial cells. Sci. Rep. 2015, 5, 9261. [Google Scholar] [CrossRef] [Green Version]
- Sarabi, A.; Kramp, B.K.; Drechsler, M.; Hackeng, T.M.; Soehnlein, O.; Weber, C.; Koenen, R.R.; Von Hundelshausen, P. CXCL4L1 inhibits angiogenesis and induces undirected endothelial cell migration without affecting endothelial cell proliferation and monocyte recruitment. J. Thromb. Haemost. 2011, 9, 209–219. [Google Scholar] [CrossRef]
- Santa-Maria, A.R.; Walter, F.R.; Figueiredo, R.; Kincses, A.; Vigh, J.P.; Heymans, M.; Culot, M.; Winter, P.; Gosselet, F.; Der, A.; et al. Flow induces barrier and glycocalyx-related genes and negative surface charge in a lab-on-a-chip human blood-brain barrier model. J. Cereb. Blood Flow Metab. 2021. [Google Scholar] [CrossRef]
- Salman, M.M.; Marsh, G.; Kusters, I.; Delince, M.; di Caprio, G.; Upadhyayula, S.; de Nola, G.; Hunt, R.; Ohashi, K.G.; Gray, T.; et al. Design and validation of a human brain endothelial microvessel-on-a-chip open microfluidic model enabling advanced optical imaging. Front. Bioeng. Biotechnol. 2020, 8, 573775. [Google Scholar] [CrossRef]
- Lopez, J.A.; Zheng, Y. Synthetic microvessels. J. Thromb. Haemost. 2013, 11, 67–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ader, M.; Tanaka, E.M. Modeling human development in 3D culture. Curr. Opin. Cell Biol. 2014, 31, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Kitchen, P.; Salman, M.M.; Abir-Awan, M.; Al-Jubair, T.; Tornroth-Horsefield, S.; Conner, A.C.; Bill, R.M. Calcein fluorescence quenching to measure plasma membrane water flux in live mammalian cells. STAR Protoc. 2020, 1, 100157. [Google Scholar] [CrossRef] [PubMed]
- Lasagni, L.; Francalanci, M.; Annunziato, F.; Lazzeri, E.; Giannini, S.; Cosmi, L.; Sagrinati, C.; Mazzinghi, B.; Orlando, C.; Maggi, E.; et al. An alternatively spliced variant of CXCR3 mediates the inhibition of endothelial cell growth induced by IP-10, Mig, and I-TAC, and acts as functional receptor for platelet factor 4. J. Exp. Med. 2003, 197, 1537–1549. [Google Scholar] [CrossRef] [PubMed]
- Campanella, G.S.; Colvin, R.A.; Luster, A.D. CXCL10 can inhibit endothelial cell proliferation independently of CXCR3. PLoS ONE 2010, 5, e12700. [Google Scholar] [CrossRef]
- Romagnani, P.; Annunziato, F.; Lasagni, L.; Lazzeri, E.; Beltrame, C.; Francalanci, M.; Uguccioni, M.; Galli, G.; Cosmi, L.; Maurenzig, L.; et al. Cell cycle-dependent expression of CXC chemokine receptor 3 by endothelial cells mediates angiostatic activity. J. Clin. Investig. 2001, 107, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Nabi, I.R.; Shankar, J.; Dennis, J.W. The galectin lattice at a glance. J. Cell Sci. 2015, 128, 2213–2219. [Google Scholar] [CrossRef] [Green Version]
- Garcia Caballero, G.; Kaltner, H.; Kutzner, T.J.; Ludwig, A.K.; Manning, J.C.; Schmidt, S.; Sinowatz, F.; Gabius, H.J. How galectins have become multifunctional proteins. Histol. Histopathol. 2020, 35, 509–539. [Google Scholar] [CrossRef]
- Dickhout, A.; Tullemans, B.M.E.; Heemskerk, J.W.M.; Thijssen, V.; Kuijpers, M.J.E.; Koenen, R.R. Galectin-1 and platelet factor 4 (CXCL4) induce complementary platelet responses in vitro. PLoS ONE 2021, 16, e0244736. [Google Scholar] [CrossRef]
- Eckardt, V.; Miller, M.C.; Blanchet, X.; Duan, R.; Leberzammer, J.; Duchene, J.; Soehnlein, O.; Megens, R.T.; Ludwig, A.K.; Dregni, A.; et al. Chemokines and galectins form heterodimers to modulate inflammation. EMBO Rep. 2020, 21, e47852. [Google Scholar] [CrossRef]
- Kuschert, G.S.; Coulin, F.; Power, C.A.; Proudfoot, A.E.; Hubbard, R.E.; Hoogewerf, A.J.; Wells, T.N. Glycosaminoglycans interact selectively with chemokines and modulate receptor binding and cellular responses. Biochemistry 1999, 38, 12959–12968. [Google Scholar] [CrossRef] [PubMed]
- del Molino del Barrio, I.; Kirby, J.; Ali, S. The role of chemokine and glycosaminoglycan interaction in chemokine-mediated migration in vitro and in vivo. Methods Enzymol. 2016, 570, 309–333. [Google Scholar] [CrossRef] [PubMed]
- Johnson, Z.; Kosco-Vilbois, M.H.; Herren, S.; Cirillo, R.; Muzio, V.; Zaratin, P.; Carbonatto, M.; Mack, M.; Smailbegovic, A.; Rose, M.; et al. Interference with heparin binding and oligomerization creates a novel anti-inflammatory strategy targeting the chemokine system. J. Immunol. 2004, 173, 5776–5785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhuri, A.; Zbrzezna, V.; Polyakova, J.; Pogo, A.O.; Hesselgesser, J.; Horuk, R. Expression of the Duffy antigen in K562 cells. Evidence that it is the human erythrocyte chemokine receptor. J. Biol. Chem. 1994, 269, 7835–7838. [Google Scholar] [CrossRef]
- Szabo, M.C.; Soo, K.S.; Zlotnik, A.; Schall, T.J. Chemokine class differences in binding to the Duffy antigen-erythrocyte chemokine receptor. J. Biol. Chem. 1995, 270, 25348–25351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Mangalmurti, N.S.; Xiong, Z.; Prakash, B.; Guo, F.; Stolz, D.B.; Lee, J.S. Duffy antigen receptor for chemokines mediates chemokine endocytosis through a macropinocytosis-like process in endothelial cells. PLoS ONE 2011, 6, e29624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, R.; Ramirez, J.C. A chemokine targets the nucleus: Cxcl12-gamma isoform localizes to the nucleolus in adult mouse heart. PLoS ONE 2009, 4, e7570. [Google Scholar] [CrossRef] [PubMed]
- Gortz, A.; Nibbs, R.J.; McLean, P.; Jarmin, D.; Lambie, W.; Baird, J.W.; Graham, G.J. The chemokine ESkine/CCL27 displays novel modes of intracrine and paracrine function. J. Immunol. 2002, 169, 1387–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaax, M.E.; Krauel, K.; Marschall, T.; Brandt, S.; Gansler, J.; Furll, B.; Appel, B.; Fischer, S.; Block, S.; Helm, C.A.; et al. Complex formation with nucleic acids and aptamers alters the antigenic properties of platelet factor 4. Blood 2013, 122, 272–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lande, R.; Lee, E.Y.; Palazzo, R.; Marinari, B.; Pietraforte, I.; Santos, G.S.; Mattenberger, Y.; Spadaro, F.; Stefanantoni, K.; Iannace, N.; et al. CXCL4 assembles DNA into liquid crystalline complexes to amplify TLR9-mediated interferon-alpha production in systemic sclerosis. Nat. Commun. 2019, 10, 1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleissner, C.A.; Shaked, I.; Little, K.M.; Ley, K. CXC chemokine ligand 4 induces a unique transcriptome in monocyte-derived macrophages. J. Immunol. 2010, 184, 4810–4818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva-Cardoso, S.C.; Tao, W.; Angiolilli, C.; Lopes, A.P.; Bekker, C.P.J.; Devaprasad, A.; Giovannone, B.; van Laar, J.; Cossu, M.; Marut, W.; et al. CXCL4 links inflammation and fibrosis by reprogramming monocyte-derived dendritic cells in vitro. Front. Immunol. 2020, 11, 2149. [Google Scholar] [CrossRef] [PubMed]
- Kapurniotu, A.; Gokce, O.; Bernhagen, J. The multitasking potential of alarmins and atypical chemokines. Front. Med. 2019, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, G.J.; Locati, M.; Mantovani, A.; Rot, A.; Thelen, M. The biochemistry and biology of the atypical chemokine receptors. Immunol. Lett. 2012, 145, 30–38. [Google Scholar] [CrossRef]
- Franic, D.; Zubcic, K.; Boban, M. Nuclear ubiquitin-proteasome pathways in proteostasis maintenance. Biomolecules 2021, 11, 54. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Blanchet, X.; Cesarek, K.; Brandt, J.; Herwald, H.; Teupser, D.; Kuchenhoff, H.; Karshovska, E.; Mause, S.F.; Siess, W.; Wasmuth, H.; et al. Inflammatory role and prognostic value of platelet chemokines in acute coronary syndrome. Thromb. Haemost. 2014, 112, 1277–1287. [Google Scholar] [CrossRef]
- Vajen, T.; Heinzmann, A.C.A.; Dickhout, A.; Zhao, Z.; Nagy, M.; Heemskerk, J.W.M.; Koenen, R.R. Laminar flow-based assays to investigate leukocyte recruitment on cultured vascular cells and adherent platelets. J. Vis. Exp. 2018, 57009. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dickhout, A.; Kaczor, D.M.; Heinzmann, A.C.A.; Brouns, S.L.N.; Heemskerk, J.W.M.; van Zandvoort, M.A.M.J.; Koenen, R.R. Rapid Internalization and Nuclear Translocation of CCL5 and CXCL4 in Endothelial Cells. Int. J. Mol. Sci. 2021, 22, 7332. https://doi.org/10.3390/ijms22147332
Dickhout A, Kaczor DM, Heinzmann ACA, Brouns SLN, Heemskerk JWM, van Zandvoort MAMJ, Koenen RR. Rapid Internalization and Nuclear Translocation of CCL5 and CXCL4 in Endothelial Cells. International Journal of Molecular Sciences. 2021; 22(14):7332. https://doi.org/10.3390/ijms22147332
Chicago/Turabian StyleDickhout, Annemiek, Dawid M. Kaczor, Alexandra C. A. Heinzmann, Sanne L. N. Brouns, Johan W. M. Heemskerk, Marc A. M. J. van Zandvoort, and Rory R. Koenen. 2021. "Rapid Internalization and Nuclear Translocation of CCL5 and CXCL4 in Endothelial Cells" International Journal of Molecular Sciences 22, no. 14: 7332. https://doi.org/10.3390/ijms22147332