
Triple Culture of Primary Human Osteoblasts, Osteoclasts and Osteocytes as an In Vitro Bone Model


Abstract
:1. Introduction
2. Results
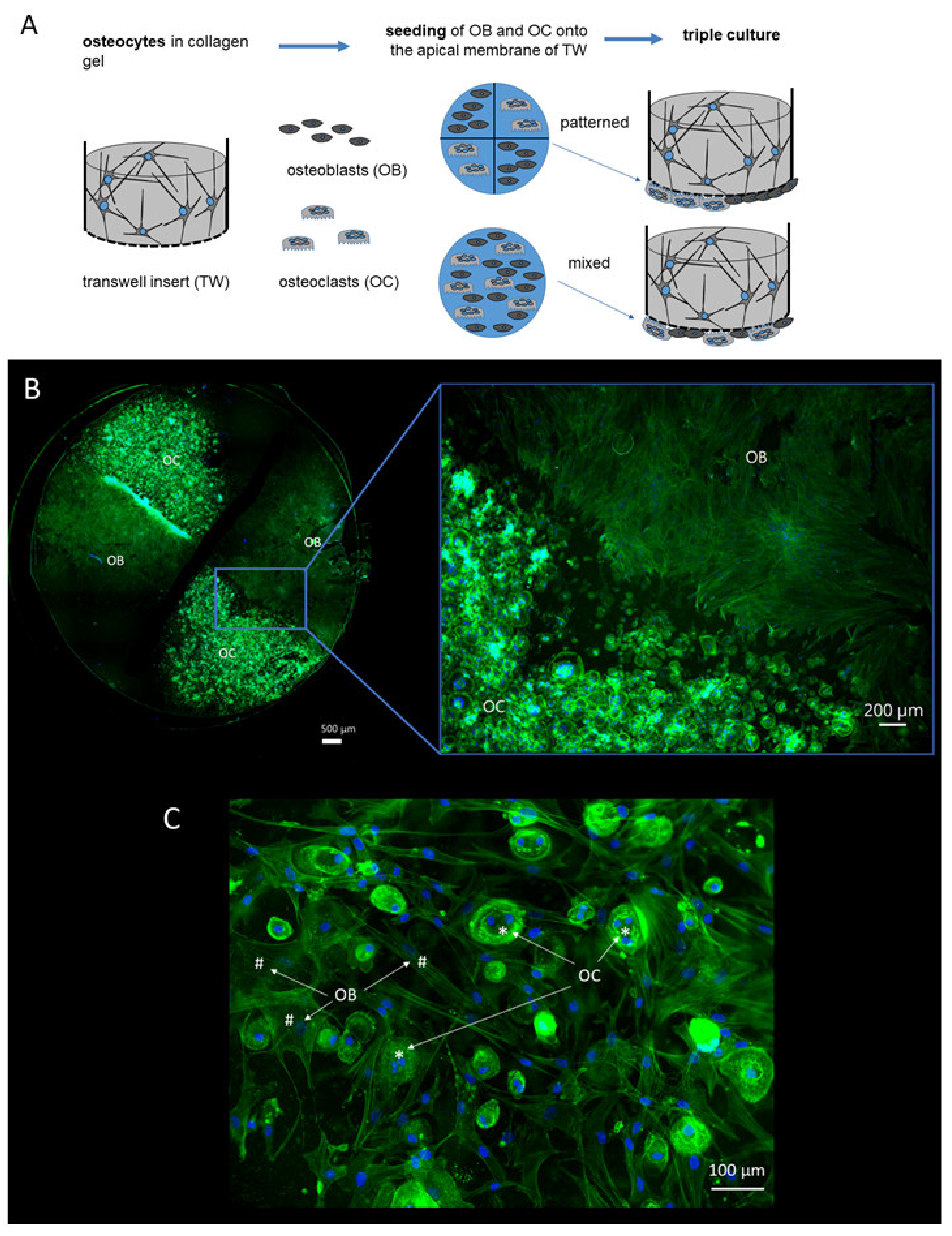
2.1. Triple Culture Setup
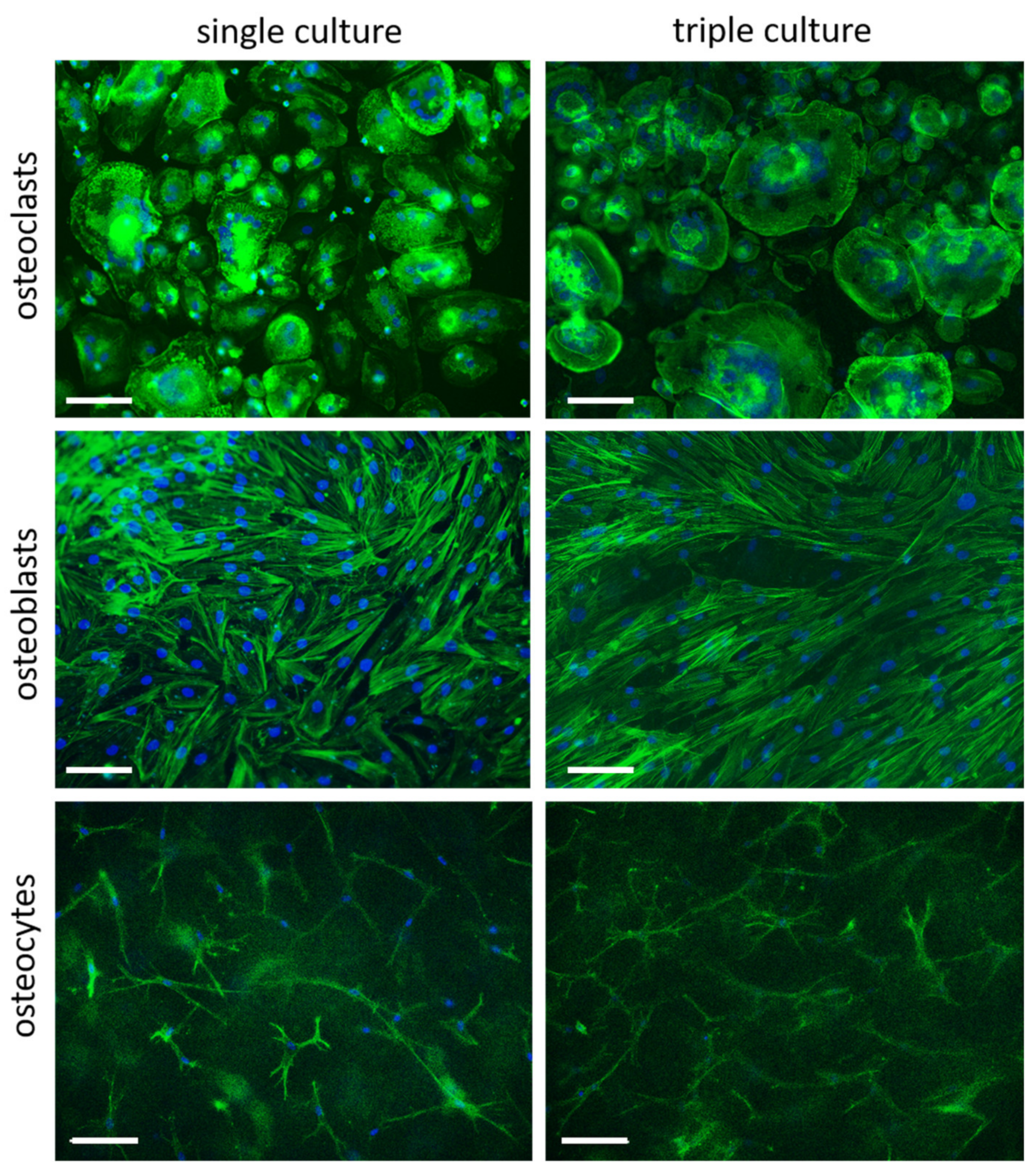
2.2. Cell Morphology in Separated Triple Cultures and Single Cultures
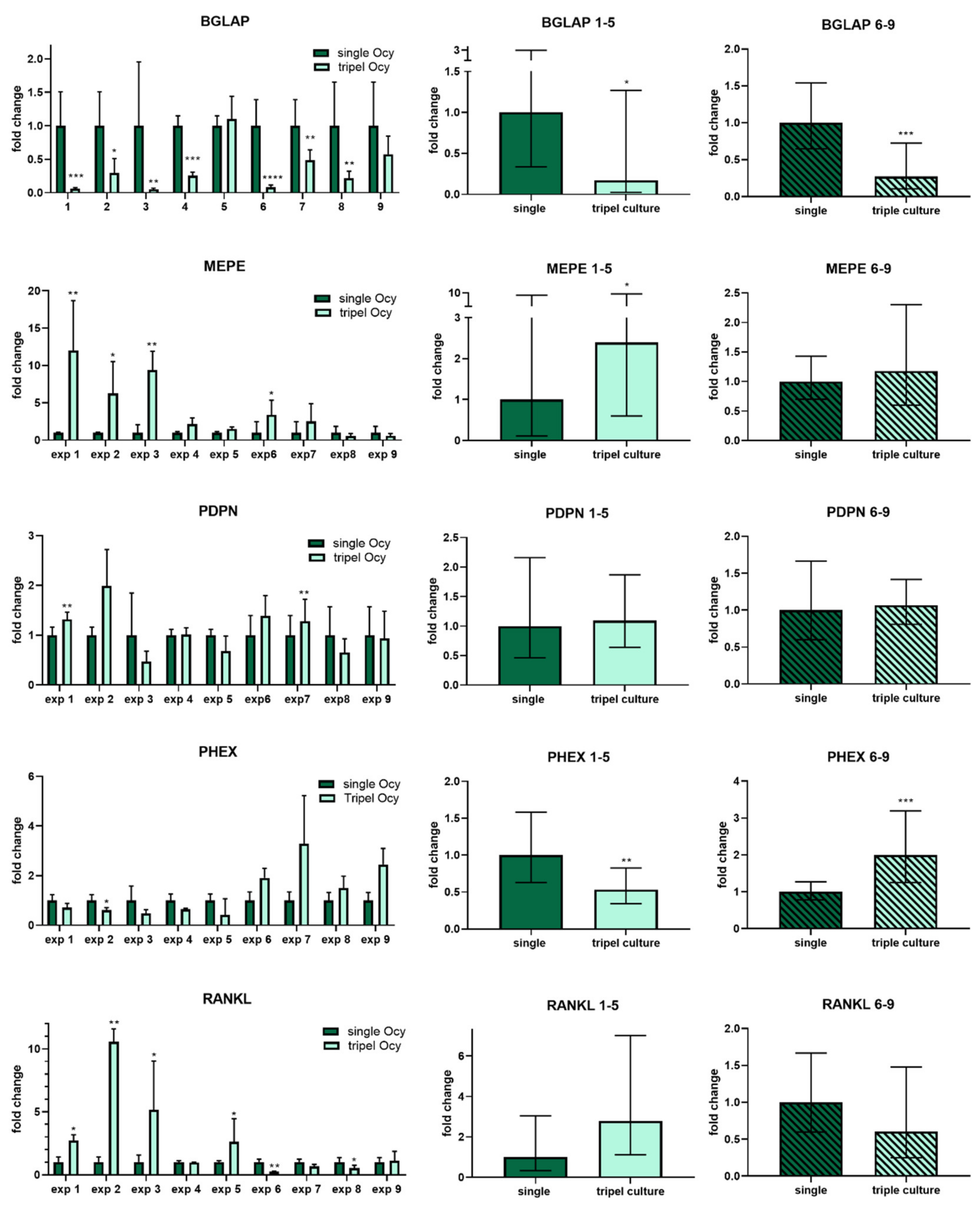
2.3. Osteocytes in Triple Culture
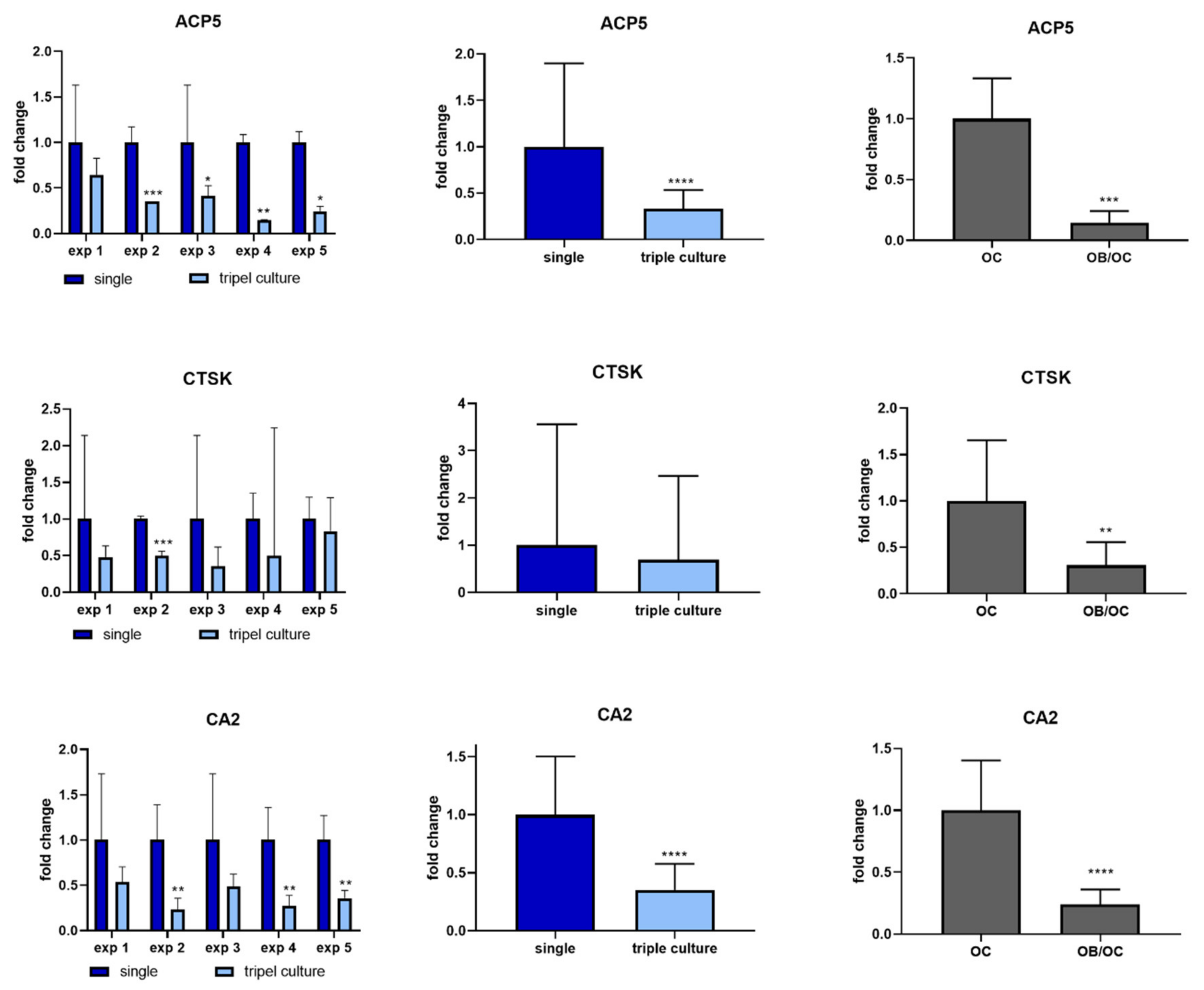
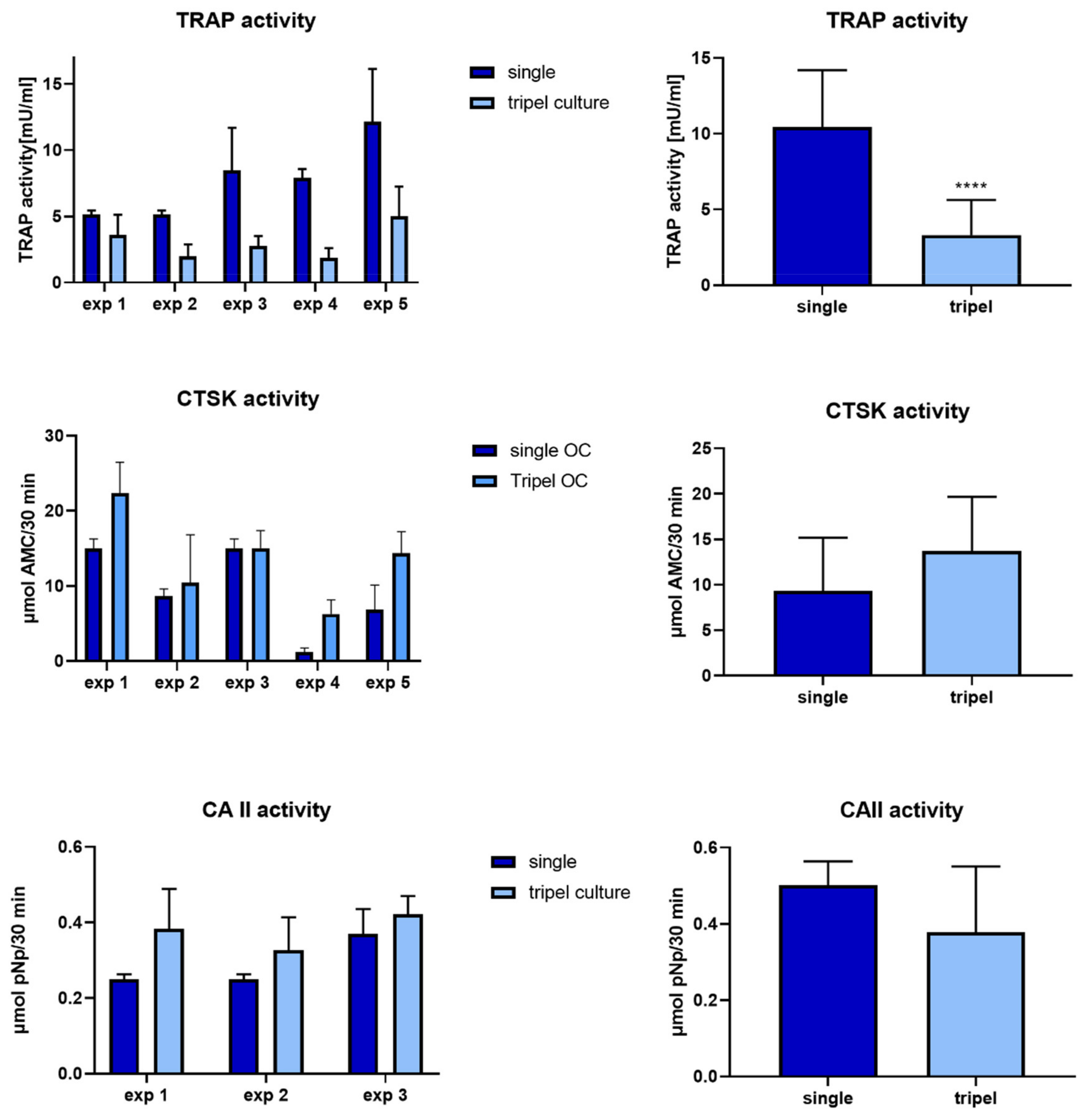
2.4. Osteoclasts in Triple Culture
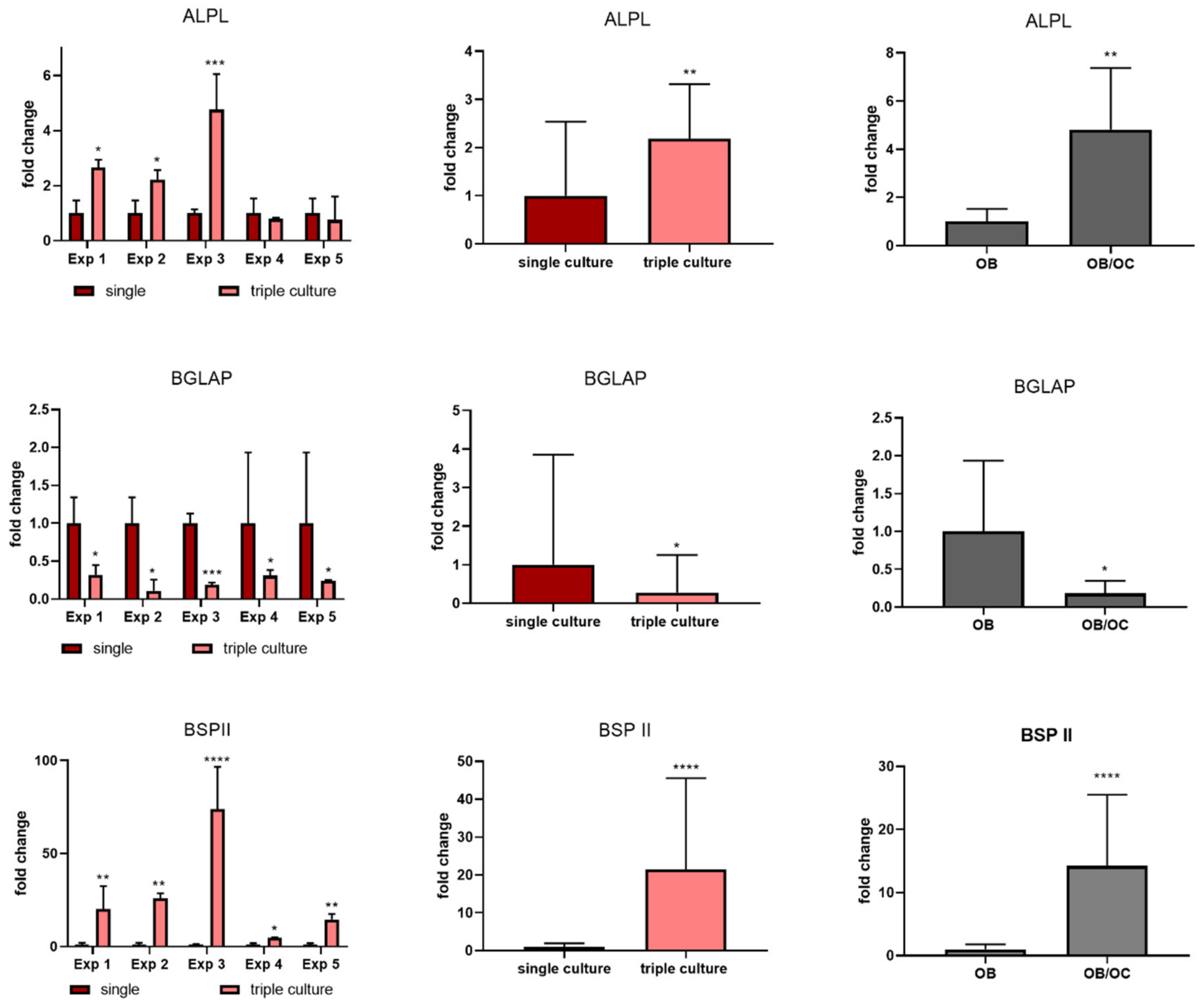
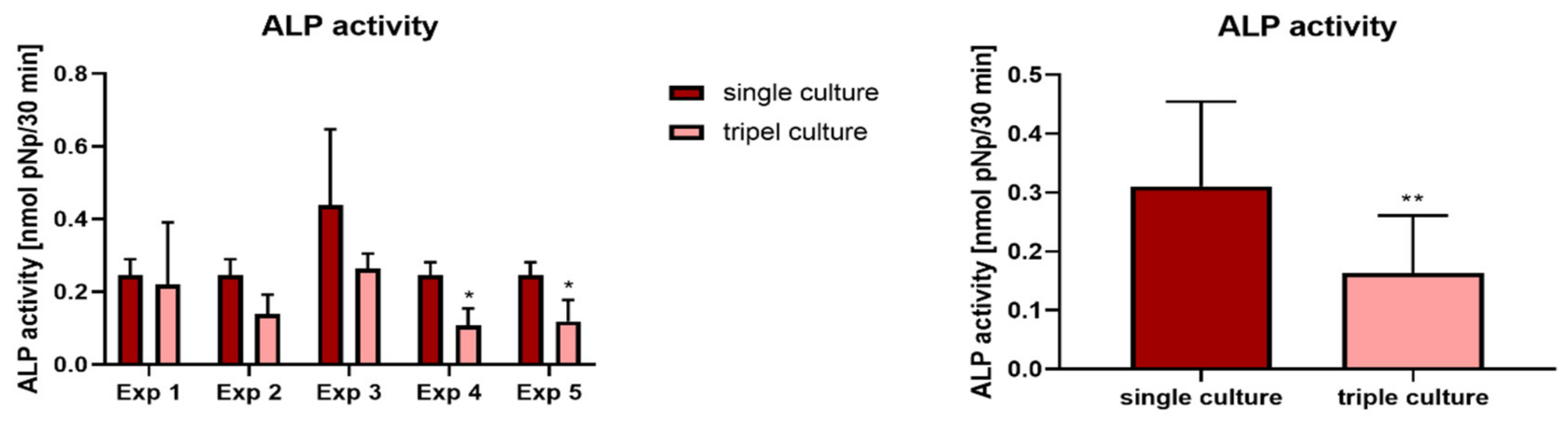
2.5. Osteoblasts in Triple Culture
3. Discussion
4. Materials and Methods
4.1. Cell Isolation and Differentiation
4.1.1. Osteoblasts
4.1.2. Osteocytes
4.1.3. Osteoclasts
4.2. Triple Cultures of Osteocytes, Osteoblasts and Osteoclasts
4.3. RNA Isolation, Preparation of cDNA and PCR
4.4. Analysis of Cell-Specific Enzyme Activities
4.5. Fluorescence Microscopy
4.6. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Detsch, R.; Boccaccini, A.R. The Role of Osteoclasts in Bone Tissue Engineering. J. Tissue Eng. Regen. Med. 2015, 9, 1133–1149. [Google Scholar] [CrossRef]
- Prideaux, M.; Findlay, D.M.; Atkins, G.J. Osteocytes: The Master Cells in Bone Remodelling. Curr. Opin. Pharmacol. 2016, 28, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, C.; Colangelo, L.; Santori, R.; Renella, M.; Mastrantonio, M.; Minisola, S.; Pepe, J. The Interplay Between Bone and Glucose Metabolism. Front. Endocrinol. 2020, 11, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kular, J.; Tickner, J.; Chim, S.M.; Xu, J. An Overview of the Regulation of Bone Remodelling at the Cellular Level. Clin. Biochem. 2012, 45, 863–873. [Google Scholar] [CrossRef]
- Terpos, E.; Ntanasis-Stathopoulos, I.; Dimopoulos, M.A. Myeloma Bone Disease: From Biology Findings to Treatment Approaches. Blood 2019, 133, 1534–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, K.; Kobayashi, Y.; Koide, M.; Uehara, S.; Okamoto, M.; Ishihara, A.; Kayama, T.; Saito, M.; Marumo, K. The Regulation of Bone Metabolism and Disorders by Wnt Signaling. Int. J. Mol. Sci. 2019, 20, 5525. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Ehnert, S.; Rouß, M.; Häussling, V.; Aspera-Werz, R.H.; Chen, T.; Nussler, A.K. From the Clinical Problem to the Basic Research-Co-Culture Models of Osteoblasts and Osteoclasts. Int. J. Mol. Sci. 2018, 19, 2284. [Google Scholar] [CrossRef] [Green Version]
- Borciani, G.; Montalbano, G.; Baldini, N.; Cerqueni, G.; Vitale-Brovarone, C.; Ciapetti, G. Co–Culture Systems of Osteoblasts and Osteoclasts: Simulating in Vitro Bone Remodeling in Regenerative Approaches. Acta Biomater. 2020, 108, 22–45. [Google Scholar] [CrossRef]
- Prideaux, M.; Schutz, C.; Wijenayaka, A.R.; Findlay, D.M.; Campbell, D.G.; Solomon, L.B.; Atkins, G.J. Isolation of Osteocytes from Human Trabecular Bone. Bone 2016, 88, 64–72. [Google Scholar] [CrossRef]
- Prideaux, M.; Stern, A.R.; Bonewald, L.F. Isolation of Murine and Human Osteocytes. In Osteoporosis and Osteoarthritis; van Wijnen, A.J., Ganshina, M.S., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2021; Volume 2221, pp. 3–13. ISBN 978-1-07-160988-0. [Google Scholar]
- Bernhardt, A.; Wolf, S.; Weiser, E.; Vater, C.; Gelinsky, M. An Improved Method to Isolate Primary Human Osteocytes from Bone. Biomed. Tech. 2019. [Google Scholar] [CrossRef]
- Li, B.; Lu, D.; Chen, Y.; Zhao, M.; Zuo, L. Unfractionated Heparin Promotes Osteoclast Formation in Vitro by Inhibiting Osteoprotegerin Activity. Int. J. Mol. Sci. 2016, 17, 613. [Google Scholar] [CrossRef] [Green Version]
- Hayashida, C.; Ito, J.; Nakayachi, M.; Okayasu, M.; Ohyama, Y.; Hakeda, Y.; Sato, T. Osteocytes Produce Interferon-β as a Negative Regulator of Osteoclastogenesis. J. Biol. Chem. 2014, 289, 11545–11555. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Zhang, Y.K.Y.; Harris, S.; Ahuja, S.S.; Bonewald, L.F. MLO-Y4 Osteocyte-like Cells Support Osteoclast Formation and Activation. J. Bone Miner. Res. 2002, 17, 2068–2079. [Google Scholar] [CrossRef]
- Honma, M.; Ikebuchi, Y.; Kariya, Y.; Hayashi, M.; Hayashi, N.; Aoki, S.; Suzuki, H. RANKL Subcellular Trafficking and Regulatory Mechanisms in Osteocytes: Rankl Subcellular Trafficking in Osteocytes. J. Bone Miner. Res. 2013, 28, 1936–1949. [Google Scholar] [CrossRef]
- Bernhardt, A.; Österreich, V.; Gelinsky, M. Three-Dimensional Co-Culture of Primary Human Osteocytes and Mature Human Osteoclasts in Collagen Gels. Tissue Eng. Part A 2020, 26, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.F.; Saunders, M.M.; Shingle, D.L.; Cimbala, J.M.; Zhou, Z.; Donahue, H.J. Mechanically Stimulated Osteocytes Regulate Osteoblastic Activity via Gap Junctions. Am. J. Physiol. Cell Physiol. 2007, 292, C545–C552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, M.; Evans, B.A.J.; Riccardi, D.; Evans, S.L.; Ralphs, J.R.; Dillingham, C.M.; Mason, D.J. A New Method to Investigate How Mechanical Loading of Osteocytes Controls Osteoblasts. Front. Endocrinol. 2014, 5, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jähn, K.; Mason, D.; Ralphs, J.; Evans, B.; Archer, C.; Richards, R.; Stoddart, M. Phenotype and Viability of MLO-Y4 Cells Is Maintained by TGFβ3 in a Serum-Dependent Manner within a 3D-Co-Culture with MG-63 Cells. Int. J. Mol. Sci. 2018, 19, 1932. [Google Scholar] [CrossRef] [Green Version]
- Skottke, J.; Gelinsky, M.; Bernhardt, A. In Vitro Co-Culture Model of Primary Human Osteoblasts and Osteocytes in Collagen Gels. Int. J. Mol. Sci. 2019, 20, 1998. [Google Scholar] [CrossRef] [Green Version]
- Vis, M.A.M.; Ito, K.; Hofmann, S. Impact of Culture Medium on Cellular Interactions in in Vitro Co-Culture Systems. Front. Bioeng. Biotechnol. 2020, 8, 911. [Google Scholar] [CrossRef]
- Clarke, M.S.F.; Sundaresan, A.; Vanderburg, C.R.; Banigan, M.G.; Pellis, N.R. A Three-Dimensional Tissue Culture Model of Bone Formation Utilizing Rotational Co-Culture of Human Adult Osteoblasts and Osteoclasts. Acta Biomater. 2013, 9, 7908–7916. [Google Scholar] [CrossRef]
- Pagani, S.; Torricelli, P.; Veronesi, F.; Salamanna, F.; Cepollaro, S.; Fini, M. An Advanced Tri-Culture Model to Evaluate the Dynamic Interplay among Osteoblasts, Osteoclasts, and Endothelial Cells. J. Cell. Physiol. 2018, 233, 291–301. [Google Scholar] [CrossRef]
- Grémare, A.; Aussel, A.; Bareille, R.; Paiva dos Santos, B.; Amédée, J.; Thébaud, N.B.; Le Nihouannen, D. A Unique Triculture Model to Study Osteoblasts, Osteoclasts, and Endothelial Cells. Tissue Eng. Part C Methods 2019, 25, 421–432. [Google Scholar] [CrossRef]
- Wein, F.; Bruinink, A. Human Triple Cell Co-Culture for Evaluation of Bone Implant Materials. Integr. Biol. 2013, 5, 703–711. [Google Scholar] [CrossRef]
- Plotkin, L.I.; Bruzzaniti, A. Molecular Signaling in Bone Cells: Regulation of Cell Differentiation and Survival. Adv. Protein Chem. Struct. Biol. 2019, 116, 237–281. [Google Scholar] [CrossRef] [PubMed]
- Scheinpflug, J.; Pfeiffenberger, M.; Damerau, A.; Schwarz, F.; Textor, M.; Lang, A.; Schulze, F. Journey into Bone Models: A Review. Genes 2018, 9, 247. [Google Scholar] [CrossRef] [Green Version]
- George, E.L.; Truesdell, S.L.; York, S.L.; Saunders, M.M. Lab-on-a-Chip Platforms for Quantification of Multicellular Interactions in Bone Remodeling. Exp. Cell Res. 2018, 365, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Truesdell, S.L.; George, E.L.; Van Vranken, C.C.; Saunders, M.M. A Lab-On-A-Chip Platform for Stimulating Osteocyte Mechanotransduction and Analyzing Functional Outcomes of Bone Remodeling. J. Vis. Exp. 2020, 61076. [Google Scholar] [CrossRef]
- Middleton, K.; Al-Dujaili, S.; Mei, X.; Günther, A.; You, L. Microfluidic Co-Culture Platform for Investigating Osteocyte-Osteoclast Signalling during Fluid Shear Stress Mechanostimulation. J. Biomech. 2017, 59, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Choudhary, S.; Mannion, C.; Kissin, Y.; Zilberberg, J.; Lee, W.Y. Ex Vivo Construction of Human Primary 3D-Networked Osteocytes. Bone 2017, 105, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Atkins, G.J.; Welldon, K.J.; Holding, C.A.; Haynes, D.R.; Howie, D.W.; Findlay, D.M. The Induction of a Catabolic Phenotype in Human Primary Osteoblasts and Osteocytes by Polyethylene Particles. Biomaterials 2009, 30, 3672–3681. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhou, Q.; Kang, H.; Yan, Y.; Qian, N.; Li, C.; Wang, F.; Yang, K.; Deng, L.; Qi, J. High Mineralization Capacity of IDG-SW3 Cells in 3D Collagen Hydrogel for Bone Healing in Estrogen-Deficient Mice. Front. Bioeng. Biotechnol. 2020, 8, 864. [Google Scholar] [CrossRef]
- Nasello, G.; Alamán-Díez, P.; Schiavi, J.; Pérez, M.Á.; McNamara, L.; García-Aznar, J.M. Primary Human Osteoblasts Cultured in a 3D Microenvironment Create a Unique Representative Model of Their Differentiation Into Osteocytes. Front. Bioeng. Biotechnol. 2020, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- Uchihashi, K.; Aoki, S.; Matsunobu, A.; Toda, S. Osteoblast Migration into Type I Collagen Gel and Differentiation to Osteocyte-like Cells within a Self-Produced Mineralized Matrix: A Novel System for Analyzing Differentiation from Osteoblast to Osteocyte. Bone 2013, 52, 102–110. [Google Scholar] [CrossRef] [PubMed]
- James Kirkpatrick, C.; Fuchs, S.; Iris Hermanns, M.; Peters, K.; Unger, R.E. Cell Culture Models of Higher Complexity in Tissue Engineering and Regenerative Medicine. Biomaterials 2007, 28, 5193–5198. [Google Scholar] [CrossRef] [PubMed]
- Ehnert, S.; Rinderknecht, H.; Aspera-Werz, R.H.; Häussling, V.; Nussler, A.K. Use of in Vitro Bone Models to Screen for Altered Bone Metabolism, Osteopathies, and Fracture Healing: Challenges of Complex Models. Arch. Toxicol. 2020, 94, 3937–3958. [Google Scholar] [CrossRef] [PubMed]
- Kreja, L.; Liedert, A.; Schmidt, C.; Claes, L.; Ignatius, A. Influence of Receptor Activator of Nuclear Factor (NF)-KappaB Ligand (RANKL), Macrophage-Colony Stimulating Factor (M-CSF) and Fetal Calf Serum on Human Osteoclast Formation and Activity. J. Mol. Histol. 2007, 38, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Honma, M.; Ikebuchi, Y.; Kariya, Y.; Suzuki, H. Establishment of Optimized in Vitro Assay Methods for Evaluating Osteocyte Functions. J. Bone Miner. Metab. 2015, 33, 73–84. [Google Scholar] [CrossRef]
- Bernhardt, A.; Thieme, S.; Domaschke, H.; Springer, A.; Rösen-Wolff, A.; Gelinsky, M. Crosstalk of Osteoblast and Osteoclast Precursors on Mineralized Collagen--towards an in Vitro Model for Bone Remodeling. J. Biomed. Mater. Res. A 2010, 95, 848–856. [Google Scholar] [CrossRef]
- Schulze, S.; Wehrum, D.; Dieter, P.; Hempel, U. A Supplement-Free Osteoclast-Osteoblast Co-Culture for Pre-Clinical Application. J. Cell. Physiol. 2018, 233, 4391–4400. [Google Scholar] [CrossRef]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-Hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for Osteocyte Regulation of Bone Homeostasis through RANKL Expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef]
- Steller, D.; Scheibert, A.; Sturmheit, T.; Hakim, S.G. Establishment and Validation of an in Vitro Co-Culture Model for Oral Cell Lines Using Human PBMC-Derived Osteoclasts, Osteoblasts, Fibroblasts and Keratinocytes. Sci. Rep. 2020, 10, 16861. [Google Scholar] [CrossRef]
- Holm, E.; Aubin, J.E.; Hunter, G.K.; Beier, F.; Goldberg, H.A. Loss of Bone Sialoprotein Leads to Impaired Endochondral Bone Development and Mineralization. Bone 2015, 71, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Licini, C.; Vitale-Brovarone, C.; Mattioli-Belmonte, M. Collagen and Non-Collagenous Proteins Molecular Crosstalk in the Pathophysiology of Osteoporosis. Cytokine Growth Factor Rev. 2019, 49, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.-H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Walker, E.C.; McGregor, N.E.; Poulton, I.J.; Pompolo, S.; Allan, E.H.; Quinn, J.M.W.; Gillespie, M.T.; Martin, T.J.; Sims, N.A. Cardiotrophin-1 Is an Osteoclast-Derived Stimulus of Bone Formation Required for Normal Bone Remodeling. J. Bone Miner. Res. 2008, 23, 2025–2032. [Google Scholar] [CrossRef] [PubMed]
- Kreja, L.; Brenner, R.E.; Tautzenberger, A.; Liedert, A.; Friemert, B.; Ehrnthaller, C.; Huber-Lang, M.; Ignatius, A. Non-Resorbing Osteoclasts Induce Migration and Osteogenic Differentiation of Mesenchymal Stem Cells. J. Cell. Biochem. 2010, 109, 347–355. [Google Scholar] [CrossRef]
- Heinemann, C.; Heinemann, S.; Worch, H.; Hanke, T. Development of an Osteoblast/Osteoclast Co-Culture Derived by Human Bone Marrow Stromal Cells and Human Monocytes for Biomaterials Testing. Eur. Cell. Mater. 2011, 21, 80–93. [Google Scholar] [CrossRef]
- Quarles, L.D. FGF23, PHEX, and MEPE Regulation of Phosphate Homeostasis and Skeletal Mineralization. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E1–E9. [Google Scholar] [CrossRef]
- Paic, F.; Igwe, J.C.; Nori, R.; Kronenberg, M.S.; Franceschetti, T.; Harrington, P.; Kuo, L.; Shin, D.-G.; Rowe, D.W.; Harris, S.E.; et al. Identification of Differentially Expressed Genes between Osteoblasts and Osteocytes. Bone 2009, 45, 682–692. [Google Scholar] [CrossRef] [Green Version]
- Prideaux, M.; Loveridge, N.; Pitsillides, A.A.; Farquharson, C. Extracellular Matrix Mineralization Promotes E11/Gp38 Glycoprotein Expression and Drives Osteocytic Differentiation. PLoS ONE 2012, 7, e36786. [Google Scholar] [CrossRef] [Green Version]
- Welldon, K.J.; Findlay, D.M.; Evdokiou, A.; Ormsby, R.T.; Atkins, G.J. Calcium Induces Pro-Anabolic Effects on Human Primary Osteoblasts Associated with Acquisition of Mature Osteocyte Markers. Mol. Cell. Endocrinol. 2013, 376, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Song, J.; Ma, H. An in Vitro Experimental Insight into the Osteoblast Responses to Vitamin D3 and Its Metabolites. Pharmacology 2018, 101, 225–235. [Google Scholar] [CrossRef]
- Bernhardt, A.; Schumacher, M.; Gelinsky, M. Formation of Osteoclasts on Calcium Phosphate Bone Cements and Polystyrene Depends on Monocyte Isolation Conditions. Tissue Eng. Part C Methods 2015, 21, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, A.; Bacova, J.; Gbureck, U.; Gelinsky, M. Influence of Cu2+ on Osteoclast Formation and Activity In Vitro. Int. J. Mol. Sci. 2021, 22, 2451. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, A.; Koperski, K.; Schumacher, M.; Gelinsky, M. Relevance of Osteoclast-Specific Enzyme Activities in Cell-Based in Vitro Resorption Assays. Eur. Cell. Mater. 2017, 33, 28–42. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exp. No. | Osteoblasts (OB)/Osteocytes (Ocy) | Osteoclasts (OC) | Seeding OB/OC |
|---|---|---|---|
| 1 | male, 65 y | PBMC donor 1 | patterned |
| 2 | male, 65 y | PBMC donor 2 | patterned |
| 3 | female, 56 y | PBMC donor 1 | patterned |
| 4 | female, 56 y | PBMC donor 3 | patterned |
| 5 | female, 56 y | PBMC donor 4 | patterned |
| 6 | female, 52 y | PBMC donor 5 | mixed |
| 7 | female, 52 y | PBMC donor 6 | mixed |
| 8 | female, 56 y | PBMC donor 5 | mixed |
| 9 | female, 56 y | PBMC donor 6 | mixed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernhardt, A.; Skottke, J.; von Witzleben, M.; Gelinsky, M. Triple Culture of Primary Human Osteoblasts, Osteoclasts and Osteocytes as an In Vitro Bone Model. Int. J. Mol. Sci. 2021, 22, 7316. https://doi.org/10.3390/ijms22147316
Bernhardt A, Skottke J, von Witzleben M, Gelinsky M. Triple Culture of Primary Human Osteoblasts, Osteoclasts and Osteocytes as an In Vitro Bone Model. International Journal of Molecular Sciences. 2021; 22(14):7316. https://doi.org/10.3390/ijms22147316
Chicago/Turabian StyleBernhardt, Anne, Jasmin Skottke, Max von Witzleben, and Michael Gelinsky. 2021. "Triple Culture of Primary Human Osteoblasts, Osteoclasts and Osteocytes as an In Vitro Bone Model" International Journal of Molecular Sciences 22, no. 14: 7316. https://doi.org/10.3390/ijms22147316