
Fibroblast Growth Factor Type 1 Ameliorates High-Glucose-Induced Oxidative Stress and Neuroinflammation in Retinal Pigment Epithelial Cells and a Streptozotocin-Induced Diabetic Rat Model



Abstract
:1. Introduction
2. Results
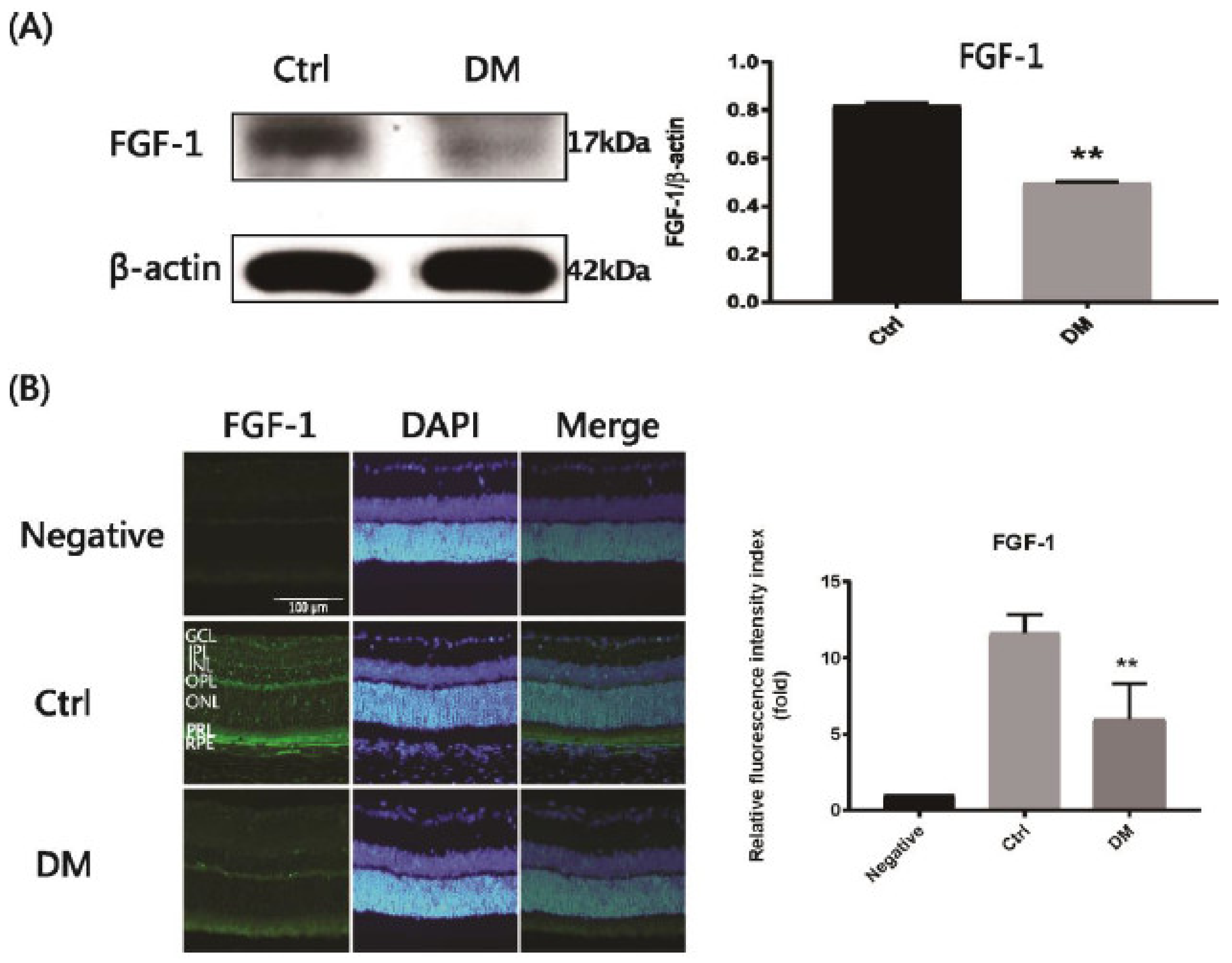
2.1. Suppression of Endogenous FGF-1 Levels in STZ-Induced Diabetic Rats
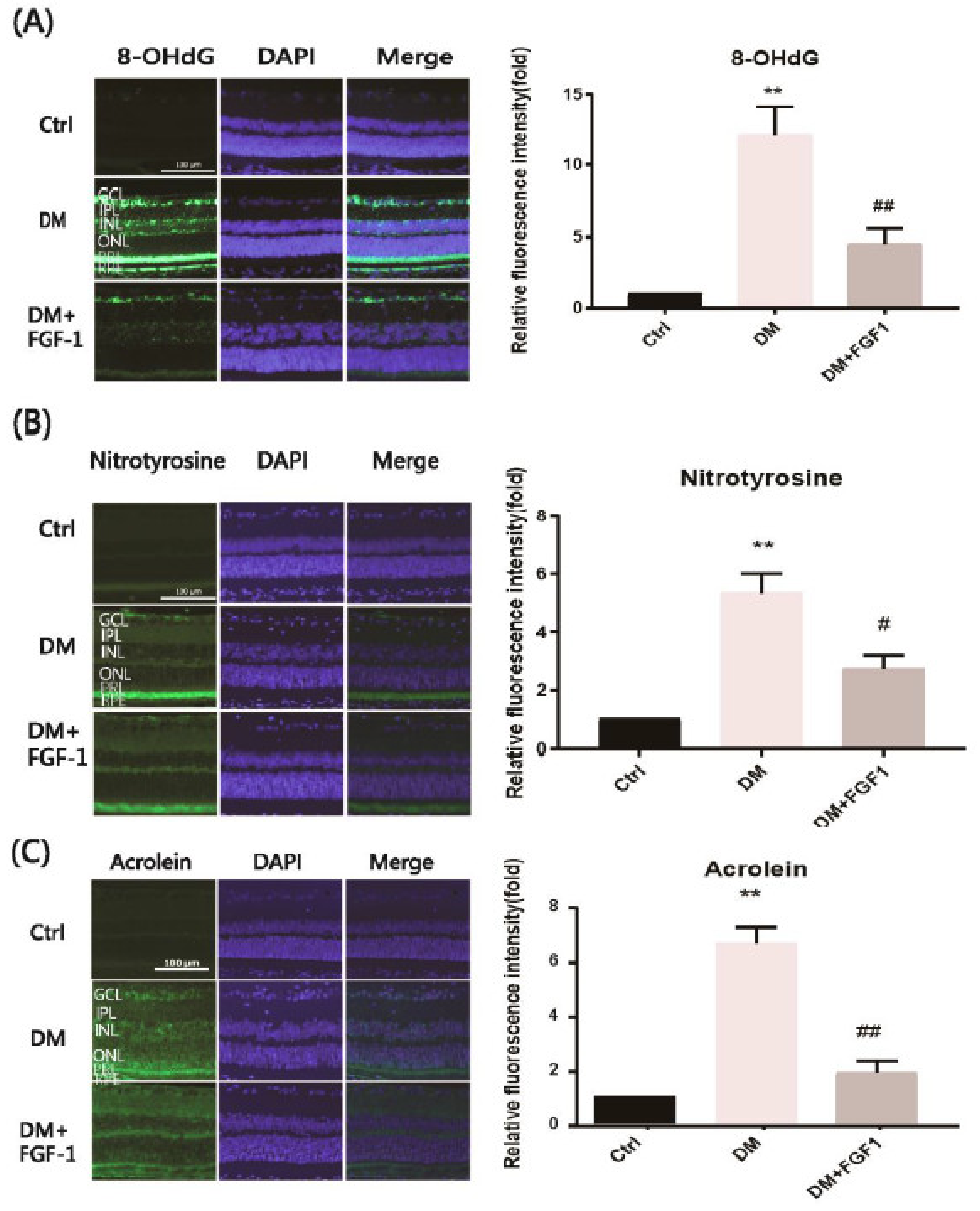
2.2. FGF-1 Treatment Prevents DR in STZ-Induced Diabetic Rats by Reducing Oxidative Stress
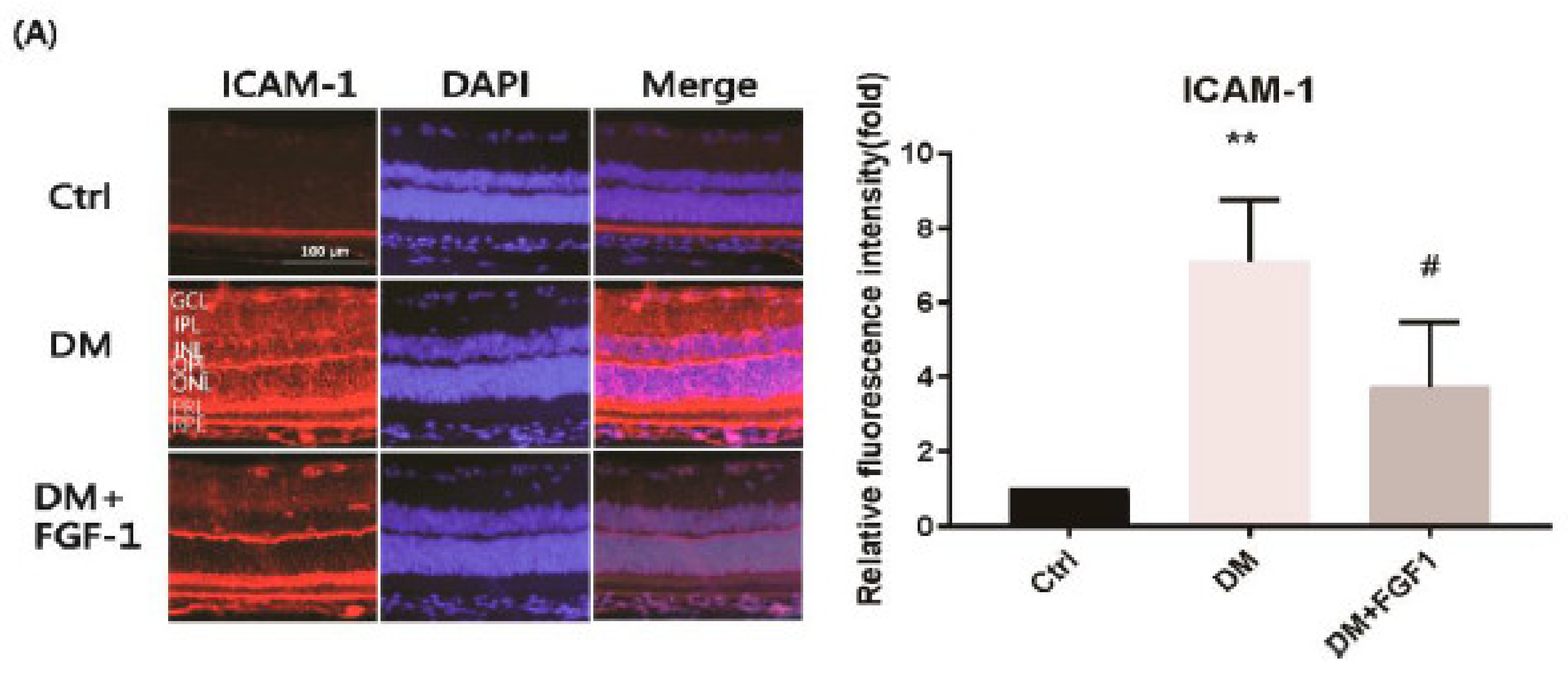
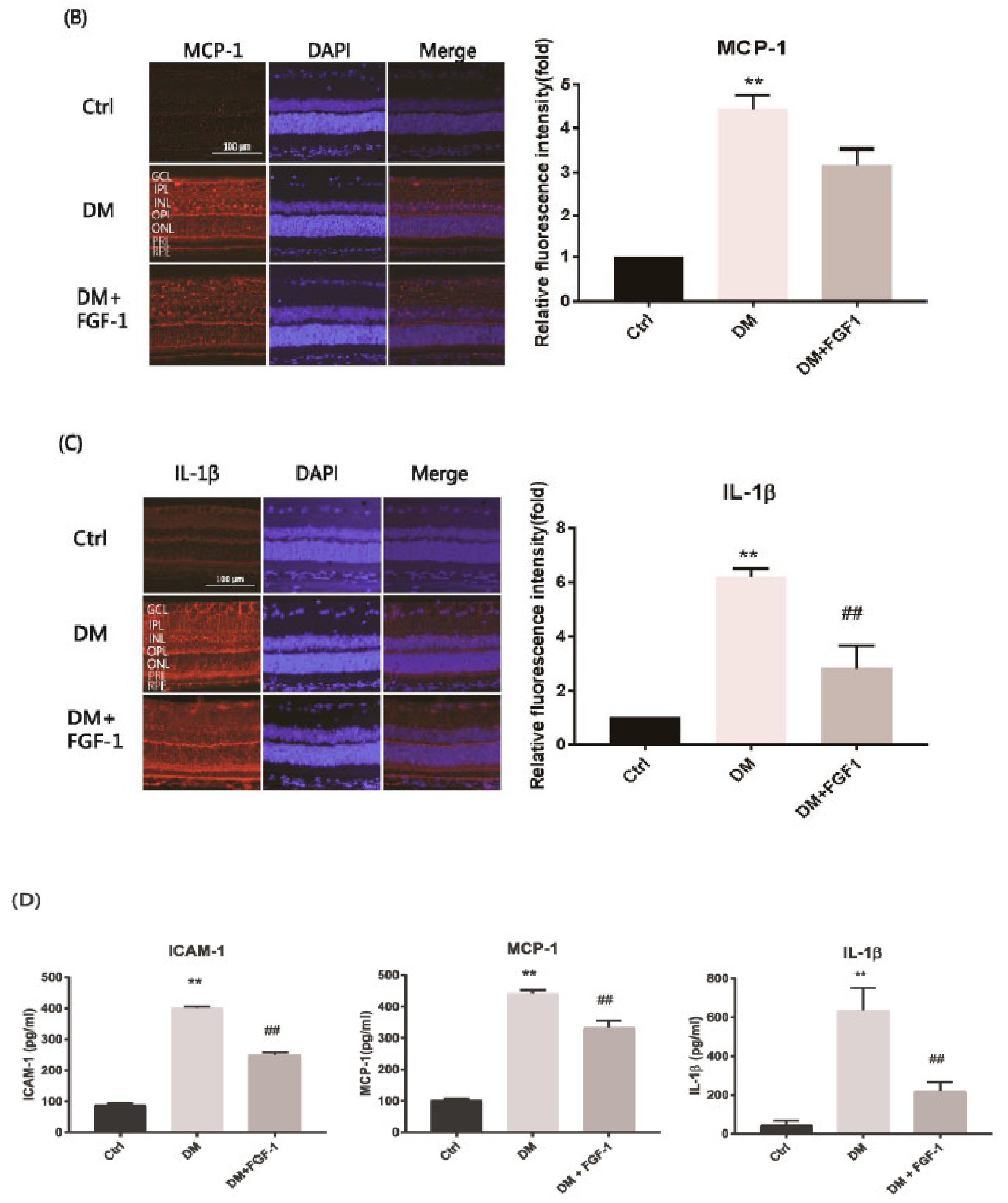
2.3. FGF-1 Treatment Attenuates Diabetes-Induced Inflammatory Mediators in the Retinas of STZ-Induced Diabetic Rats
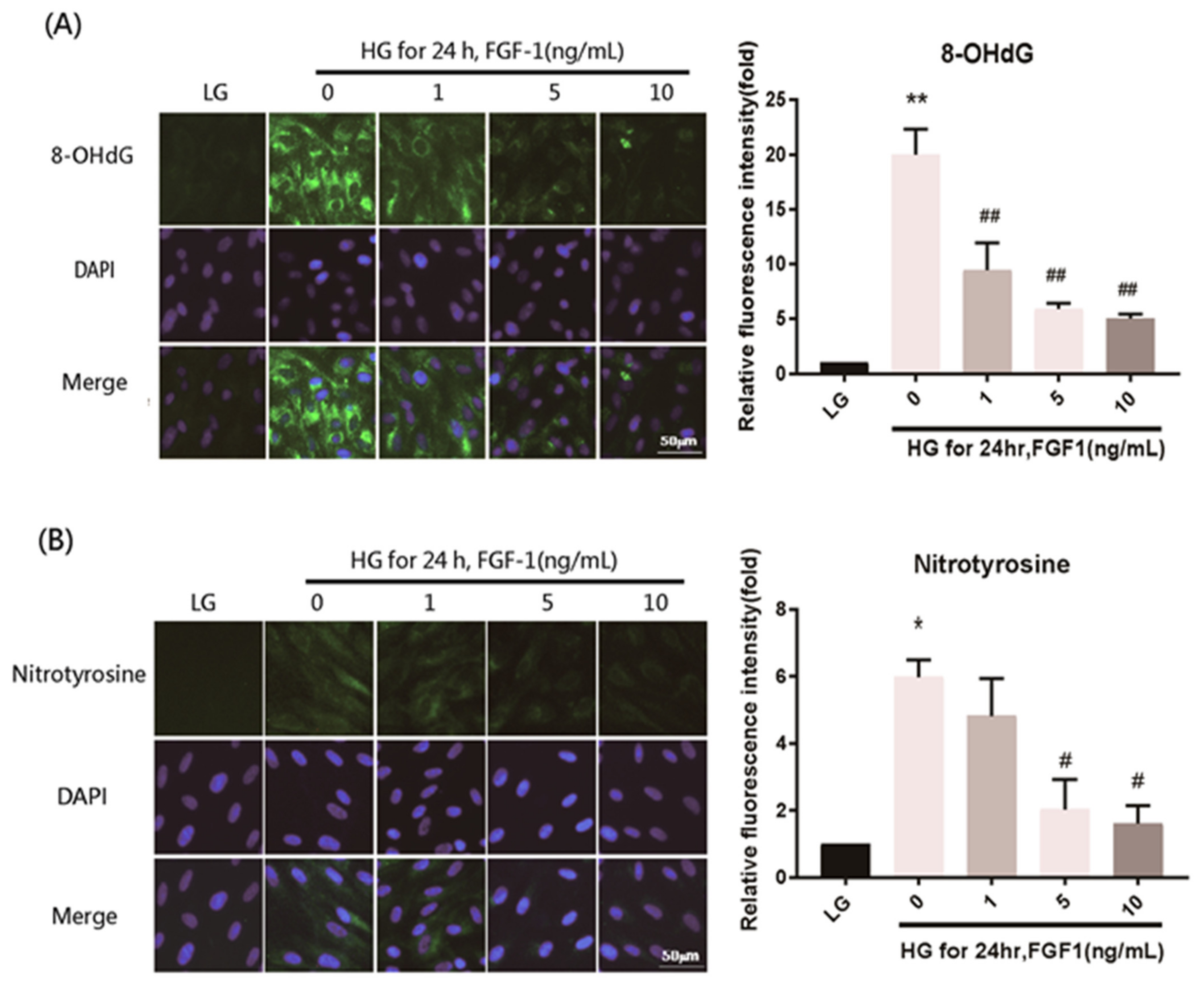
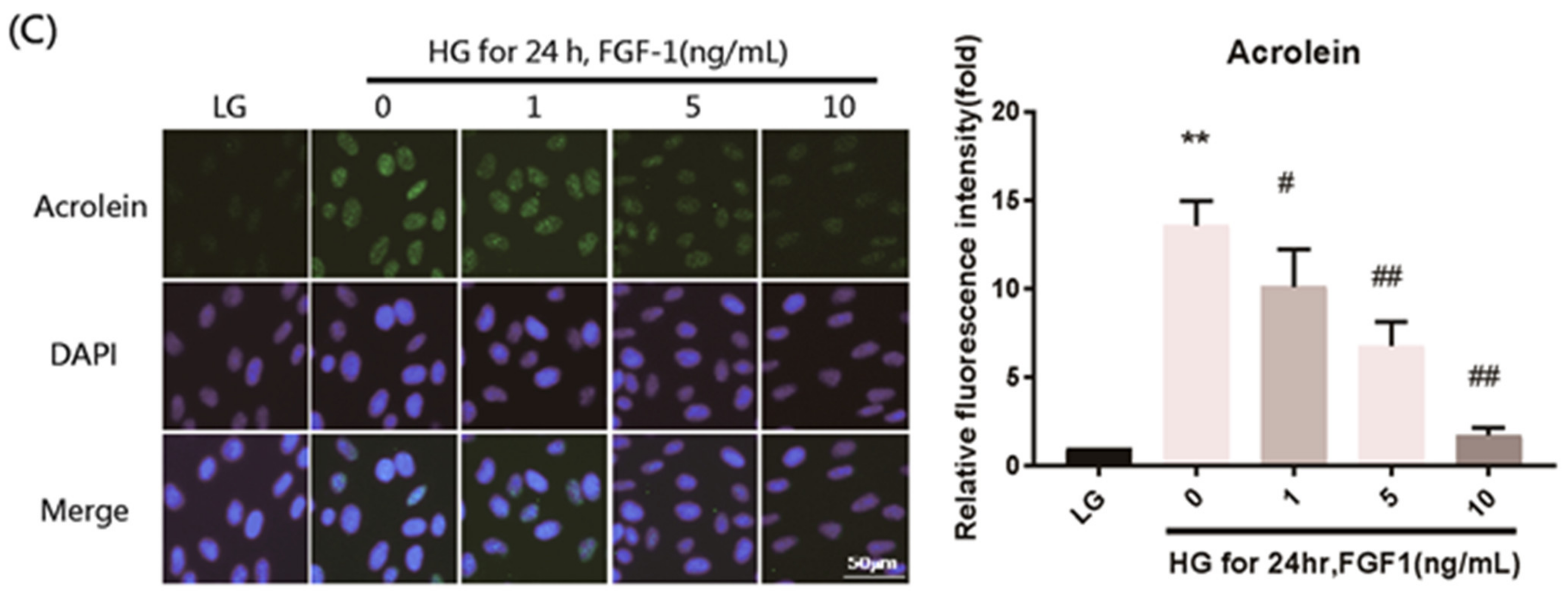
2.4. FGF-1 Treatment Ameliorates Oxidative Stress Damage in ARPE-19 Cells Exposed to Elevated Glucose Levels
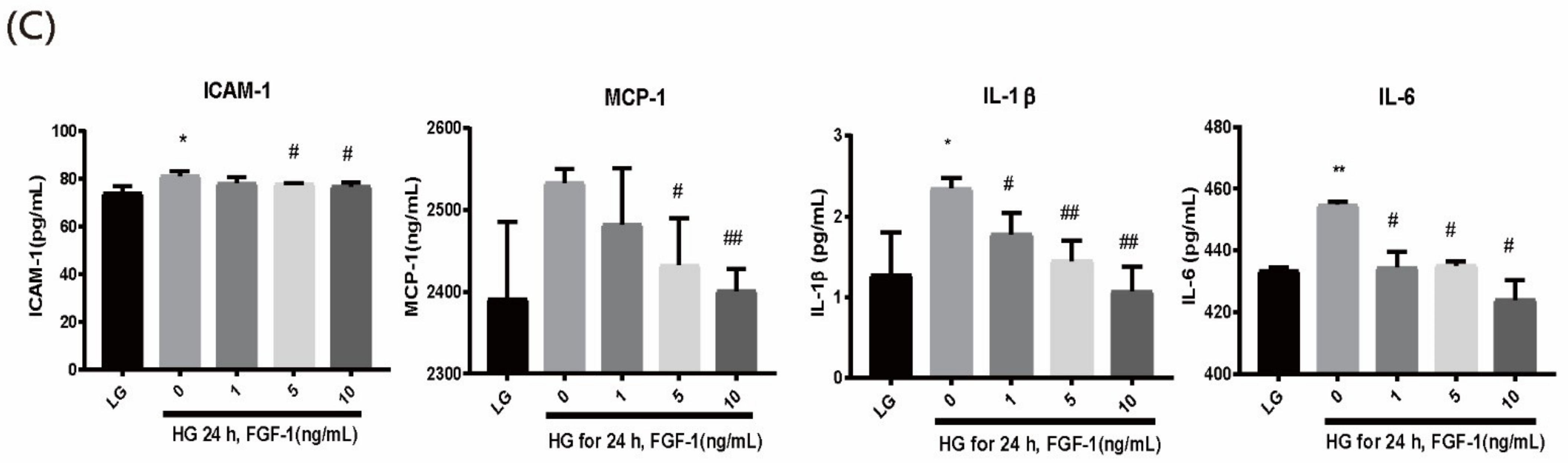
2.5. FGF-1 Treatment Alleviates High-Glucose-Stimulated Inflammatory Mediators in ARPE-19 Retinal Cells
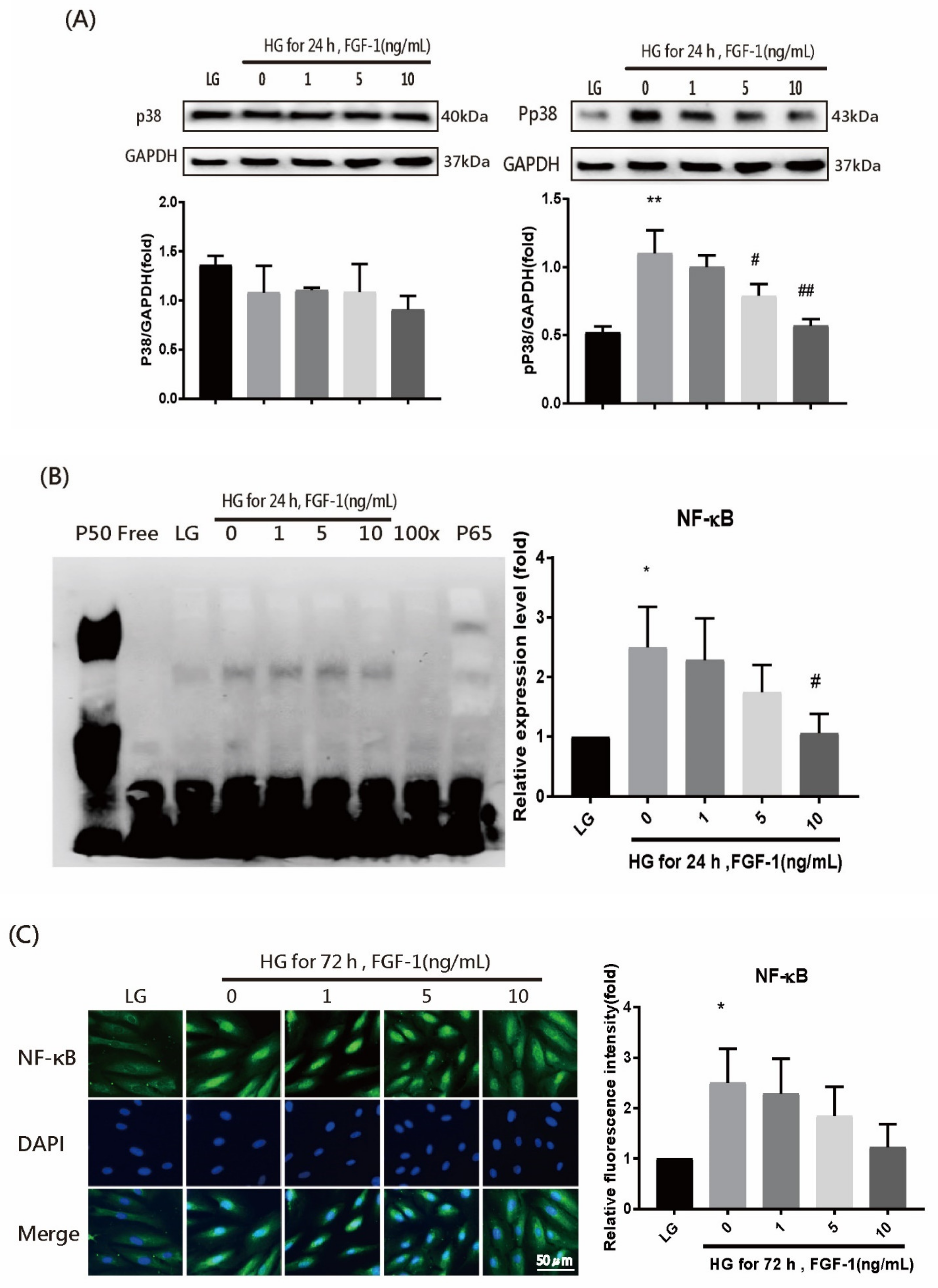
2.6. FGF-1 Suppresses the Activation of P38 and NF-κB Signaling in ARPE-19 Cells Incubated in High-Glucose Medium
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Animal Experiments
4.3. Ethics Statement
4.4. Cell Culture and Treatment
4.5. IF Detection of Oxidative Stress Markers and Inflammatory Mediators
4.6. Total RNA Extraction and Real-Time PCR Analysis
4.7. WB
4.8. ELISA
4.9. Nuclear Protein Extraction and NF-κB EMSA
4.10. IF Staining of NF-κB
4.11. Statistical Analyses
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Engelgau, M.M.; Geiss, L.S.; Saaddine, J.B.; Boyle, J.P.; Benjamin, S.M.; Gregg, E.W.; Tierney, E.F.; Rios-Burrows, N.; Mokdad, A.H.; Ford, E.S.; et al. The Evolving Diabetes Burden in the United States. Ann. Intern. Med. 2004, 140, 945–950. [Google Scholar] [CrossRef]
- Kobrin Klein, B.E. Overview of Epidemiologic Studies of Diabetic Retinopathy. Ophthalmic Epidemiol. 2007, 14, 179–183. [Google Scholar] [CrossRef]
- Madsen-Bouterse, S.A.; Kowluru, R.A. Oxidative stress and diabetic retinopathy: Pathophysiological mechanisms and treatment perspectives. Rev. Endocr. Metab. Disord. 2008, 9, 315–327. [Google Scholar] [CrossRef]
- Eisma, J.H.; Dulle, J.E.; Fort, P.E. Current knowledge on diabetic retinopathy from human donor tissues. World J. Diabetes 2015, 6, 312–320. [Google Scholar] [CrossRef]
- Kansora, M.B.; Goldhardt, R. Decision Making in Proliferative Diabetic Retinopathy Treatment. Curr. Ophthalmol. Rep. 2019, 7, 45–50. [Google Scholar] [CrossRef]
- Vaziri, K.; Schwartz, S.G.; Relhan, N.; Kishor, K.S.; Flynn, H.W., Jr. New Therapeutic Approaches in Diabetic Retinopathy. Rev. Diabet. Stud. 2015, 12, 196–210. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Bressler, N.M. Aflibercept, bevacizumab or ranibizumab for diabetic macular oedema: Recent clinically relevant findings from DRCR.net Protocol T. Curr. Opin. Ophthalmol. 2017, 28, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, R.; Cicinelli, M.V.; Bandello, F. Intravitreal Steroids in Diabetic Macular Edema. Dev. Ophthalmol. 2017, 60, 78–90. [Google Scholar]
- Shah, C.P.; Chen, C. Review of therapeutic advances in diabetic retinopathy. Ther. Adv. Endocrinol. Metab. 2011, 2, 39–53. [Google Scholar] [PubMed]
- El Annan, J.; Carvounis, P.E. Current management of vitreous hemorrhage due to proliferative diabetic retinopathy. Int. Ophthalmol. Clin. 2014, 54, 141–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.Y.; Zhao, K.K.; Li, J.K.; Rossmiller, B.; Zhao, P.Q. Four-port bimanual 23-gauge vitrectomy for diabetic tractional retinal detachment. Acta Ophthalmol. 2016, 94, 365–372. [Google Scholar] [CrossRef]
- Solomon, S.D.; Chew, E.; Duh, E.J.; Sobrin, L.; Sun, J.K.; VanderBeek, B.L.; Wykoff, C.C.; Gardner, T.W. Diabetic Retinopathy: A Position Statement by the American Diabetes Association. Diabetes Care 2017, 40, 412–418. [Google Scholar] [CrossRef] [Green Version]
- Giugliano, D.; Ceriello, A.; Paolisso, G. Oxidative stress and diabetic vascular complications. Diabetes Care 1996, 19, 257–267. [Google Scholar] [CrossRef]
- Rösen, P.; Nawroth, P.P.; King, G.; Möller, W.; Tritschler, H.J.; Packer, L. The role of oxidative stress in the onset and progression of diabetes and its complications: A summary of a Congress Series sponsored by UNESCO-MCBN, the American Diabetes Association and the German Diabetes Society. Diabetes Metab. Res. Rev. 2001, 17, 189–212. [Google Scholar] [CrossRef]
- Stitt, A.W.; Frizzell, N.; Thorpe, S.R. Advanced glycation and advanced lipoxidation: Possible role in initiation and progression of diabetic retinopathy. Curr. Pharm. Des. 2004, 10, 3349–3360. [Google Scholar] [CrossRef]
- Chen, M.; Curtis, T.M.; Stitt, A.W. Advanced glycation end products and diabetic retinopathy. Curr. Med. Chem. 2013, 20, 3234–3240. [Google Scholar] [CrossRef] [PubMed]
- Augustine, J.; Troendle, E.P.; Barabas, P.; McAleese, C.A.; Friedel, T.; Stitt, A.W.; Curtis, T.M. The Role of Lipoxidation in the Pathogenesis of Diabetic Retinopathy. Front. Endocrinol. 2021, 11, 1146. [Google Scholar] [CrossRef] [PubMed]
- Fang, I.M.; Yang, C.H.; Yang, C.M.; Chen, M.S. Chitosan oligosaccharides attenuates oxidative-stress related retinal degeneration in rats. PLoS ONE 2013, 8, e77323. [Google Scholar]
- Yeh, P.-T.; Huang, H.-W.; Yang, C.-M.; Yang, W.-S.; Yang, C.-H. Astaxanthin Inhibits Expression of Retinal Oxidative Stress and Inflammatory Mediators in Streptozotocin-Induced Diabetic Rats. PLoS ONE 2016, 11, e0146438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraswathy, S.; Nguyen, A.M.; Rao, N.A. The role of TLR4 in photoreceptor {alpha}a crystallin upregulation during early experimental autoimmune uveitis. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3680–3686. [Google Scholar] [CrossRef]
- Nag, T.C.; Kathpalia, P.; Gorla, S.; Wadhwa, S. Localization of nitro-tyrosine immunoreactivity in human retina. Ann. Anat. 2019, 223, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Noda, K.; Yoshida, S.; Saito, M.; Fujiya, A.; Kanda, A.; Ishida, S. Unsaturated Aldehyde Acrolein Promotes Retinal Glial Cell Migration. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4425–4435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joussen, A.M.; Murata, T.; Tsujikawa, A.; Kirchhof, B.; Bursell, S.E.; Adamis, A.P. Leukocyte-mediated endothelial cell injury and death in the diabetic retina. Am. J. Pathol. 2001, 158, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Joussen, A.M.; Poulaki, V.; Le, M.L.; Koizumi, K.; Esser, C.; Janicki, H.; Schraermeyer, U.; Kociok, N.; Fauser, S.; Kirchhof, B.; et al. A central role for inflammation in the pathogenesis of diabetic retinopathy. FASEB J. 2004, 18, 1450–1452. [Google Scholar] [CrossRef]
- Rübsam, A.; Parikh, S.; Fort, P.E. Role of Inflammation in Diabetic Retinopathy. Int. J. Mol. Sci. 2018, 19, 942. [Google Scholar] [CrossRef] [Green Version]
- Ornitz, D.M.; Itoh, N. The Fibroblast Growth Factor signaling pathway. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 215–266. [Google Scholar] [CrossRef] [Green Version]
- Gospodarowicz, D. Purification of a fibroblast growth factor from bovine pituitary. J. Biol. Chem. 1975, 250, 2515–2520. [Google Scholar] [CrossRef]
- Kan, M.; Huang, J.S.; Mansson, P.E.; Yasumitsu, H.; Carr, B.; McKeehan, W.L. Heparin-binding growth factor type 1 (acidic fibroblast growth factor): A potential biphasic autocrine and paracrine regulator of hepatocyte regeneration. Proc. Natl. Acad. Sci. USA 1989, 86, 7432–7436. [Google Scholar] [CrossRef] [Green Version]
- Banai, S.; Jaklitsch, M.T.; Casscells, W.; Shou, M.; Shrivastav, S.; Correa, R.; Epstein, S.E.; Unger, E.F. Effects of acidic fibroblast growth factor on normal and ischemic myocardium. Circ. Res. 1991, 69, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, M.; Simons, M. Fibroblast growth factor regulation of neovascularization. Curr. Opin. Hematol. 2008, 15, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Min, D.; Guo, G.; Liao, X.; Fu, Z. Experimental study of epidermal growth factor and acidic fibroblast growth factor in the treatment of diabetic foot wounds. Exp. Ther. Med. 2018, 15, 5365–5370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves, L.M. Fibroblast growth factor-mediated angiogenesis for the treatment of ischemia. Lessons learned from experimental models and early human experience. Rev. Port. Cardiol. Orgao Of. Soc. Port. Cardiol. 1998, 17 (Suppl 2), II11–II20. [Google Scholar]
- Palmen, M.; Daemen, M.J.; De Windt, L.J.; Willems, J.; Dassen, W.R.; Heeneman, S.; Zimmermann, R.; Van Bilsen, M.; Doevendans, P.A. Fibroblast growth factor-1 improves cardiac functional recovery and enhances cell survival after ischemia and reperfusion: A fibroblast growth factor receptor, protein kinase C, and tyrosine kinase-dependent mechanism. J. Am. Coll. Cardiol. 2004, 44, 1113–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Liu, Y.; Beenken, A.; Jiang, L.; Gao, X.; Huang, Z.; Hsu, A.; Gross, G.J.; Wang, Y.G.; Mohammadi, M.; et al. A novel fibroblast growth factor-1 ligand with reduced heparin binding protects the heart against ischemia-reperfusion injury in the presence of heparin co-administration. Cardiovasc. Res. 2017, 113, 1585–1602. [Google Scholar] [CrossRef]
- Wu, J.C.; Huang, W.C.; Tsai, Y.A.; Chen, Y.C.; Cheng, H. Nerve repair using acidic fibroblast growth factor in human cervical spinal cord injury: A preliminary Phase I clinical study. J. Neurosurg. Spine 2008, 8, 208–214. [Google Scholar] [CrossRef]
- Suh, J.M.; Jonker, J.W.; Ahmadian, M.; Goetz, R.; Lackey, D.; Osborn, O.; Huang, Z.; Liu, W.; Yoshihara, E.; van Dijk, T.H.; et al. Endocrinization of FGF1 produces a neomorphic and potent insulin sensitizer. Nature 2014, 513, 436–439. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, L.; Chen, S.; Feng, B.; Lu, X.; Bai, Y.; Liang, G.; Tan, Y.; Shao, M.; Skibba, M.; et al. The prevention of diabetic cardiomyopathy by non-mitogenic acidic fibroblast growth factor is probably mediated by the suppression of oxidative stress and damage. PLoS ONE 2013, 8, e82287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Yu, W.Z.; Shen, X.T.; Xiang, Q.; Xu, J.; Yang, J.J.; Chen, P.P.; Fan, Z.L.; Xiao, J.; Zhao, Y.Z.; et al. Advanced Interfere Treatment of Diabetic Cardiomyopathy Rats by aFGF-Loaded Heparin-Modified Microbubbles and UTMD Technique. Cardiovasc. Drugs Ther. 2016, 30, 247–261. [Google Scholar] [CrossRef]
- Liu, W.; Struik, D.; Nies, V.J.; Jurdzinski, A.; Harkema, L.; de Bruin, A.; Verkade, H.J.; Downes, M.; Evans, R.M.; van Zutphen, T.; et al. Effective treatment of steatosis and steatohepatitis by fibroblast growth factor 1 in mouse models of nonalcoholic fatty liver disease. Proc. Natl. Acad. Sci. USA 2016, 113, 2288–2293. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.; Song, L.; Chen, Z.; Qian, Y.; Xie, J.; Zhao, L.; Lin, Q.; Zhu, G.; Tan, Y.; Li, X.; et al. Fibroblast growth factor 1 ameliorates diabetic nephropathy by an anti-inflammatory mechanism. Kidney Int. 2018, 93, 95–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.; Zhang, M.; Dong, B.; Guan, M.; Lu, M.; Huang, Z.; Gao, H.; Li, X. Improved refractory wound healing with administration of acidic fibroblast growth factor in diabetic rats. Diabetes Res. Clin. Pract. 2011, 93, 396–403. [Google Scholar] [CrossRef]
- Noji, S.; Matsuo, T.; Koyama, E.; Yamaai, T.; Nohno, T.; Matsuo, N.; Taniguchi, S. Expression pattern of acidic and basic fibroblast growth factor genes in adult rat eyes. Biochem. Biophys. Res. Commun. 1990, 168, 343–349. [Google Scholar] [CrossRef]
- Jacquemin, E.; Jonet, L.; Oliver, L.; Bugra, K.; Laurent, M.; Courtois, Y.; Jeanny, J.C. Developmental regulation of acidic fibroblast growth factor (aFGF) expression in bovine retina. Int. J. Dev. Biol. 1993, 37, 417–423. [Google Scholar] [PubMed]
- Caruelle, D.; Groux-Muscatelli, B.; Gaudric, A.; Sestier, C.; Coscas, G.; Caruelle, J.P.; Barritault, D. Immunological study of acidic fibroblast growth factor (aFGF) distribution in the eye. J. Cell. Biochem. 1989, 39, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Catalani, E.; Tomassini, S.; Dal Monte, M.; Bosco, L.; Casini, G. Localization patterns of fibroblast growth factor 1 and its receptors FGFR1 and FGFR2 in postnatal mouse retina. Cell Tissue Res. 2009, 336, 423–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Flannery, J.G.; LaVail, M.M.; Steinberg, R.H.; Xu, J.; Simon, M.I. bcl-2 overexpression reduces apoptotic photoreceptor cell death in three different retinal degenerations. Proc. Natl. Acad. Sci. USA 1996, 93, 7042–7047. [Google Scholar] [CrossRef] [Green Version]
- Guillonneau, X.; Régnier-Ricard, F.; Dupuis, C.; Courtois, Y.; Mascarelli, F. Paracrine effects of phosphorylated and excreted FGF1 by retinal pigmented epithelial cells. Growth Factors 1998, 15, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Bryckaert, M.; Guillonneau, X.; Hecquet, C.; Perani, P.; Courtois, Y.; Mascarelli, F. Regulation of proliferation-survival decisions is controlled by FGF1 secretion in retinal pigmented epithelial cells. Oncogene 2000, 19, 4917–4929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullins, R.F.; Skeie, J.M.; Malone, E.A.; Kuehn, M.H. Macular and peripheral distribution of ICAM-1 in the human choriocapillaris and retina. Mol. Vis. 2006, 12, 224–235. [Google Scholar]
- Yoshida, S.; Yoshida, A.; Ishibashi, T.; Elner, S.G.; Elner, V.M. Role of MCP-1 and MIP-1alpha in retinal neovascularization during postischemic inflammation in a mouse model of retinal neovascularization. J. Leukoc. Biol. 2003, 73, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, T.; Hisatomi, T.; Nakazawa, C.; Noda, K.; Maruyama, K.; She, H.; Matsubara, A.; Miyahara, S.; Nakao, S.; Yin, Y.; et al. Monocyte chemoattractant protein 1 mediates retinal detachment-induced photoreceptor apoptosis. Proc. Natl. Acad. Sci. USA 2007, 104, 2425–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natoli, R.; Fernando, N.; Madigan, M.; Chu-Tan, J.A.; Valter, K.; Provis, J.; Rutar, M. Microglia-derived IL-1β promotes chemokine expression by Müller cells and RPE in focal retinal degeneration. Mol. Neurodegener. 2017, 12, 31. [Google Scholar] [CrossRef] [PubMed]
- Wooff, Y.; Man, S.M.; Aggio-Bruce, R.; Natoli, R.; Fernando, N. IL-1 Family Members Mediate Cell Death, Inflammation and Angiogenesis in Retinal Degenerative Diseases. Front. Immunol. 2019, 10, 1618. [Google Scholar] [CrossRef] [PubMed]
- Kitaoka, Y.; Munemasa, Y.; Nakazawa, T.; Ueno, S. NMDA-induced interleukin-1beta expression is mediated by nuclear factor-kappa B p65 in the retina. Brain Res. 2007, 1142, 247–255. [Google Scholar] [CrossRef]
- Ting, D.S.; Cheung, G.C.; Wong, T.Y. Diabetic retinopathy: Global prevalence, major risk factors, screening practices and public health challenges: A review. Clin. Exp. Ophthalmol. 2016, 44, 260–277. [Google Scholar] [CrossRef] [Green Version]
- Kadłubowska, J.; Malaguarnera, L.; Wąż, P.; Zorena, K. Neurodegeneration and Neuroinflammation in Diabetic Retinopathy: Potential Approaches to Delay Neuronal Loss. Curr. Neuropharmacol. 2016, 14, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Kowluru, R.A.; Menon, B.; Gierhart, D.L. Beneficial effect of zeaxanthin on retinal metabolic abnormalities in diabetic rats. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1645–1651. [Google Scholar] [CrossRef]
- Vieira-Potter, V.J.; Karamichos, D.; Lee, D.J. Ocular Complications of Diabetes and Therapeutic Approaches. Biomed. Res. Int. 2016, 2016, 3801570. [Google Scholar] [CrossRef] [Green Version]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxid. Med. Cell. Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [Green Version]
- Simó, R.; Villarroel, M.; Corraliza, L.; Hernández, C.; Garcia-Ramírez, M. The retinal pigment epithelium: Something more than a constituent of the blood-retinal barrier--implications for the pathogenesis of diabetic retinopathy. J. Biomed. Biotechnol. 2010, 2010, 190724. [Google Scholar] [CrossRef]
- Li, C.; Miao, X.; Li, F.; Wang, S.; Liu, Q.; Wang, Y.; Sun, J. Oxidative Stress-Related Mechanisms and Antioxidant Therapy in Diabetic Retinopathy. Oxid. Med. Cell. Longev. 2017, 2017, 9702820. [Google Scholar] [CrossRef] [PubMed]
- Willermain, F.; Scifo, L.; Weber, C.; Caspers, L.; Perret, J.; Delporte, C. Potential Interplay between Hyperosmolarity and Inflammation on Retinal Pigmented Epithelium in Pathogenesis of Diabetic Retinopathy. Int. J. Mol. Sci. 2018, 19, 1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang-Fischer, Y.; Garyantes, T. Improving the Reliability and Utility of Streptozotocin-Induced Rat Diabetic Model. J. Diabetes Res. 2018, 2018, 8054073. [Google Scholar] [CrossRef]
- Yuan, Z.; Feng, W.; Hong, J.; Zheng, Q.; Shuai, J.; Ge, Y. p38MAPK and ERK promote nitric oxide production in cultured human retinal pigmented epithelial cells induced by high concentration glucose. Nitric Oxide 2009, 20, 9–15. [Google Scholar] [CrossRef]
- Khuu, L.-A.; Tayyari, F.; Sivak, J.M.; Flanagan, J.G.; Singer, S.; Brent, M.H.; Huang, D.; Tan, O.; Hudson, C. Aqueous humour concentrations of TGF-β, PLGF and FGF-1 and total retinal blood flow in patients with early non-proliferative diabetic retinopathy. Acta Ophthalmol. 2017, 95, e206–e211. [Google Scholar] [CrossRef] [Green Version]
- Kowluru, R.A.; Chan, P.-S. Oxidative stress and diabetic retinopathy. Exp. Diabetes Res. 2007, 2007, 43603. [Google Scholar] [CrossRef] [Green Version]
- Cecilia, O.-M.; José Alberto, C.-G.; José, N.-P.; Ernesto Germán, C.-M.; Ana Karen, L.-C.; Luis Miguel, R.-P.; Ricardo Raúl, R.-R.; Adolfo Daniel, R.-C. Oxidative Stress as the Main Target in Diabetic Retinopathy Pathophysiology. J. Diabetes Res. 2019, 2019, 8562408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Li, Y.; Jiang, T.; Yuan, Y.; Li, R.; Xu, Z.; Zhong, X.; Jia, G.; Liu, Y.; Xie, L.; et al. Reduction of cellular stress is essential for Fibroblast growth factor 1 treatment for diabetic nephropathy. J. Cell. Mol. Med. 2018, 22, 6294–6303. [Google Scholar] [CrossRef]
- Pena, A.M.; Chen, S.; Feng, B.; Cai, L.; Li, X.; Liang, G.; Chakrabarti, S. Prevention of Diabetic Nephropathy by Modified Acidic Fibroblast Growth Factor. Nephron 2017, 137, 221–236. [Google Scholar] [CrossRef]
- Van Reyk, D.M.; Gillies, M.C.; Davies, M.J. The retina: Oxidative stress and diabetes. Redox Rep. 2003, 8, 187–192. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Jin, M.; Zhao, X.; Zhao, T.; Lin, W.; He, Z.; Fan, M.; Jin, W.; Zhou, J.; Jin, L.; et al. FGF1ΔHBS ameliorates chronic kidney disease via PI3K/AKT mediated suppression of oxidative stress and inflammation. Cell Death Dis. 2019, 10, 464. [Google Scholar] [CrossRef]
- Wu, Y.; Jia, G.; Wang, B.; Xiong, J.; Xu, J.; Zheng, P.; Yuan, Y.; Li, Y.; Jiang, T.; Al Mamun, A.; et al. Fibroblast growth factor 1 ameliorates diabetes-induced splenomegaly via suppressing inflammation and oxidative stress. Biochem. Biophys. Res. Commun. 2020, 528, 249–255. [Google Scholar] [CrossRef]
- Meleth, A.D.; Agrón, E.; Chan, C.C.; Reed, G.F.; Arora, K.; Byrnes, G.; Csaky, K.G.; Ferris, F.L., III; Chew, E.Y. Serum inflammatory markers in diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4295–4301. [Google Scholar] [CrossRef]
- Koleva-Georgieva, D.N.; Sivkova, N.P.; Terzieva, D. Serum inflammatory cytokines IL-1beta, IL-6, TNF-alpha and VEGF have influence on the development of diabetic retinopathy. Folia Med. 2011, 53, 44–50. [Google Scholar]
- Feng, S.; Yu, H.; Yu, Y.; Geng, Y.; Li, D.; Yang, C.; Lv, Q.; Lu, L.; Liu, T.; Li, G.; et al. Levels of Inflammatory Cytokines IL-1β, IL-6, IL-8, IL-17A, and TNF-α in Aqueous Humour of Patients with Diabetic Retinopathy. J. Diabetes Res. 2018, 2018, 8546423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raju, R.; Palapetta, S.M.; Sandhya, V.K.; Sahu, A.; Alipoor, A.; Balakrishnan, L.; Advani, J.; George, B.; Kini, K.R.; Geetha, N.P.; et al. A Network Map of FGF-1/FGFR Signaling System. J. Signal Transduct. 2014, 2014, 962962. [Google Scholar] [CrossRef] [Green Version]
- Guillonneau, X.; Bryckaert, M.; Launay-Longo, C.; Courtois, Y.; Mascarelli, F. Endogenous FGF1-induced activation and synthesis of extracellular signal-regulated kinase 2 reduce cell apoptosis in retinal-pigmented epithelial cells. J. Biol. Chem. 1998, 273, 22367–22373. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Du, S.; Wu, Q.; Hu, J.; Li, T. Decorin Prevents Retinal Pigment Epithelial Barrier Breakdown under Diabetic Conditions by Suppressing p38 MAPK Activation. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2971–2979. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, M.; Wakasaki, H.; Takahara, N.; Ishii, H.; Jiang, Z.Y.; Yamauchi, T.; Kuboki, K.; Meier, M.; Rhodes, C.J.; King, G.L. Glucose or diabetes activates p38 mitogen-activated protein kinase via different pathways. J. Clin. Investig. 1999, 103, 185–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.G.; Kim, K.S.; Park, H.C.; Choi, K.H.; Lee, H.Y.; Han, D.S.; Kang, S.W. High glucose activates the p38 MAPK pathway in cultured human peritoneal mesothelial cells. Kidney Int. 2003, 63, 958–968. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Yu, H.; Huang, S.; Zhu, P. Resveratrol Protects against TNF-α-Induced Injury in Human Umbilical Endothelial Cells through Promoting Sirtuin-1-Induced Repression of NF-KB and p38 MAPK. PLoS ONE 2016, 11, e0147034. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, J.; Wang, L.; Chen, C.; Yang, D.; Jin, M.; Bai, C.; Song, Y. Urban particulate matter triggers lung inflammation via the ROSMAPK- NF-κB signaling pathway. J. Thorac. Dis. 2017, 9, 4398–4412. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.; Santani, D. Role of NF-κB in the pathogenesis of diabetes and its associated complications. Pharmacol. Rep. 2009, 61, 595–603. [Google Scholar] [CrossRef]
- Olson, C.M.; Hedrick, M.N.; Izadi, H.; Bates, T.C.; Olivera, E.R.; Anguita, J. p38 Mitogen-Activated Protein Kinase Controls NF-κB Transcriptional Activation and Tumor Necrosis Factor Alpha Production through RelA Phosphorylation Mediated by Mitogen- and Stress-Activated Protein Kinase 1 in Response to Borrelia burgdorferi Antigens. Infect. Immun. 2007, 75, 270–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, R.N.; Jana, M.; Pahan, K. MAPK p38 regulates transcriptional activity of NF-kappaB in primary human astrocytes via acetylation of p65. J. Immunol. 2007, 179, 7101–7109. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Liu, X.; Qu, Y.; Wang, L.; Geng, D.; Chen, W.; Li, L.; Tian, Y.; Chang, S.; Zhao, C.; et al. The roles of p38 MAPK → COX2 and NF-κB → COX2 signal pathways in age-related testosterone reduction. Sci. Rep. 2019, 9, 10556. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.M.; Xu, W.M.; Lin, J.C.; Mo, L.Q.; Hua, X.X.; Chen, P.X.; Wu, K.; Zheng, D.D.; Feng, J.Q. Activation of the p38 MAPK/NF-kappaB pathway contributes to doxorubicin-induced inflammation and cytotoxicity in H9c2 cardiac cells. Mol. Med. Rep. 2013, 8, 603–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Company | Catalogue Number | Concentration |
|---|---|---|---|
| Primary Antibody | |||
| anti-FGF1 | Abcam | ab179455 | 1:200 |
| anti-8OHdG | Abcam | ab62623 | 1:200 |
| anti-nitrotyrosine | Abcam | ab42789 | 1:200 |
| anti-acrolein | Abcam | ab37110 | 1:200 |
| anti-ICAM-1 | Abcam | ab282575 | 1:200 |
| anti-MCP-1 | Abcam | ab7202 | 1:100 |
| anti-IL-1β | Cell Signaling | 12242S | 1:200 |
| Secondary Antibody | |||
| DyLight 594 Antibody Kit | Vector | DI-1794 | 1:200 |
| DyLight 488 Antibody Kit | Vector | DI-1788 | 1:200 |
| Gene | Sequence (5′→3′) |
|---|---|
| ICAM-1 | forward 5′-TCAGAAGGGACCGAGGTGAT-3′ |
| reverse 5′-TTTTCTGGCCACGTCCAGT-3′ | |
| MCP-1 | forward 5′-CCAGATGCAATCAATGCCCC-3′ |
| reverse 5′-TCCTTGGCCACAATGGTCTT-3′ | |
| IL-1β | forward 5′-ACCTGAGCTCGCCAGTGAAA-3′ |
| reverse 5′-CAACAACTGACACGGCCTGC-3′ | |
| IL-6 | forward 5′-AAGCCAGAGCTGTGCAAATGAG-3′ |
| reverse 5′-TCGTCAGCAGGCTGGCATTT-3′ | |
| GAPDH | forward 5′- TTCGAGAG-TCAGCCGCATTT-3′ |
| reverse 5′- GACTCCGACCTTCACCTTCC-3′ |
| Name | Company | Catalogue Number | Concentration |
|---|---|---|---|
| Primary Antibody | |||
| anti-FGF1 | Abcam | ab179455 | 1:1000 |
| anti-ICAM-1 | proteintech | 10831-1-AP | 1:500 |
| anti-MCP-1 | BioLegend | 502601 | 1:1000 |
| anti-IL-1β | Cell Signaling | 12242S | 1:1000 |
| anti-IL-6 | Abcam | ab6672 | 1:1000 |
| anti-p38 | Cell Signaling | #9212s | 1:1000 |
| anti-phospho-p38 | Cell Signaling | #9211 | 1:1000 |
| anti-GAPDH | Millipore | MAB374 | 1:2000 |
| anti-β-actin | Abcam | ab8226 | 1:2000 |
| Secondary Antibody | |||
| anti-mouse IgG | Cell Signaling | 7076S | 1:5000 |
| anti-rabbit IgG | Cell Signaling | 7074S | 1:5000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, H.-W.; Yang, C.-M.; Yang, C.-H. Fibroblast Growth Factor Type 1 Ameliorates High-Glucose-Induced Oxidative Stress and Neuroinflammation in Retinal Pigment Epithelial Cells and a Streptozotocin-Induced Diabetic Rat Model. Int. J. Mol. Sci. 2021, 22, 7233. https://doi.org/10.3390/ijms22137233
Huang H-W, Yang C-M, Yang C-H. Fibroblast Growth Factor Type 1 Ameliorates High-Glucose-Induced Oxidative Stress and Neuroinflammation in Retinal Pigment Epithelial Cells and a Streptozotocin-Induced Diabetic Rat Model. International Journal of Molecular Sciences. 2021; 22(13):7233. https://doi.org/10.3390/ijms22137233
Chicago/Turabian StyleHuang, Hsin-Wei, Chung-May Yang, and Chang-Hao Yang. 2021. "Fibroblast Growth Factor Type 1 Ameliorates High-Glucose-Induced Oxidative Stress and Neuroinflammation in Retinal Pigment Epithelial Cells and a Streptozotocin-Induced Diabetic Rat Model" International Journal of Molecular Sciences 22, no. 13: 7233. https://doi.org/10.3390/ijms22137233