
Altered Induction of Reactive Oxygen Species by X-rays in Hematopoietic Cells of C57BL/6-Tg (CAG-EGFP) Mice

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. General Physiological Conditions of the Mice
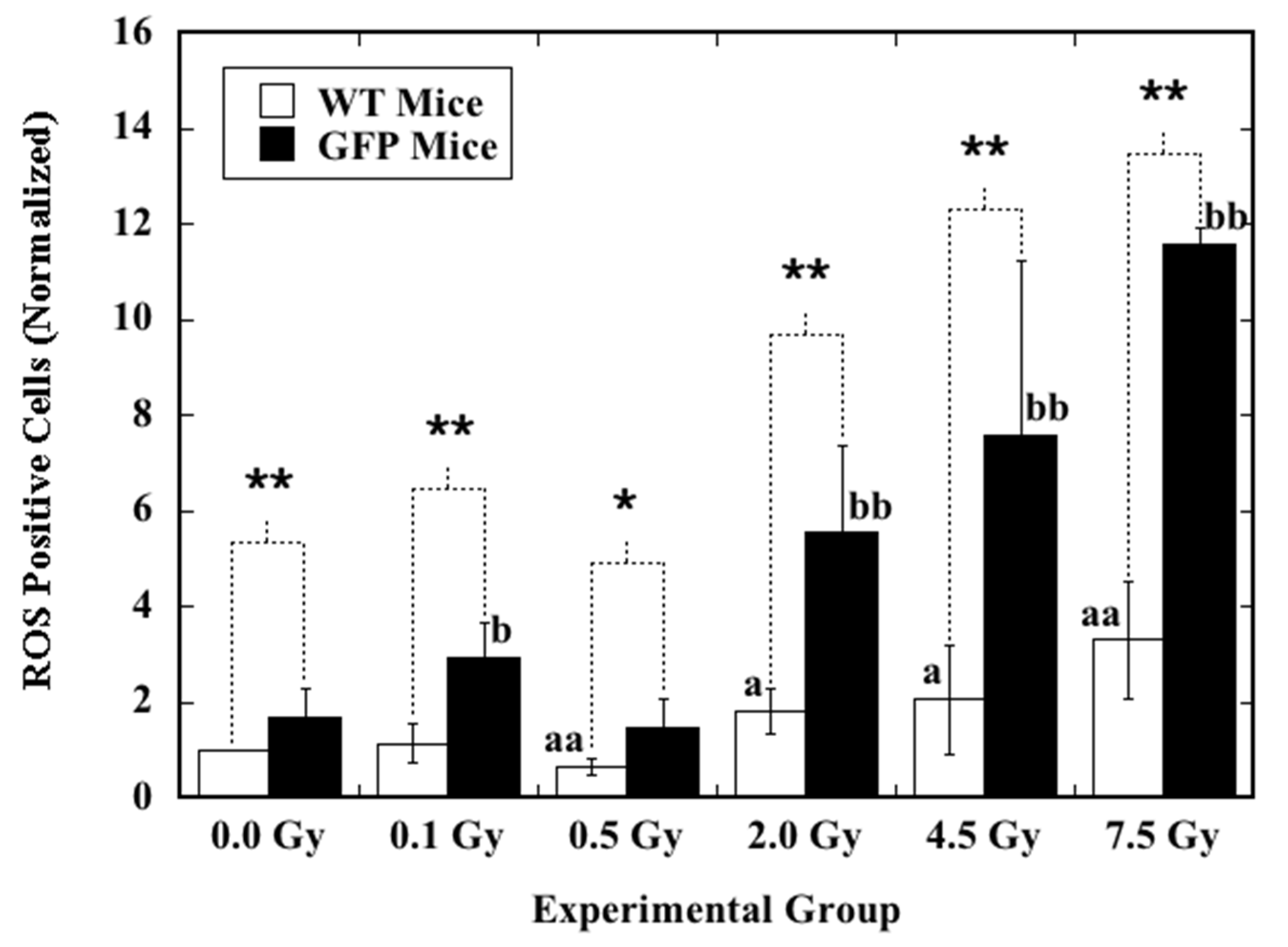
2.2. Induction of ROS by X-rays in the Hematopoietic Cells
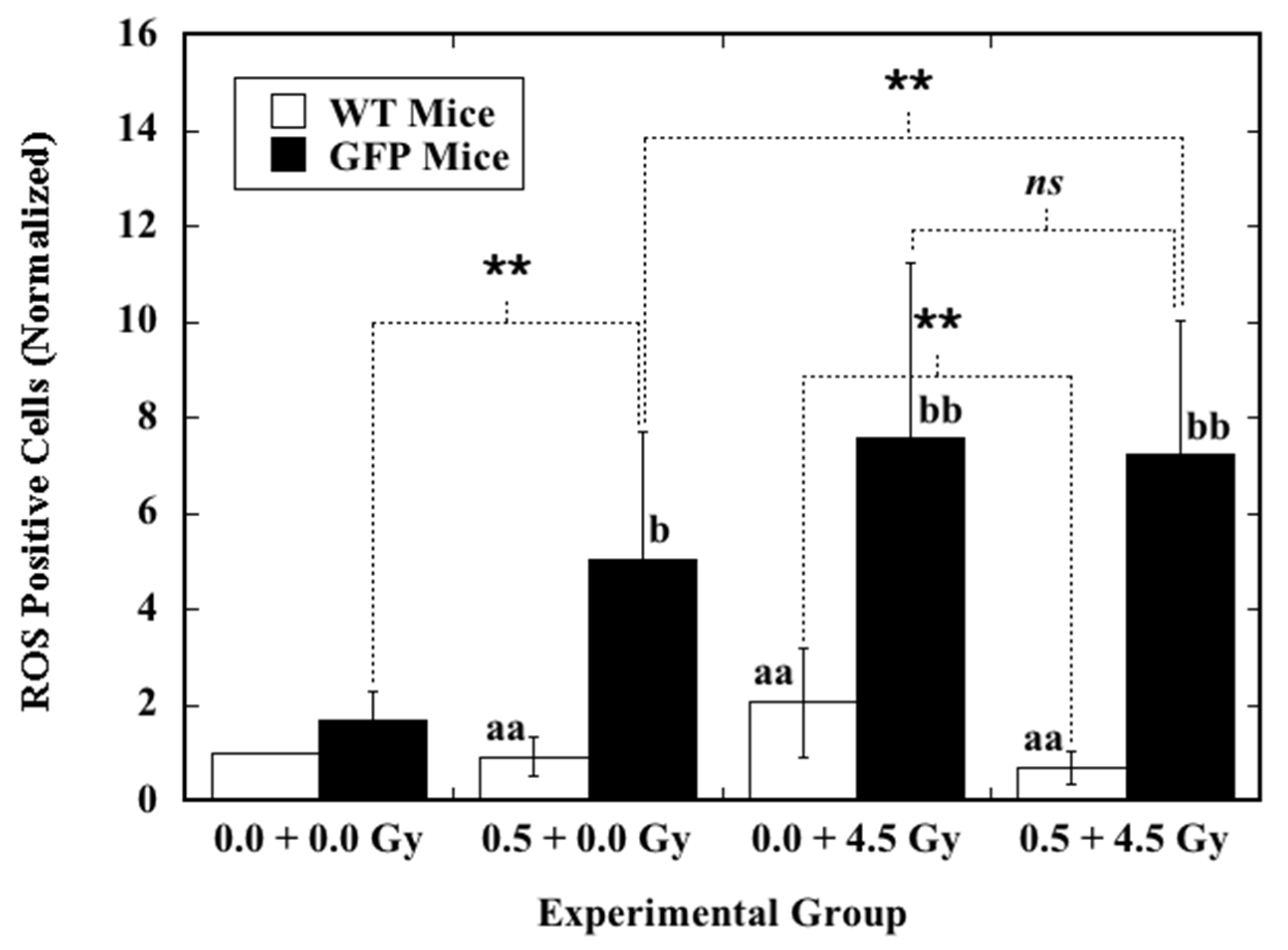
2.3. Effect of Priming TBI on Induction of ROS by Ex Vivo X-Irradiation in the Hematopoietic Cells
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Collection of the Nucleated Bone Marrow Cells
4.3. Irradiation
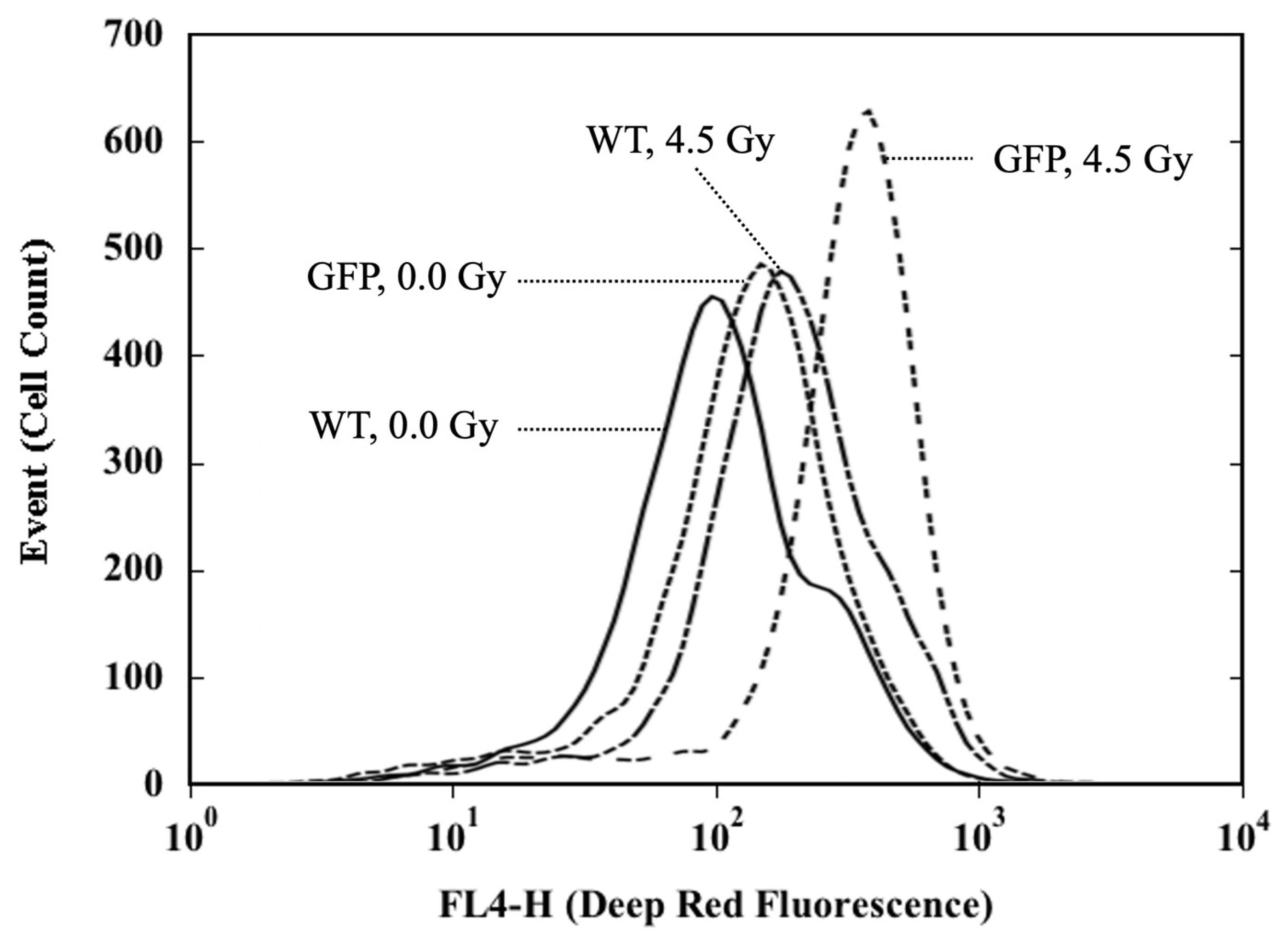
4.4. Flow Cytometric Analysis of ROS
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Godwin, A.R.; Stadler, H.S.; Nakamura, K.; Capecchi, M.R. Detection of targeted GFP-Hox gene fusions during mouse embryogenesis. Proc. Natl. Acad. Sci. USA 1998, 95, 13042–13047. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Campbell, R.E.; Ting, A.Y.; Tsien, R.Y. Creating new fluorescent probes for cell biology. Nat. Rev. Mol. Cell Biol. 2002, 3, 906–918. [Google Scholar] [CrossRef]
- Shaner, N.C.; Steinbach, P.A.; Tsien, R.Y. A guide to choosing fluorescent proteins. Nat. Methods 2005, 2, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Gurtu, V.; Kain, S.R. An enhanced green fluorescent protein allows sensitive detection of gene transfer in mammalian cells. Biochem. Biophys. Res. Commun. 1996, 227, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.A.; Szalay, A.A.; Wang, G.; Oberg, K. Visualization of molecular and cellular events with green fluorescent proteins in developing embryos: A review. Luminescence 2003, 18, 1–18. [Google Scholar] [CrossRef]
- Baumstark-Khan, C.; Hellweg, C.E.; Palm, M.; Horneck, G. Enhanced green fluorescent protein (EGFP) for space radiation research using mammalian cells in the International Space Station. Phys. Med. 2001, 17, 210–214. [Google Scholar] [PubMed]
- Baumstark-Khan, C.; Hellweg, C.E.; Arenz, A.; Meier, M.M. Cellular monitoring of the nuclear factor kappaB pathway for assessment of space environmental radiation. Radiat. Res. 2005, 164, 527–530. [Google Scholar] [CrossRef]
- Tanno, Y.; Kobayashi, K.; Tatsuka, M.; Gotoh, E.; Takakura, K. Mitotic arrest caused by an X-ray microbeam in a single cell expressing EGFP-aurora kinase B. Radiat. Protect. Dosi. 2006, 122, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Lee, C.; Hayashi, K.; Yamauchi, K.; Yamamoto, N.; Tsuchiya, H.; Tomita, K.; Bouvet, M.; Hoffman, R.M. UV light killing efficacy of fluorescent protein-expressing cancer cells In Vitro and In Vivo. J. Cell. Biochem. 2010, 110, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, K.; Yori, K.; Kaminaga, C.; Yamashita, T.; Kinoshita, M.; Oda, S.; Mitani, H. Responses of embryonic germ cells of the radiation-sensitive Medaka mutant to gamma-irradiation. J. Radiat. Res. 2007, 48, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, K.; Iwanami, N.; Maruyama-Hayakawa, T.; Doi, K.; Wang, B. A small fish model for quantitative analysis of radiation effects using visualized thymus responses in GFP transgenic medaka. Int. J. Radiat. Biol. 2019, 95, 1144–1149. [Google Scholar] [CrossRef]
- Anaganti, N.; Basu, B.; Apte, S.K. In situ real-time evaluation of radiation-responsive promoters in the extremely radioresistant microbe Deinococcus radiodurans. J. Biosci. 2016, 41, 193–203. [Google Scholar] [CrossRef]
- Petrova, A.S.; Lukonina, A.A.; Badun, G.A.; Kudryasheva, N.S. Fluorescent coelenteramide-containing protein as a color bioindicator for low-dose radiation effect. Anal. Bioanal. Chem. 2017, 409, 4377–4381. [Google Scholar] [CrossRef]
- Sarkar, P.; Chattopadhyay, A. GFP fluorescence: A few lesser known nuggets that make it work. J. Biosci. 2018, 43, 421–430. [Google Scholar] [CrossRef]
- Heim, R.; Prasher, D.C.; Tsien, R.Y. Wavelength mutations and posttranslational autoxidation of green fluorescence protein. Proc. Natl. Acad. Sci. USA 1994, 91, 12501–12504. [Google Scholar] [CrossRef] [Green Version]
- Okabe, M.; Ikawa, M.; Kominami, K.; Nakanishi, T.; Nishimune, Y. ‘Green mice’ as a source of ubiquitous green cells. FEBS Lett. 1997, 407, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.S.O.D.E.; Pinho, T.M.G.; Rachid, M.A.; Barbosa-Stancioli, D.F.; Da Fonseca, F.G. The perennial use of the green fluorescent protein marker in a live vaccinia virus Ankara recombinant platform shows no acute adverse effects in mice. Braz. J. Microbiol. 2019, 50, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Ansari, A.M.; Ahmed, A.K.; Matsangos, A.E.; Lay, F.; Born, L.J.; Marti, G.; Harmon, J.W.; Sun, Z. Cellular GFP toxicity and immunogenicity: Potential confounders in In Vivo cell tracking experiments. Stem Cell Rev. Rep. 2016, 12, 553–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.S.; Jan, M.S.; Chou, C.K.; Chen, P.H.; Ke, N.J. Is green fluorescent protein toxic to the living cells? Biochem. Biophys. Res. Commun. 1999, 260, 712–717. [Google Scholar] [CrossRef] [Green Version]
- Kam, W.W.; Middleton, R.; Lake, V.; Banati, R.B. Green fluorescent protein alters the transcriptional regulation of human mitochondrial genes after gamma irradiation. J. Fluoresc. 2013, 23, 613–619. [Google Scholar] [CrossRef]
- Makanae, K.; Kintaka, R.; Makino, T.; Kitano, H.; Moriya, H. Identification of dosage-sensitive genes in Saccharomyces cerevisiae using the genetic tug-of-war method. Genome Res. 2013, 23, 300–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, L.M.; Moreo, A.; Clark, K.R.; Harper, S.Q. Dose-dependent toxicity of humanized renilla reniformis GFP (hrGFP) limits its utility as a reporter gene in mouse muscle. Mol. Ther. Nucleic Acids. 2013, 2, e86. [Google Scholar] [CrossRef]
- Coumans, J.V.F.; Gau, C.D.; Poljak, A.; Wasinger, V.; Roy, P.; Moens, P. Green fluorescent protein expression triggers proteome changes in breast cancer cells. Exp. Cell Res. 2014, 320, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Avey, S.R.; Ojehomon, M.; Dawson, J.F.; Gillis, T.E. How the expression of green fluorescent protein and human cardiac actin in the heart influences cardiac function and aerobic performance in zebrafish Danio rerio. J. Fish Biol. 2017, 92, 177–189. [Google Scholar] [CrossRef]
- Huang, W.Y.; Aramburu, J.; Douglas, P.S.; Izumo, S. Transgenic expression of green fluorescence protein can cause dilated cardiomyopathy. Nat. Med. 2000, 6, 482–483. [Google Scholar] [CrossRef]
- Li, H.; Wei, H.; Wang, Y.; Tang, H.; Wang, Y. Enhanced green fluorescent protein transgenic expression In Vivo is not biologically inert. J. Proteome Res. 2013, 12, 3801–3808. [Google Scholar] [CrossRef] [PubMed]
- Goto, H.; Yang, B.; Petersen, D.; Pepper, K.A.; Alfaro, P.A.; Kohn, D.B.; Reynolds, C.P. Transduction of green fluorescent protein increased oxidative stress and enhanced sensitivity to cytotoxic drugs in neuroblastoma cell lines. Mol. Cancer Ther. 2003, 2, 911–917. [Google Scholar] [PubMed]
- Tsien, R.Y. The green fluorescent protein. Annu. Rev. Biochem. 1998, 67, 509–544. [Google Scholar] [CrossRef]
- Liu, C.; Tanaka, K.; Katsube, T.; Varès, G.; Maruyama, K.; Ninomiya, Y.; Fardous, Z.; Sun, C.; Fujimori, A.; Moreno, S.G.; et al. Altered response to total body irradiation of C57BL/6-Tg (CAG-EGFP) mice. Dose Res. 2020, 18, 1559325820951332. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Wang, B.; Wang, H.; Meng, L.; Zhao, Q.; Li, X.; Xin, Y.; Jiang, X. Radiation-induced normal tissue damage: Oxidative stress and epigenetic mechanisms. Oxid. Med. Cell. Longev. 2019, 2019, 3010342. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Lee, S.; Seo, D.; Kim, D.; Kim, K.; Kim, E.; Kang, J.; Seong, K.M.; Youn, H.; Youn, B. Cellular stress responses in radiotherapy. Cells 2019, 8, 1105. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Lyu, X.; Yuan, S.; Wang, S.; Li, W.; Chen, Z.; Yu, H.; Li, F.; Jiang, Q. Oxidative stress: A critical hint in ionizing radiation induced pyroptosis. Radiat. Med. Prot. 2020, 1, 179–185. [Google Scholar] [CrossRef]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Jia, Z.; Trush, M.A. Defining ROS in biology and medicine. React. Oxyg. Species 2016, 1, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, F. Chemical basis of reactive oxygen species reactivity and involvement in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Auten, R.; Davis, J. Oxygen toxicity and reactive oxygen species: The devil is in the details. Pediatr. Res. 2009, 66, 121–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobashigawa, S.; Kashino, G.; Suzuki, K.; Yamashita, S.; Mori, H. Ionizing radiation-induced cell death is partly caused by increase of mitochondrial reactive oxygen species in normal human fibroblast cells. Radiat. Res. 2015, 183, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Albano, C.R.; Bentley, W.E.; Rao, G. Quantitative and kinetic study of oxidative stress regulons using green fluorescent protein. Biotechnol. Bioeng. 2005, 89, 574–587. [Google Scholar] [CrossRef]
- Zhang, L.; Patel, H.N.; Lappe, J.W.; Wachter, R.M. Reaction progress of chromophore biogenesis in green fluorescent protein. J. Am. Chem. Soc. 2006, 128, 4766–4772. [Google Scholar] [CrossRef]
- Ganini, D.; Leinisch, F.; Kumar, A.; Jiang, J.; Tokar, E.; Malone, C.C.; Petrovich, R.M.; Mason, R.P. Fluorescent proteins such as eGFP lead to catalytic oxidative stress in cells. Redox Biol. 2017, 12, 462–468. [Google Scholar] [CrossRef]
- Sobotta, M.C.; Liou, W.; Stöcker, S.; Talwar, D.; Oehler, M.; Ruppert, T.; Scharf, A.N.D.; Dick, T.P. Peroxiredoxin-2 and STAT3 form a redox relay for H2O2 signaling. Nat. Chem. Biol. 2015, 11, 64–70. [Google Scholar] [CrossRef]
- Murakami, S.; Motohashi, H. Roles of Nrf2 in cell proliferation and differentiation. Free. Radic. Biol. Med. 2015, 88, 168–178. [Google Scholar] [CrossRef] [Green Version]
- Mikhed, Y.; Görlach, A.; Knaus, U.G.; Daiber, A. Redox regulation of genome stability by effects on gene expression, epigenetic pathways and DNA damage/repair. Redox Biol. 2015, 5, 275–289. [Google Scholar] [CrossRef] [Green Version]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Kalyanaraman, B. Green fluorescent proteins induce oxidative stress in cells: A worrisome new wrinkle in the application of the GFP reporter system to biological systems? Redox Biol. 2017, 12, 755–757. [Google Scholar] [CrossRef]
- Nenoi, M.; Wang, B.; Varès, G. In Vivo radioadaptive response: A review of studies relevant to radiation-induced cancer risk. Human Exp. Toxicol. 2015, 34, 272–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapio, S.; Jacob, V. Radioadaptive response revisited. Radiat. Environ. Biophys. 2007, 46, 1–12. [Google Scholar] [CrossRef]
- Belli, M.; Indovina, L. The response of living organisms to low radiation environment and its implications in radiation protection. Front. Public Health 2020, 8, 601711. [Google Scholar] [CrossRef] [PubMed]
- Coleman, C.N.; Eke, I.; Makinde, A.Y.; Chopra, S.; Demaria, S.; Formenti, S.C.; Martello, S.; Bylicky, M.; Mitchell, J.B.; Aryankalayi, M.J. Radiation-induced adaptive response: New potential for cancer treatment. Clin. Cancer Res. 2020, 26, 5781–5790. [Google Scholar] [CrossRef] [PubMed]
- Dimova, E.G.; Bryant, P.E.; Chankova, S.G. “Adaptive response”—Some underlying mechanisms and open questions. Genet. Mol. Biol. 2008, 31, 396–408. [Google Scholar] [CrossRef] [Green Version]
- Sisakht, M.; Darabian, M.; Mahmoodzadeh, A.; Bazi, A.; Shafiee, S.M.; Mokarram, P.; Khoshdel, Z. The role of radiation induced oxidative stress as a regulator of radio-adaptive responses. Int. J. Radiat. Biol. 2020, 96, 561–576. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef]
- Verbon, E.H.; Post, J.A.; Boonstra, J. The influence of reactive oxygen species on cell cycle progression in mammalian cells. Gene 2012, 511, 1–6. [Google Scholar] [CrossRef]
- Zhang, M.; Du, Y.; Lu, R.; Shu, Y.; Zhao, W.; Li, Z.; Zhang, Y.; Liu, R.; Yang, T.; Luo, S.; et al. Cholesterol retards senescence in bone marrow mesenchymal stem cells by modulating autophagy and ROS/p53/p21Cip1/Waf1 pathway. Oxid. Med. Cell. Longev. 2016, 2016, 7524308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, M.; Pickrell, A.M.; Wang, X.; Bacman, S.R.; Yu, A.; Hida, A.; Dillon, L.M.; Morton, P.D.; Malek, T.R.; Williams, S.L.; et al. Transient mitochondrial DNA double strand breaks in mice cause accelerated aging phenotypes in a ROS-dependent but p53/p21-independent manner. Cell Death Differ. 2017, 24, 288–299. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Huang, C.; Sun, A.; Qiao, L.; Zhang, X.; Huang, J.; Sun, X.; Yang, X.; Sun, S. Hydrogen alleviates cellular senescence via regulation of ROS/p53/p21 pathway in bone marrow-derived mesenchymal stem cells In Vivo. Biomed. Pharmacother. 2018, 106, 1126–1134. [Google Scholar] [CrossRef]
- Patterson, J.C.; Joughin, B.A.; van de Kooij, B.; Lim, D.C.; Lauffenburger, D.A.; Yaffe, M.B. ROS and oxidative stress are elevated in mitosis during asynchronous cell cycle progression and are exacerbated by mitotic arrest. Cell Syst. 2019, 8, 163–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plautz, J.; Day, R.N.; Dailey, G.M.; Welsh, S.B.; Hall, J.C.; Halpain, S.; Kay, S.A. Green fluorescent protein and its derivatives as versatile markers for gene expression in living Drosophila melanogaster, plant and mammalian cells. Gene 1996, 173, 83–87. [Google Scholar] [CrossRef]
- Gubin, A.; Reddy, B.; Njoroge, J.M.; Miller, J.L. Long-term, stable expression of GFP in mammalian cells. Biochem. Biophys. Res. Comm. 1997, 236, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Roy, N.S.; Wang, S.; Jiang, L.; Kang, J.; Benraiss, A.; Harrison-Restelli, C.; Fraser, R.A.R.; Couldwell, W.T.; Kawaguchi, A.; Okano, H.; et al. In Vitro neurogenesis by progenitor cells isolated from the adult human hippocampus. Nat. Med. 2000, 6, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Goldman, S.; Roy, N. Reply to “Human neural progenitor cells: Better blue than green?”. Nat. Med. 2000, 6, 483–484. [Google Scholar] [CrossRef] [PubMed]
- Hadjantonakis, A.K.; Gertsenstein, M.; Ikawa, M.; Okabe, M.; Nagy, A. Generating green fluorescent mice by germline transmission of green fluorescent ES cells. Mech. Dev. 1998, 76, 79–90. [Google Scholar] [CrossRef]
- Hadjantonakis, A.K.; Macmaster, S.; Nagy, A. Embryonic stem cells and mice expressing different GFP variants for multiple non-invasive reporter usage within a single animal. BMC Biotechnol. 2002, 2, 11. [Google Scholar] [CrossRef]
- Khoo, M.S.C.; Grueter, C.E.; Eren, M.; Yang, J.; Zhang, R.; Bass, M.A.; Lwin, S.T.; Mendes, L.A.; Vaughan, D.E.; Colbran, R.J.; et al. Calmodulin kinase II inhibition disrupts cardiomyopathic effects of enhanced green fluorescent protein. J. Mol. Cell. Cardiol. 2008, 44, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, M.; Connole, M.; Glickman, R.; Yue, S.P.; Noren, B.; DeMaria, M.; Johnson, R.P. Induction of cytotoxic T lymphocyte and antibody responses to enhanced green fluorescent protein following transplantation of transduced CD34(+) hematopoietic cells. Blood 2001, 97, 1951–1959. [Google Scholar] [CrossRef] [Green Version]
- Riddell, S.R.; Elliott, M.; Lewinsohn, D.A.; Gilbert, M.J.; Wilson, L.; Manley, S.A.; Lupton, S.D.; Overell, R.W.; Reynolds, T.C.; Corey, L.; et al. T cell-mediated rejection of gene-modified HIV specific cytotoxic T lymphocytes in HIV-infected patients. Nat. Med. 1996, 2, 216–223. [Google Scholar] [CrossRef]
- Re, F.; Srinivasan, R.; Igarashi, T.; Marincola, F.; Childs, R. Green fluorescent protein expression in dendritic cells enhances their immunogenicity and elicits specific cytotoxic T-cell responses in humans. Exp. Hematol. 2004, 32, 210–217. [Google Scholar] [CrossRef]
- Gambotto, A.; Dworacki, G.; Cicinnati, V.; Kenniston, T.; Steitz, J.; Tüting, T.; Robbins, P.D.; DeLeo, A.B. Immunogenicity of enhanced green fluorescent protein (EGFP) in BALB/c mice: Identification of an H2-Kd-restricted CTL epitope. Gene Ther. 2000, 7, 2036–2040. [Google Scholar] [CrossRef] [Green Version]
- Han, W.G.H.; Unger, W.W.J.; Wauben, M.H.M. Identification of the immunodominant CTL epitope of EGFP in C57BL/6 mice. Gene Ther. 2008, 15, 700–701. [Google Scholar] [CrossRef] [Green Version]
- Bresser, K.; Dijkgraaf, F.E.; Pritchard, C.E.J.; Huijbers, I.J.; Song, J.Y.; Rohr, J.C.; Scheeren, F.A.; Schumacher, T.N. A mouse model that is immunologically tolerant to reporter and modifier proteins. Commun. Biol. 2020, 3, 273. [Google Scholar] [CrossRef] [PubMed]
- Stripecke, R.; del Carmen Villacres, M.; Skelton, D.C.; Satake, N.; Halene, S.; Kohn, D.B. Immune response to green fluorescent protein: Implications for gene therapy. Gene Ther. 1999, 6, 1305–1312. [Google Scholar] [CrossRef]
- Aoyama, N.; Miyoshi, H.; Miyachi, H.; Sonoshita, M.; Okabe, M.; Taketo, M.M. Transgenic mice that accept Luciferase- or GFP-expressing syngeneic tumor cells at high efficiencies. Genes Cells 2018, 23, 580–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipták, N.; Bősze, Z.; Hiripi, L. GFP transgenic animals in biomedical research: A review of potential disadvantages. Physiol. Res. 2019, 68, 525–530. [Google Scholar] [CrossRef]
- Dalmazi, G.D.; Hirshberg, J.; Lyle, D.; Freij, J.B.; Caturegli, P. Reactive oxygen species in organ-specific autoimmunity. Autoimmun. Highlights 2016, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.H.; Griffiths, H.R. The dual role of reactive oxygen species in autoimmune and inflammatory diseases: Evidence from preclinical models. Free Radic. Biol. Med. 2018, 125, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Awad, E.M.; Khan, S.Y.; Sokolikova, B.; Brunner, P.M.; Olcaydu, D.; Wojta, J.; Breuss, J.M.; Uhrin, P. Cold induces reactive oxygen species production and activation of the NF-kappa B response in endothelial cells and inflammation In Vivo. J. Thromb. Haemost. 2013, 11, 1716–1726. [Google Scholar] [CrossRef]
- Sun, W.; Wang, Z.; Cao, J.; Cui, H.; Ma, Z. Cold stress increases reactive oxygen species formation via TRPA1 activation in A549 cells. Cell Stress Chaperones 2016, 21, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, T.; Kuroiwa, A.; Yamada, S.; Isotani, A.; Yamashita, A.; Tairaka, A.; Hayashi, T.; Takagi, T.; Ikawa, M.; Matsuda, Y.; et al. FISH analysis of 142 EGFP transgene integration sites into the mouse genome. Genomics 2002, 80, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Ikawa, M.; Kominami, K.; Yoshimura, Y.; Tanaka, K.; Nishimune, Y.; Okabe, M. Green fluorescent protein as a marker in transgenic mice. Dev. Growth Differ. 1995, 37, 455–459. [Google Scholar] [CrossRef]
- Xue, Y.; Wang, A.-Z. DJ-1 plays a neuroprotective role in SH-SY5Y cells by modulating Nrf2 signaling in response to lidocaine-mediated oxidative stress and apoptosis. Kaohsiung J. Med. Sci. 2020, 36, 630–639. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Mice | Body Weight (g) at 6 Postnatal Weeks | Body Weight (g) at 8 Postnatal Weeks | Body Weight (g) at 8 Postnatal Weeks (with 0.5 Gy TBI at 6 Postnatal Weeks) |
|---|---|---|---|
| WT | 16.7 ± 0.6 | 18.4 ± 0.5 | 18.7 ± 1.0 |
| GFP | 16.5 ± 0.7 | 17.8 ± 0.5 * | 16.4 ± 0.6 *,# |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Hirakawa, H.; Katsube, T.; Fang, Y.; Tanaka, K.; Nenoi, M.; Fujimori, A.; Wang, B. Altered Induction of Reactive Oxygen Species by X-rays in Hematopoietic Cells of C57BL/6-Tg (CAG-EGFP) Mice. Int. J. Mol. Sci. 2021, 22, 6929. https://doi.org/10.3390/ijms22136929
Liu C, Hirakawa H, Katsube T, Fang Y, Tanaka K, Nenoi M, Fujimori A, Wang B. Altered Induction of Reactive Oxygen Species by X-rays in Hematopoietic Cells of C57BL/6-Tg (CAG-EGFP) Mice. International Journal of Molecular Sciences. 2021; 22(13):6929. https://doi.org/10.3390/ijms22136929
Chicago/Turabian StyleLiu, Cuihua, Hirokazu Hirakawa, Takanori Katsube, Yaqun Fang, Kaoru Tanaka, Mitsuru Nenoi, Akira Fujimori, and Bing Wang. 2021. "Altered Induction of Reactive Oxygen Species by X-rays in Hematopoietic Cells of C57BL/6-Tg (CAG-EGFP) Mice" International Journal of Molecular Sciences 22, no. 13: 6929. https://doi.org/10.3390/ijms22136929