
Efficacy of a Three Drug-Based Therapy for Neuroblastoma in Mice

 ,
, 
 , , , and
, , , and 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
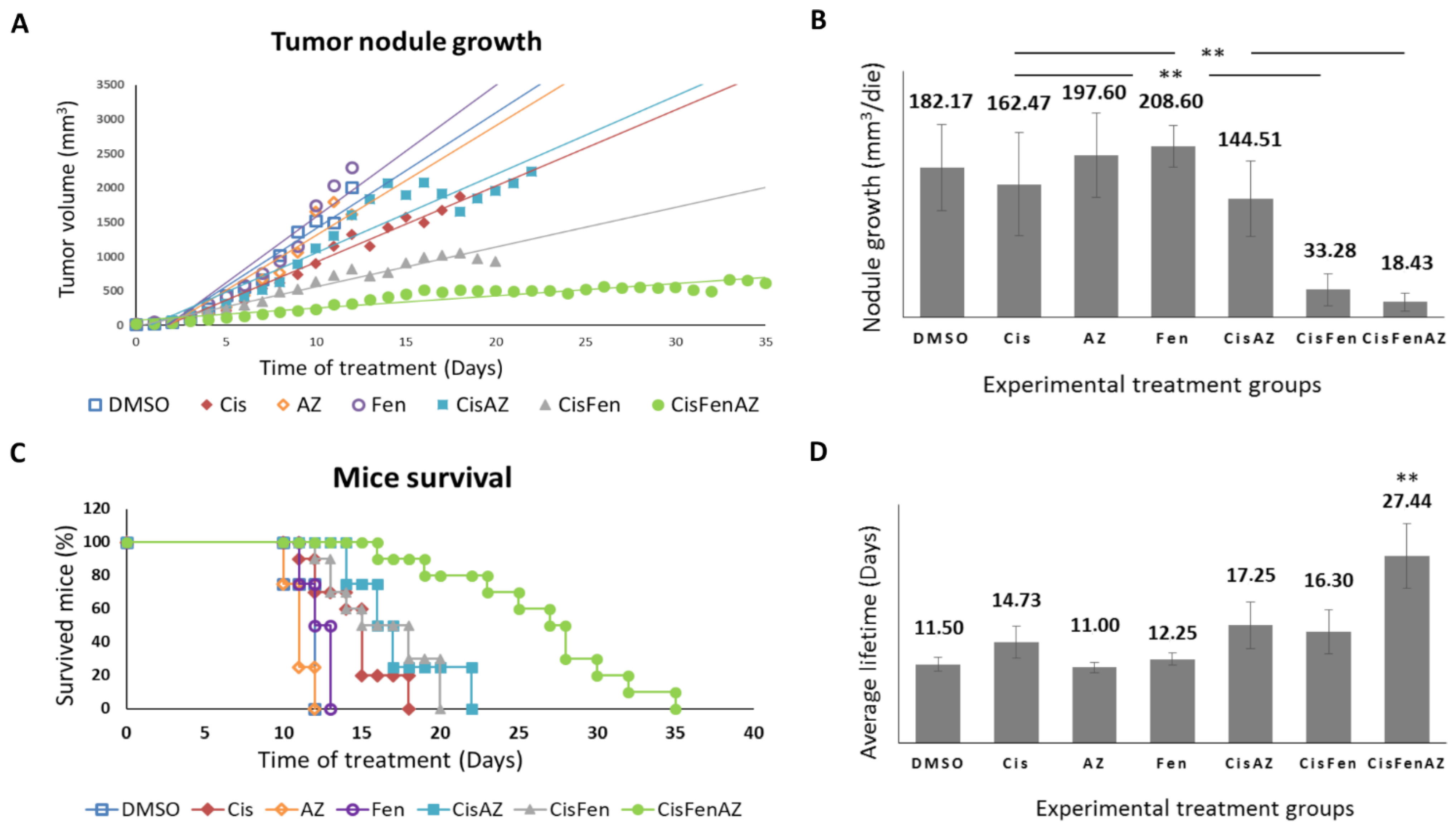
2.1. The Co-Administration of Acetazolamide with Cis/Fen Therapy Increased the Mice’s Survival and Inhibits Tumor Nodule Growth
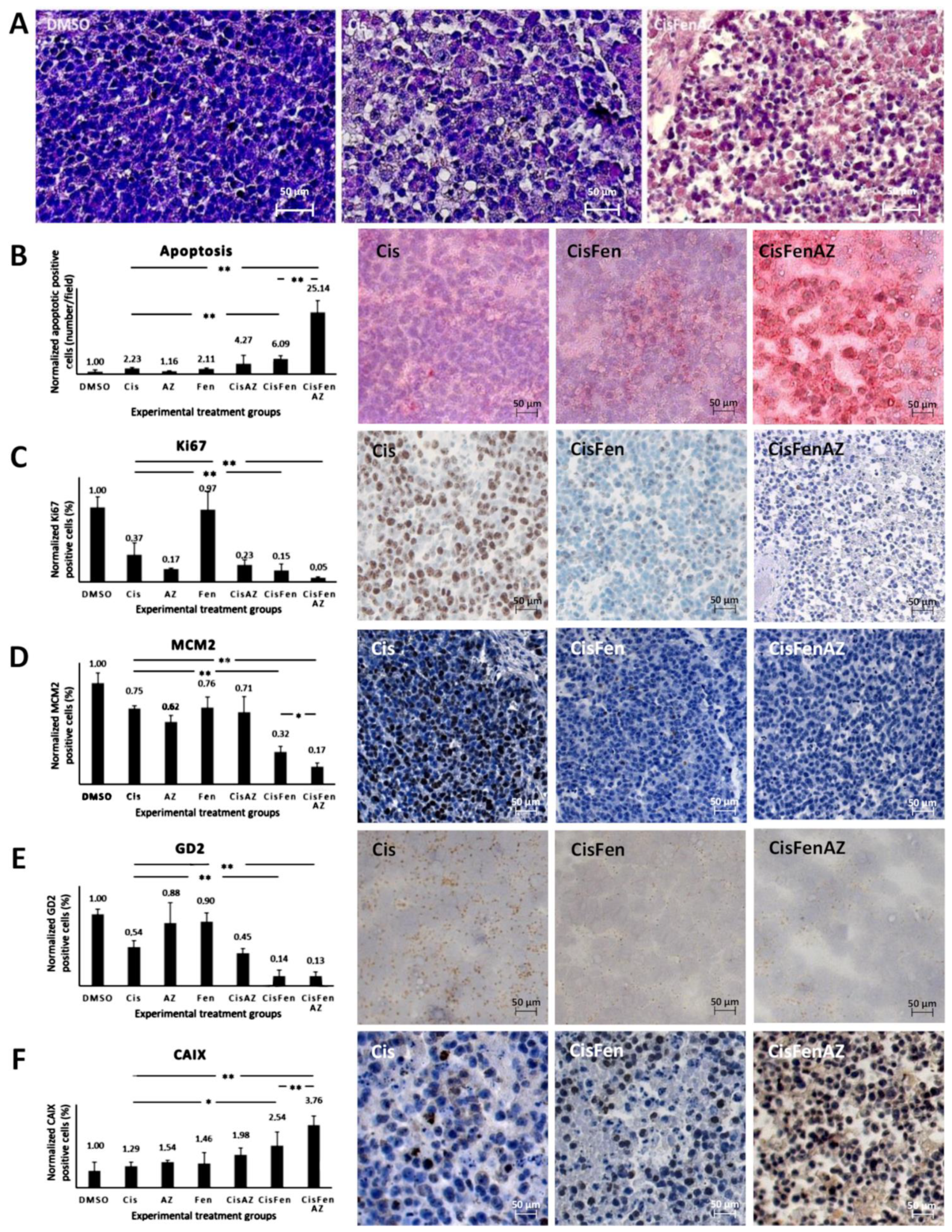
2.2. Acetazolamide in Combination with Cisplatin and Fendiline Hydrochloride Reduces the Expression of Malignancy Markers of NB Xenografts In Vivo
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Cultures
4.3. In Vivo Tumor Formation
4.4. Immunohistochemistry
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Maris, J.M. Recent Advances in Neuroblastoma. N. Engl. J. Med. 2010, 362, 2202–2211. [Google Scholar] [CrossRef] [Green Version]
- Maris, J.M.; Hogarty, M.D.; Bagatell, R.; Cohn, S.L. Neuroblastoma. Lancet 2007, 369, 2106–2120. [Google Scholar] [CrossRef]
- Whittle, S.B.; Smith, V.; Doherty, E.; Zhao, S.; McCarty, S.; Zage, P.E. Overview and recent advances in the treatment of neuroblastoma. Expert Rev. Anticancer Ther. 2017, 17, 369–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Würth, R.; Thellung, S.; Bajetto, A.; Mazzanti, M.; Florio, T.; Barbieri, F. Drug-repositioning opportunities for cancer therapy: Novel molecular targets for known compounds. Drug Discov. Today 2016, 21, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Sidarovich, V.; De Mariano, M.; Aveic, S.; Pancher, M.; Adami, V.; Gatto, P.; Pizzini, S.; Pasini, L.; Croce, M.; Parodi, F.; et al. A high-content screening of anticancer compounds suggests the multiple tyrosine kinase inhibitor ponatinib for repurposing in neuroblastoma therapy. Mol. Cancer Ther. 2018, 17, 1405–1415. [Google Scholar] [CrossRef] [Green Version]
- Vella, S.; Penna, I.; Longo, L.; Pioggia, G.; Garbati, P.; Florio, T.; Rossi, F.; Pagano, A. Perhexiline maleate enhances antitumor efficacy of cisplatin in neuroblastoma by inducing over-expression of NDM29 ncRNA. Sci. Rep. 2015, 5, 18144. [Google Scholar] [CrossRef] [Green Version]
- Brizzolara, A.; Garbati, P.; Vella, S.; Calderoni, M.; Quattrone, A.; Tonini, G.P.; Capasso, M.; Longo, L.; Barbieri, R.; Florio, T.; et al. Co-Administration of Fendiline Hydrochloride Enhances Chemotherapeutic Efficacy of Cisplatin in Neuroblastoma Treatment. Molecules 2020, 25, 5234. [Google Scholar] [CrossRef] [PubMed]
- Gavazzo, P.; Vella, S.; Marchetti, C.; Nizzari, M.; Cancedda, R.; Pagano, A. Acquisition of neuron-like electrophysiological properties in neuroblastoma cells by controlled expression of NDM29 ncRNA. J. Neurochem. 2011, 119, 989–1001. [Google Scholar] [CrossRef] [PubMed]
- Castelnuovo, M.; Massone, S.; Tasso, R.; Fiorino, G.; Gatti, M.; Robello, M.; Gatta, E.; Berger, A.; Strub, K.; Florio, T.; et al. An Alu-like RNA promotes cell differentiation and reduces malignancy of human neuroblastoma cells. FASEB J. 2010, 24, 4033–4046. [Google Scholar] [CrossRef] [Green Version]
- Garbati, P.; Barbieri, R.; Cangelosi, D.; Zanon, C.; Costa, D.; Eva, A.; Thellung, S.; Calderoni, M.; Baldini, F.; Tonini, G.P.; et al. MCM2 and Carbonic Anhydrase 9 Are Novel Potential Targets for Neuroblastoma Pharmacological Treatment. Biomedicines 2020, 8, 471. [Google Scholar] [CrossRef]
- Robertson, N.; Potter, C.; Harris, A.L. Role of carbonic anhydrase IX in human tumor cell growth, survival, and invasion. Cancer Res. 2004, 64, 6160–6165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiry, A.; Dogné, J.M.; Masereel, B.; Supuran, C.T. Targeting tumor-associated carbonic anhydrase IX in cancer therapy. Trends Pharmacol. Sci. 2006, 27, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Vukovic, V.; Tannock, I.F. Influence of low pH on cytotoxicity of paclitaxel, mitoxantrone and topotecan. Br. J. Cancer 1997, 75, 1167–1172. [Google Scholar] [CrossRef] [Green Version]
- Raghunand, N.; He, X.; Van Sluis, R.; Mahoney, B.; Baggett, B.; Taylor, C.W.; Paine-Murrieta, G.; Roe, D.; Bhujwalla, Z.M.; Gillies, R.J. Enhancement of chemotherapy by manipulation of tumour pH. Br. J. Cancer 1999, 80, 1005–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stubbs, M.; McSheehy, P.M.J.; Griffiths, J.R.; Bashford, C.L. Causes and consequences of tumour acidity and implications for treatment. Mol. Med. Today 2000, 6, 15–19. [Google Scholar] [CrossRef]
- Wykoff, C.C.; Beasley, N.J.P.; Watson, P.H.; Turner, K.J.; Pastorek, J.; Sibtain, A.; Wilson, G.D.; Turley, H.; Talks, K.L.; Maxwell, P.H.; et al. Hypoxia-inducible expression of tumor-associated carbonic anhydrases. Cancer Res. 2000, 60, 7075–7083. [Google Scholar]
- Giatromanolaki, A.; Harris, A.L. Tumour hypoxia, hypoxia signaling pathways and hypoxia inducible factor expression in human cancer. Anticancer Res. 2001, 21, 4317–4324. [Google Scholar]
- Dungwa, J.V.; Hunt, L.P.; Ramani, P. Carbonic anhydrase IX up-regulation is associated with adverse clinicopathologic and biologic factors in neuroblastomas. Hum. Pathol. 2012, 43, 1651–1660. [Google Scholar] [CrossRef]
- Benej, M.; Pastorekova, S.; Pastorek, J. Carbonic anhydrase IX: Regulation and role in cancer. Subcell. Biochem. 2014, 75, 199–219. [Google Scholar]
- Islam, S.S.; Mokhtari, R.B.; Akbari, P.; Hatina, J.; Yeger, H.; Farhat, W.A. Simultaneous Targeting of Bladder Tumor Growth, Survival, and Epithelial-to-Mesenchymal Transition with a Novel Therapeutic Combination of Acetazolamide (AZ) and Sulforaphane (SFN). Target. Oncol. 2016, 11, 209–227. [Google Scholar] [CrossRef]
- Kong, B.; Zhao, S.P. Acetazolamide inhibits aquaporin-1 expression and colon cancer xenograft tumor growth. Hepatogastroenterology 2011, 58, 1502–1506. [Google Scholar]
- Bagheri, F.; Safarian, S.; Eslaminejad, M.B.; Sheibani, N. Stable overexpression of DNA fragmentation factor in T-47D cells: Sensitization of breast cancer cells to apoptosis in response to acetazolamide and sulfabenzamide. Mol. Biol. Rep. 2014, 41, 7387–7394. [Google Scholar] [CrossRef] [Green Version]
- Mokhtari, R.B.; Kumar, S.; Islam, S.S.; Yazdanpanah, M.; Adeli, K.; Cutz, E.; Yeger, H. Combination of carbonic anhydrase inhibitor, acetazolamide, and sulforaphane, reduces the viability and growth of bronchial carcinoid cell lines. BMC Cancer 2013, 13, 378. [Google Scholar] [CrossRef] [Green Version]
- Ameis, H.M.; Drenckhan, A.; Freytag, M.; Izbicki, J.R.; Supuran, C.T.; Reinshagen, K.; Holland-Cunz, S.; Gros, S.J. Carbonic anhydrase IX correlates with survival and is a potential therapeutic target for neuroblastoma. J. Enzyme Inhib. Med. Chem. 2016, 31, 404–409. [Google Scholar] [CrossRef]
- Giaginis, C.; Vgenopoulou, S.; Vielh, P.; Theocharis, S. MCM proteins as diagnostic and prognostic tumor markers in the clinical setting. Histol. Histopathol. 2010, 25, 351–370. [Google Scholar] [PubMed]
- Mujoo, K.; Cheresh, D.A.; Yang, H.M.; Reisfeld, R.A. Disialoganglioside GD2 on Human Neuroblastoma Cells: Target Antigen for Monoclonal Antibody-mediated Cytolysis and Suppression of Tumor Growth. Cancer Res. 1987, 47, 1098–1104. [Google Scholar]
- McFarland, B.C.; Benveniste, E.N. Reactive astrocytes foster brain metastases via STAT3 signaling. Ann. Transl. Med. 2019, 7, S83. [Google Scholar] [CrossRef]
- Kamijo, T. Role of stemness-related molecules in neuroblastoma. Pediatr. Res. 2012, 71, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, N.K.V.; Heller, G. Chemotherapy dose intensity correlates strongly with response, median survival, and median progression-free survival in metastatic neuroblastoma. J. Clin. Oncol. 1991, 9, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Troncone, L.; Rufini, V.; Luzi, S.; Mastrangelo, R.; Riccardi, R. The treatment of neuroblastoma with [131I]MIBG at diagnosis. Q. J. Nucl. Med. 1995, 39, 65–68. [Google Scholar] [PubMed]
- Mastrangelo, S.; Tornesello, A.; Diociaiuti, L.; Pession, A.; Prete, A.; Rufini, V.; Troncone, L.; Mastrangelo, R. Treatment of advanced neuroblastoma: Feasibility and therapeutic potential of a novel approach combining 131-I-MIBG and multiple drug chemotherapy. Br. J. Cancer 2001, 84, 460–464. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.W.; Jo, Y.H.; Jee, H.K.; Wu, H.G.; Chae, S.R.; Chol, H.L.; Kim, T.Y.; Dae, S.H.; Bang, Y.J.; Noe, K.K. Neoadjuvant etoposide, ifosfamide, and cisplatin for the treatment of olfactory neuroblastoma. Cancer 2004, 101, 2257–2260. [Google Scholar] [CrossRef]
- Norris, M.D.; Bordow, S.B.; Haber, P.S.; Marshall, G.M.; Kavallaris, M.; Madafiglio, J.; Cohn, S.L.; Salwen, H.; Schmidt, M.L.; Hipfner, D.R.; et al. Evidence that the MYCN oncogene regulates MRP gene expression in neuroblastoma. Eur. J. Cancer 1997, 33, 1911–1916. [Google Scholar] [CrossRef]
- Casinelli, G.; LaRosa, J.; Sharma, M.; Cherok, E.; Banerjee, S.; Branca, M.; Edmunds, L.; Wang, Y.; Sims-Lucas, S.; Churley, L.; et al. N-Myc overexpression increases cisplatin resistance in neuroblastoma via deregulation of mitochondrial dynamics. Cell Death Discov. 2016, 2, 16082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vella, S.; Conaldi, P.G.; Florio, T.; Pagano, A. PPAR Gamma in Neuroblastoma: The Translational Perspectives of Hypoglycemic Drugs. PPAR Res. 2016, 2016, 3038164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, D.; Gigoni, A.; Würth, R.; Cancedda, R.; Florio, T.; Pagano, A. Metformin inhibition of neuroblastoma cell proliferation is differently modulated by cell differentiation induced by retinoic acid or overexpression of NDM29 non-coding RNA. Cancer Cell Int. 2014, 14, 59. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T.; Briganti, F.; Tilli, S.; Chegwidden, W.R.; Scozzafava, A. Carbonic anhydrase inhibitors: Sulfonamides as antitumor agents? Bioorganic Med. Chem. 2001, 9, 703–714. [Google Scholar] [CrossRef]
- Bolden, J.E.; Peart, M.J.; Johnstone, R.W. Anticancer activities of histone deacetylase inhibitors. Nat. Rev. Drug Discov. 2006, 5, 769–784. [Google Scholar] [CrossRef] [PubMed]
- Bayat Mokhtari, R.; Baluch, N.; Ka Hon Tsui, M.; Kumar, S.; Homayouni, S.T.; Aitken, K.; Das, B.; Baruchel, S.; Yeger, H. Acetazolamide potentiates the anti-tumor potential of HDACi, MS-275, in neuroblastoma. BMC Cancer 2017, 17, 156. [Google Scholar] [CrossRef] [Green Version]
- Jan, C.R.; Lee, K.C.; Chou, K.J.; Cheng, J.S.; Wang, J.L.; Lo, Y.K.; Chang, H.T.; Tang, K.Y.; Yu, C.C.; Huang, J.K. Fendiline, an anti-anginal drug, increases intracellular Ca2+ in PC3 human prostate cancer cells. Cancer Chemother. Pharmacol. 2001, 48, 37–41. [Google Scholar]
- Huang, C.C.; Huang, C.J.; Cheng, J.S.; Liu, S.I.; Chen, I.S.; Tsai, J.Y.; Chou, C.T.; Tseng, P.L.; Jan, C.R. Fendiline-evoked [Ca2+]i rises and non-Ca2+-triggered cell death in human oral cancer cells. Hum. Exp. Toxicol. 2009, 28, 41–48. [Google Scholar] [CrossRef] [PubMed]
- van der Hoeven, D.; Cho, K.-J.; Ma, X.; Chigurupati, S.; Parton, R.G.; Hancock, J.F. Fendiline Inhibits K-Ras Plasma Membrane Localization and Blocks K-Ras Signal Transmission. Mol. Cell. Biol. 2013, 33, 237–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, N.; Trevino, J.; Coppola, D.; Chellappan, S.; Yang, S.; Padmanabhan, J. Fendiline inhibits proliferation and invasion of pancreatic cancer cells by interfering with ADAM10 activation and β-catenin signaling. Oncotarget 2015, 6, 35931–35948. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Dong, H.; Li, G.; Jin, H. Combined treatment with acetazolamide and cisplatin enhances chemosensitivity in laryngeal carcinoma Hep-2 cells. Oncol. Lett. 2018, 15, 9299–9306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, D.; Estlin, E.J.; Dive, C.; Makin, G.W.J. Chronic hypoxia promotes hypoxia-inducible factor-1-α-dependent resistance to etoposide and vincristine in neuroblastoma cells. Mol. Cancer Ther. 2006, 5, 2241–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Tuominen, V.J.; Ruotoistenmäki, S.; Viitanen, A.; Jumppanen, M.; Isola, J. ImmunoRatio: A publicly available web application for quantitative image analysis of estrogen receptor (ER), progesterone receptor (PR), and Ki-67. Breast Cancer Res. 2010, 12, R56. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garbati, P.; Barbieri, R.; Calderoni, M.; Baldini, F.; Nizzari, M.; Modesto, P.; Florio, T.; Pagano, A. Efficacy of a Three Drug-Based Therapy for Neuroblastoma in Mice. Int. J. Mol. Sci. 2021, 22, 6753. https://doi.org/10.3390/ijms22136753
Garbati P, Barbieri R, Calderoni M, Baldini F, Nizzari M, Modesto P, Florio T, Pagano A. Efficacy of a Three Drug-Based Therapy for Neuroblastoma in Mice. International Journal of Molecular Sciences. 2021; 22(13):6753. https://doi.org/10.3390/ijms22136753
Chicago/Turabian StyleGarbati, Patrizia, Raffaella Barbieri, Matilde Calderoni, Francesca Baldini, Mario Nizzari, Paola Modesto, Tullio Florio, and Aldo Pagano. 2021. "Efficacy of a Three Drug-Based Therapy for Neuroblastoma in Mice" International Journal of Molecular Sciences 22, no. 13: 6753. https://doi.org/10.3390/ijms22136753