
Corticosterone Administration Alters White Matter Tract Structure and Reduces Gliosis in the Sub-Acute Phase of Experimental Stroke

 , ,
, ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
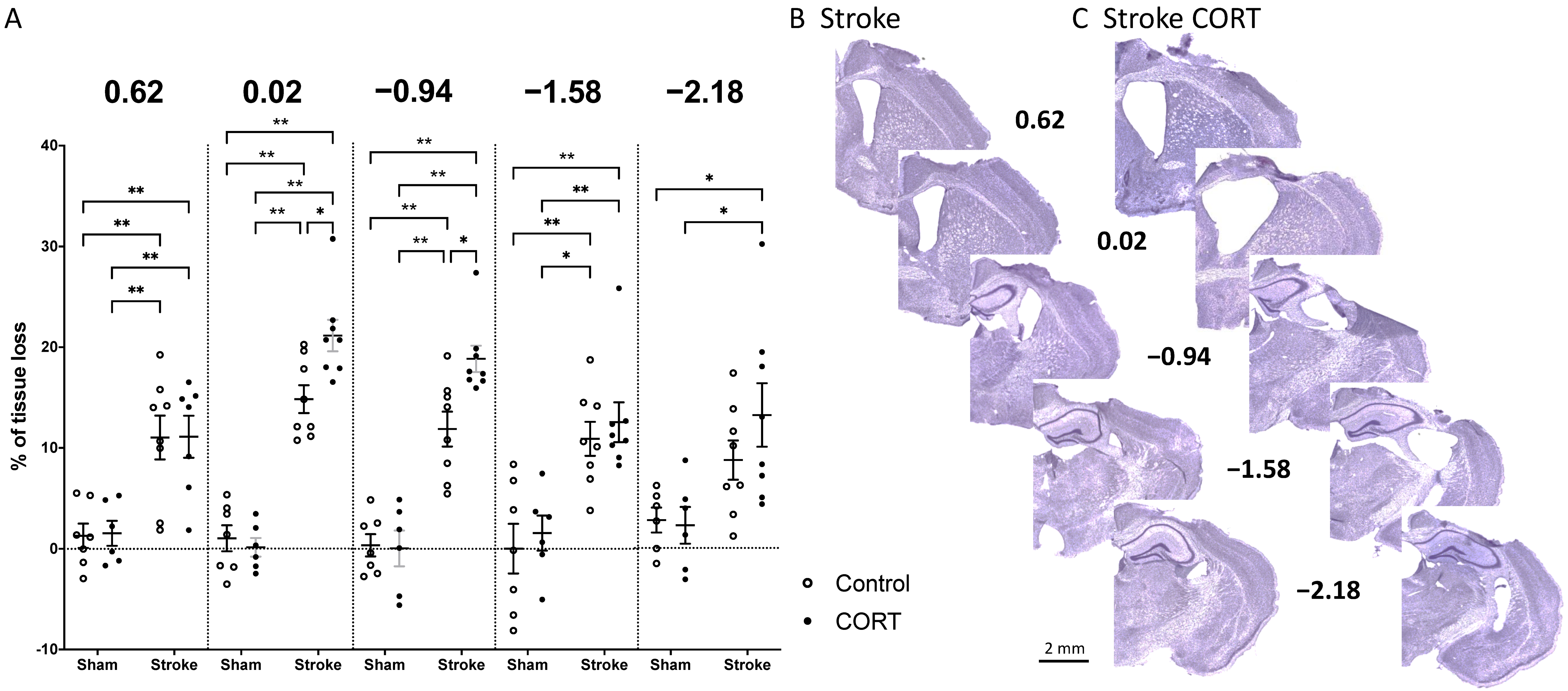
2.1. Oral CORT Administration Post-Stroke Exacerbates Tissue Loss
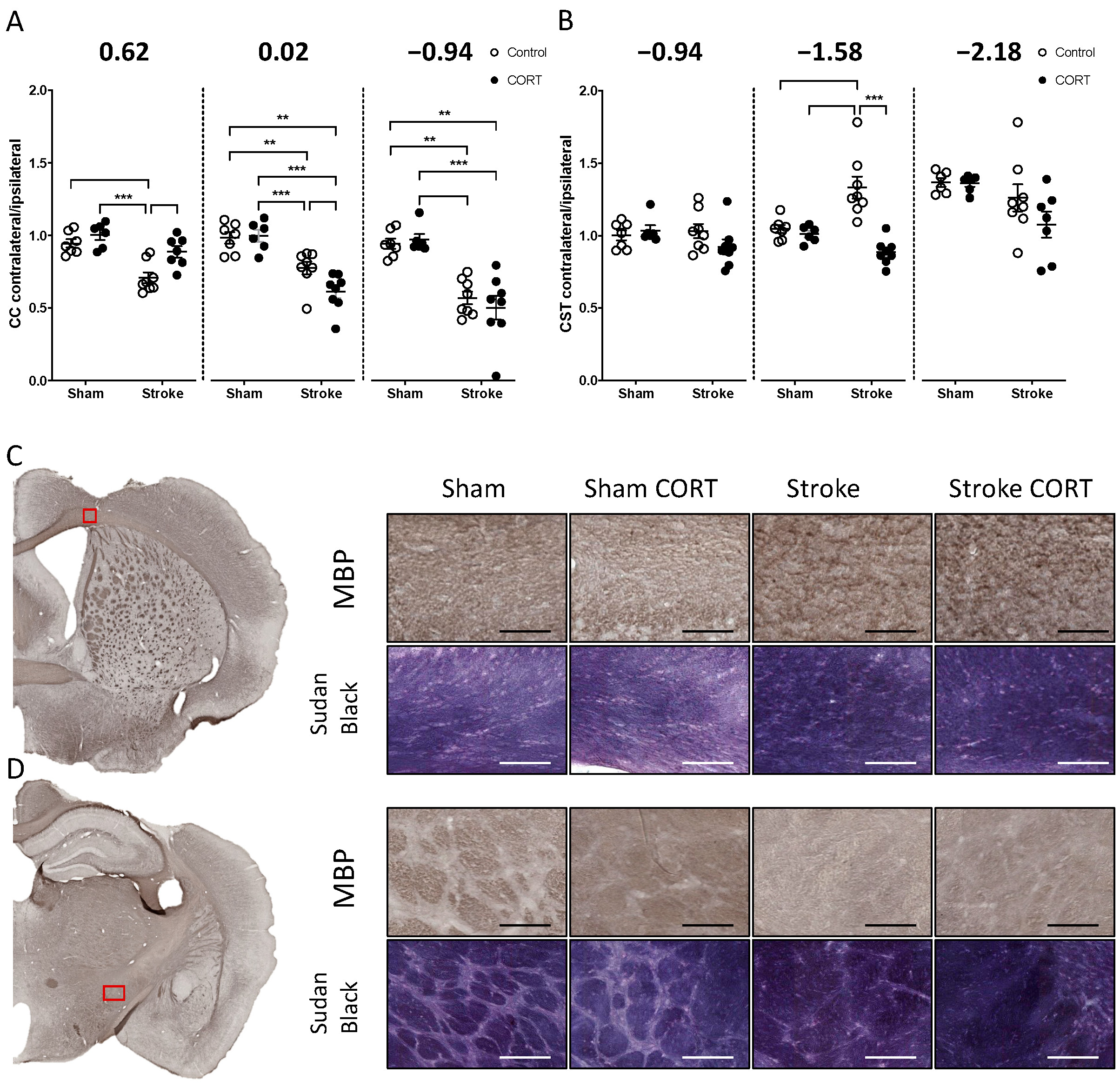
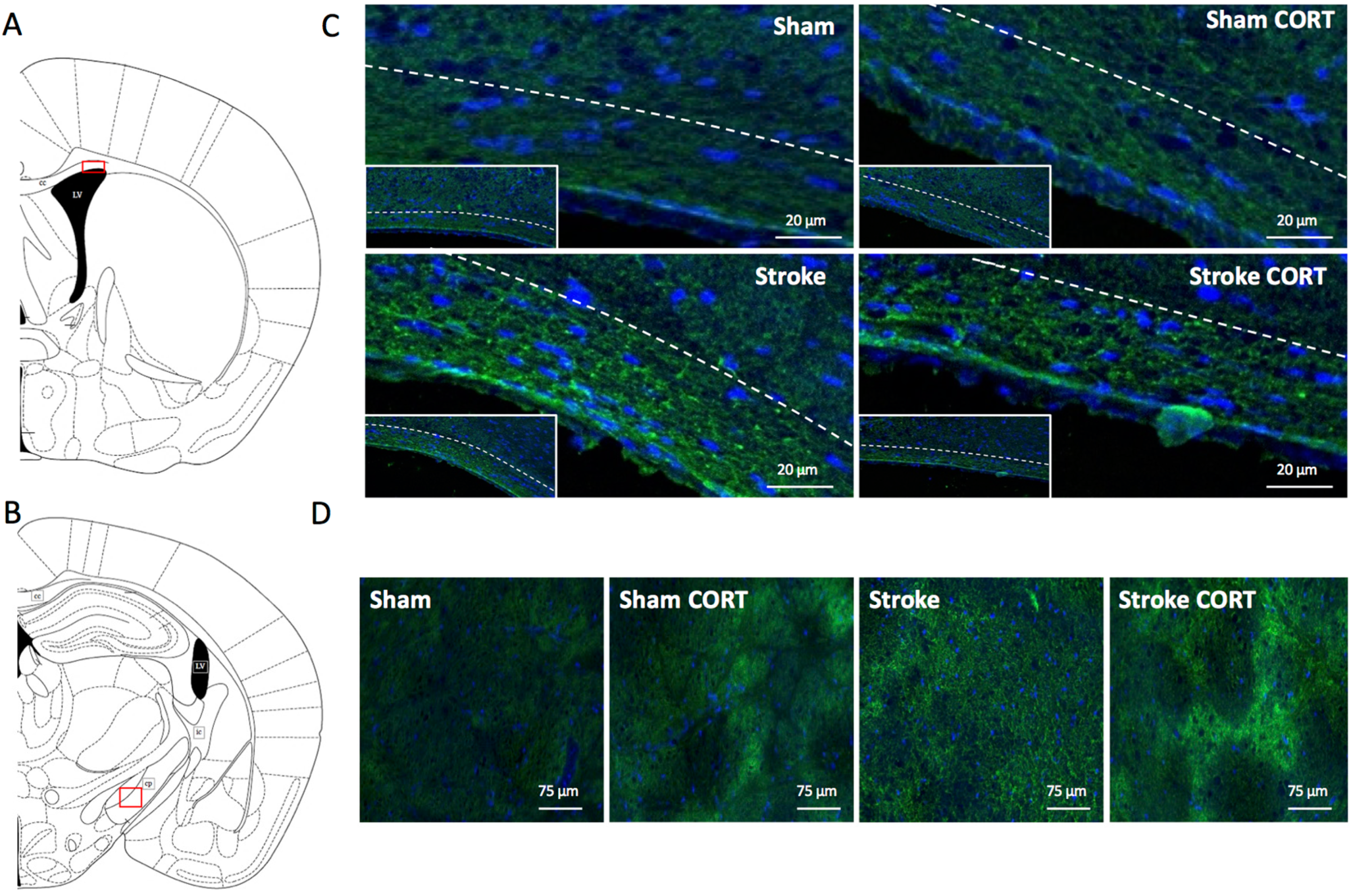
2.2. Oral CORT Administration Alters the Structure of the Corpus Callosum (CC) and Corticospinal Tract (CST)
2.3. Both CORT and Stroke Alter Myelin Structure
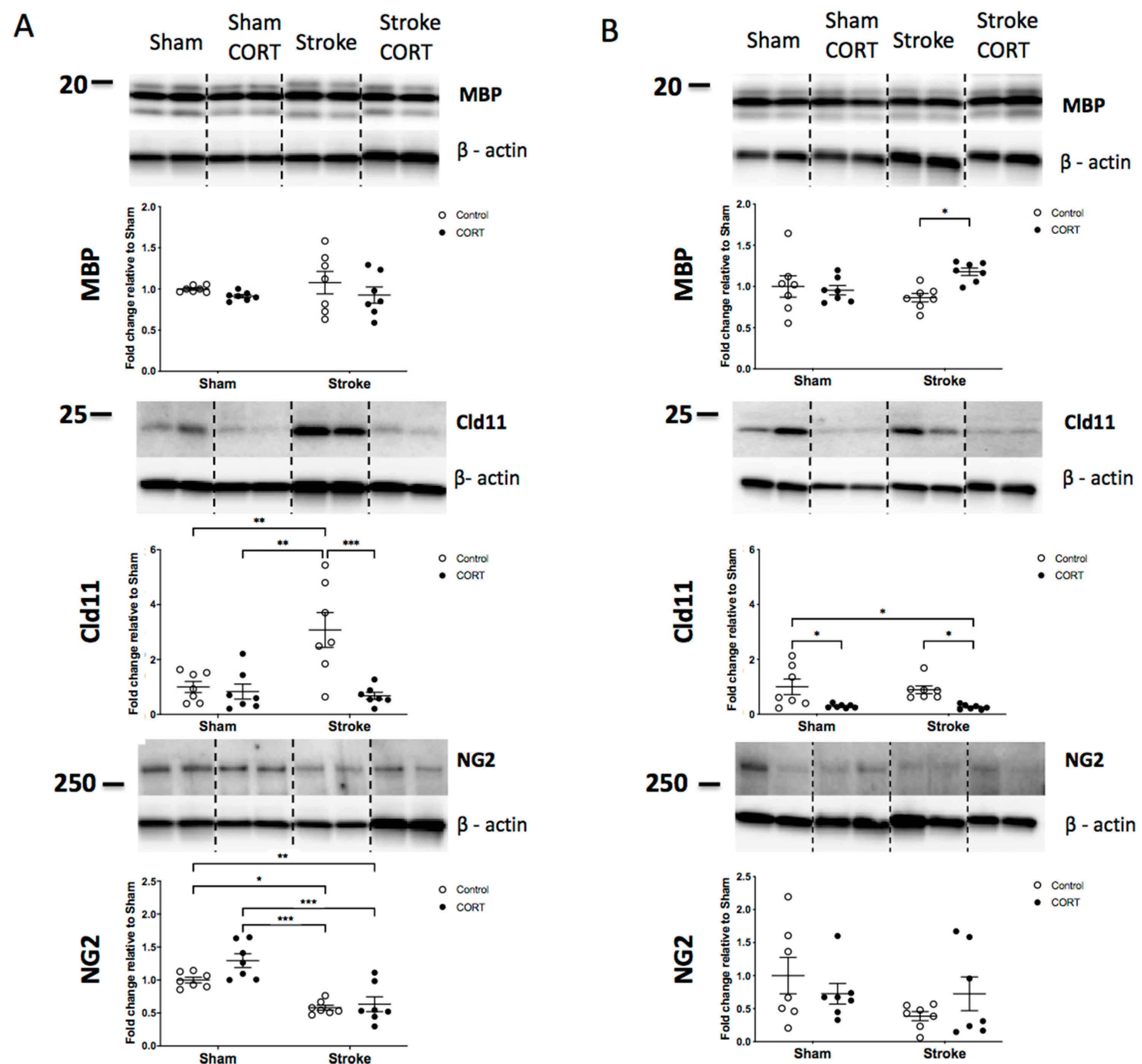
2.4. Both CORT and Stroke Have no Effect on Total Level of MBP Protein
2.5. Oral CORT Administration Reduces Expression of Claudin-11
2.6. Stroke Induction Reduces Level of NG2
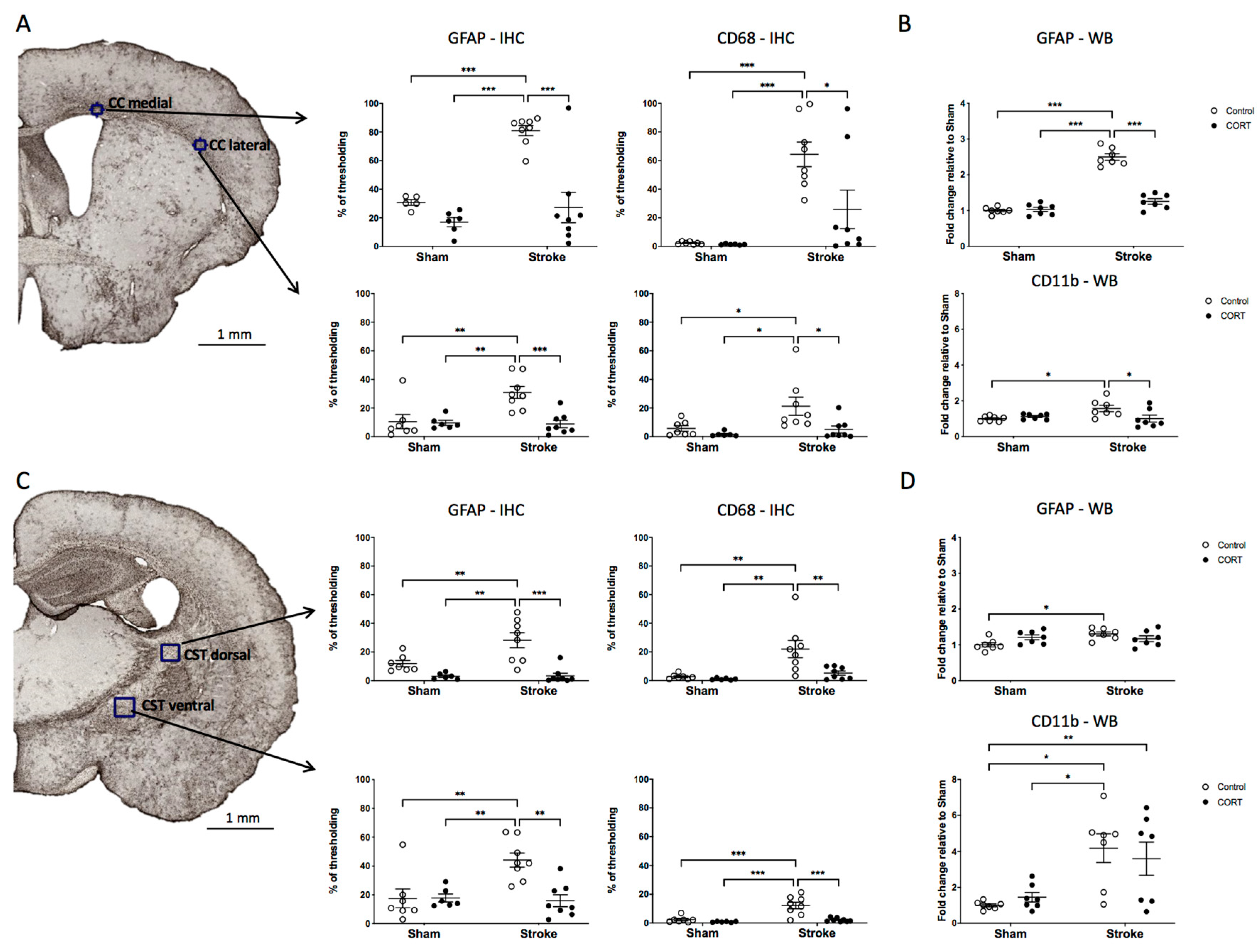
2.7. Oral CORT Administration Reduces the Expression of Microglial and Astrocytic Markers
2.8. Axonal Degeneration is Enhanced in Stroke Animals Treated with CORT in the CST; However, Not the CC
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Animals
4.3. Experimental Design
4.4. Experimental Stroke and Oral Delivery of Corticosterone (CORT)
4.5. Tissue Processing
4.6. Histology and Immunohistochemistry
4.7. Image Acquisition and Analysis
4.8. Protein Extraction and Western Blotting
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Liu, G.; Hong, D.; Chen, F.; Ji, X.; Cao, G. White matter injury in ischemic stroke. Prog. Neurobiol. 2016, 141, 45–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Gao, H.; Zeng, P.; Jia, Y.; Kong, X.; Xu, K.; Bai, R. Secondary degeneration of white matter after focal sensorimotor cortical ischemic stroke in rats. Front. Neurosci. 2021, 14, 611696. [Google Scholar] [CrossRef] [PubMed]
- Visser, M.M.; Yassi, N.; Campbell, B.C.; Desmond, P.M.; Davis, S.M.; Spratt, N.; Parsons, M.; Bivard, A. White matter degeneration after ischemic stroke: A longitudinal diffusion tensor imaging study. J. Neuroimaging 2018, 29, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Thomalla, G.; Glauche, V.; Weiller, C.; Röther, J. Time course of wallerian degeneration after ischaemic stroke revealed by diffusion tensor imaging. J. Neurol. Neurosurg. Psychiatry 2005, 76, 266–268. [Google Scholar] [CrossRef] [Green Version]
- Pantoni, L.; Garcia, J.H.; Gutierrez, J.A. Cerebral white matter is highly vulnerable to ischemia. Stroke 1996, 27, 1641–1646. [Google Scholar] [CrossRef] [PubMed]
- Pietrogrande, G.; Zalewska, K.; Zhao, Z.; Abdolhoseini, M.; Chow, W.Z.; Sanchez-Bezanilla, S.; Ong, L.K.; Johnson, S.J.; Nilsson, M.; Walker, F.R. Low oxygen post conditioning prevents thalamic secondary neuronal loss caused by excitotoxicity after cortical stroke. Sci. Rep. 2019, 9, 4841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Bezanilla, S.; Åberg, N.D.; Crock, P.; Walker, F.R.; Nilsson, M.; Isgaard, J.; Ong, L.K. Growth hormone treatment promotes remote hippocampal plasticity after experimental cortical stroke. Int. J. Mol. Sci. 2020, 21, 4563. [Google Scholar] [CrossRef]
- Sanchez-Bezanilla, S.; Hood, R.J.; Collins-Praino, L.E.; Turner, R.J.; Walker, F.R.; Nilsson, M.; Ong, L.K. More than motor impairment: A spatiotemporal analysis of cognitive impairment and associated neuropathological changes following cortical photothrombotic stroke. J. Cereb. Blood Flow Metab. 2021, 271678X211005877. [Google Scholar]
- Lindberg, P.G.; Skejø, P.H.B.; Rounis, E.; Nagy, Z.; Schmitz, C.; Wernegren, H.; Bring, A.; Engardt, M.; Forssberg, H.; Borg, J. Wallerian Degeneration of the Corticofugal Tracts in Chronic Stroke: A Pilot Study Relating Diffusion Tensor Imaging, Transcranial Magnetic Stimulation, and Hand Function. Neurorehabilit. Neural Repair 2007, 21, 551–560. [Google Scholar] [CrossRef]
- Yu, C.; Zhu, C.; Zhang, Y.; Chen, H.; Qin, W.; Wang, M.; Li, K. A longitudinal diffusion tensor imaging study on Wallerian degeneration of corticospinal tract after motor pathway stroke. NeuroImage 2009, 47, 451–458. [Google Scholar] [CrossRef]
- Guo, A.H.; Hao, F.L.; Liu, L.F.; Wang, B.J.; Jiang, X.F. An assessment of the correlation between early postinfarction pyramidal tract Wallerian degeneration and nerve function recovery using diffusion tensor imaging. Genet. Mol. Res. 2017, 16, gmr16019035. [Google Scholar] [CrossRef]
- Puig, J.; Pedraza, S.; Blasco, G.; Daunis-I-Estadella, P.; Prats-Galino, A.; Prados, F.; Boada, I.; Castellanos, M.D.M.; Sánchez-González, J.; Remollo, S.; et al. Wallerian degeneration in the corticospinal tract evaluated by diffusion tensor imaging correlates with motor deficit 30 days after middle cerebral artery ischemic stroke. Am. J. Neuroradiol. 2010, 31, 1324–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jokinen, H.; Kalska, H.; Mäntylä, R.; Ylikoski, R.; Hietanen, M.; Pohjasvaara, T.; Kaste, M.; Erkinjuntti, T. White matter hyperintensities as a predictor of neuropsychological deficits post-stroke. J. Neurol. Neurosurg. Psychiatry 2005, 76, 1229–1233. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.K.; Miller, B.L.; Cummings, J.L.; Mehringer, C.M.; Goldberg, M.A.; Howng, S.L.; Benson, D.F. A quantitative MRI study of vascular dementia. Neurology 1992, 42, 138. [Google Scholar] [CrossRef] [PubMed]
- Grau-Olivares, M.; Arboix, A. Mild cognitive impairment in stroke patients with ischemic cerebral small-vessel disease: A forerunner of vascular dementia? Expert Rev. Neurother. 2009, 9, 1201–1217. [Google Scholar] [CrossRef] [PubMed]
- Walker, F.R.; Jones, K.; Patience, M.J.; Zhao, Z.; Nilsson, M. Stress as Necessary component of realistic recovery in animal models of experimental stroke. J. Cereb. Blood Flow Metab. 2014, 34, 208–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilari, K.; Northcott, S.; Roy, P.; Marshall, J.; Wiggins, R.D.; Chataway, J.; Ames, D. Psychological distress after stroke and aphasia: The first six months. Clin. Rehabil. 2010, 24, 181–190. [Google Scholar] [CrossRef]
- Gyawali, P.; Chow, W.Z.; Hinwood, M.; Kluge, M.; English, C.; Ong, L.K.; Nilsson, M.; Walker, F.R. Opposing associations of stress and resilience with functional outcomes in stroke survivors in the chronic phase of stroke: A cross-sectional study. Front. Neurol. 2020, 11, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feibel, J.H.; Hardy, P.M.; Campbell, R.G.; Goldstein, M.N.; Joynt, R.J. Prognostic value of the stress response following stroke. JAMA 1977, 238, 1374–1376. [Google Scholar] [CrossRef]
- Ostwald, S.K.; Swank, P.R.; Khan, M.M. Predictors of functional independence and stress level of stroke survivors at discharge from inpatient rehabilitation. J. Cardiovasc. Nurs. 2008, 23, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Barugh, A.J.; Gray, P.; Shenkin, S.D.; MacLullich, A.M.J.; Mead, G.E. Cortisol levels and the severity and outcomes of acute stroke: A systematic review. J. Neurol. 2014, 261, 533–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Assayag, E.; Korczyn, A.D.; Giladi, N.; Goldbourt, U.; Berliner, A.S.; Shenhar-Tsarfaty, S.; Kliper, E.; Hallevi, H.; Shopin, L.; Hendler, T.; et al. Predictors for poststroke outcomes: The Tel Aviv brain acute stroke cohort (TABASCO) study protocol. Int. J. Stroke 2011, 7, 341–347. [Google Scholar] [CrossRef]
- Jones, K.; Zouikr, I.; Patience, M.; Clarkson, A.N.; Isgaard, J.; Johnson, S.J.; Spratt, N.; Nilsson, M.; Walker, F.R. Chronic stress exacerbates neuronal loss associated with secondary neurodegeneration and suppresses microglial-like cells following focal motor cortex ischemia in the mouse. Brain Behav. Immun. 2015, 48, 57–67. [Google Scholar] [CrossRef]
- Ong, L.K.; Zhao, Z.; Kluge, M.; Walker, F.R.; Nilsson, M. Chronic stress exposure following photothrombotic stroke is associated with increased levels of Amyloid beta accumulation and altered oligomerisation at sites of thalamic secondary neurodegeneration in mice. J. Cereb. Blood Flow Metab. 2016, 37, 1338–1348. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Ong, L.K.; Johnson, S.; Nilsson, M.; Walker, F.R. Chronic stress induced disruption of the peri-infarct neurovascular unit following experimentally induced photothrombotic stroke. J. Cereb. Blood Flow Metab. 2017, 37, 3709–3724. [Google Scholar] [CrossRef] [Green Version]
- Zalewska, K.; Ong, L.K.; Johnson, S.J.; Nilsson, M.; Walker, F.R. Oral administration of corticosterone at stress-like levels drives microglial but not vascular disturbances post-stroke. Neuroscience 2017, 352, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Zalewska, K.; Pietrogrande, G.; Ong, L.K.; Abdolhoseini, M.; Kluge, M.; Johnson, S.J.; Walker, F.R.; Nilsson, M. Sustained administration of corticosterone at stress-like levels after stroke suppressed glial reactivity at sites of thalamic secondary neurodegeneration. Brain Behav. Immun. 2018, 69, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, R.; Bourgin, J.; Boumezbeur, F.; Marques, P.; Bottlaender, M.; Poupon, C.; Djemaï, B.; Duchesnay, E.; Mériaux, S.; Sousa, N.; et al. White matter changes in microstructure associated with a maladaptive response to stress in rats. Transl. Psychiatry 2017, 7, e1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniels, J.K.; Lamke, J.-P.; Gaebler, M.; Walter, H.; Scheel, M. White matter integrity and its relationship to PTSD and childhood trauma—A systematic review and meta-analysis. Depress. Anxiety 2013, 30, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.R.; Bastin, M.E.; Ferguson, K.J.; Maniega, S.M.; MacPherson, S.E.; Deary, I.J.; Wardlaw, J.M.; MacLullich, A. Brain white matter integrity and cortisol in older men: The Lothian Birth Cohort 1936. Neurobiol. Aging 2015, 36, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Cox, S.R.; MacPherson, S.E.; Ferguson, K.J.; Royle, N.A.; Maniega, S.M.; Hernández, M.D.C.V.; Bastin, M.E.; MacLullich, A.M.; Wardlaw, J.M.; Deary, I.J. Does white matter structure or hippocampal volume mediate associations between cortisol and cognitive ageing? Psychoneuroendocrinology 2015, 62, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Watanabe, K.; Kakeda, S.; Yoshimura, R.; Abe, O.; Ide, S.; Hayashi, K.; Katsuki, A.; Umene-Nakano, W.; Watanabe, R.; et al. Relationship between white matter integrity and serum cortisol levels in drug-naive patients with major depressive disorder: Diffusion tensor imaging study using tract-based spatial statistics. Br. J. Psychiatry 2016, 208, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Po, C.; Kalthoff, D.; Kim, Y.B.; Nelles, M.; Hoehn, M. White matter reorganization and functional response after focal cerebral ischemia in the rat. PLoS ONE 2012, 7, e45629. [Google Scholar] [CrossRef] [Green Version]
- Kongsui, R.; Johnson, S.J.; Graham, B.A.; Nilsson, M.; Walker, F.R. A combined cumulative threshold spectra and digital reconstruction analysis reveal structural alterations of microglia within the prefrontal cortex following low-dose LPS administration. Neuroscience 2015, 310, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Ehara, A.; Ueda, S. Application of fluoro-jade C in acute and chronic neurodegeneration models: Utilities and staining differences. Acta Histochem. ET Cytochem. 2009, 42, 171–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frid, K.; Einstein, O.; Friedman-Levi, Y.; Binyamin, O.; Ben-Hur, T.; Gabizon, R. Aggregation of MBP in chronic demyelination. Ann. Clin. Transl. Neurol. 2015, 2, 711–721. [Google Scholar] [CrossRef]
- Zhu, X.; Hill, R.A.; Nishiyama, A. NG2 cells generate oligodendrocytes and gray matter astrocytes in the spinal cord. Neuron Glia Biol. 2008, 4, 19–26. [Google Scholar] [CrossRef]
- Chetty, S.; Friedman, A.R.; Taravosh-Lahn, K.; Kirby, E.D.; Mirescu, C.; Guo, F.; Krupik, D.; Nicholas, A.; Geraghty, A.C.; Krishnamurthy, A.; et al. Stress and glucocorticoids promote oligodendrogenesis in the adult hippocampus. Mol. Psychiatry 2014, 19, 1275–1283. [Google Scholar] [CrossRef] [Green Version]
- Wilber, A.A.; Walker, A.G.; Southwood, C.J.; Farrell, M.R.; Lin, G.L.; Rebec, G.V.; Wellman, C.L. Chronic stress alters neural activity in medial prefrontal cortex during retrieval of extinction. Neuroscience 2011, 174, 115–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borstad, A.L.; Choi, S.; Schmalbrock, P.; Nichols-Larsen, D.S. Frontoparietal white matter integrity predicts haptic performance in chronic stroke. NeuroImage Clin. 2016, 10, 129–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakhani, B.; Hayward, K.; Boyd, L.A. Hemispheric asymmetry in myelin after stroke is related to motor impairment and function. NeuroImage Clin. 2017, 14, 344–353. [Google Scholar] [CrossRef]
- Hafsteinsdottir, T.; Grypdonck, M. Being a stroke patient: A review of the literature. J. Adv. Nurs. 1997, 26, 580–588. [Google Scholar] [CrossRef]
- Nugent, K.L.; Chiappelli, J.; Sampath, H.; Rowland, L.; Thangavelu, K.; Davis, B.; Du, X.; Muellerklein, F.; Daughters, S.; Kochunov, P.; et al. Cortisol reactivity to stress and its association with white matter integrity in adults with schizophrenia. Psychosom. Med. 2015, 77, 733–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheikh, H.I.; Joanisse, M.F.; Mackrell, S.M.; Kryski, K.R.; Smith, H.J.; Singh, S.M.; Hayden, E.P. Links between white matter microstructure and cortisol reactivity to stress in early childhood: Evidence for moderation by parenting. NeuroImage Clin. 2014, 6, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Arboix, A.; Cartanyà, A.; Lowak, M.; García-Eroles, L.; Parra, O.; Oliveres, M.; Massons, J. Gender differences and woman-specific trends in acute stroke: Results from a hospital-based registry (1986–2009). Clin. Neurol. Neurosurg. 2014, 127, 19–24. [Google Scholar] [CrossRef]
- Kudielka, B.M.; Kirschbaum, C. Sex differences in HPA axis responses to stress: A review. Biol. Psychol. 2005, 69, 113–132. [Google Scholar] [CrossRef] [PubMed]
- Kluge, M.G.; Kracht, L.; Abdolhoseini, M.; Ong, L.K.; Johnson, S.J.; Nilsson, M.; Walker, F.R. Impaired microglia process dynamics post-stroke are specific to sites of secondary neurodegeneration. Glia 2017, 65, 1885–1899. [Google Scholar] [CrossRef]
- Pietrogrande, G.; Mabotuwana, N.; Zhao, Z.; Abdolhoseini, M.; Johnson, S.J.; Nilsson, M.; Walker, F.R. Chronic stress induced disturbances in Laminin: A significant contributor to modulating microglial pro-inflammatory tone? Brain Behav. Immun. 2018, 68, 23–33. [Google Scholar] [CrossRef]
- Perry, A.; Brat, D.J. Practical Surgical Neuropathology: A Diagnostic Approach; Churchill Livingstone/Elsevier: Philadelphia, PA, USA, 2010. [Google Scholar]
- Kiernan, J. Histochemistry of Staining methods for normal and degenerating myelin in the central and peripheral nervous systems. J. Histotechnol. 2007, 30, 87–106. [Google Scholar] [CrossRef]
- Ong, L.K.; Zhao, Z.; Kluge, M.; TeBay, C.; Zalewska, K.; Dickson, P.W.; Johnson, S.J.; Nilsson, M.; Walker, F.R. Reconsidering the role of glial cells in chronic stress-induced dopaminergic neurons loss within the substantia nigra? Friend or foe? Brain Behav. Immun. 2017, 60, 117–125. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zalewska, K.; Hood, R.J.; Pietrogrande, G.; Sanchez-Bezanilla, S.; Ong, L.K.; Johnson, S.J.; Young, K.M.; Nilsson, M.; Walker, F.R. Corticosterone Administration Alters White Matter Tract Structure and Reduces Gliosis in the Sub-Acute Phase of Experimental Stroke. Int. J. Mol. Sci. 2021, 22, 6693. https://doi.org/10.3390/ijms22136693
Zalewska K, Hood RJ, Pietrogrande G, Sanchez-Bezanilla S, Ong LK, Johnson SJ, Young KM, Nilsson M, Walker FR. Corticosterone Administration Alters White Matter Tract Structure and Reduces Gliosis in the Sub-Acute Phase of Experimental Stroke. International Journal of Molecular Sciences. 2021; 22(13):6693. https://doi.org/10.3390/ijms22136693
Chicago/Turabian StyleZalewska, Katarzyna, Rebecca J. Hood, Giovanni Pietrogrande, Sonia Sanchez-Bezanilla, Lin Kooi Ong, Sarah J. Johnson, Kaylene M. Young, Michael Nilsson, and Frederick R. Walker. 2021. "Corticosterone Administration Alters White Matter Tract Structure and Reduces Gliosis in the Sub-Acute Phase of Experimental Stroke" International Journal of Molecular Sciences 22, no. 13: 6693. https://doi.org/10.3390/ijms22136693