
Using Diatom and Apicomplexan Models to Study the Heme Pathway of Chromera velia

 , , ,
, , , 
Abstract
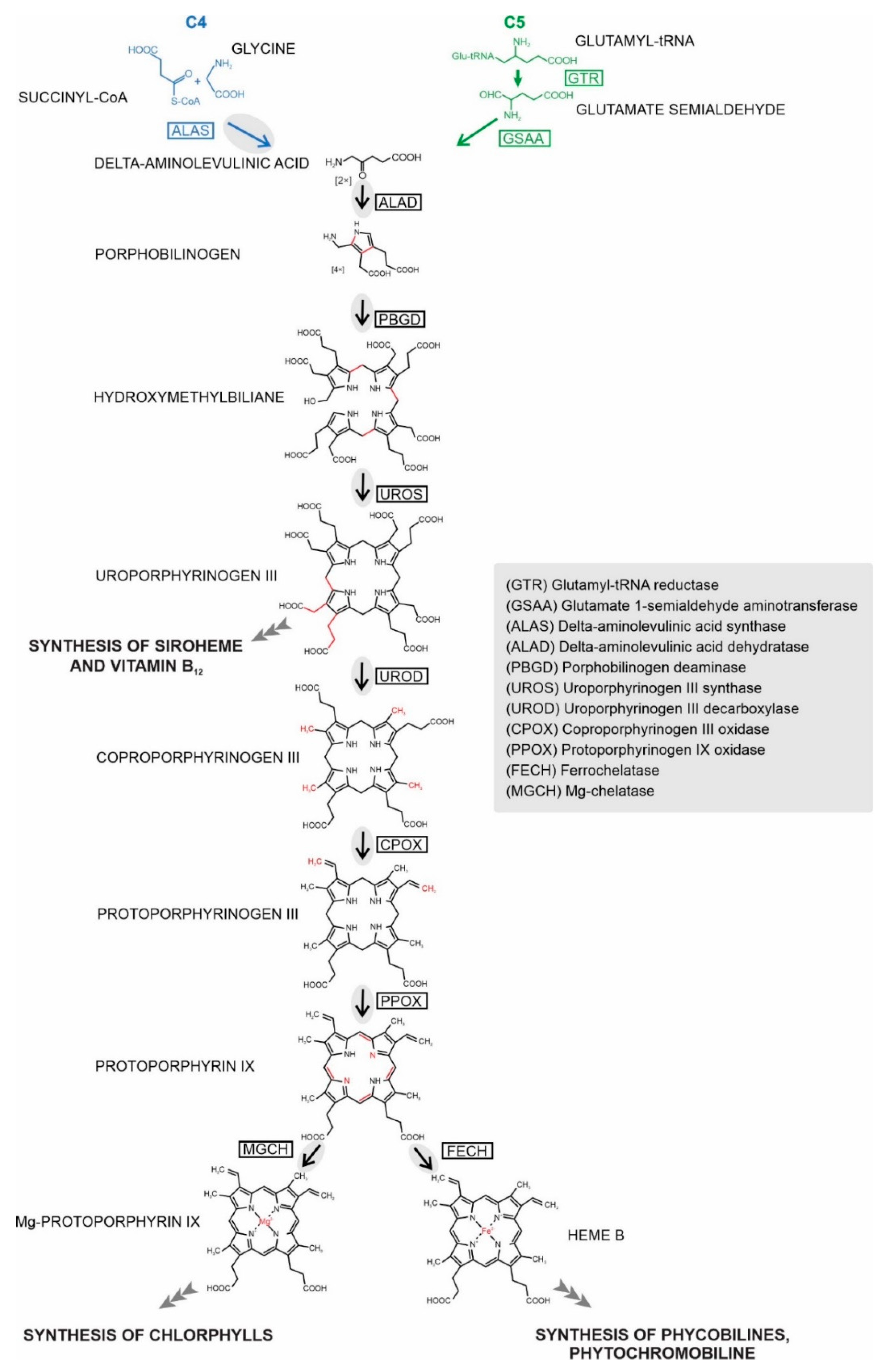
:1. Introduction
2. Results
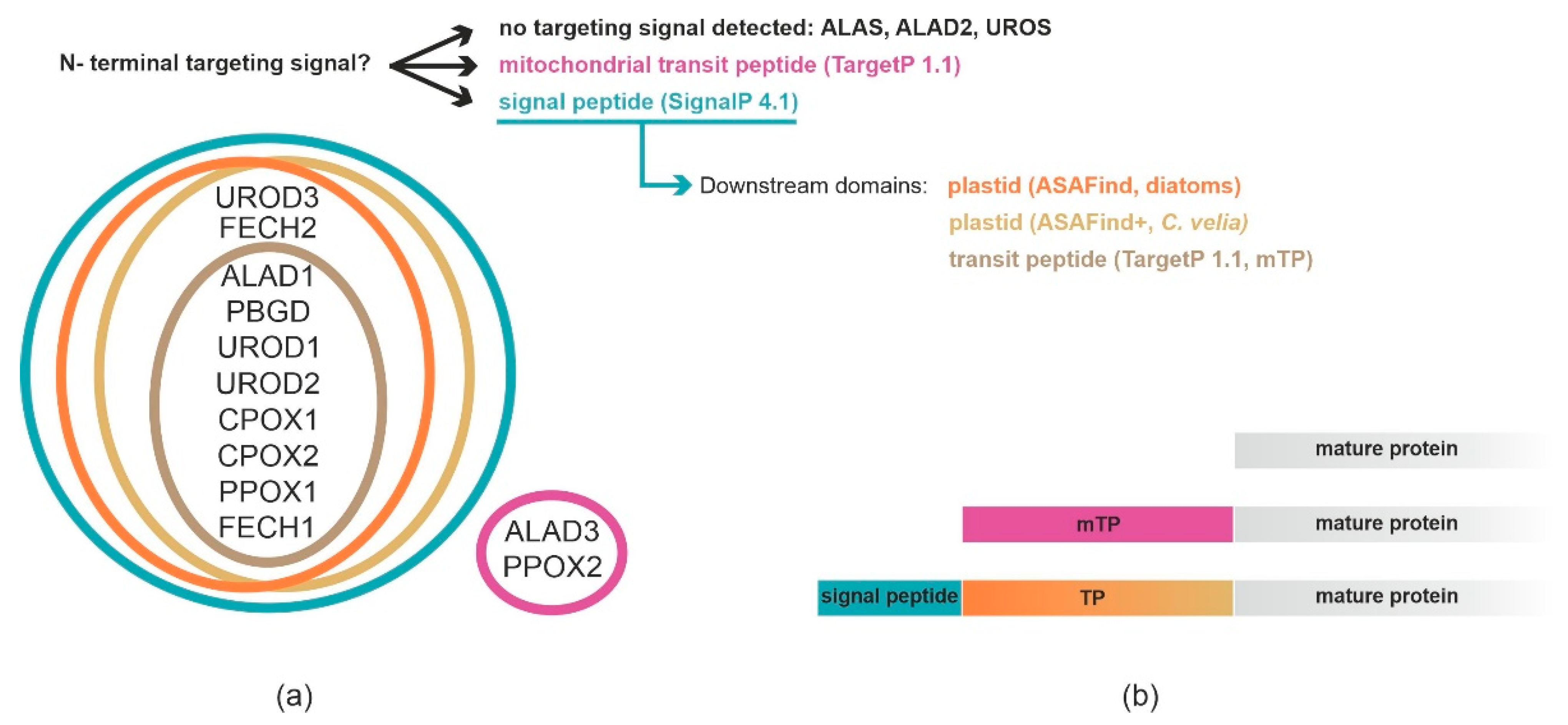
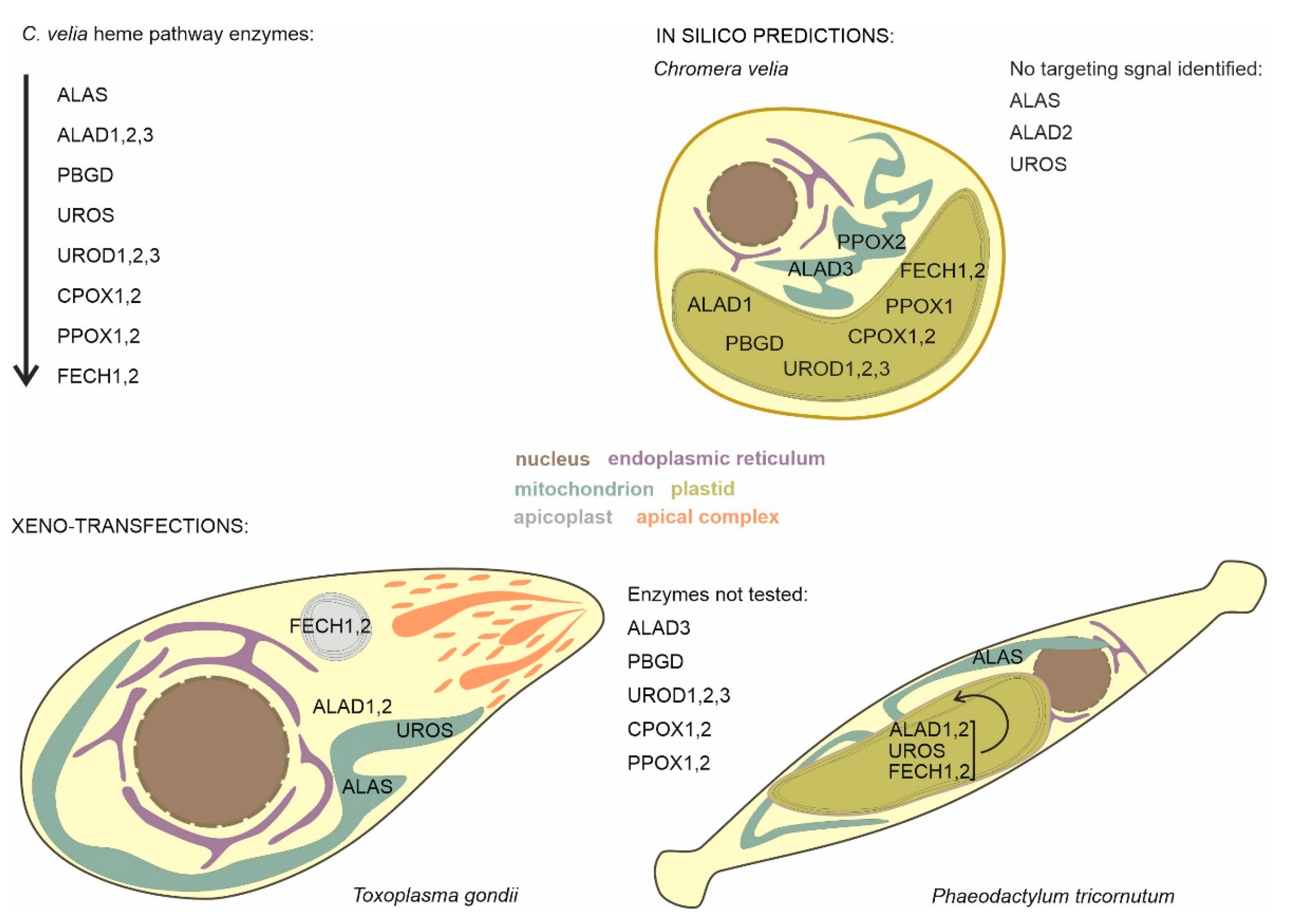
2.1. Prediction of Localization of Heme Synthesis Enzymes in C. velia
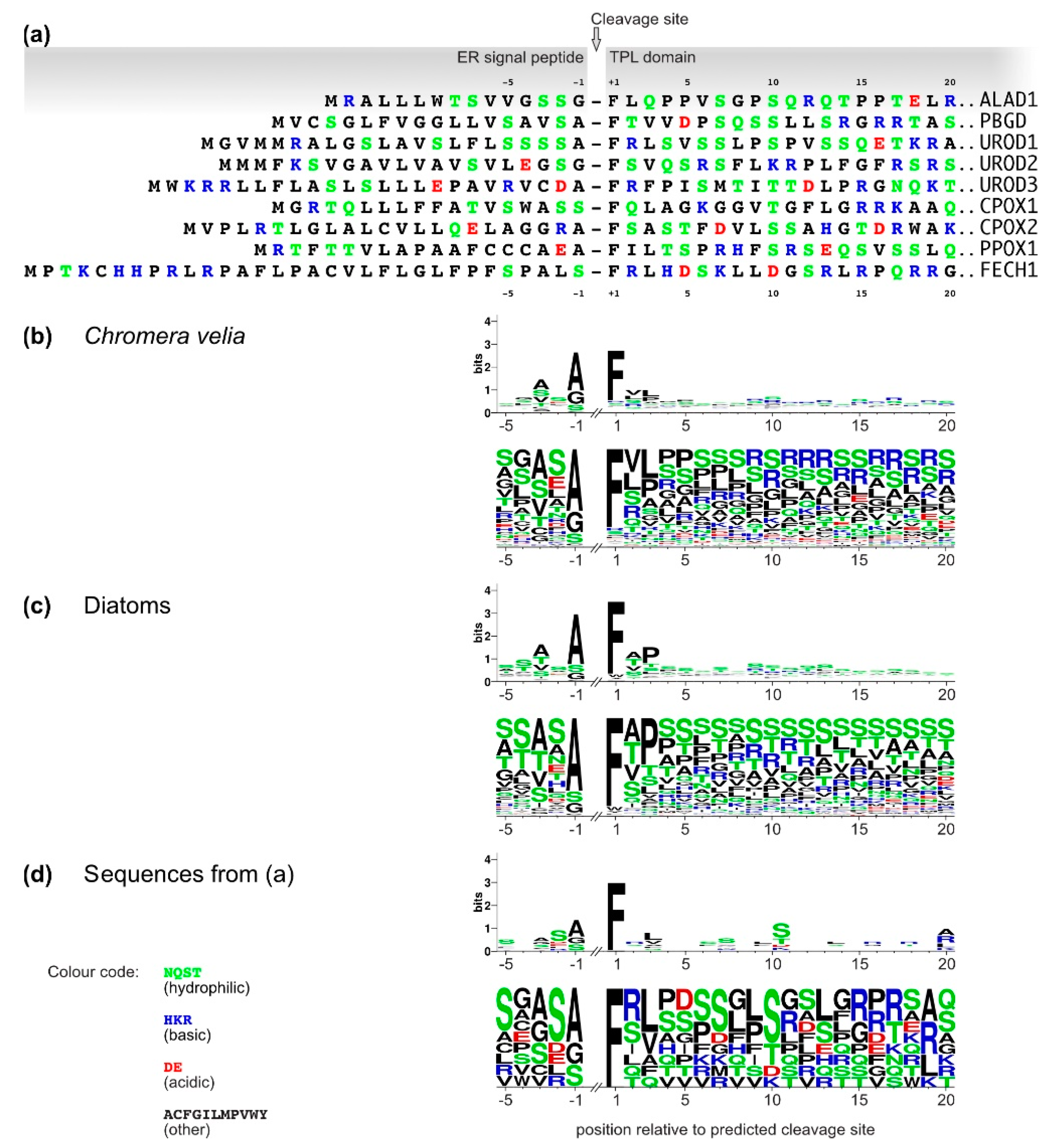
2.2. Analyses of C. velia Heme Pathway Enzymes N-termini Sequence
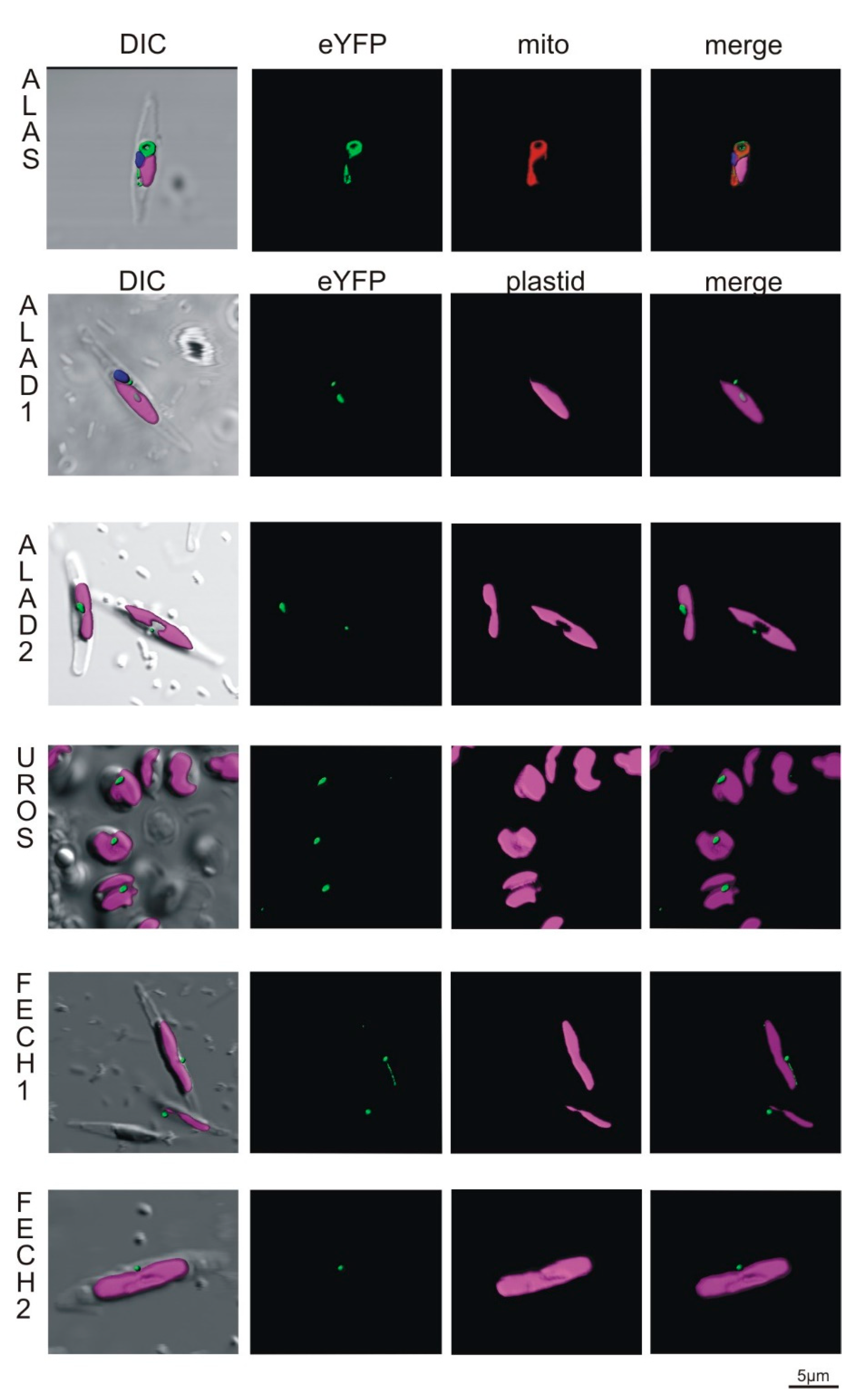
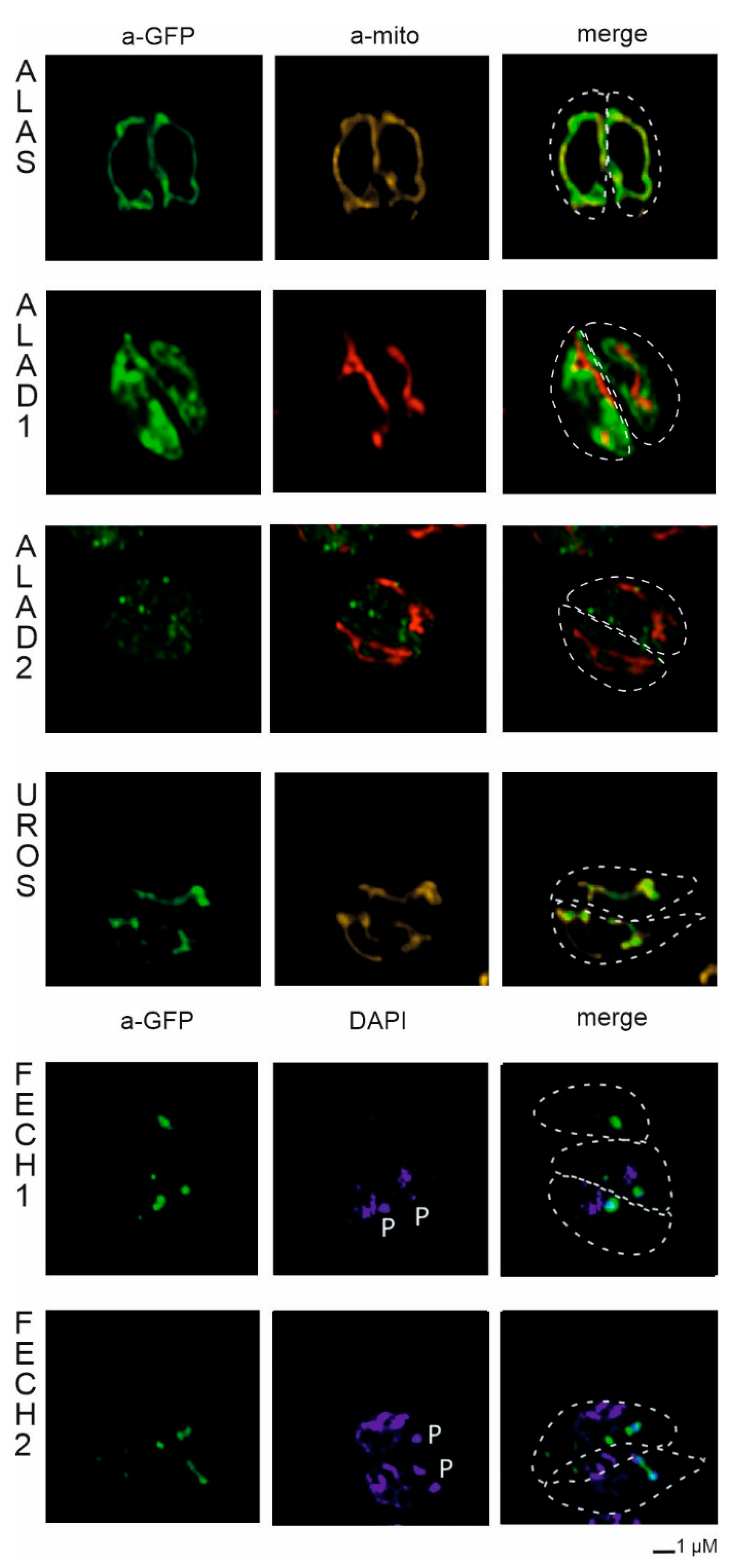
2.3. Localization of C. velia Heme Synthesis Enzymes via Heterologous Expression
2.3.1. Localization in Phaeodactylum tricornutum
2.3.2. Localization in Toxoplasma gondii
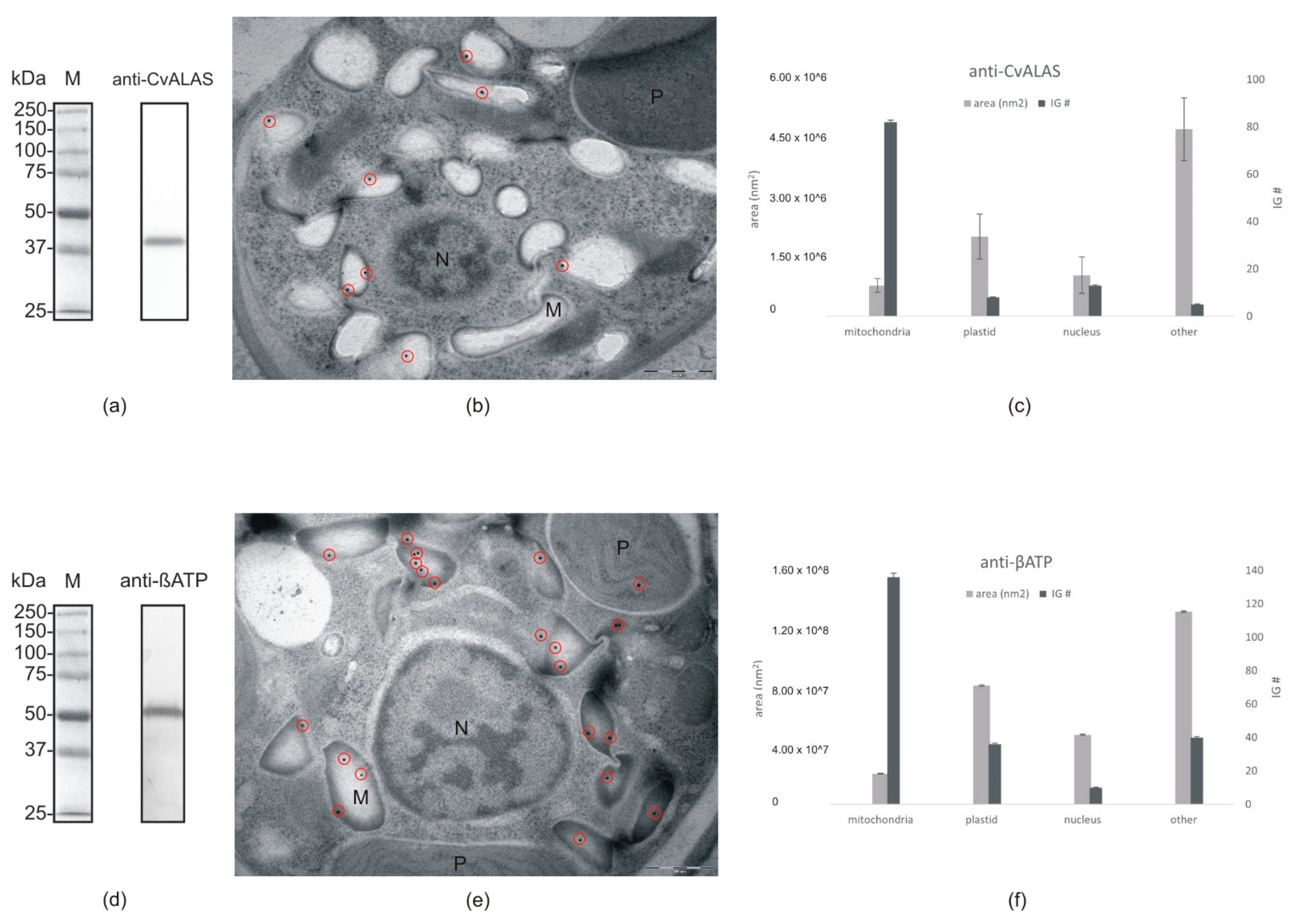
2.4. Direct ALAS Localization via Immune Gold Labeling of C. velia Cells
3. Discussion
4. Conclusions
5. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tanaka, R.; Tanaka, A. Tetrapyrrole biosynthesis in higher plants. Annu. Rev. Plant Biol. 2007, 58, 321–346. [Google Scholar] [CrossRef]
- Koreny, L.; Sobotka, R.; Kovarova, J.; Gnipova, A.; Flegontov, P.; Horvath, A.; Obornik, M.; Ayala, F.J.; Lukes, J. Aerobic kinetoplastid flagellate Phytomonas does not require heme for viability. Proc. Natl. Acad. Sci. USA 2012, 109, 3808–3813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, P.M. Highlights in haem biosynthesis. Curr. Opin. Struct. Biol. 1994, 4, 902–911. [Google Scholar] [CrossRef]
- Oborník, M.; Green, B.R. Mosaic origin of the heme biosynthesis pathway in photosynthetic eukaryotes. Mol. Biol. Evol. 2005, 22, 2343–2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kořený, L.; Oborník, M.; Lukeš, J. Make it, take it, or leave it: Heme metabolism of parasites. PLoS Pathog. 2013, 9, e1003088. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T.; Fujita, Y. Regulation and evolution of chlorophyll metabolism. Photochem. Photobiol. Sci. 2008, 7, 1131–1149. [Google Scholar] [CrossRef] [PubMed]
- Czarnecki, O.; Grimm, B. Post-translational control of tetrapyrrole biosynthesis in plants, algae, and cyanobacteria. J. Exp. Bot. 2012, 63, 1675–1687. [Google Scholar] [CrossRef] [Green Version]
- Ikushiro, H.; Nagami, A.; Takai, T.; Sawai, T.; Shimeno, Y.; Hori, H.; Miyahara, I.; Kamiya, N.; Yano, T. heme-dependent inactivation of 5-aminolevulinate synthase from Caulobacter crescentus. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Wißbrock, A.; George, A.A.P.; Brewitz, H.H.; Kühl, T.; Imhof, D. The molecular basis of transient heme-protein interactions: Analysis, concept and implementation. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- Kloehn, J.; Harding, C.R.; Soldati-Favre, D. Supply and demand—Heme synthesis, salvage and utilization by Apicomplexa. FEBS J. 2021, 288, 382–404. [Google Scholar] [CrossRef]
- Kořený, L.; Sobotka, R.; Janouškovec, J.; Keeling, P.J.; Oborník, M. Tetrapyrrole synthesis of photosynthetic chromerids is likely homologous to the unusual pathway of apicomplexan parasites. Plant Cell 2011, 23, 3454–3462. [Google Scholar] [CrossRef] [Green Version]
- Hamza, I. Intracellular trafficking of porphyrins. ACS Chem. Biol. 2006, 1, 627–629. [Google Scholar] [CrossRef] [Green Version]
- Swenson, S.A.; Moore, C.M.; Marcero, J.R.; Medlock, A.E.; Reddi, A.R.; Khalimonchuk, O. From synthesis to utilization: The ins and outs of mitochondrial heme. Cells 2020, 9, 579. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki, N.; Tanaka, R.; Grimm, B.; Masuda, T.; Moulin, M.; Smith, A.G.; Tanaka, A.; Terry, M.J. The cell biology of tetrapyrroles: A life and death struggle. Trends Plant Sci. 2010, 15, 488–498. [Google Scholar] [CrossRef]
- Kořený, L.; Oborník, M. Sequence evidence for the presence of two tetrapyrrole pathways in Euglena gracilis. Genome Biol. Evol. 2011, 3, 359–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cihlář, J.; Füssy, Z.; Horak, A.; Oborník, M. Evolution of the tetrapyrrole biosynthetic pathway in secondary algae: Conservation, redundancy and replacement. PLoS ONE 2016, 11, e0166338. [Google Scholar] [CrossRef] [PubMed]
- Votýpka, J.; Modrý, D.; Oborník, M.; Šlapeta, J.; Lukeš, J. Apicomplexa. In Handbook of the Protists; Archibald, J.M., Simpson, A.G.B., Slamovits, C.H., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 567–624. [Google Scholar] [CrossRef]
- Ralph, S.A.; Van Dooren, G.G.; Waller, R.; Crawford, M.J.; Fraunholz, M.; Foth, B.J.; Tonkin, C.J.; Roos, D.; McFadden, G.I. Metabolic maps and functions of the Plasmodium falciparum apicoplast. Nat. Rev. Microbiol. 2004, 2, 203–216. [Google Scholar] [CrossRef]
- Seeber, F.; Limenitakis, J.; Soldati-Favre, D. Apicomplexan mitochondrial metabolism: A story of gains, losses and retentions. Trends Parasitol. 2008, 24, 468–478. [Google Scholar] [CrossRef]
- Van Dooren, G.G.; Kennedy, A.T.; McFadden, G.I. The use and abuse of heme in apicomplexan parasites. Antioxid. Redox Signal. 2012, 17, 634–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tjhin, E.T.; Hayward, J.A.; McFadden, G.I.; van Dooren, G.G. Characterization of the apicoplast-localized enzyme TgUroD in Toxoplasma gondii reveals a key role of the apicoplast in heme biosynthesis. J. Biol. Chem. 2020, 295, 1539–1550. [Google Scholar] [CrossRef]
- Dalbey, R.; von Heijne, G. Protein Targeting, Transport, and Translocation, 1st ed.; Academic Press: Cambridge, MA, USA, 2002; 336p. [Google Scholar]
- Kroth, P.G. Protein transport into secondary plastids and the evolution of primary and secondary plastids. Int. Rev. Cytol. 2002, 221, 191–255. [Google Scholar] [CrossRef] [PubMed]
- Gould, S.B.; Waller, R.; McFadden, G.I. Plastid evolution. Annu. Rev. Plant Biol. 2008, 59, 491–517. [Google Scholar] [CrossRef] [Green Version]
- Bolte, K.; Bullmann, L.; Hempel, F.; Bozarth, A.; Zauner, S.; Maier, U.G. Protein targeting into secondary plastids. J. Eukaryot. Microbiol. 2009, 56, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Maier, U.G.; Zauner, S.; Hempel, F. Protein import into complex plastids: Cellular organization of higher complexity. Eur. J. Cell Biol. 2015, 94, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Cavalier-Smith, T. Kingdom Chromista and its eight phyla: A new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences. Protoplasma 2018, 255, 297–357. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.B.; Oborník, M.; Janouškovec, J.; Chrudimský, T.; Vancová, M.; Green, D.H.; Wright, S.W.; Davies, N.W.; Bolch, C.J.S.; Heimann, K.; et al. A photosynthetic alveolate closely related to apicomplexan parasites. Nature 2008, 451, 959–963. [Google Scholar] [CrossRef] [PubMed]
- Oborník, M.; Modrý, D.; Lukeš, M.; Černotíková-Stříbrná, E.; Cihlář, J.; Tesařová, M.; Kotabová, E.; Vancová, M.; Prasil, O.; Lukeš, J. Morphology, ultrastructure and life cycle of Vitrella brassicaformis n. sp., n. gen., a novel chromerid from the Great Barrier Reef. Protist 2012, 163, 306–323. [Google Scholar] [CrossRef]
- Janouškovec, J.; Tikhonenkov, D.; Burki, F.; Howe, A.T.; Kolísko, M.; Mylnikov, A.P.; Keeling, P.J. Factors mediating plastid dependency and the origins of parasitism in apicomplexans and their close relatives. Proc. Natl. Acad. Sci. USA 2015, 112, 10200–10207. [Google Scholar] [CrossRef] [Green Version]
- Oborník, M.; Kručinská, J.; Esson, H. Life cycles of chromerids resemble those of colpodellids and apicomplexan parasites. Perspect. Phycol. 2016, 3, 21–27. [Google Scholar] [CrossRef]
- Füssy, Z.; Masařová, P.; Kručinská, J.; Esson, H.J.; Oborník, M. Budding of the alveolate alga Vitrella brassicaformis resembles sexual and asexual processes in apicomplexan parasites. Protist 2017, 168, 80–91. [Google Scholar] [CrossRef]
- Oborník, M. Endosymbiotic evolution of algae, secondary heterotrophy and parasitism. Biomolecules 2019, 9, 266. [Google Scholar] [CrossRef] [Green Version]
- Oborník, M. Photoparasitism as an intermediate state in the evolution of apicomplexan parasites. Trends Parasitol. 2020, 36, 727–734. [Google Scholar] [CrossRef]
- Kilian, O.; Kroth, P.G. Identification and characterization of a new conserved motif within the presequence of proteins targeted into complex diatom plastids. Plant J. 2005, 41, 175–183. [Google Scholar] [CrossRef]
- Patron, N.J.; Waller, R. Transit peptide diversity and divergence: A global analysis of plastid targeting signals. BioEssays 2007, 29, 1048–1058. [Google Scholar] [CrossRef] [PubMed]
- Apt, K.E.; Grossman, A.R.; Kroth-Pancic, P.G. Stable nuclear transformation of the diatom Phaeodactylum tricornutum. Mol. Gen. Genet. MGG 1996, 252, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Striepen, B.; He, C.Y.; Matrajt, M.; Soldati-Favre, D.; Roos, D. Expression, selection, and organellar targeting of the green fluorescent protein in Toxoplasma gondii. Mol. Biochem. Parasitol. 1998, 92, 325–338. [Google Scholar] [CrossRef]
- Poulsen, N.; Chesley, P.M.; Kröger, N. MOLECULAR genetic manipulation of the diatom Thalassiosira pseudonana (bacillariophyceae). J. Phycol. 2006, 42, 1059–1065. [Google Scholar] [CrossRef]
- Striepen, B.; Soldati, D. Genetic manipulation of Toxoplasma gondii. In Toxoplasma Gondii; Academic Press: Cambridge, MA, USA, 2007; pp. 391–418. [Google Scholar] [CrossRef]
- Zhang, C.; Hu, H. High-efficiency nuclear transformation of the diatom Phaeodactylum tricornutum by electroporation. Mar. Genom. 2014, 16, 63–66. [Google Scholar] [CrossRef]
- McFadden, G.I. Plastids and protein targeting. J. Eukaryot. Microbiol. 1999, 46, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Roos, D.S.; Crawford, M.J.; Donald, R.G.; Kissinger, J.; Klimczak, L.J.; Striepen, B. Origin, targeting, and function of the apicomplexan plastid. Curr. Opin. Microbiol. 1999, 2, 426–432. [Google Scholar] [CrossRef]
- DeRocher, A.; Hagen, C.B.; Froehlich, J.E.; Feagin, J.E.; Parsons, M. Analysis of targeting sequences demonstrates that trafficking to the Toxoplasma gondii plastid branches off the secretory system. J. Cell Sci. 2000, 113, 3969–3977. [Google Scholar] [CrossRef]
- Waller, R.F.; Reed, M.B.; Cowman, A.F.; McFadden, G.I. Protein trafficking to the plastid of Plasmodium falciparum is via the secretory pathway. EMBO J. 2000, 19, 1794–1802. [Google Scholar] [CrossRef] [Green Version]
- Apt, K.E.; Zaslavkaia, L.; Lippmeier, J.C.; Lang, M.; Kilian, O.; Wetherbee, R.; Grossman, A.R.; Kroth, P.G. In vivo characterization of diatom multipartite plastid targeting signals. J. Cell Sci. 2002, 115, 4061–4069. [Google Scholar] [CrossRef] [Green Version]
- Sheiner, L.; Demerly, J.L.; Poulsen, N.; Beatty, W.L.; Lucas, O.; Behnke, M.; White, M.W.; Striepen, B. A systematic screen to discover and analyze apicoplast proteins identifies a conserved and essential protein import factor. PLOS Pathog. 2011, 7, e1002392. [Google Scholar] [CrossRef] [Green Version]
- Huesgen, P.F.; Alami, M.; Lange, P.F.; Foster, L.J.; Schröder, W.P.; Overall, C.M.; Green, B.R. Proteomic amino-termini profiling reveals targeting information for protein import into complex plastids. PLoS ONE 2013, 8, e74483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Füssy, Z.; Oborník, M. Chromerids and their plastids. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2017; Volume 84, pp. 187–218. [Google Scholar] [CrossRef]
- Oborník, M.; Lukeš, J. Cell biology of chromerids. In International Review of Cell and Molecular Biology; Academic Press: Cambridge, MA, USA, 2013; Volume 306, pp. 333–369. [Google Scholar] [CrossRef]
- Nielsen, H. Predicting secretory proteins with SignalP. In Protein Function Prediction; Kihara, D., Ed.; Humana Press: New York, NY, USA, 2017; pp. 59–73. [Google Scholar] [CrossRef] [Green Version]
- Emanuelsson, O.; Brunak, S.; Von Heijne, G.; Nielsen, H.A. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef] [PubMed]
- Gruber, A.; Rocap, G.; Kroth, P.G.; Armbrust, E.V.; Mock, T. Plastid proteome prediction for diatoms and other algae with secondary plastids of the red lineage. Plant J. 2015, 81, 519–528. [Google Scholar] [CrossRef] [Green Version]
- Füssy, Z.; Faitová, T.; Oborník, M. subcellular compartments interplay for carbon and nitrogen allocation in Chromera velia and Vitrella brassicaformis. Genome Biol. Evol. 2019, 11, 1765–1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukasawa, Y.; Tsuji, J.; Fu, S.-C.; Tomii, K.; Horton, P.; Imai, K. MitoFates: Improved prediction of mitochondrial targeting sequences and their cleavage sites. Mol. Cell. Proteom. 2015, 14, 1113–1126. [Google Scholar] [CrossRef] [Green Version]
- Gruber, A.; McKay, C.; Kroth, P.G.; Armbrust, E.V.; Mock, T. Comparison of different versions of SignalP and TargetP for diatom plastid protein predictions with ASAFind. Matters 2020, 81, 519–528. [Google Scholar] [CrossRef] [Green Version]
- Bártulos, C.R.; Rogers, M.B.; Williams, T.; Gentekaki, E.; Brinkmann, H.; Cerff, R.; Liaud, M.-F.; Hehl, A.; Yarlett, N.R.; Gruber, A.; et al. Mitochondrial glycolysis in a major lineage of eukaryotes. Genome Biol. Evol. 2018, 10, 2310–2325. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; De Martino, A.; Amato, A.; Montsant, A.; Mathieu, B.; Rostaing, P.; Tirichine, L.; Bowler, C. ultrastructure and membrane traffic during cell division in the marine pennate diatom Phaeodactylum tricornutum. Protist 2015, 166, 506–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, S.B.; Sommer, M.S.; Hadfi, K.; Zauner, S.; Kroth, P.G.; Maier, U.G. Protein targeting into the complex plastid of cryptophytes. J. Mol. Evol. 2006, 62, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Gould, S.B.; Sommer, M.S.; Kroth, P.G.; Gile, G.H.; Keeling, P.J.; Maier, U.G. Nucleus-to-nucleus gene transfer and protein retargeting into a remnant cytoplasm of cryptophytes and diatoms. Mol. Biol. Evol. 2006, 23, 2413–2422. [Google Scholar] [CrossRef] [PubMed]
- Ovciarikova, J.; Lemgruber, L.; Stilger, K.L.; Sullivan, W.J.; Sheiner, L. Mitochondrial behaviour throughout the lytic cycle of Toxoplasma gondii. Sci. Rep. 2017, 7, 42746. [Google Scholar] [CrossRef] [Green Version]
- Šubrtová, K.; Panicucci, B.; Zíková, A. ATPaseTb2, a unique membrane-bound FoF1-ATPase component, is essential in bloodstream and dyskinetoplastic trypanosomes. PLOS Pathog. 2015, 11, e1004660. [Google Scholar] [CrossRef]
- Brzezowski, P.; Richter, A.S.; Grimm, B. Regulation and function of tetrapyrrole biosynthesis in plants and algae. Biochim. Biophys. Acta Bioenerg. 2015, 1847, 968–985. [Google Scholar] [CrossRef] [Green Version]
- Cihlář, J.; Füssy, Z.; Oborník, M. Evolution of tetrapyrrole pathway in eukaryotic phototrophs. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2019; Volume 90, pp. 273–309. [Google Scholar] [CrossRef]
- Zaslavskaia, L.A.; Lippmeier, J.C.; Grossman, A.R.; Kroth, P.G.; Apt, K.E. Transformation of the diatom Phaeodactylum tricornutum (Bacillariophyceae) with a variety of selectable marker and reporter genes. J. Phycol. 2000, 36, 379–386. [Google Scholar] [CrossRef]
- Niu, Y.-F.; Yang, Z.; Zhang, M.-H.; Zhu, C.-C.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. Transformation of diatom Phaeodactylum tricornutum by electroporation and establishment of inducible selection marker. Biotechniques 2012, 52. [Google Scholar] [CrossRef]
- Weiss, L.M.; Kim, K. Toxoplasma Gondii: The Model Apicomplexan—Perspectives and Methods, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2013; 1085p. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Kleczkowski, L.A. Optimization of ATP synthase function in mitochondria and chloroplasts via the adenylate kinase equilibrium. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Warren, M.J.; Smith, A.G.; Schubert, H.L.; Erskine, P.T.; Cooper, J.B. 5-Aminolaevulinic acid dehydratase, porphobilinogen deaminase and uroporphyrinogen III synthase. In Tetrapyrroles; Springer: New York, NY, USA, 2009; pp. 43–73. [Google Scholar] [CrossRef]
- Woo, Y.H.; Ansari, H.; Otto, T.; Klinger, C.M.; Kolisko, M.; Michálek, J.; Saxena, A.; Shanmugam, D.; Tayyrov, A.; Veluchamy, A.; et al. Chromerid genomes reveal the evolutionary path from photosynthetic algae to obligate intracellular parasites. eLife 2015, 4, e06974. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, M.; Medlock, A.; Dailey, H. Porphyrin metabolism. In Encyclopedia of Biological Chemistry, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 544–549. [Google Scholar] [CrossRef]
- Bhagavan, N.; Ha, C.-E. Metabolism of iron and heme. In Essentials of Medical Biochemistry; Academic Press: Cambridge, MA, USA, 2015; pp. 511–529. [Google Scholar] [CrossRef]
- Masoumi, A.; Heinemann, I.U.; Rohde, M.; Koch, M.; Jahn, M.; Jahn, D. Complex formation between protoporphyrinogen IX oxidase and ferrochelatase during haem biosynthesis in Thermosynechococcus elongatus. Microbiology 2009, 154, 3707–3714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbst, J.; Hey, D.; Grimm, B. Posttranslational control of tetrapyrrole biosynthesis: Interacting proteins, chaperones, auxiliary factors. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2019; pp. 163–194. [Google Scholar] [CrossRef]
- Gonzalez, N.H.; Felsner, G.; Schramm, F.D.; Klingl, A.; Maier, U.-G.; Bolte, K. A single peroxisomal targeting signal mediates matrix protein import in diatoms. PLoS ONE 2011, 6, e25316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stork, S.; Lau, J.; Moog, D.; Maier, U.-G. Three old and one new: Protein import into red algal-derived plastids surrounded by four membranes. Protoplasma 2013, 250, 1013–1023. [Google Scholar] [CrossRef]
- Gruber, A.; Vugrinec, S.; Hempel, F.; Gould, S.B.; Maier, U.-G.; Kroth, P.G. Protein targeting into complex diatom plastids: Functional characterisation of a specific targeting motif. Plant Mol. Biol. 2007, 64, 519–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsner, G.; Sommer, M.S.; Maier, U.G. The physical and functional borders of transit peptide-like sequences in secondary endosymbionts. BMC Plant Biol. 2010, 10, 223. [Google Scholar] [CrossRef] [Green Version]
- Blume, M.; Nitzsche, R.; Sternberg, U.; Gerlic, M.; Masters, S.L.; Gupta, N.; McConville, M.J. A Toxoplasma gondii gluconeogenic enzyme contributes to robust central carbon metabolism and is essential for replication and virulence. Cell Host Microbe 2015, 18, 210–220. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.M.; Long, S.; Sibley, L.D. Plasma membrane association by N-Acylation governs PKG function in Toxoplasma gondii. mBio 2017, 8, e00375-17. [Google Scholar] [CrossRef] [Green Version]
- Sheiner, L.; Striepen, B. Protein sorting in complex plastids. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 352–359. [Google Scholar] [CrossRef] [Green Version]
- Boucher, M.J.; Yeh, E. Plastid–endomembrane connections in apicomplexan parasites. PLoS Pathog. 2019, 15, e1007661. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S.; van Dooren, G.G.; Beatty, W.L.; Striepen, B. Genetic evidence that an endosymbiont-derived endoplasmic reticulum-associated protein degradation (ERAD) system functions in import of apicoplast proteins. J. Biol. Chem. 2009, 284, 33683–33691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hempel, F.; Bullmann, L.; Lau, J.; Zauner, S.; Maier, U.G. ERAD-derived preprotein transport across the second outermost plastid membrane of diatoms. Mol. Biol. Evol. 2009, 26, 1781–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hempel, F.; Bolte, K.; Klingl, A.; Zauner, S.; Maier, U.-G. Protein transport into plastids of secondarily evolved organisms. In Plastid Biology; Springer: New York, NY, USA, 2014; pp. 291–303. [Google Scholar]
- Bullmann, L.; Haarmann, R.; Mirus, O.; Bredemeier, R.; Hempel, F.; Maier, U.G.; Schleiff, E. Filling the gap, evolutionarily conserved Omp85 in plastids of chromalveolates. J. Biol. Chem. 2010, 285, 6848–6856. [Google Scholar] [CrossRef] [Green Version]
- Glaser, S.; van Dooren, G.G.; Agrawal, S.; Brooks, C.F.; McFadden, G.I.; Striepen, B.; Higgins, M.K. Tic22 is an essential chaperone required for protein import into the apicoplast. J. Biol. Chem. 2012, 287, 39505–39512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheiner, L.; Fellows, J.D.; Ovciarikova, J.; Brooks, C.F.; Agrawal, S.; Holmes, Z.C.; Bietz, I.; Flinner, N.; Heiny, S.; Mirus, O.; et al. Toxoplasma gondii Toc75 functions in import of stromal but not peripheral apicoplast proteins. Traffic 2015, 16, 1254–1269. [Google Scholar] [CrossRef] [Green Version]
- Roger, A.J.; Muñoz-Gómez, S.A.; Kamikawa, R. The origin and diversification of mitochondria. Curr. Biol. 2017, 27, R1177–R1192. [Google Scholar] [CrossRef] [Green Version]
- Wiedemann, N.; Pfanner, N. Mitochondrial machineries for protein import and assembly. Annu. Rev. Biochem. 2017, 86, 685–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maréchal, E.; Cesbron-Delauw, M.-F. The apicoplast: A new member of the plastid family. Trends Plant. Sci. 2001, 6, 200–205. [Google Scholar] [CrossRef]
- Smith, A.; Santana, M.; Wallace-Cook, A.; Roper, J.; Labbe-Bois, R. Isolation of a cDNA encoding chloroplast ferrochelatase from Arabidopsis thaliana by functional complementation of a yeast mutant. J. Biol. Chem. 1994, 269, 13405–13413. [Google Scholar] [CrossRef]
- Tanaka, R.; Kobayashi, K.; Masuda, T. Tetrapyrrole metabolism in Arabidopsis thaliana. Arab. Book 2011, 9, e0145. [Google Scholar] [CrossRef] [Green Version]
- Nagai, S.; Koide, M.; Takahashi, S.; Kikuta, A.; Aono, M.; Sasaki-Sekimoto, Y.; Ohta, H.; Takamiya, K.-I.; Masuda, T. Induction of isoforms of tetrapyrrole biosynthetic enzymes, AtHEMA2 and AtFC1, under stress conditions and their physiological functions in arabidopsis. Plant Physiol. 2007, 144, 1039–1051. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Masuda, T. Transcriptional regulation of tetrapyrrole biosynthesis in Arabidopsis thaliana. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Sobotka, R.; Tichy, M.; Wilde, A.; Hunter, C.N. Functional assignments for the carboxyl-terminal domains of the ferrochelatase from synechocystis PCC 6803: The CAB domain plays a regulatory role, and region II is essential for catalysis. Plant Physiol. 2011, 155, 1735–1747. [Google Scholar] [CrossRef] [Green Version]
- Pazderník, M.; Mareš, J.; Pilný, J.; Sobotka, R. The antenna-like domain of the cyanobacterial ferrochelatase can bind chlorophyll and carotenoids in an energy-dissipative configuration. J. Biol. Chem. 2019, 294, 11131–11143. [Google Scholar] [CrossRef] [PubMed]
- Franken, A.C.W.; Lokman, B.C.; Ram, A.F.J.; Punt, P.J.; van den Hondel, C.A.M.J.J.; de Weert, S. Heme biosynthesis and its regulation: Towards understanding and improvement of heme biosynthesis in filamentous fungi. Appl. Microbiol. Biotechnol. 2011, 91, 447–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Kang, Z.; Chen, J.; Du, G. Optimization of the heme biosynthesis pathway for the production of 5-aminolevulinic acid in Escherichia coli. Sci. Rep. 2015, 5, 8584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medlock, A.E.; Shiferaw, M.T.; Marcero, J.R.; Vashisht, A.A.; Wohlschlegel, J.A.; Phillips, J.D.; Dailey, H.A. Identification of the mitochondrial heme metabolism complex. PLoS ONE 2015, 10, e0135896. [Google Scholar] [CrossRef] [Green Version]
- Petersen, T.N.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- Schneider, T.D.; Stephens, R.M. Sequence logos: A new way to display consensus sequences. Nucleic Acids Res. 1990, 18, 6097–6100. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roos, D.; Donald, R.G.K.; Morrissette, N.S.; Moulton, A.L.C. Molecular tools for genetic dissection of the protozoan parasite Toxoplasma gondii. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 1995; Volume 45, pp. 27–63. [Google Scholar] [CrossRef]
- Donald, R.G.; Roos, D. Stable molecular transformation of Toxoplasma gondii: A selectable dihydrofolate reductase-thymidylate synthase marker based on drug-resistance mutations in malaria. Proc. Natl. Acad. Sci. USA 1993, 90, 11703–11707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacot, D.; Meissner, M.; Sheiner, L.; Soldati-Favre, D.; Striepen, B. Genetic manipulation of Toxoplasma gondii. In Toxoplasma Gondii, The Model Apicomplexan—Perspectives and Methods, 2nd ed.; Elevier: Amsterdam, The Netherlands, 2014; pp. 577–611. [Google Scholar] [CrossRef]
- Lucocq, J.M.; Habermann, A.; Watt, S.; Backer, J.M.; Mayhew, T.M.; Griffiths, G. A rapid method for assessing the distribution of gold labeling on thin sections. J. Histochem. Cytochem. 2004, 52, 991–1000. [Google Scholar] [CrossRef]
- Lucocq, J.M.; Gawden-Bone, C. Quantitative assessment of specificity in immunoelectron microscopy. J. Histochem. Cytochem. 2010, 58, 917–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Accession (CryptoDB) | Evolutionary Origin | Targeting Prediction | Localization T. gondii | Localization P. tricornutum | Conclusion |
|---|---|---|---|---|---|---|
| ALAS | Cvel_28814.t1 | Alphaproteobacteria | No targeting signal identified | Mitochondria | Mitochondria | Mitochondria |
| ALAD1 | Cvel_108.t1 | Cyanobacteria | Plastid | Cytosol | PPC/ER | Plastid |
| ALAD2 | Cvel_13826.t1 | Primary alga | No targeting signal identified | Cytosol | PPC/ER | Uncertain location |
| ALAD3 | Cvel_36189.t1 | Proteobacterial | Mitochondria | Not tested | Not tested | Uncertain location |
| PBGD | Cvel_26028.t1 | Alphaproteobacteria | Plastid | Not tested | Not tested | Plastid |
| UROS | Cvel_15018.t1 | Uncertain origin in primary alga | No targeting signal identified | Mitochondria | PPC/ER | Uncertain location |
| UROD1 | Cvel_14720.t1 | Cyanobacteria | Plastid | Not tested | Not tested | Plastid |
| UROD2 | Cvel_5098.t1 | Endosymbiont nucleus | Plastid | Not tested | Not tested | Plastid |
| UROD3 | Cvel_31936.t1 | Secondary host nucleus | Plastid | Not tested | Not tested | Plastid |
| CPOX1 | Cvel_21486.t1 | Secondary host nucleus | Plastid | Not tested | Not tested | Plastid |
| CPOX2 | Cvel_2641.t1 | Uncertain origin in primary alga | Plastid | Not tested | Not tested | Plastid |
| PPOX1 | Cvel_13840.t1 | Cyanobacteria | Plastid | Not tested | Not tested | Plastid |
| PPOX2 | Cvel_18037.t1 | Eukaryotic origin | Mitochondria | Not tested | Not tested | Uncertain location |
| FECH1 | Cvel_18167.t1 | Cyanobacteria | Plastid | Apicoplast | PPC/ER | Plastid |
| FECH2 | Cvel_26873.t1 | Alphaproteobacteria | Signal peptide positive | Apicoplast | PPC/ER | Uncertain location |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richtová, J.; Sheiner, L.; Gruber, A.; Yang, S.-M.; Kořený, L.; Striepen, B.; Oborník, M. Using Diatom and Apicomplexan Models to Study the Heme Pathway of Chromera velia. Int. J. Mol. Sci. 2021, 22, 6495. https://doi.org/10.3390/ijms22126495
Richtová J, Sheiner L, Gruber A, Yang S-M, Kořený L, Striepen B, Oborník M. Using Diatom and Apicomplexan Models to Study the Heme Pathway of Chromera velia. International Journal of Molecular Sciences. 2021; 22(12):6495. https://doi.org/10.3390/ijms22126495
Chicago/Turabian StyleRichtová, Jitka, Lilach Sheiner, Ansgar Gruber, Shun-Min Yang, Luděk Kořený, Boris Striepen, and Miroslav Oborník. 2021. "Using Diatom and Apicomplexan Models to Study the Heme Pathway of Chromera velia" International Journal of Molecular Sciences 22, no. 12: 6495. https://doi.org/10.3390/ijms22126495