
The AtCRK5 Protein Kinase Is Required to Maintain the ROS NO Balance Affecting the PIN2-Mediated Root Gravitropic Response in Arabidopsis

 , , , , and
, , , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
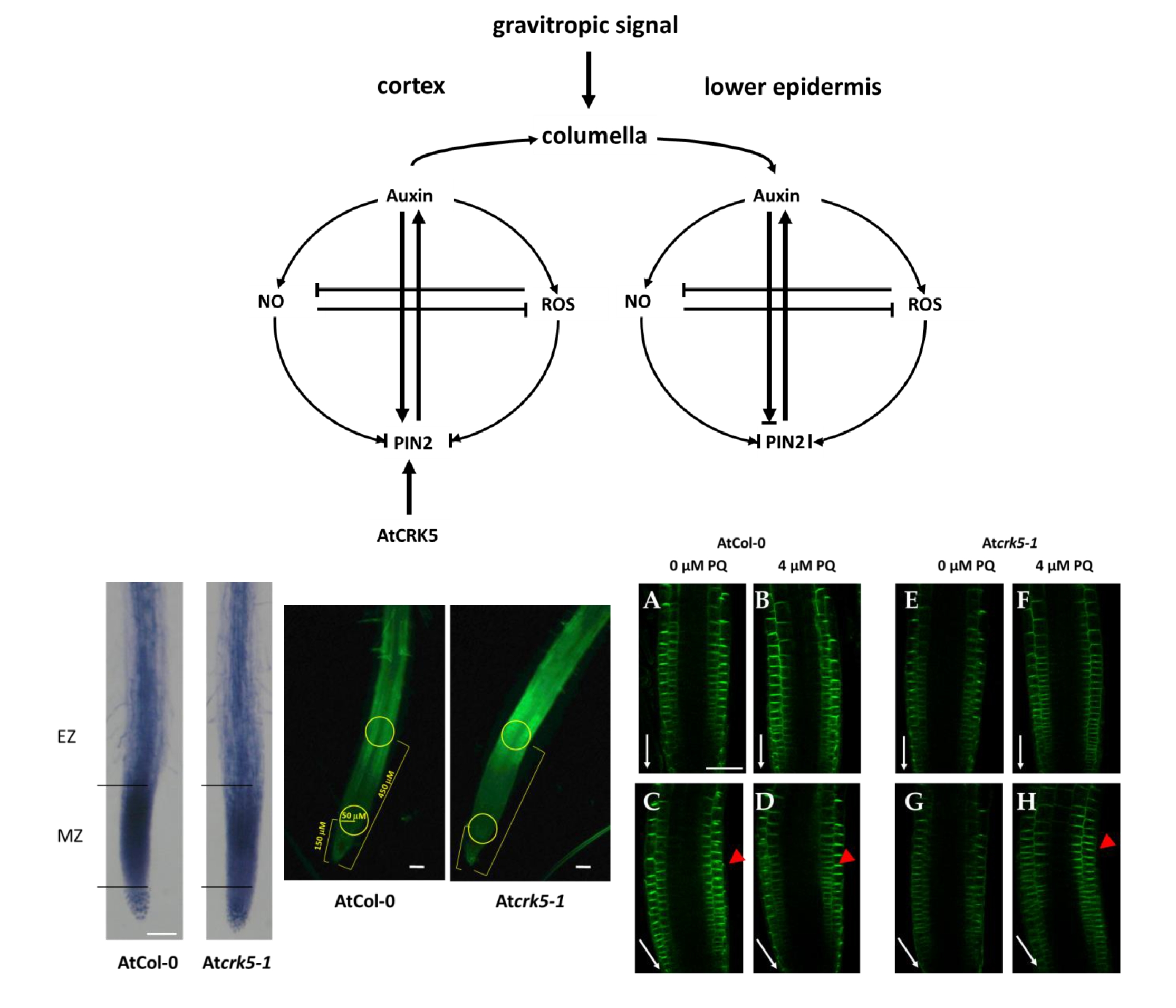
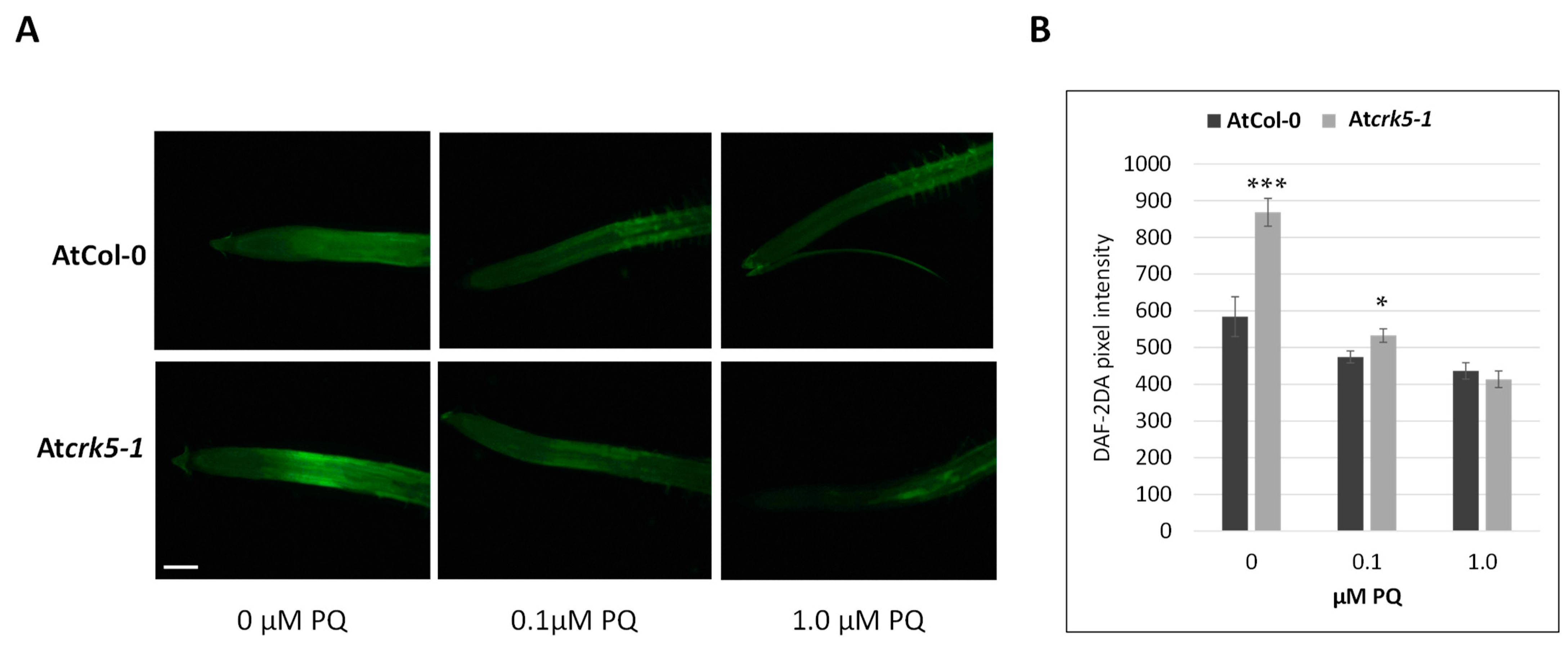
2.1. The Seedlings of the Atcrk5-1 Mutant Arabidopsis Have O2•− and H2O2 Deficiencies in Root Tips
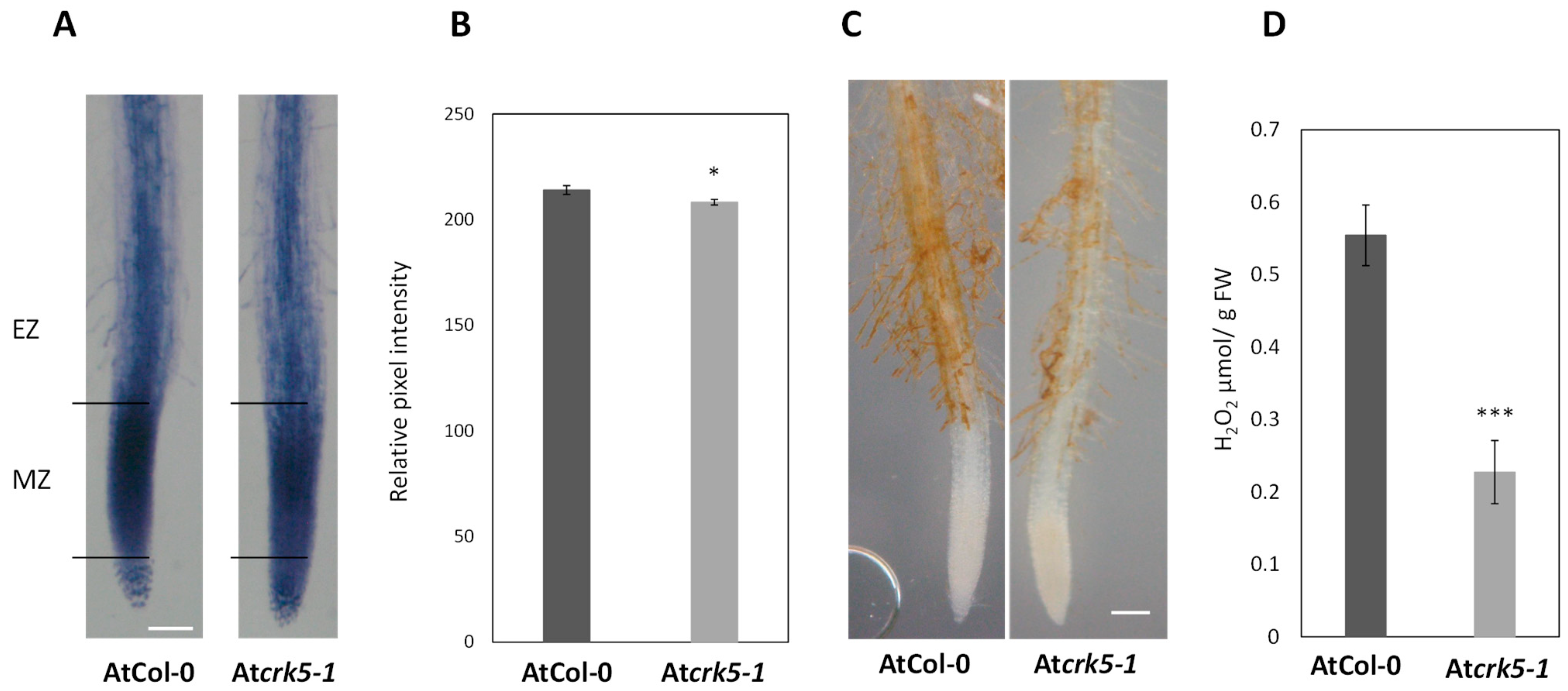
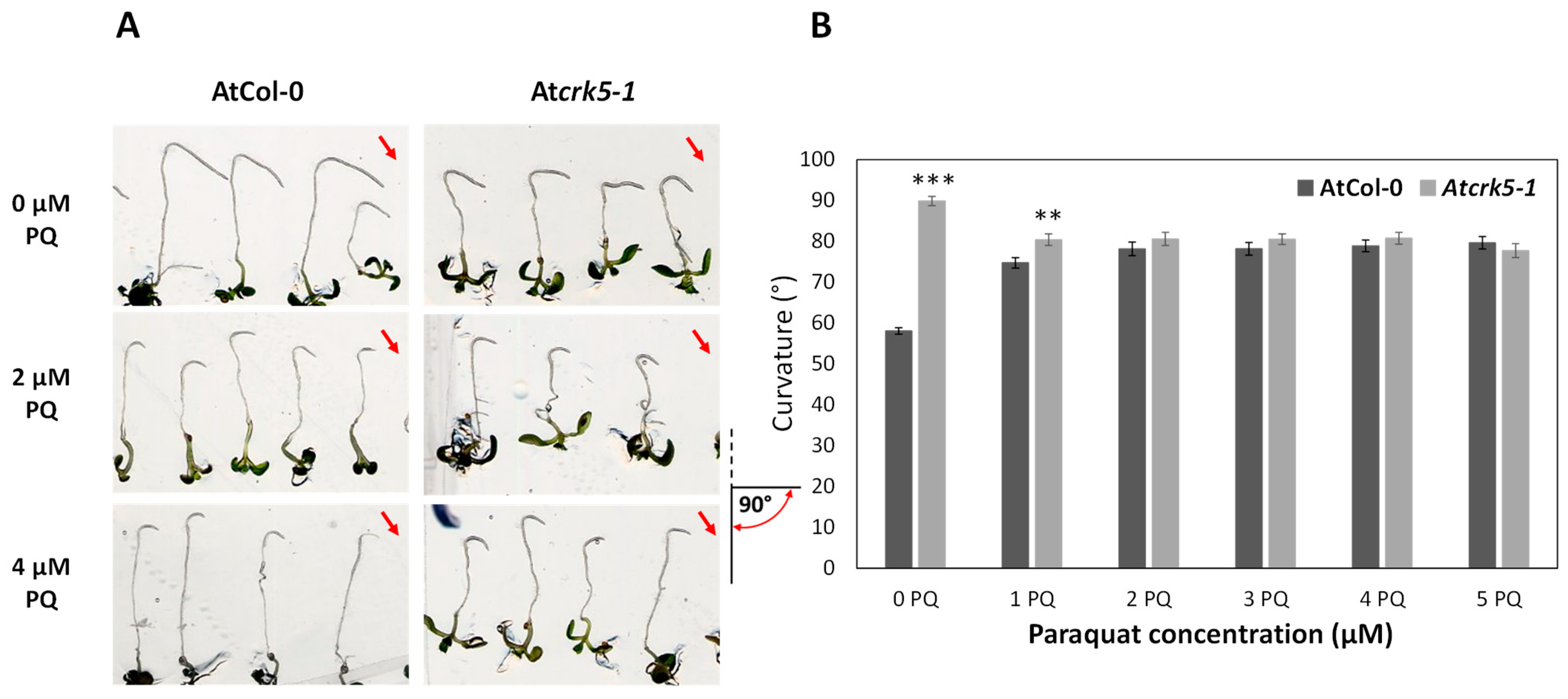
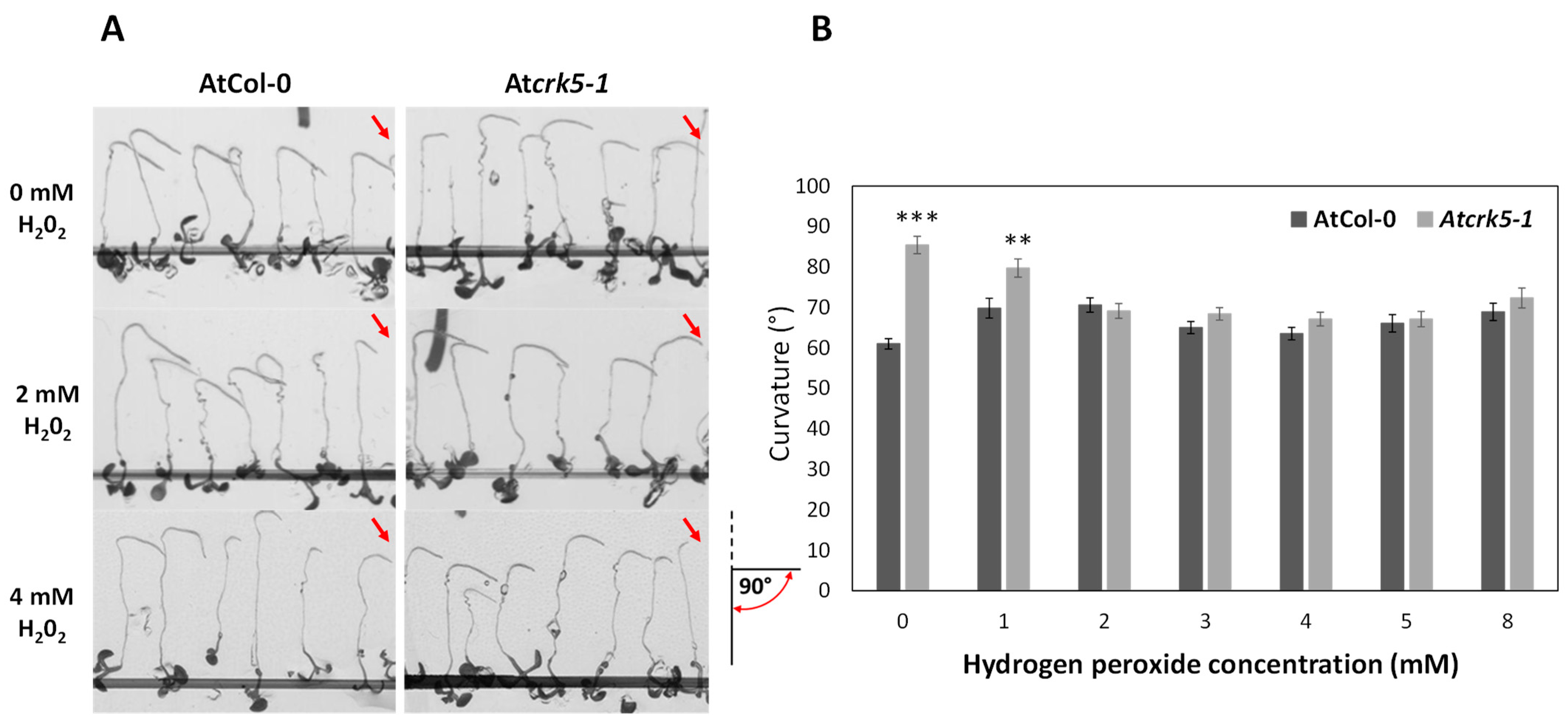
2.2. Paraquat and H2O2 Treatments Restore the Gravitropic Response of Atcrk5-1 Roots
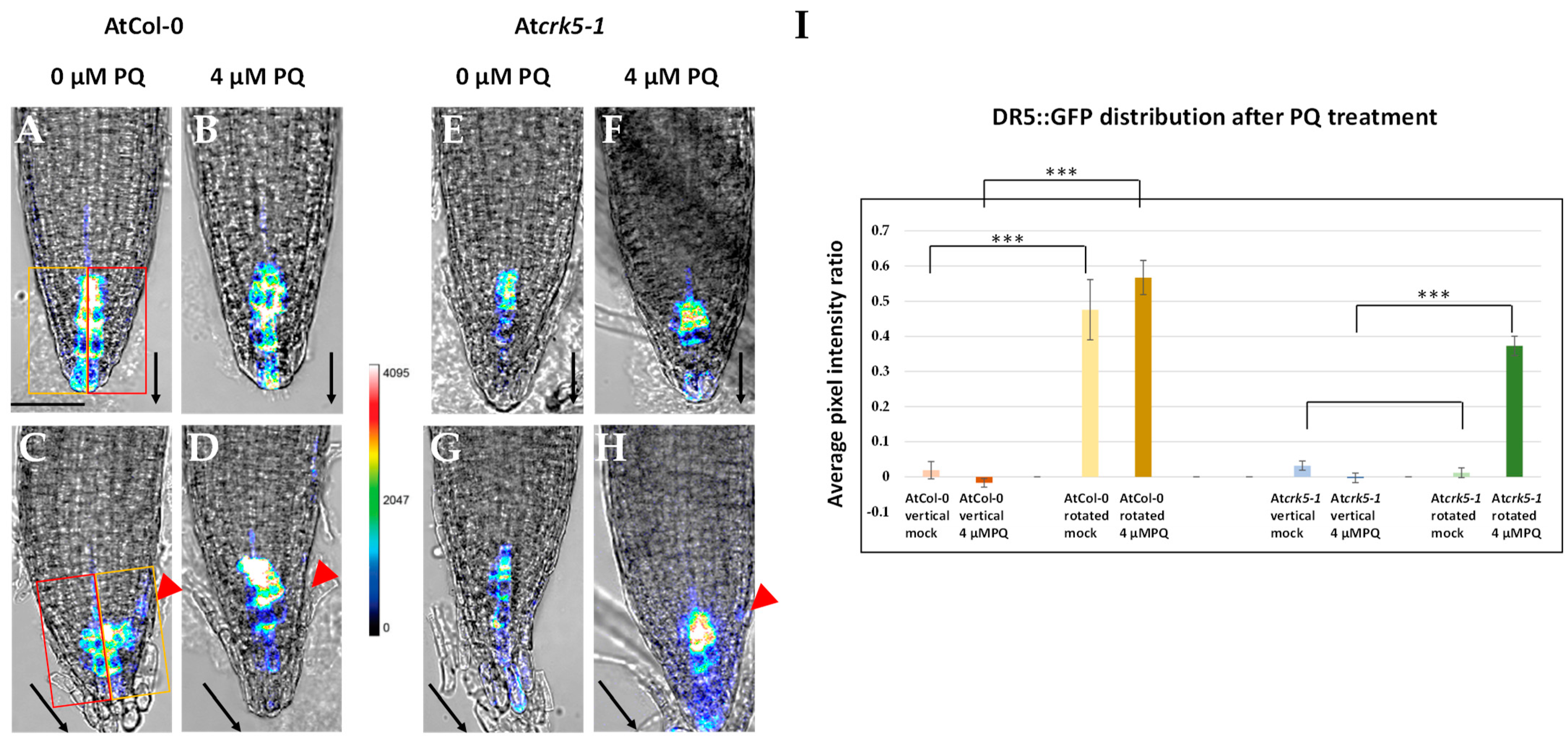
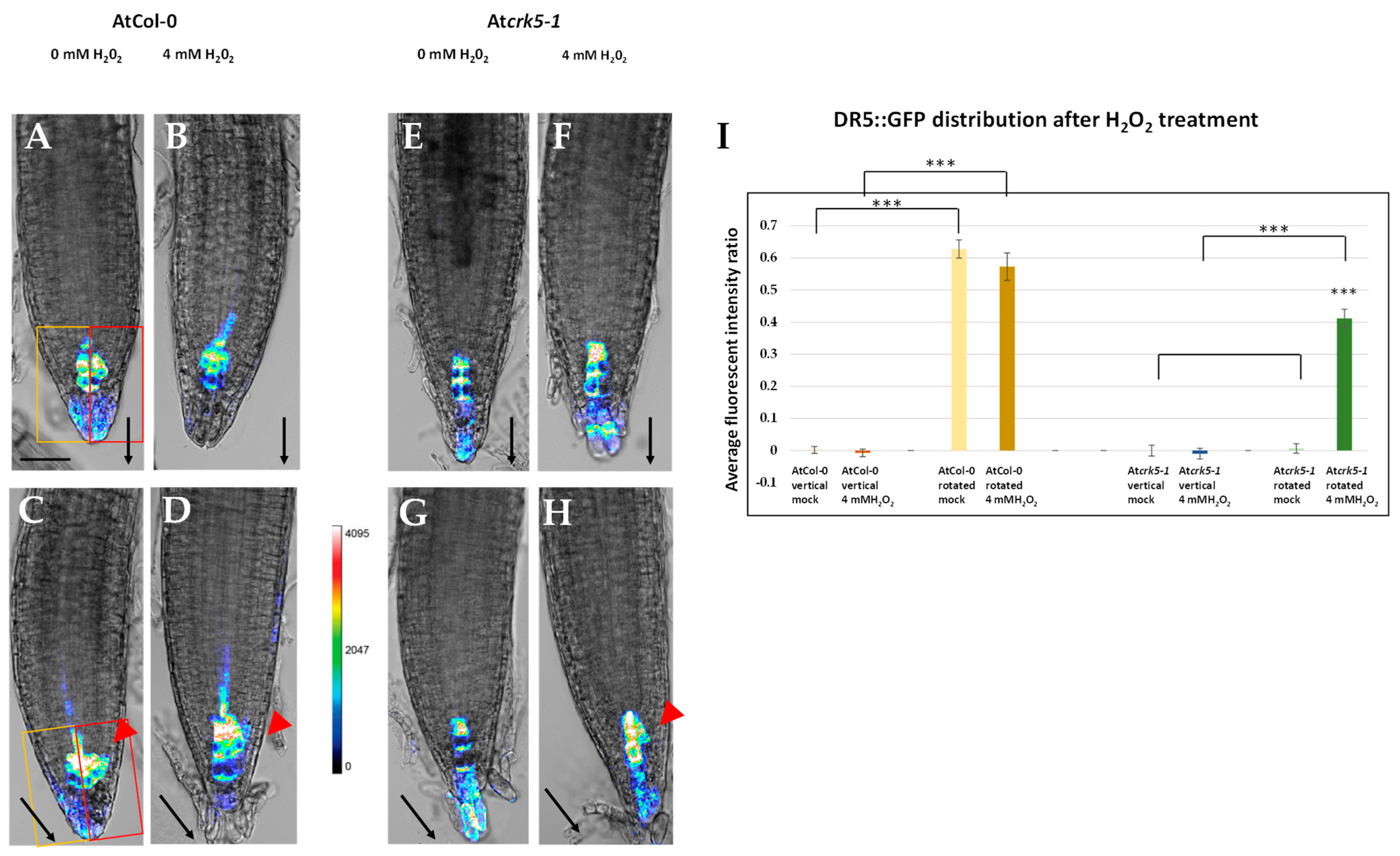
2.3. PQ or H2O2 Treatments Restore the Auxin Distribution in the Root Meristem of the Atcrk5-1 Mutant during Gravistimulation
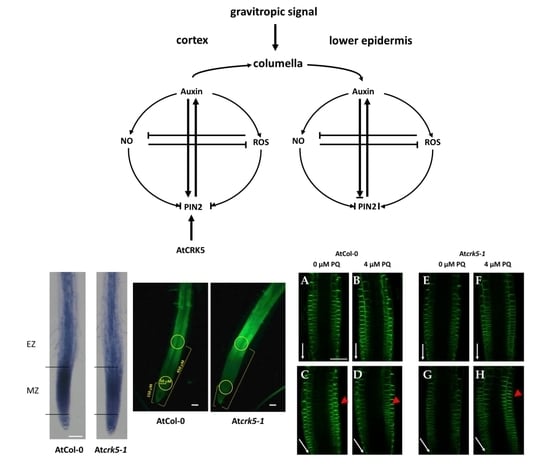
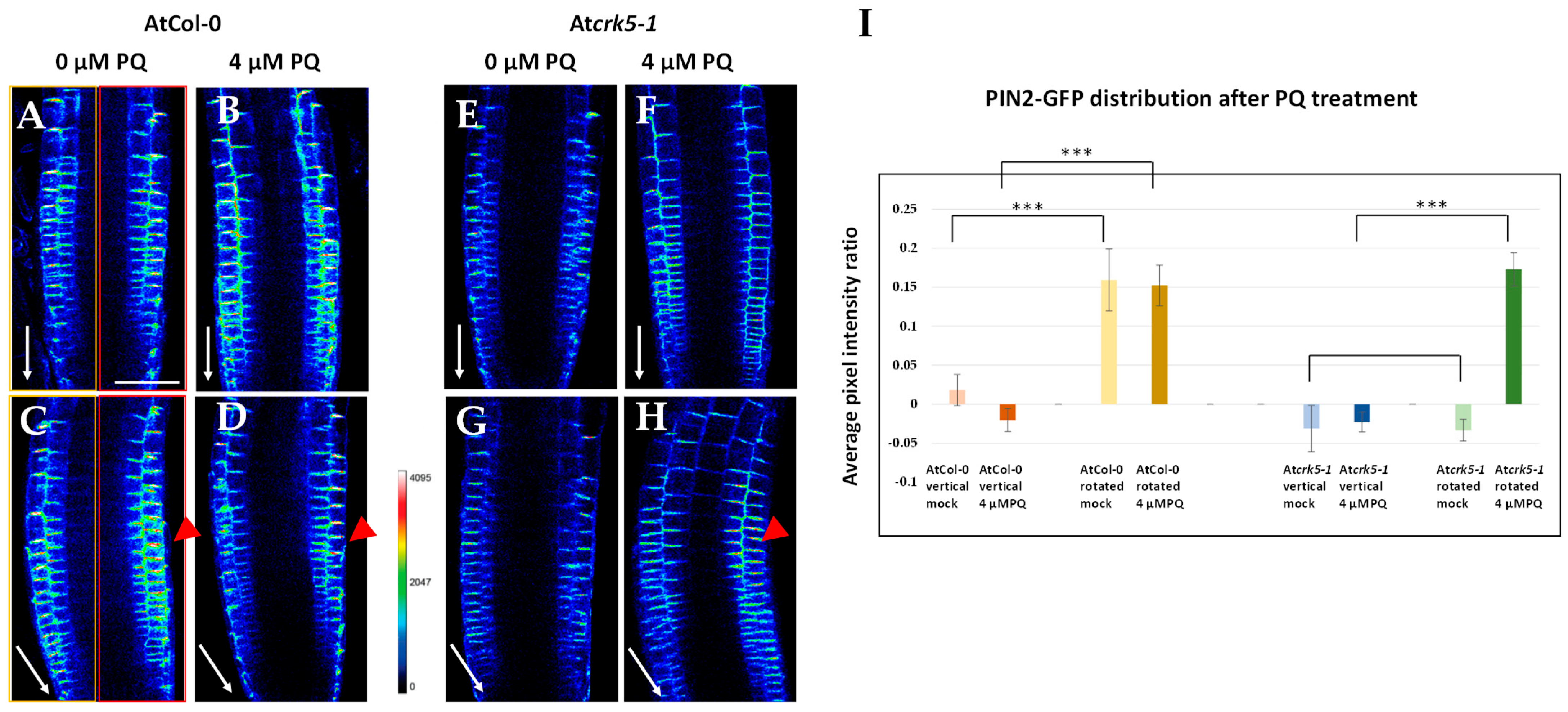
2.4. PQ Restores the PIN2-GFP Distribution in the Root Meristem of the Atcrk5-1 Mutant during Gravistimulation
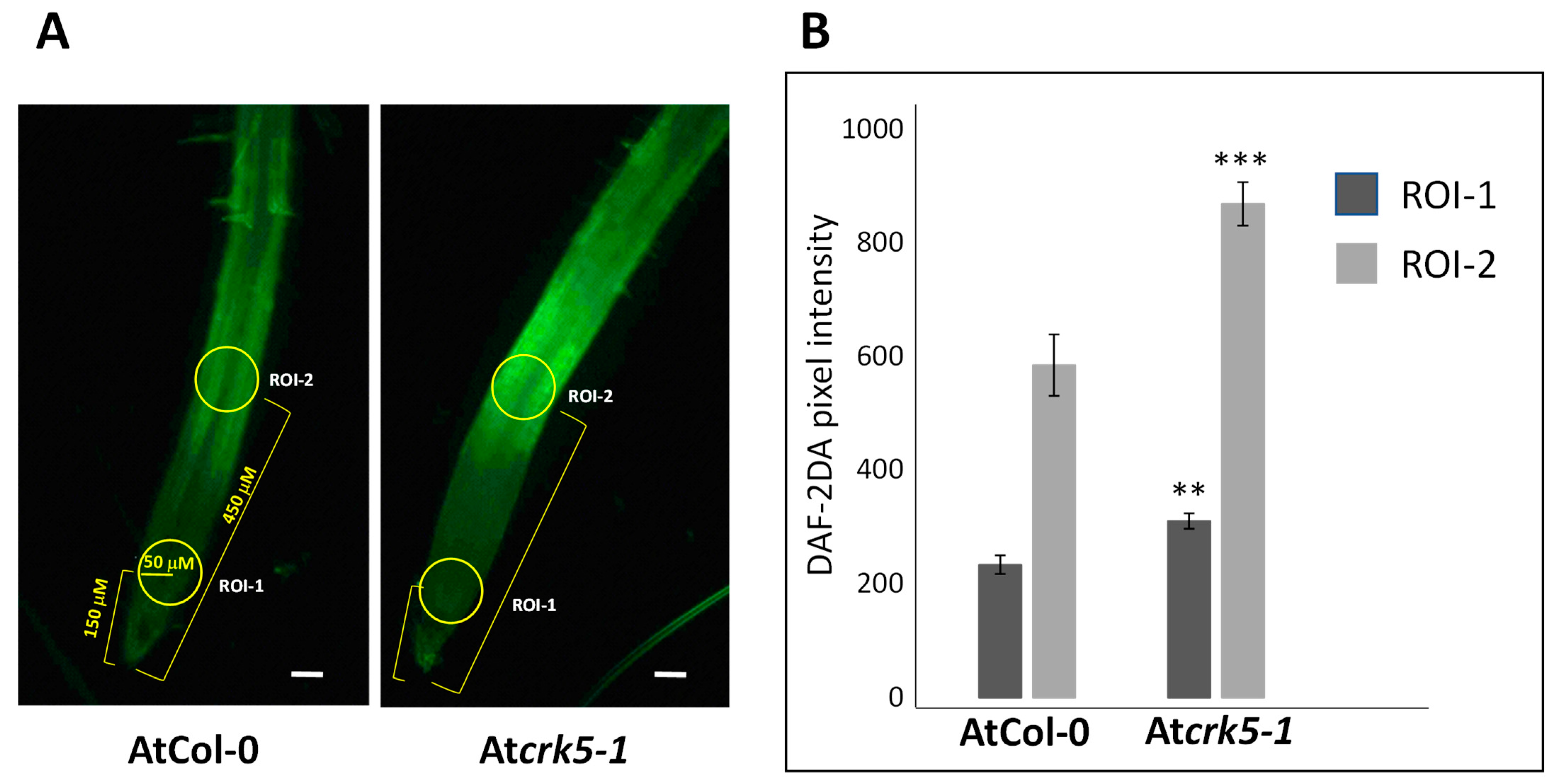
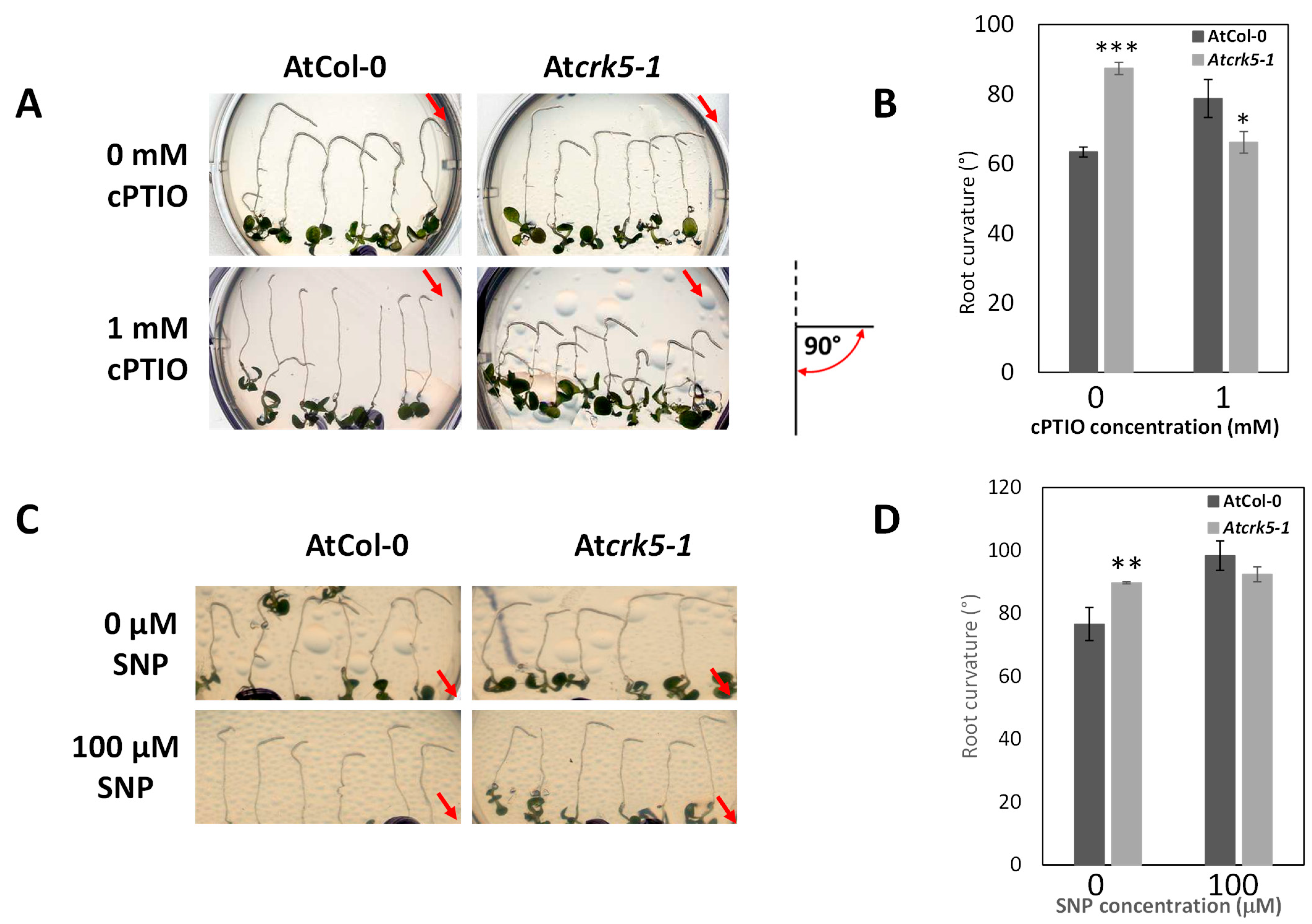
2.5. Increased Nitric Oxide Level in Its Root Apices Contributes to the Delayed Gravitropic Bending of the Atcrk5-1 Mutant
2.6. PQ Treatment Restores the Wild Type NO Level in Atcrk5-1 Mutant Root Tips
3. Discussion
3.1. The AtCRK5 Kinase Controls the Root Gravitropic Response Facilitating the Redistribution of Auxin
3.2. AtCRK5 Is Required to Maintain the ROS Homeostasis at the Root Tip Region That Is Needed for Proper Root Growth and Gravitropic Response
3.3. Nitric Oxide Might Mediate the Effect of AtCRK5-Dependent ROS Generation on Auxin Transport
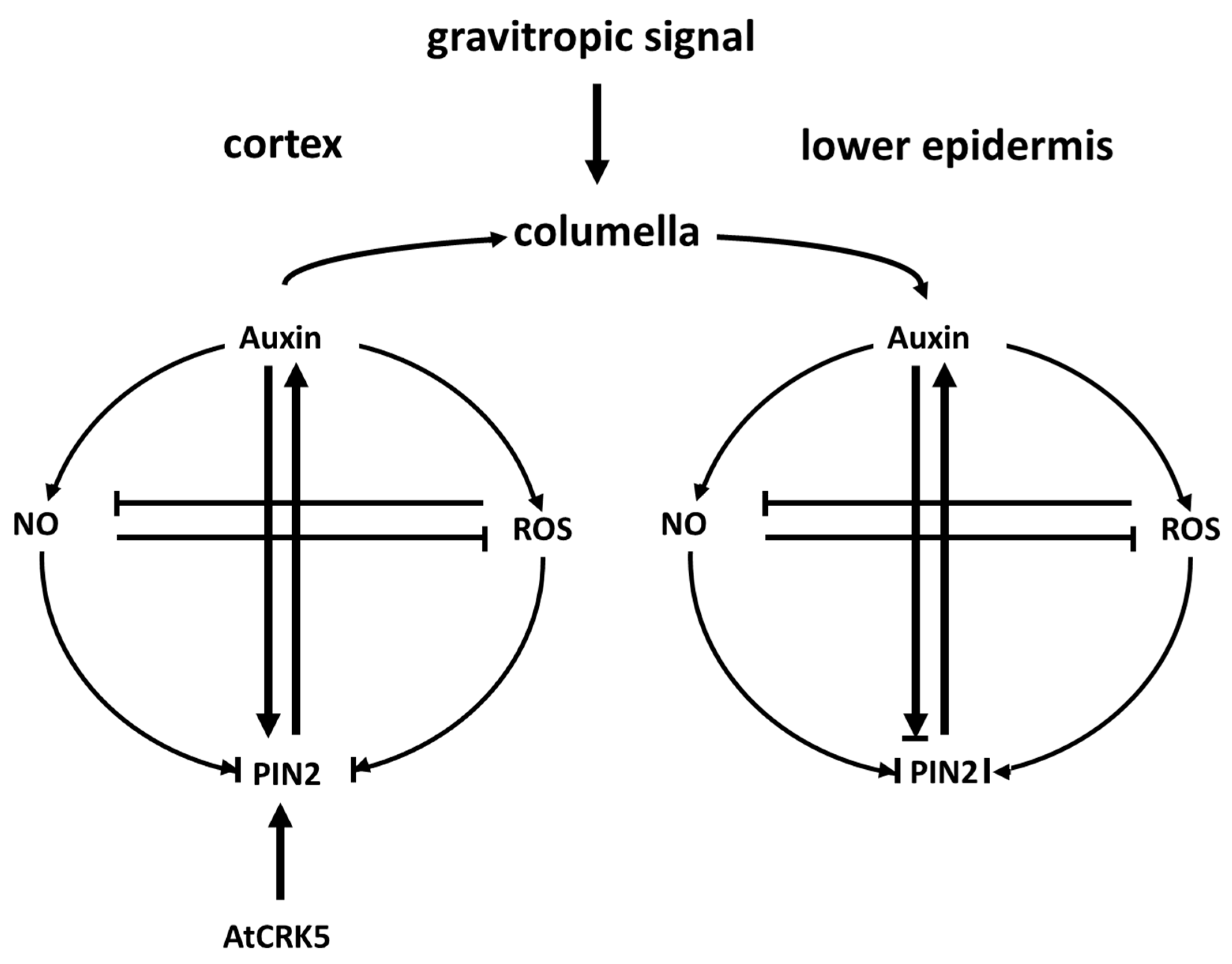
3.4. The AtCRK5 Kinase Fine Tunes the Feedback Loop Including PIN2, Auxin, ROS, and NO That Controls the Gravitropic Response of Arabidopsis Roots
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Root Gravitropic Assays
4.3. Paraquat and Hydrogen Peroxide Tissue Culture Tests
4.4. Histochemical In Situ Detection of ROS
4.5. DR5::GFP and PIN2-GFP Distribution under Oxidative Tests in Roots
4.6. Confocal Laser Scanning Microscopy
4.7. Statistical Analysis
4.8. Accession Numbers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, I. Gravitropism: Lateral Thinking in Auxin Transport. Curr. Biol. 2002, 12, R452–R454. [Google Scholar] [CrossRef] [Green Version]
- Esmon, C.A.; Pedmale, U.V.; Liscum, E. Plant Tropisms: Providing the Power of Movement to a Sessile Organism. Int. J. Dev. Biol. 2004, 49, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.T. Happy End in Sight after 70 Years of Controversy. Trends Plant Sci. 2003, 8, 359–360. [Google Scholar] [CrossRef]
- Swarup, R.; Bennett, M.J. Root Gravitropism. In Annual Plant Reviews Online; American Cancer Society: Atlanta, GA, USA, 2018; pp. 157–174. ISBN 978-1-119-31299-4. [Google Scholar]
- Su, S.-H.; Gibbs, N.M.; Jancewicz, A.L.; Masson, P.H. Molecular Mechanisms of Root Gravitropism. Curr. Biol. 2017, 27, R964–R972. [Google Scholar] [CrossRef] [Green Version]
- Péret, B.; Swarup, K.; Ferguson, A.; Seth, M.; Yang, Y.; Dhondt, S.; James, N.; Casimiro, I.; Perry, P.; Syed, A.; et al. AUX/LAX Genes Encode a Family of Auxin Influx Transporters That Perform Distinct Functions during Arabidopsis Development. Plant Cell 2012, 24, 2874–2885. [Google Scholar] [CrossRef] [Green Version]
- Křeček, P.; Skůpa, P.; Libus, J.; Naramoto, S.; Tejos, R.; Friml, J.; Zažímalová, E. The PIN-FORMED (PIN) Protein Family of Auxin Transporters. Genome Biol. 2009, 10, 249. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.; Cho, H. The Function of ABCB Transporters in Auxin Transport. Plant Signal. Behav. 2013, 8, e22990. [Google Scholar] [CrossRef] [Green Version]
- Blakeslee, J.J.; Peer, W.A.; Murphy, A.S. Auxin Transport. Curr. Opin. Plant Biol. 2005, 8, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN Auxin Efflux Facilitator Network Controls Growth and Patterning in Arabidopsis Roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef]
- Abas, L.; Benjamins, R.; Malenica, N.; Paciorek, T.; Wišniewska, J.; Moulinier–Anzola, J.C.; Sieberer, T.; Friml, J.; Luschnig, C. Intracellular Trafficking and Proteolysis of the Arabidopsis Auxin-Efflux Facilitator PIN2 Are Involved in Root Gravitropism. Nat. Cell Biol. 2006, 8, 249–256. [Google Scholar] [CrossRef]
- Müller, A.; Guan, C.; Gälweiler, L.; Tänzler, P.; Huijser, P.; Marchant, A.; Parry, G.; Bennett, M.; Wisman, E.; Palme, K. AtPIN2 Defines a Locus of Arabidopsis for Root Gravitropism Control. EMBO J. 1998, 17, 6903–6911. [Google Scholar] [CrossRef]
- Adamowski, M.; Friml, J. PIN-Dependent Auxin Transport: Action, Regulation, and Evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunewald, W.; Friml, J. The March of the PINs: Developmental Plasticity by Dynamic Polar Targeting in Plant Cells. EMBO J. 2010, 29, 2700–2714. [Google Scholar] [CrossRef] [PubMed]
- Baster, P.; Robert, S.; Kleine-Vehn, J.; Vanneste, S.; Kania, U.; Grunewald, W.; De Rybel, B.; Beeckman, T.; Friml, J. SCFTIR1/AFB-Auxin Signalling Regulates PIN Vacuolar Trafficking and Auxin Fluxes during Root Gravitropism. EMBO J. 2013, 32, 260–274. [Google Scholar] [CrossRef] [Green Version]
- Sato, E.M.; Hijazi, H.; Bennett, M.J.; Vissenberg, K.; Swarup, R. New Insights into Root Gravitropic Signalling. J. Exp. Bot. 2015, 66, 2155–2165. [Google Scholar] [CrossRef]
- Barbosa, I.C.R.; Hammes, U.Z.; Schwechheimer, C. Activation and Polarity Control of PIN-FORMED Auxin Transporters by Phosphorylation. Trends Plant Sci. 2018, 23, 523–538. [Google Scholar] [CrossRef]
- Michniewicz, M.; Zago, M.K.; Abas, L.; Weijers, D.; Schweighofer, A.; Meskiene, I.; Heisler, M.G.; Ohno, C.; Zhang, J.; Huang, F.; et al. Antagonistic Regulation of PIN Phosphorylation by PP2A and PINOID Directs Auxin Flux. Cell 2007, 130, 1044–1056. [Google Scholar] [CrossRef] [Green Version]
- Sukumar, P.; Edwards, K.S.; Rahman, A.; DeLong, A.; Muday, G.K. PINOID Kinase Regulates Root Gravitropism through Modulation of PIN2-Dependent Basipetal Auxin Transport in Arabidopsis. Plant Physiol. 2009, 150, 722–735. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Mulkey, T.J.; Evans, M.L. Reversible Loss of Gravitropic Sensitivity in Maize Roots after Tip Application of Calcium Chelators. Science 1983, 220, 1375–1376. [Google Scholar] [CrossRef]
- Monshausen, G.; Miller, N.; Murphy, A.; Gilroy, S. Dynamics of Auxin-Dependent Ca2+ and PH Signaling in Root Growth Revealed by Integrating High-Resolution Imaging with Automated Computer Vision-Based Analysis. Plant J. 2011, 65, 309–318. [Google Scholar] [CrossRef]
- Plieth, C.; Trewavas, A. Reorientation of Seedlings in the Earth’s Gravitational Field Induces Cytosolic Calcium Transients. Plant Physiol. 2002, 129, 786–796. [Google Scholar] [CrossRef] [Green Version]
- Tatsumi, H.; Toyota, M.; Furuichi, T.; Sokabe, M. Calcium Mobilizations in Response to Changes in the Gravity Vector in Arabidopsis Seedlings. Possible Cellular Mechanisms. Plant Signal. Behav. 2014, 9, 29099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, Y.; Jia, W.-J.; Chu, Y.-J.; Xue, H.-W. Arabidopsis Phosphatidylinositol Monophosphate 5-Kinase 2 Is Involved in Root Gravitropism through Regulation of Polar Auxin Transport by Affecting the Cycling of PIN Proteins. Cell Res. 2012, 22, 581–597. [Google Scholar] [CrossRef] [Green Version]
- Joo, J.H.; Yoo, H.J.; Hwang, I.; Lee, J.S.; Nam, K.H.; Bae, Y.S. Auxin-Induced Reactive Oxygen Species Production Requires the Activation of Phosphatidylinositol 3-Kinase. FEBS Lett. 2005, 579, 1243–1248. [Google Scholar] [CrossRef] [Green Version]
- Perera, I.; Hung, C.-Y.; Brady, S.; Muday, G.; Boss, W. A Universal Role for Inositol 1,4,5-Trisphosphate-Mediated Signaling in Plant Gravitropism. Plant Physiol. 2006, 140, 746–760. [Google Scholar] [CrossRef] [Green Version]
- Perera, I.Y.; Heilmann, I.; Boss, W.F. Transient and Sustained Increases in Inositol 1,4,5-Trisphosphate Precede the Differential Growth Response in Gravistimulated Maize Pulvini. Proc. Natl. Acad. Sci. USA 1999, 96, 5838–5843. [Google Scholar] [CrossRef] [Green Version]
- Scott, A.C.; Allen, N.S. Changes in Cytosolic PH within Arabidopsis Root Columella Cells Play a Key Role in the Early Signaling Pathway for Root Gravitropism. Plant Physiol. 1999, 121, 1291–1298. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Neill, S.J.; Tang, Z.; Cai, W. Nitric Oxide Mediates Gravitropic Bending in Soybean Roots. Plant Physiol. 2005, 137, 663–670. [Google Scholar] [CrossRef] [Green Version]
- París, R.; Vazquez, M.M.; Graziano, M.; Terrile, M.C.; Miller, N.D.; Spalding, E.P.; Otegui, M.S.; Casalongué, C.A. Distribution of Endogenous NO Regulates Early Gravitropic Response and PIN2 Localization in Arabidopsis Roots. Front. Plant Sci. 2018, 9, 495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, J.H.; Bae, Y.S.; Lee, J.S. Role of Auxin-Induced Reactive Oxygen Species in Root Gravitropism. Plant Physiol. 2001, 126, 1055–1060. [Google Scholar] [CrossRef] [Green Version]
- Krieger, G.; Shkolnik, D.; Miller, G.; Fromm, H. Reactive Oxygen Species Tune Root Tropic Responses. Plant Phyisol. 2016, 172, 1209–1220. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Hou, H.; Yang, T.; Lian, Y.; Sun, Y.; Bian, Z.; Wang, C. Exogenous Hydrogen Peroxide Inhibits Primary Root Gravitropism by Regulating Auxin Distribution during Arabidopsis Seed Germination. Plant Physiol. Biochem. 2018, 128, 126–133. [Google Scholar] [CrossRef]
- Baba, A.; Rigó, G.; Ayaydin, F.; Rehman, A.; Andrási, N.; Zsigmond, L.; Valkai, I.; Urbancsok, J.; Vass, I.; Pasternak, T.; et al. Functional Analysis of the Arabidopsis Thaliana CDPK-Related Kinase Family: AtCRK1 Regulates Responses to Continuous Light. Int. J. Mol. Sci. 2018, 19, 1282. [Google Scholar] [CrossRef] [Green Version]
- Rigó, G.; Ayaydin, F.; Tietz, O.; Zsigmond, L.; Kovács, H.; Páy, A.; Salchert, K.; Darula, Z.; Medzihradszky, K.F.; Szabados, L.; et al. Inactivation of Plasma Membrane-Localized CDPK-RELATED KINASE5 Decelerates PIN2 Exocytosis and Root Gravitropic Response in Arabidopsis. Plant Cell 2013, 25, 1592–1608. [Google Scholar] [CrossRef] [Green Version]
- Delormel, T.Y.; Boudsocq, M. Properties and Functions of Calcium-Dependent Protein Kinases and Their Relatives in Arabidopsis Thaliana. New Phytol. 2019, 224, 585–604. [Google Scholar] [CrossRef] [Green Version]
- Baba, A.I.; Andrási, N.; Valkai, I.; Gorcsa, T.; Koczka, L.; Darula, Z.; Medzihradszky, K.F.; Szabados, L.; Fehér, A.; Rigó, G.; et al. AtCRK5 Protein Kinase Exhibits a Regulatory Role in Hypocotyl Hook Development during Skotomorphogenesis. Int. J. Mol. Sci. 2019, 20, 3432. [Google Scholar] [CrossRef] [Green Version]
- Baba, A.I.; Valkai, I.; Labhane, N.; Koczka, L.; Andrasi, N.; Klement, E.; Darula, Z.; Medzihradszky, K.; Szabados, L.; Fehér, A.; et al. CRK5 Protein Kinase Contributes to the Progression of Embryogenesis of Arabidopsis Thaliana. Int. J. Mol. Sci. 2019, 20, 6120. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Xiang, Y.; Li, C.; Yu, G. Modulatory Role of Reactive Oxygen Species in Root Development in Model Plant of Arabidopsis Thaliana. Front. Plant Sci. 2020, 11, 485932. [Google Scholar] [CrossRef]
- Liszkay, A.; van der Zalm, E.; Schopfer, P. Production of Reactive Oxygen Intermediates (O2-, H2O2, and OH) by Maize Roots and Their Role in Wall Loosening and Elongation Growth. Plant Physiol. 2004, 136, 3114–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peer, W.A.; Cheng, Y.; Murphy, A.S. Evidence of Oxidative Attenuation of Auxin Signalling. J. Exp. Bot. 2013, 64, 2629–2639. [Google Scholar] [CrossRef] [PubMed]
- Zwiewka, M.; Bielach, A.; Tamizhselvan, P.; Madhavan, S.; Ryad, E.E.; Tan, S.; Hrtyan, M.N.; Dobrev, P.; Vankovï, R.; Friml, J.; et al. Root Adaptation to H2O2-Induced Oxidative Stress by ARF-GEF BEN1- and Cytoskeleton-Mediated PIN2 Trafficking. Plant Cell Physiol. 2019, 60, 255–273. [Google Scholar] [CrossRef] [PubMed]
- Correa-Aragunde, N.; Foresi, N.; Lamattina, L. Chapter Three—Auxin and Nitric Oxide: A Counterbalanced Partnership Ensures the Redox Cue Control Required for Determining Root Growth Pattern. In Advances in Botanical Research; Nitric Oxide and Signaling in Plants; Wendehenne, D., Ed.; Academic Press: Cambridge, MA, USA, 2016; Volume 77, pp. 41–54. [Google Scholar]
- Correa-Aragunde, N.; París, R.; Foresi, N.; Terrile, C.; Casalongué, C.; Lamattina, L. The Auxin-Nitric Oxide Highway: A Right Direction in Determining the Plant Root System. In Gasotransmitters in Plants: The Rise of a New Paradigm in Cell Signaling; Signaling and Communication in Plants; Lamattina, L., García-Mata, C., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 117–136. ISBN 978-3-319-40713-5. [Google Scholar]
- Lanteri, M.L.; Graziano, M.; Correa-Aragunde, N.; Lamattina, L. From Cell Division to Organ Shape: Nitric Oxide Is Involved in Auxin-Mediated Root Development. In Communication in Plants: Neuronal Aspects of Plant Life; Baluška, F., Mancuso, S., Volkmann, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 123–136. ISBN 978-3-540-28516-8. [Google Scholar]
- Sanz, L.; Fernández-Marcos, M.; Modrego, A.; Lewis, D.R.; Muday, G.K.; Pollmann, S.; Dueñas, M.; Santos-Buelga, C.; Lorenzo, O. Nitric Oxide Plays a Role in Stem Cell Niche Homeostasis through Its Interaction with Auxin. Plant Physiol. 2014, 166, 1972–1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Marcos, M.; Sanz, L.; Lewis, D.R.; Muday, G.K.; Lorenzo, O. Nitric Oxide Causes Root Apical Meristem Defects and Growth Inhibition While Reducing PIN-FORMED 1 (PIN1)-Dependent Acropetal Auxin Transport. Proc. Natl. Acad. Sci. USA 2011, 108, 18506–18511. [Google Scholar] [CrossRef] [Green Version]
- Terrile, M.; Parıs, R.; Calderon-Villalobos, L.; Iglesias, M.; Lamattina, L.; Estelle, M.; Casalongue, C. Nitric Oxide Influences Auxin Signaling through S-Nitrosylation of the Arabidopsis Transport Inhibitor Response 1 Auxin Receptor. Plant J. 2012, 70, 492–500. [Google Scholar] [CrossRef] [Green Version]
- Prakash, V.; Vishwakarma, K.; Singh, V.P.; Rai, P.; Ramawat, N.; Tripathi, D.K.; Sharma, S. NO and ROS Implications in the Organization of Root System Architecture. Physiol. Plant 2020, 168, 473–489. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; Barroso, J.B. Functions of Nitric Oxide (NO) in Roots during Development and under Adverse Stress Conditions. Plants 2015, 4, 240–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, M.S.; Fukaki, H.; Mori, I.C.; Nakahara, K.; Mano, J. Reactive Oxygen Species and Reactive Carbonyl Species Constitute a Feed-Forward Loop in Auxin Signaling for Lateral Root Formation. Plant J. 2019, 100, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Tsukagoshi, H. Control of Root Growth and Development by Reactive Oxygen Species. Curr. Opin. Plant Biol. 2016, 29, 57–63. [Google Scholar] [CrossRef]
- Dunand, C.; Crèvecoeur, M.; Penel, C. Distribution of Superoxide and Hydrogen Peroxide in Arabidopsis Root and Their Influence on Root Development: Possible Interaction with Peroxidases. New Phytol. 2007, 174, 332–341. [Google Scholar] [CrossRef]
- Kagenishi, T.; Yokawa, K.; Baluška, F. MES Buffer Affects Arabidopsis Root Apex Zonation and Root Growth by Suppressing Superoxide Generation in Root Apex. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Lascano, R.; Munoz, N.; Robert, G.; Rodriguez, M.; Melchiorre, M.; Trippi, V.; Quero, G. Paraquat: An Oxidative Stress Inducer. In Herbicide—Properties, Synthesis and Control of Weeds; Hasaneen, M.N., Ed.; InTech: London, UK, 2012; ISBN 978-953-307-803-8. [Google Scholar]
- Cochemé, H.M.; Murphy, M.P. Chapter 22 The Uptake and Interactions of the Redox Cycler Paraquat with Mitochondria. In Methods in Enzymology; Mitochondrial Function, Part A: Mitochondrial Electron Transport Complexes and Reactive Oxygen Species; Academic Press: Cambridge, MA, USA, 2009; Volume 456, pp. 395–417. [Google Scholar]
- Band, L.R.; Wells, D.M.; Larrieu, A.; Sun, J.; Middleton, A.M.; French, A.P.; Brunoud, G.; Sato, E.M.; Wilson, M.H.; Péret, B.; et al. Root Gravitropism Is Regulated by a Transient Lateral Auxin Gradient Controlled by a Tipping-Point Mechanism. Proc. Natl. Acad. Sci. USA 2012, 109, 4668–4673. [Google Scholar] [CrossRef] [Green Version]
- Konstantinova, N.; Korbei, B.; Luschnig, C. Auxin and Root Gravitropism: Addressing Basic Cellular Processes by Exploiting a Defined Growth Response. Int. J. Mol. Sci. 2021, 22, 2749. [Google Scholar] [CrossRef]
- Ottenschläger, I.; Wolff, P.; Wolverton, C.; Bhalerao, R.P.; Sandberg, G.; Ishikawa, H.; Evans, M.; Palme, K. Gravity-Regulated Differential Auxin Transport from Columella to Lateral Root Cap Cells. Proc. Natl. Acad. Sci. USA 2003, 100, 2987–2991. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Takahashi, M.; Shibasaki, K.; Wu, S.; Inaba, T.; Tsurumi, S.; Baskin, T.I. Gravitropism of Arabidopsis Thaliana Roots Requires the Polarization of PIN2 toward the Root Tip in Meristematic Cortical Cells. Plant Cell 2010, 22, 1762–1776. [Google Scholar] [CrossRef] [Green Version]
- Rashotte, A.M.; Brady, S.R.; Reed, R.C.; Ante, S.J.; Muday, G.K. Basipetal Auxin Transport Is Required for Gravitropism in Roots of Arabidopsis. Plant Physiol. 2000, 122, 481–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, M.M.; Casalongué, C.A.; París, R. Nitrate Reductase Mediates Nitric Oxide-Dependent Gravitropic Response in Arabidopsis Thaliana Roots. Plant Signal. Behav. 2019, 14, e1578631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolbert, Z.; Feigl, G. Cross-talk of Reactive Oxygen Species and Nitric Oxide in Various Processes of Plant Development. In Reactive Oxygen Species in Plants; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 261–289. ISBN 978-1-119-32492-8. [Google Scholar]
- Lewis, D.; Miller, N.; Splitt, B.; Wu, G.; Spalding, E. Separating the Roles of Acropetal and Basipetal Auxin Transport on Gravitropism with Mutations in Two Arabidopsis Multidrug Resistance-like ABC Transporter Genes. Plant Cell 2007, 19, 1838–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, G.A.; Okuyama, H. Lipid-Linked Proteins of Plants. Prog. Lipid Res. 2000, 39, 19–39. [Google Scholar] [CrossRef]
- Shin, H.; Shin, H.-S.; Guo, Z.; Blancaflor, E.B.; Masson, P.H.; Chen, R. Complex Regulation of Arabidopsis AGR1/PIN2-Mediated Root Gravitropic Response and Basipetal Auxin Transport by Cantharidin-Sensitive Protein Phosphatases. Plant J. 2005, 42, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Dunand, C.; Penel, C. Localization of Superoxide in the Root Apex of Arabidopsis. Plant Signal. Behav. 2007, 2, 131–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livanos, P.; Galatis, B.; Apostolakos, P. The Interplay between ROS and Tubulin Cytoskeleton in Plants. Plant Signal. Behav. 2014, 9, e28069. [Google Scholar] [CrossRef]
- Ambrose, C.; Ruan, Y.; Gardiner, J.; Tamblyn, L.M.; Catching, A.; Kirik, V.; Marc, J.; Overall, R.; Wasteneys, G.O. CLASP Interacts with Sorting Nexin 1 to Link Microtubules and Auxin Transport via PIN2 Recycling in Arabidopsis Thaliana. Dev. Cell 2013, 24, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Fujioka, S.; Peng, J.; Chen, J.; Li, G.; Chen, R. The E3 Ubiquitin Ligase SCFTIR1/AFB and Membrane Sterols Play Key Roles in Auxin Regulation of Endocytosis, Recycling, and Plasma Membrane Accumulation of the Auxin Efflux Transporter PIN2 in Arabidopsis Thaliana. Plant Cell 2009, 21, 568–580. [Google Scholar] [CrossRef] [Green Version]
- Kolbert, Z.; Bartha, B.; Erdei, L. Exogenous Auxin-Induced NO Synthesis Is Nitrate Reductase-Associated in Arabidopsis Thaliana Root Primordia. J. Plant Physiol. 2008, 165, 967–975. [Google Scholar] [CrossRef]
- Lombardo, M.C.; Graziano, M.; Polacco, J.C.; Lamattina, L. Nitric Oxide Functions as a Positive Regulator of Root Hair Development. Plant Signal. Behav. 2006, 1, 28–33. [Google Scholar] [CrossRef] [Green Version]
- Pagnussat, G.C.; Simontacchi, M.; Puntarulo, S.; Lamattina, L. Nitric Oxide Is Required for Root Organogenesis. Plant Physiol. 2002, 129, 954–956. [Google Scholar] [CrossRef] [Green Version]
- Kolbert, Z.; Menesi, D.; Molnár, Á.; Kénesi, E.; Valkai, I.; Feigl, G.; Rigó, G.; Cséplő, Á.; Lindermayr, C.; Fehér, A. S-Nitrosation of ROP2 Is Involved in Nitric Oxide (NO)-Induced Root Shortening in Arabidopsis. J. Exp. Bot. 2021. submitted. [Google Scholar]
- Astier, J.; Besson-Bard, A.; Wawer, I.; Parent, C.; Rasul, S.; Jeandroz, S.; Dat, J.; Wendehenne, D. Nitric Oxide Signalling in Plants: Cross-Talk with Ca2+, Protein Kinases and Reactive Oxygen Species. In Annual Plant Reviews Online; American Cancer Society: Atlanta, GA, USA, 2018; pp. 147–170. ISBN 978-1-119-31299-4. [Google Scholar]
- Lindermayr, C. Crosstalk between Reactive Oxygen Species and Nitric Oxide in Plants: Key Role of S-Nitrosoglutathione Reductase. Free Radic. Biol. Med. 2018, 122, 110–115. [Google Scholar] [CrossRef]
- Lindermayr, C.; Durner, J. Interplay of Reactive Oxygen Species and Nitric Oxide: Nitric Oxide Coordinates Reactive Oxygen Species Homeostasis. Plant Physiol. 2015, 167, 1209–1210. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Ziguo Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Pető, A.; Lehotai, N.; Lozano-Juste, J.; León, J.; Tari, I.; Erdei, L.; Kolbert, Z. Involvement of nitric oxide (NO) in signal transduction of copper-induced morphological responses in Arabidopsis seedlings. Ann. Bot. 2011, 108, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinf. 2017, 18, 529. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cséplő, Á.; Zsigmond, L.; Andrási, N.; Baba, A.I.; Labhane, N.M.; Pető, A.; Kolbert, Z.; Kovács, H.E.; Steinbach, G.; Szabados, L.; et al. The AtCRK5 Protein Kinase Is Required to Maintain the ROS NO Balance Affecting the PIN2-Mediated Root Gravitropic Response in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 5979. https://doi.org/10.3390/ijms22115979
Cséplő Á, Zsigmond L, Andrási N, Baba AI, Labhane NM, Pető A, Kolbert Z, Kovács HE, Steinbach G, Szabados L, et al. The AtCRK5 Protein Kinase Is Required to Maintain the ROS NO Balance Affecting the PIN2-Mediated Root Gravitropic Response in Arabidopsis. International Journal of Molecular Sciences. 2021; 22(11):5979. https://doi.org/10.3390/ijms22115979
Chicago/Turabian StyleCséplő, Ágnes, Laura Zsigmond, Norbert Andrási, Abu Imran Baba, Nitin M. Labhane, Andrea Pető, Zsuzsanna Kolbert, Hajnalka E. Kovács, Gábor Steinbach, László Szabados, and et al. 2021. "The AtCRK5 Protein Kinase Is Required to Maintain the ROS NO Balance Affecting the PIN2-Mediated Root Gravitropic Response in Arabidopsis" International Journal of Molecular Sciences 22, no. 11: 5979. https://doi.org/10.3390/ijms22115979