
Current Strategies for the Regeneration of Skeletal Muscle Tissue


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
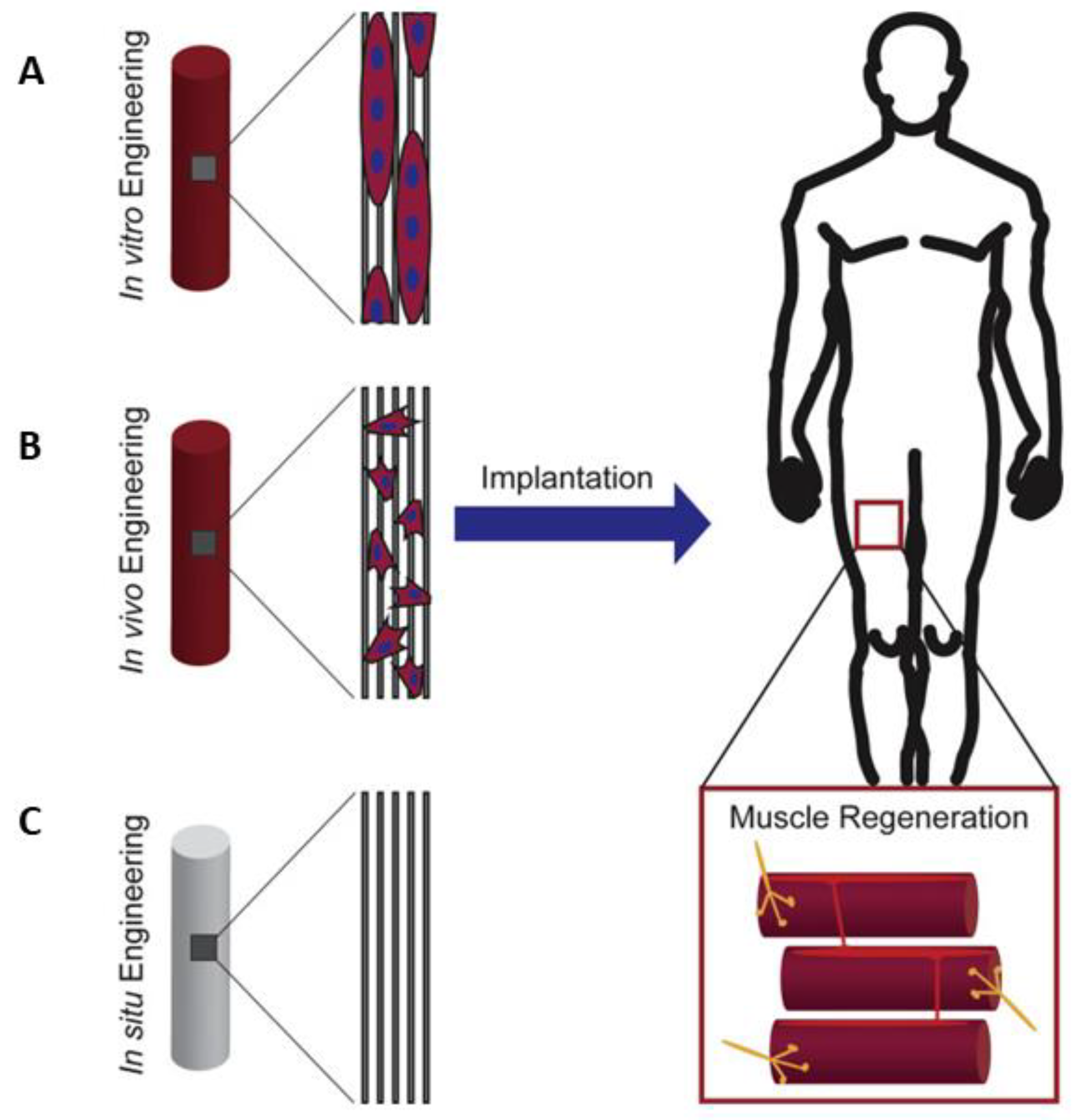
2. Strategies for Skeletal Muscle Regeneration
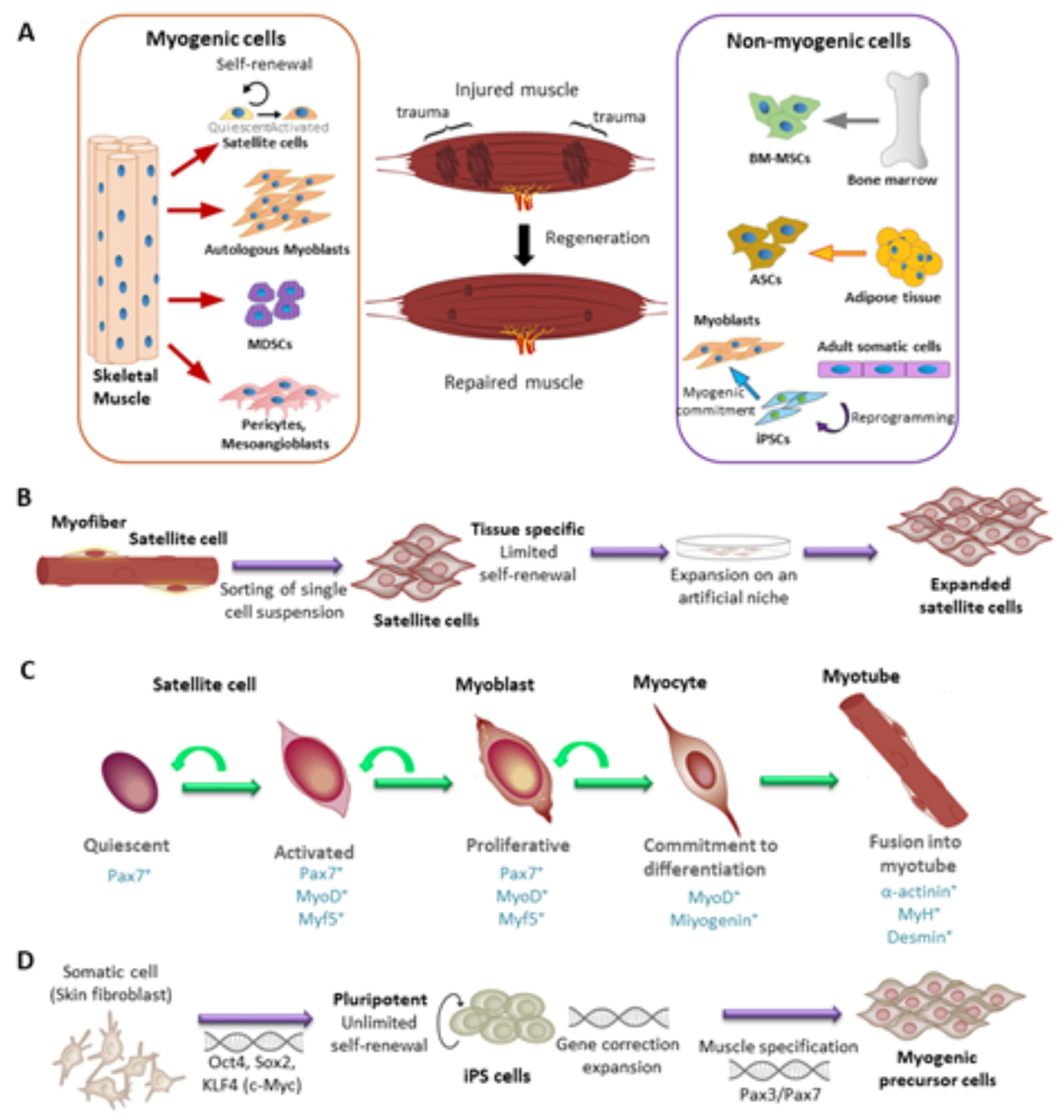
3. Cell Types for the Regeneration of Skeletal Muscle
3.1. Satellite Cells (SCs) and Myoblasts
3.2. Mesenchymal Stromal Cells (MSCs)
3.3. Muscle Derived Stem Cells (MDSCs)
3.4. Pericytes
3.5. Induced Pluripotent Stem Cells (iPSCs)
4. Biomaterials for the Transplantation of Striated Muscle Cells
4.1. Natural Hydrogels
4.2. Synthetic Hydrogels
4.3. Composite (Hybrid) Hydrogels
5. Biomechanical Strategies for the Generation of Skeletal Muscle Tissue
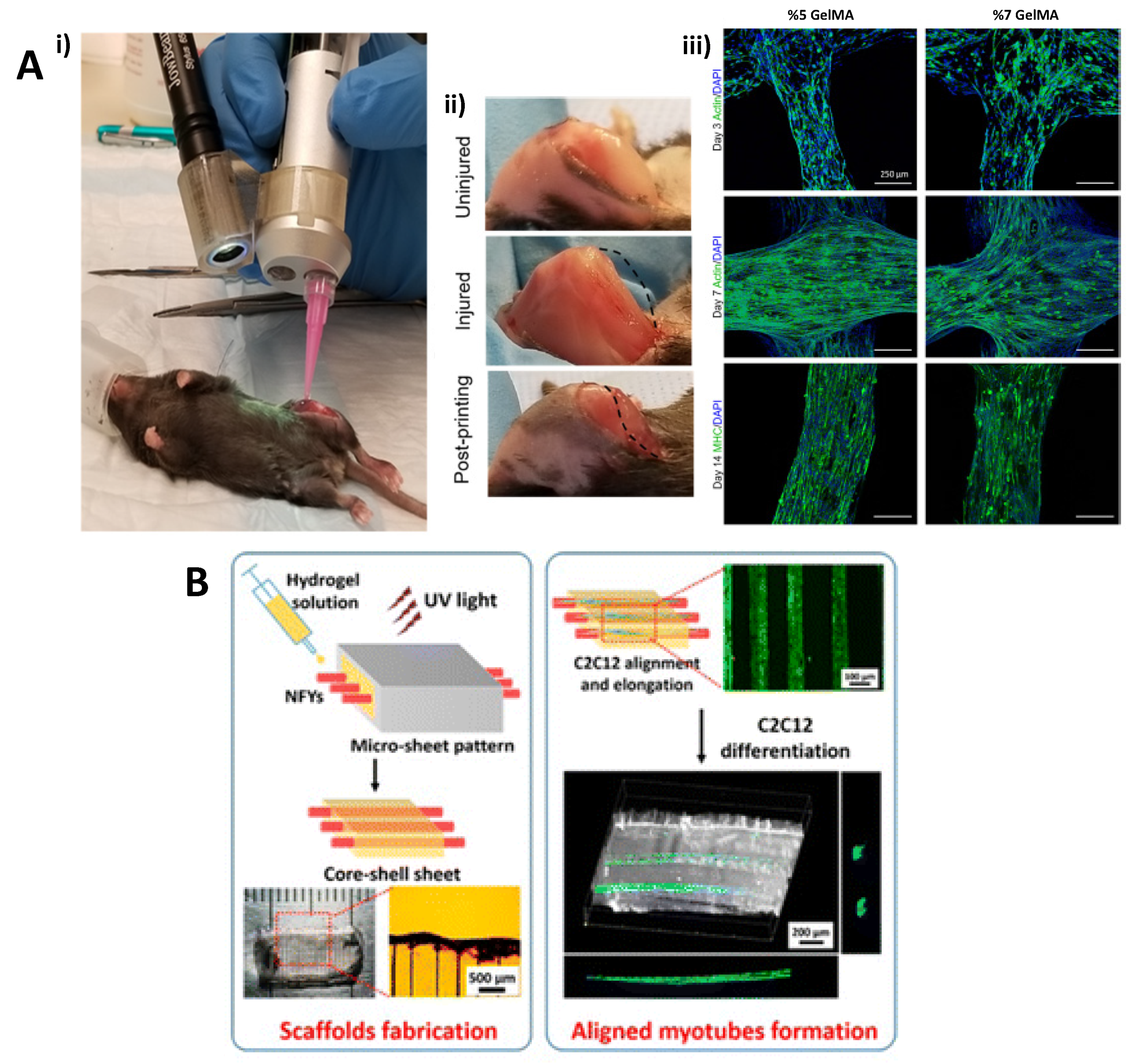
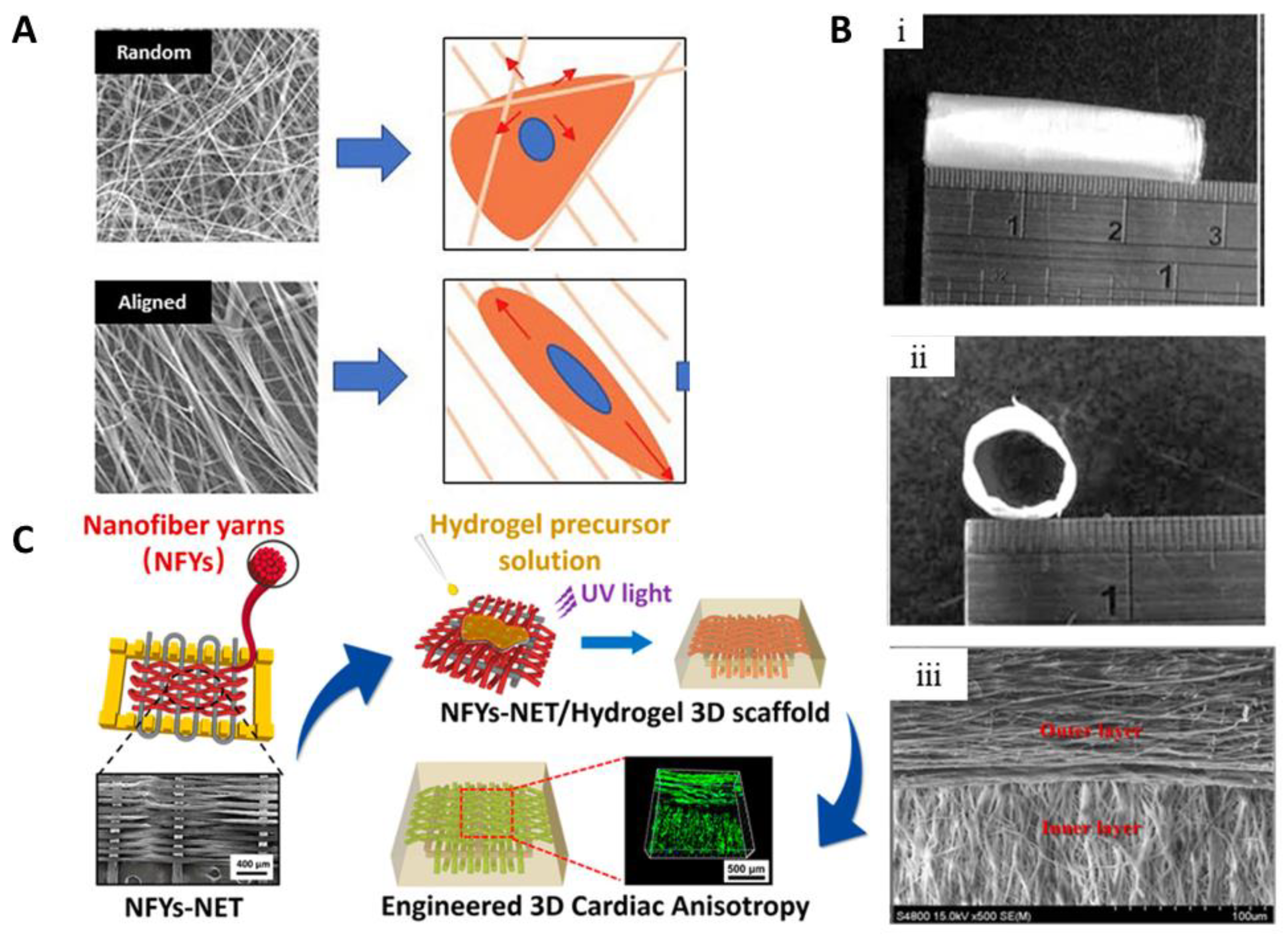
6. Scaffold Topographies for Improved Generation of Muscle Tissue
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qazi, T.H.; Mooney, D.J.; Pumberger, M.; Geissler, S.; Duda, G.N. Biomaterials based strategies for skeletal muscle tissue engineering: Existing technologies and future trends. Biomaterials 2015, 53, 502–521. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Saul, D.; Boker, K.O.; Ernst, J.; Lehman, W.; Schilling, A.F. Current Methods for Skeletal Muscle Tissue Repair and Regeneration. Biomed. Res. Int. 2018, 2018, 1984879. [Google Scholar] [CrossRef] [PubMed]
- Carnes, M.E.; Pins, G.D. Skeletal Muscle Tissue Engineering: Biomaterials-Based Strategies for the Treatment of Volumetric Muscle Loss. Bioengineering 2020, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Forcina, L.; Cosentino, M.; Musaro, A. Mechanisms Regulating Muscle Regeneration: Insights into the Interrelated and Time-Dependent Phases of Tissue Healing. Cells 2020, 9, 1297. [Google Scholar] [CrossRef] [PubMed]
- Basurto, I.M.; Mora, M.T.; Gardner, G.M.; Christ, G.J.; Caliari, S.R. Aligned and Conductive 3D Collagen Scaffolds for Skeletal Muscle Tissue Engineering. Biomater. Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Cezar, C.A.; Mooney, D.J. Biomaterial-based delivery for skeletal muscle repair. Adv. Drug Deliv. Rev. 2015, 84, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Xing, F.; Li, L.; Zhou, C.; Long, C.; Wu, L.; Lei, H.; Kong, Q.; Fan, Y.; Xiang, Z.; Zhang, X. Regulation and Directing Stem Cell Fate by Tissue Engineering Functional Microenvironments: Scaffold Physical and Chemical Cues. Stem Cells Int. 2019, 2019, 2180925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smoak, M.M.; Mikos, A.G. Advances in biomaterials for skeletal muscle engineering and obstacles still to overcome. Mater. Today Bio. 2020, 100069. [Google Scholar] [CrossRef]
- Jarvinen, T.A.; Jarvinen, M.; Kalimo, H. Regeneration of injured skeletal muscle after the injury. Muscles Ligaments Tendons J. 2013, 3, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Gilbert-Honick, J.; Iyer, S.R.; Somers, S.M.; Lovering, R.M.; Wagner, K.; Mao, H.-Q.; Grayson, W.L. Engineering functional and histological regeneration of vascularized skeletal muscle. Biomaterials 2018, 164, 70–79. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, I.; Seol, Y.-J.; Ko, I.K.; Yoo, J.J.; Atala, A.; Lee, S.J. Neural cell integration into 3D bioprinted skeletal muscle constructs accelerates restoration of muscle function. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Yang, W.; Hu, P. Skeletal muscle regeneration is modulated by inflammation. J. Orthop. Transl. 2018, 13, 25–32. [Google Scholar] [CrossRef] [PubMed]
- McCullen, S.D.; Chow, A.G.Y.; Stevens, M.M. In vivo tissue engineering of musculoskeletal tissues. Curr. Opin. Biotechnol. 2011, 22, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.S.; Mostafavi, A.; Quint, J.P.; Panayi, A.C.; Baldino, K.; Williams, T.J.; Daubendiek, J.G.; Hugo Sánchez, V.; Bonick, Z.; Trujillo-Miranda, M. In situ printing of adhesive hydrogel scaffolds for the treatment of skeletal muscle injuries. ACS Appl. Bio. Mater. 2020, 3, 1568–1579. [Google Scholar] [CrossRef]
- Qazi, T.H.; Duda, G.N.; Ort, M.J.; Perka, C.; Geissler, S.; Winkler, T. Cell therapy to improve regeneration of skeletal muscle injuries. J. Cachexia Sarcopenia Muscle 2019, 10, 501–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantelic, M.N.; Larkin, L.M. Stem cells for skeletal muscle tissue engineering. Tissue Eng. Part. B Rev. 2018, 24, 373–391. [Google Scholar] [CrossRef]
- McCullagh, K.J.A.; Perlingeiro, R.C.R. Coaxing stem cells for skeletal muscle repair. Adv. Drug Deliv. Rev. 2015, 84, 198–207. [Google Scholar] [CrossRef] [Green Version]
- Collins, C.A.; Olsen, I.; Zammit, P.S.; Heslop, L.; Petrie, A.; Partridge, T.A.; Morgan, J.E. Stem cell function, self-renewal, and behavioral heterogeneity of cells from the adult muscle satellite cell niche. Cell 2005, 122, 289–301. [Google Scholar] [CrossRef] [Green Version]
- Dunn, A.; Talovic, M.; Patel, K.; Patel, A.; Marcinczyk, M.; Garg, K. Biomaterial and stem cell-based strategies for skeletal muscle regeneration. J. Orthop. Res. 2019, 37, 1246–1262. [Google Scholar] [CrossRef]
- Relaix, F.; Zammit, P.S. Satellite cells are essential for skeletal muscle regeneration: The cell on the edge returns centre stage. Development 2012, 139, 2845–2856. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [Green Version]
- Tedesco, F.S.; Dellavalle, A.; Diaz-Manera, J.; Messina, G.; Cossu, G. Repairing skeletal muscle: Regenerative potential of skeletal muscle stem cells. J. Clin. Investig. 2010, 120, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Fishman, J.M.; Tyraskis, A.; Maghsoudlou, P.; Urbani, L.; Totonelli, G.; Birchall, M.A.; De Coppi, P. Skeletal muscle tissue engineering: Which cell to use? Tissue Eng. Part B Rev. 2013, 19, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Yablonka-Reuveni, Z. The skeletal muscle satellite cell: Still young and fascinating at 50. J. Histochem. Cytochem. 2011, 59, 1041–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Grand, F.; Rudnicki, M.A. Skeletal muscle satellite cells and adult myogenesis. Curr. Opin. Cell Biol. 2007, 19, 628–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motohashi, N.; Asakura, Y.; Asakura, A. Isolation, culture, and transplantation of muscle satellite cells. J. Vis. Exp. JOVE 2014, 50846. [Google Scholar] [CrossRef] [PubMed]
- Grefte, S.; Kuijpers-Jagtman, A.M.; Torensma, R.; Von den Hoff, J.W. Skeletal muscle development and regeneration. Stem Cells Dev. 2007, 16, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, F.; Perlingeiro, R.C.R. Stem cells for skeletal muscle regeneration: Therapeutic potential and roadblocks. Transl. Res. 2014, 163, 409–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sass, F.A.; Fuchs, M.; Pumberger, M.; Geissler, S.; Duda, G.N.; Perka, C.; Schmidt-Bleek, K. Immunology guides skeletal muscle regeneration. Int. J. Mol. Sci. 2018, 19, 835. [Google Scholar] [CrossRef] [Green Version]
- Judson, R.N.; Rossi, F.M.V. Towards stem cell therapies for skeletal muscle repair. NPJ Regen. Med. 2020, 5, 1–6. [Google Scholar] [CrossRef]
- Murphy, M.M.; Lawson, J.A.; Mathew, S.J.; Hutcheson, D.A.; Kardon, G. Satellite cells, connective tissue fibroblasts and their interactions are crucial for muscle regeneration. Development 2011, 138, 3625–3637. [Google Scholar] [CrossRef] [Green Version]
- Czajka, C.A.; Calder, B.W.; Yost, M.J.; Drake, C.J. Implanted scaffold-free prevascularized constructs promote tissue repair. Ann. Plast. Surg. 2015, 74, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montarras, D.; Morgan, J.; Collins, C.; Relaix, F.; Zaffran, S.; Cumano, A.; Partridge, T.; Buckingham, M. Direct isolation of satellite cells for skeletal muscle regeneration. Science 2005, 309, 2064–2067. [Google Scholar] [CrossRef]
- Sacco, A.; Doyonnas, R.; Kraft, P.; Vitorovic, S.; Blau, H.M. Self-renewal and expansion of single transplanted muscle stem cells. Nature 2008, 456, 502–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arpke, R.W.; Darabi, R.; Mader, T.L.; Zhang, Y.; Toyama, A.; Lonetree, C.l.; Nash, N.; Lowe, D.A.; Perlingeiro, R.C.R.; Kyba, M. A New Immuno-, Dystrophin-Deficient Model, the NSG-mdx4Cv Mouse, Provides Evidence for Functional Improvement Following Allogeneic Satellite Cell Transplantation. Stem Cells 2013, 31, 1611–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, J.E.; Watt, D.J.; Sloper, J.C.; Partridge, T.A. Partial correction of an inherited biochemical defect of skeletal muscle by grafts of normal muscle precursor cells. J. Neurol. Sci. 1988, 86, 137–147. [Google Scholar] [CrossRef]
- Watt, D.J.; Lambert, K.; Morgan, J.E.; Partridge, T.A.; Sloper, J.C. Incorporation of donor muscle precursor cells into an area of muscle regeneration in the host mouse. J. Neurol. Sci. 1982, 57, 319–331. [Google Scholar] [CrossRef]
- Partridge, T.A.; Morgan, J.E.; Coulton, G.R.; Hoffman, E.P.; Kunkel, L.M. Conversion of mdx myofibres from dystrophin-negative to-positive by injection of normal myoblasts. Nature 1989, 337, 176–179. [Google Scholar] [CrossRef]
- Kinoshita, I.; Vilquin, J.T.; GuéRette, B.; Asselin, I.; Roy, R.; Tremblay, J.P. Very efficient myoblast allotransplantation in mice under FK506 immunosuppression. Muscle Nerve Off. J. Am. Assoc. Electrodiagn. Med. 1994, 17, 1407–1415. [Google Scholar] [CrossRef]
- Kinoshita, I.; Huard, J.; Tremblay, J.P. Utilization of myoblasts from transgenic mice to evaluate the efficacy of myoblast transplantation. Muscle Nerve Off. J. Am. Assoc. Electrodiagn. Med. 1994, 17, 975–980. [Google Scholar] [CrossRef]
- Huard, J.; Verreault, S.; Roy, R.; Tremblay, M.; Tremblay, J.P. High efficiency of muscle regeneration after human myoblast clone transplantation in SCID mice. J. Clin. Investig. 1994, 93, 586–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, C.A.; Flaibani, M.; Blaauw, B.; Pozzobon, M.; Figallo, E.; Reggiani, C.; Vitiello, L.; Elvassore, N.; De Coppi, P. In vivo tissue engineering of functional skeletal muscle by freshly isolated satellite cells embedded in a photopolymerizable hydrogel. FASEB J. 2011, 25, 2296–2304. [Google Scholar] [CrossRef]
- Prüller, J.; Mannhardt, I.; Eschenhagen, T.; Zammit, P.S.; Figeac, N. Satellite cells delivered in their niche efficiently generate functional myotubes in three-dimensional cell culture. PLoS ONE 2018, 13, e0202574. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, P.M.; Havenstrite, K.L.; Magnusson, K.E.G.; Sacco, A.; Leonardi, N.A.; Kraft, P.; Nguyen, N.K.; Thrun, S.; Lutolf, M.P.; Blau, H.M. Substrate elasticity regulates skeletal muscle stem cell self-renewal in culture. Science 2010, 329, 1078–1081. [Google Scholar] [CrossRef] [Green Version]
- Pavlath, G.K.; Thaloor, D.; Rando, T.A.; Cheong, M.; English, A.W.; Zheng, B. Heterogeneity among muscle precursor cells in adult skeletal muscles with differing regenerative capacities. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1998, 212, 495–508. [Google Scholar] [CrossRef]
- Price, F.D.; Kuroda, K.; Rudnicki, M.A. Stem cell based therapies to treat muscular dystrophy. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2007, 1772, 272–283. [Google Scholar] [CrossRef] [Green Version]
- Shadrach, J.L.; Wagers, A.J. Stem cells for skeletal muscle repair. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2297–2306. [Google Scholar] [CrossRef] [PubMed]
- Péault, B.; Rudnicki, M.; Torrente, Y.; Cossu, G.; Tremblay, J.P.; Partridge, T.; Gussoni, E.; Kunkel, L.M.; Huard, J. Stem and progenitor cells in skeletal muscle development, maintenance, and therapy. Mol. Ther. 2007, 15, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Karpati, G.; Ajdukovic, D.; Arnold, D.; Gledhill, R.B.; Guttmann, R.; Holland, P.; Koch, P.A.; Shoubridge, E.; Spence, D.; Vanasse, M. Myoblast transfer in Duchenne muscular dystrophy. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child. Neurol. Soc. 1993, 34, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Mendell, J.R.; Kissel, J.T.; Amato, A.A.; King, W.; Signore, L.; Prior, T.W.; Sahenk, Z.; Benson, S.; McAndrew, P.E.; Rice, R. Myoblast transfer in the treatment of Duchenne’s muscular dystrophy. N. Engl. J. Med. 1995, 333, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Skuk, D.; Roy, B.; Goulet, M.; Tremblay, J.P. Successful myoblast transplantation in primates depends on appropriate cell delivery and induction of regeneration in the host muscle. Exp. Neurol. 1999, 155, 22–30. [Google Scholar] [CrossRef]
- Hill, E.; Boontheekul, T.; Mooney, D.J. Designing scaffolds to enhance transplanted myoblast survival and migration. Tissue Eng. 2006, 12, 1295–1304. [Google Scholar] [CrossRef] [Green Version]
- Machingal, M.A.; Corona, B.T.; Walters, T.J.; Kesireddy, V.; Koval, C.N.; Dannahower, A.; Zhao, W.; Yoo, J.J.; Christ, G.J. A tissue-engineered muscle repair construct for functional restoration of an irrecoverable muscle injury in a murine model. Tissue Eng. Part A 2011, 17, 2291–2303. [Google Scholar] [CrossRef] [Green Version]
- Corona, B.T.; Machingal, M.A.; Criswell, T.; Vadhavkar, M.; Dannahower, A.C.; Bergman, C.; Zhao, W.; Christ, G.J. Further development of a tissue engineered muscle repair construct in vitro for enhanced functional recovery following implantation in vivo in a murine model of volumetric muscle loss injury. Tissue Eng. Part A 2012, 18, 1213–1228. [Google Scholar] [CrossRef] [Green Version]
- Corona, B.T.; Ward, C.L.; Baker, H.B.; Walters, T.J.; Christ, G.J. Implantation of in vitro tissue engineered muscle repair constructs and bladder acellular matrices partially restore in vivo skeletal muscle function in a rat model of volumetric muscle loss injury. Tissue Eng. Part A 2014, 20, 705–715. [Google Scholar]
- Nakamura, Y.; Miyaki, S.; Ishitobi, H.; Matsuyama, S.; Nakasa, T.; Kamei, N.; Akimoto, T.; Higashi, Y.; Ochi, M. Mesenchymal-stem-cell-derived exosomes accelerate skeletal muscle regeneration. FEBS Lett. 2015, 589, 1257–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A.I. Mesenchymal stem cells: Cell–based reconstructive therapy in orthopedics. Tissue Eng. 2005, 11, 1198–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dezawa, M.; Ishikawa, H.; Itokazu, Y.; Yoshihara, T.; Hoshino, M.; Takeda, S.; Ide, C.; Nabeshima, Y. Bone marrow stromal cells generate muscle cells and repair muscle degeneration. Science 2005, 309, 314–317. [Google Scholar] [CrossRef] [PubMed]
- Krampera, M.; Pizzolo, G.; Aprili, G.; Franchini, M. Mesenchymal stem cells for bone, cartilage, tendon and skeletal muscle repair. Bone 2006, 39, 678–683. [Google Scholar] [CrossRef]
- Kern, S.; Eichler, H.; Stoeve, J.; Klüter, H.; Bieback, K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef]
- Mara, C.S.d.; Sartori, A.R.; Duarte, A.S.; Andrade, A.L.L.; Pedro, M.A.C.; Coimbra, I.B. Periosteum as a source of mesenchymal stem cells: The effects of TGF-β3 on chondrogenesis. Clinics 2011, 66, 487–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.S.; Hung, S.C.; Peng, S.T.; Huang, C.C.; Wei, H.M.; Guo, Y.J.; Fu, Y.S.; Lai, M.C.; Chen, C.C. Mesenchymal stem cells in the Wharton’s jelly of the human umbilical cord. Stem Cells 2004, 22, 1330–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, S.A.; Roobrouck, V.D.; Verfaillie, C.M.; Van Gool, S.W. Immunological characteristics of human mesenchymal stem cells and multipotent adult progenitor cells. Immunol. Cell Biol. 2013, 91, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Ankrum, J.A.; Ong, J.F.; Karp, J.M. Mesenchymal stem cells: Immune evasive, not immune privileged. Nat. Biotechnol. 2014, 32, 252. [Google Scholar] [CrossRef] [Green Version]
- Galli, D.; Vitale, M.; Vaccarezza, M. Bone marrow-derived mesenchymal cell differentiation toward myogenic lineages: Facts and perspectives. Biomed. Res. Int. 2014, 2014, 762695. [Google Scholar] [CrossRef] [Green Version]
- Sicari, B.M.; Dearth, C.L.; Badylak, S.F. Tissue engineering and regenerative medicine approaches to enhance the functional response to skeletal muscle injury. Anat. Rec. 2014, 297, 51–64. [Google Scholar] [CrossRef]
- Ferrari, G.; Angelis, D.; Coletta, M.; Paolucci, E.; Stornaiuolo, A.; Cossu, G.; Mavilio, F. Muscle regeneration by bone marrow-derived myogenic progenitors. Science 1998, 279, 1528–1530. [Google Scholar] [CrossRef]
- Feng, S.W.; Lu, X.L.; Liu, Z.S.; Zhang, Y.N.; Liu, T.Y.; Li, J.L.; Yu, M.J.; Zeng, Y.; Zhang, C. Dynamic distribution of bone marrow-derived mesenchymal stromal cells and change of pathology after infusing into mdx mice. Cytotherapy 2008, 10, 254–264. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, Z.; Terry, T.; McNatt, J.M.; Willerson, J.T.; Zoldhelyi, P. Intra-arterial transplantation of adult bone marrow cells restores blood flow and regenerates skeletal muscle in ischemic limbs. Vasc. Endovasc. Surg. 2009, 43, 433–443. [Google Scholar] [CrossRef]
- Andrade, B.M.; Baldanza, M.R.; Ribeiro, K.C.; Porto, A.; Pecanha, R.; Fortes, F.S.A.; Zapata-Sudo, G.; Campos-de-Carvalho, A.C.; Goldenberg, R.C.S.; Werneck-de-Castro, J.P. Bone marrow mesenchymal cells improve muscle function in a skeletal muscle re-injury model. PLoS ONE 2015, 10, e0127561. [Google Scholar] [CrossRef]
- Helal, M.A.M.; Shaheen, N.E.M.; Abu Zahra, F.A. Immunomodulatory capacity of the local mesenchymal stem cells transplantation after severe skeletal muscle injury in female rats. Immunopharmacol. Immunotoxicol. 2016, 38, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Winkler, T.; von Roth, P.; Matziolis, G.; Mehta, M.; Perka, C.; Duda, G.N. Dose–response relationship of mesenchymal stem cell transplantation and functional regeneration after severe skeletal muscle injury in rats. Tissue Eng. Part A 2009, 15, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Yonemochi, Y.; Nakajyo, Y.; Hidaka, H.; Ikeda, T.; Ando, Y. CXCL12 and osteopontin from bone marrow-derived mesenchymal stromal cells improve muscle regeneration. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sassoli, C.; Vallone, L.; Tani, A.; Chellini, F.; Nosi, D.; Zecchi-Orlandini, S. Combined use of bone marrow-derived mesenchymal stromal cells (BM-MSCs) and platelet rich plasma (PRP) stimulates proliferation and differentiation of myoblasts in vitro: New therapeutic perspectives for skeletal muscle repair/regeneration. Cell Tissue Res. 2018, 372, 549–570. [Google Scholar] [CrossRef] [PubMed]
- Natsu, K.; Ochi, M.; Mochizuki, Y.; Hachisuka, H.; Yanada, S.; Yasunaga, Y. Allogeneic bone marrow-derived mesenchymal stromal cells promote the regeneration of injured skeletal muscle without differentiation into myofibers. Tissue Eng. 2004, 10, 1093–1112. [Google Scholar] [CrossRef] [PubMed]
- Merritt, E.K.; Cannon, M.V.; Hammers, D.W.; Le, L.N.; Gokhale, R.; Sarathy, A.; Song, T.J.; Tierney, M.T.; Suggs, L.J.; Walters, T.J. Repair of traumatic skeletal muscle injury with bone-marrow-derived mesenchymal stem cells seeded on extracellular matrix. Tissue Eng. Part A 2010, 16, 2871–2881. [Google Scholar] [CrossRef] [Green Version]
- Kozlowska, U.; Krawczenko, A.; Futoma, K.; Jurek, T.; Rorat, M.; Patrzalek, D.; Klimczak, A. Similarities and differences between mesenchymal stem/progenitor cells derived from various human tissues. World J. Stem Cells 2019, 11, 347. [Google Scholar] [CrossRef]
- Huri, P.Y.; Wang, A.; Spector, A.A.; Grayson, W.L. Multistage adipose-derived stem cell myogenesis: An experimental and modeling study. Cell. Mol. Bioeng. 2014, 7, 497–509. [Google Scholar] [CrossRef]
- Di Rocco, G.; Iachininoto, M.G.; Tritarelli, A.; Straino, S.; Zacheo, A.; Germani, A.; Crea, F.; Capogrossi, M.C. Myogenic potential of adipose-tissue-derived cells. J. Cell Sci. 2006, 119, 2945–2952. [Google Scholar] [CrossRef] [Green Version]
- da Justa Pinheiro, C.H.; de Queiroz, J.C.F.; Guimarães-Ferreira, L.; Vitzel, K.F.; Nachbar, R.T.; de Sousa, L.G.O.; de Souza-Jr, A.L.; Nunes, M.T.; Curi, R. Local injections of adipose-derived mesenchymal stem cells modulate inflammation and increase angiogenesis ameliorating the dystrophic phenotype in dystrophin-deficient skeletal muscle. Stem Cell Rev. Rep. 2012, 8, 363–374. [Google Scholar] [CrossRef]
- Kesireddy, V. Evaluation of adipose-derived stem cells for tissue-engineered muscle repair construct-mediated repair of a murine model of volumetric muscle loss injury. Int. J. Nanomed. 2016, 11, 1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meligy, F.Y.; Shigemura, K.; Behnsawy, H.M.; Fujisawa, M.; Kawabata, M.; Shirakawa, T. The efficiency of in vitro isolation and myogenic differentiation of MSCs derived from adipose connective tissue, bone marrow, and skeletal muscle tissue. Vitr. Cell. Dev. Biol. Anim. 2012, 48, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Zheng, B.; Jankowski, R.J.; Kimura, S.; Ikezawa, M.; Deasy, B.; Cummins, J.; Epperly, M.; Qu-Petersen, Z.; Huard, J. Muscle stem cells differentiate into haematopoietic lineages but retain myogenic potential. Nat. Cell Biol. 2003, 5, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Qu-Petersen, Z.; Cao, B.; Kimura, S.; Jankowski, R.; Cummins, J.; Usas, A.; Gates, C.; Robbins, P.; Wernig, A. Clonal isolation of muscle-derived cells capable of enhancing muscle regeneration and bone healing. J. Cell Biol. 2000, 150, 1085–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhagavati, S. Stem cell based therapy for skeletal muscle diseases. Curr. Stem Cell Res. Ther. 2008, 3, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Matthias, N.; Hunt, S.D.; Wu, J.; Lo, J.; Callahan, L.A.S.; Li, Y.; Huard, J.; Darabi, R. Volumetric muscle loss injury repair using in situ fibrin gel cast seeded with muscle-derived stem cells (MDSCs). Stem Cell Res. 2018, 27, 65–73. [Google Scholar] [CrossRef]
- Usas, A.; Huard, J. Muscle-derived stem cells for tissue engineering and regenerative therapy. Biomaterials 2007, 28, 5401–5406. [Google Scholar] [CrossRef] [Green Version]
- Deasy, B.M.; Jankowski, R.J.; Huard, J. Muscle-derived stem cells: Characterization and potential for cell-mediated therapy. Blood Cells Mol. Dis. 2001, 27, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Torrente, Y.; Tremblay, J.P.; Pisati, F.; Belicchi, M.; Rossi, B.; Sironi, M.; Fortunato, F.; El Fahime, M.; D’Angelo, M.G.; Caron, N.J.; et al. Intraarterial injection of muscle-derived CD34+Sca-1+ stem cells restores dystrophin in mdx mice. J. Cell Biol. 2001, 152, 335–348. [Google Scholar] [CrossRef] [Green Version]
- Qu-Petersen, Z.; Deasy, B.; Jankowski, R.; Ikezawa, M.; Cummins, J.; Pruchnic, R.; Mytinger, J.; Cao, B.; Gates, C.; Wernig, A. Identification of a novel population of muscle stem cells in mice potential for muscle regeneration. J. Cell Biol. 2002, 157, 851–864. [Google Scholar] [CrossRef]
- Rouger, K.; Larcher, T.; Dubreil, L.; Deschamps, J.-Y.; Le Guiner, C.; Jouvion, G.; Delorme, B.; Lieubeau, B.; Carlus, M.; Fornasari, B. Systemic delivery of allogenic muscle stem cells induces long-term muscle repair and clinical efficacy in duchenne muscular dystrophy dogs. Am. J. Pathol. 2011, 179, 2501–2518. [Google Scholar] [CrossRef] [PubMed]
- Lorant, J.; Saury, C.; Schleder, C.; Robriquet, F.; Lieubeau, B.; Négroni, E.; Leroux, I.; Chabrand, L.; Viau, S.; Babarit, C. Skeletal muscle regenerative potential of human MuStem cells following transplantation into injured mice muscle. Mol. Ther. 2018, 26, 618–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skuk, D.; Goulet, M.; Roy, B.; Piette, V.; Côté, C.H.; Chapdelaine, P.; Hogrel, J.-Y.; Paradis, M.; Bouchard, J.-P.; Sylvain, M. First test of a “high-density injection” protocol for myogenic cell transplantation throughout large volumes of muscles in a Duchenne muscular dystrophy patient: Eighteen months follow-up. Neuromuscul. Disord. 2007, 17, 38–46. [Google Scholar] [CrossRef]
- Klimczak, A.; Zimna, A.; Malcher, A.; Kozlowska, U.; Futoma, K.; Czarnota, J.; Kemnitz, P.; Bryl, A.; Kurpisz, M. Co-Transplantation of Bone Marrow-MSCs and Myogenic Stem/Progenitor Cells from Adult Donors Improves Muscle Function of Patients with Duchenne Muscular Dystrophy. Cells 2020, 9, 1119. [Google Scholar] [CrossRef] [PubMed]
- Farini, A.; Razini, P.; Erratico, S.; Torrente, Y.; Meregalli, M. Cell based therapy for Duchenne muscular dystrophy. J. Cell. Physiol. 2009, 221, 526–534. [Google Scholar] [CrossRef]
- Dellavalle, A.; Sampaolesi, M.; Tonlorenzi, R.; Tagliafico, E.; Sacchetti, B.; Perani, L.; Innocenzi, A.; Galvez, B.G.; Messina, G.; Morosetti, R. Pericytes of human skeletal muscle are myogenic precursors distinct from satellite cells. Nat. Cell Biol. 2007, 9, 255–267. [Google Scholar] [CrossRef] [Green Version]
- Birbrair, A.; Delbono, O. Pericytes are essential for skeletal muscle formation. Stem Cell Rev. Rep. 2015, 11, 547–548. [Google Scholar] [CrossRef]
- Dellavalle, A.; Maroli, G.; Covarello, D.; Azzoni, E.; Innocenzi, A.; Perani, L.; Antonini, S.; Sambasivan, R.; Brunelli, S.; Tajbakhsh, S. Pericytes resident in postnatal skeletal muscle differentiate into muscle fibres and generate satellite cells. Nat. Commun. 2011, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Birbrair, A.; Zhang, T.; Wang, Z.-M.; Messi, M.L.; Enikolopov, G.N.; Mintz, A.; Delbono, O. Role of pericytes in skeletal muscle regeneration and fat accumulation. Stem Cells Dev. 2013, 22, 2298–2314. [Google Scholar] [CrossRef] [Green Version]
- Fuoco, C.; Sangalli, E.; Vono, R.; Testa, S.; Sacchetti, B.; Latronico, M.V.; Bernardini, S.; Madeddu, P.; Cesareni, G.; Seliktar, D. 3D hydrogel environment rejuvenates aged pericytes for skeletal muscle tissue engineering. Front. Physiol. 2014, 5, 203. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [Green Version]
- van der Wal, E.; Herrero-Hernandez, P.; Wan, R.; Broeders, M.; In’t Groen, S.L.M.; van Gestel, T.J.M.; van Ijcken, W.F.J.; Cheung, T.H.; van der Ploeg, A.T.; Schaaf, G.J. Large-scale expansion of human iPSC-derived skeletal muscle cells for disease modeling and cell-based therapeutic strategies. Stem Cell Rep. 2018, 10, 1975–1990. [Google Scholar] [CrossRef]
- Steinle, H.; Weber, M.; Behring, A.; Mau-Holzmann, U.; von Ohle, C.; Popov, A.F.; Schlensak, C.; Wendel, H.P.; Avci-Adali, M. Reprogramming of Urine-Derived Renal Epithelial Cells into iPSCs Using srRNA and Consecutive Differentiation into Beating Cardiomyocytes. Mol. Nucleic Acids 2019, 17, 907–921. [Google Scholar] [CrossRef] [Green Version]
- Del Carmen Ortuno-Costela, M.; Garcia-Lopez, M.; Cerrada, V.; Gallardo, M.E. iPSCs: A powerful tool for skeletal muscle tissue engineering. J. Cell Mol. Med. 2019, 23, 3784–3794. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, U.; Benvenisty, N. The tumorigenicity of human embryonic and induced pluripotent stem cells. Nat. Rev. Cancer 2011, 11, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Roca, I.; Requena, J.; Edel, M.J.; Alvarez-Palomo, A.B. Myogenic precursors from iPS cells for skeletal muscle cell replacement therapy. J. Clin. Med. 2015, 4, 243–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, I.Y.; Lim, H.; Estrellas, K.; Mula, J.; Cohen, T.V.; Zhang, Y.; Donnelly, C.J.; Richard, J.-P.; Kim, Y.J.; Kim, H. Concordant but varied phenotypes among Duchenne muscular dystrophy patient-specific myoblasts derived using a human iPSC-based model. Cell Rep. 2016, 15, 2301–2312. [Google Scholar] [CrossRef] [Green Version]
- Chal, J.; Al Tanoury, Z.; Hestin, M.; Gobert, B.; Aivio, S.; Hick, A.; Cherrier, T.; Nesmith, A.P.; Parker, K.K.; Pourquié, O. Generation of human muscle fibers and satellite-like cells from human pluripotent stem cells in vitro. Nat. Protoc. 2016, 11, 1833–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goudenege, S.; Lebel, C.; Huot, N.B.; Dufour, C.; Fujii, I.; Gekas, J.; Rousseau, J.; Tremblay, J.P. Myoblasts derived from normal hESCs and dystrophic hiPSCs efficiently fuse with existing muscle fibers following transplantation. Mol. Ther. 2012, 20, 2153–2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, Y.; Chang, H.; Umeda, K.; Niwa, A.; Iwasa, T.; Awaya, T.; Fukada, S.i.; Yamamoto, H.; Yamanaka, S.; Nakahata, T. Generation of skeletal muscle stem/progenitor cells from murine induced pluripotent stem cells. FASEB J. 2010, 24, 2245–2253. [Google Scholar] [CrossRef]
- Rao, L.; Qian, Y.; Khodabukus, A.; Ribar, T.; Bursac, N. Engineering human pluripotent stem cells into a functional skeletal muscle tissue. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Maffioletti, S.M.; Sarcar, S.; Henderson, A.B.H.; Mannhardt, I.; Pinton, L.; Moyle, L.A.; Steele-Stallard, H.; Cappellari, O.; Wells, K.E.; Ferrari, G. Three-dimensional human iPSC-derived artificial skeletal muscles model muscular dystrophies and enable multilineage tissue engineering. Cell Rep. 2018, 23, 899–908. [Google Scholar] [CrossRef] [Green Version]
- Darabi, R.; Pan, W.; Bosnakovski, D.; Baik, J.; Kyba, M.; Perlingeiro, R.C.R. Functional myogenic engraftment from mouse iPS cells. Stem Cell Rev. Rep. 2011, 7, 948–957. [Google Scholar] [CrossRef] [Green Version]
- Darabi, R.; Arpke, R.W.; Irion, S.; Dimos, J.T.; Grskovic, M.; Kyba, M.; Perlingeiro, R.C.R. Human ES-and iPS-derived myogenic progenitors restore DYSTROPHIN and improve contractility upon transplantation in dystrophic mice. Cell Stem Cell 2012, 10, 610–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoji, E.; Woltjen, K.; Sakurai, H. Directed myogenic differentiation of human induced pluripotent stem cells. Methods Mol. Biol. 2016, 1353, 89–99. [Google Scholar] [PubMed]
- Hosoyama, T.; McGivern, J.V.; Van Dyke, J.M.; Ebert, A.D.; Suzuki, M. Derivation of myogenic progenitors directly from human pluripotent stem cells using a sphere-based culture. Stem Cells Transl. Med. 2014, 3, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Jiwlawat, S.; Lynch, E.; Glaser, J.; Smit-Oistad, I.; Jeffrey, J.; Van Dyke, J.M.; Suzuki, M. Differentiation and sarcomere formation in skeletal myocytes directly prepared from human induced pluripotent stem cells using a sphere-based culture. Differentiation 2017, 96, 70–81. [Google Scholar] [CrossRef]
- Borchin, B.; Chen, J.; Barberi, T. Derivation and FACS-mediated purification of PAX3+/PAX7+ skeletal muscle precursors from human pluripotent stem cells. Stem Cell Rep. 2013, 1, 620–631. [Google Scholar] [CrossRef] [Green Version]
- van der Wal, E.; Bergsma, A.J.; Van Gestel, T.J.M.; Lm, S.; Zaehres, H.; Araúzo-Bravo, M.J.; Schöler, H.R.; van der Ploeg, A.T.; Pijnappel, W.W.M.P. GAA deficiency in Pompe disease is alleviated by exon inclusion in iPSC-derived skeletal muscle cells. Mol. Ther. Nucleic Acids 2017, 7, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Baci, D.; Chirivì, M.; Pace, V.; Maiullari, F.; Milan, M.; Rampin, A.; Somma, P.; Presutti, D.; Garavelli, S.; Bruno, A. Extracellular Vesicles from Skeletal Muscle Cells Efficiently Promote Myogenesis in Induced Pluripotent Stem Cells. Cells 2020, 9, 1527. [Google Scholar] [CrossRef]
- Zhou, L.; Ge, J.; Wang, M.; Chen, M.; Cheng, W.; Ji, W.; Lei, B. Injectable muscle-adhesive antioxidant conductive photothermal bioactive nanomatrix for efficiently promoting full-thickness skeletal muscle regeneration. Bioact. Mater. 2020, 6, 1605–1617. [Google Scholar] [CrossRef]
- Lev, R.; Seliktar, D. Hydrogel biomaterials and their therapeutic potential for muscle injuries and muscular dystrophies. J. R. Soc. Interface 2018, 15, 20170380. [Google Scholar] [CrossRef]
- Mei, X.; Cheng, K. Recent Development in Therapeutic Cardiac Patches. Front. Cardiovasc. Med. 2020, 7, 294. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.H.; Shayan, M.; Huang, N.F. Engineering biomimetic materials for skeletal muscle repair and regeneration. Adv. Healthc. Mater. 2019, 8, 1801168. [Google Scholar] [CrossRef]
- Fischer, K.M.; Scott, T.E.; Browe, D.P.; McGaughey, T.A.; Wood, C.; Wolyniak, M.J.; Freeman, J.W. Hydrogels for Skeletal Muscle Regeneration. Regen. Eng. Transl. Med. 2020, 1–9. [Google Scholar] [CrossRef]
- Camci-Unal, G.; Annabi, N.; Dokmeci, M.R.; Liao, R.; Khademhosseini, A. Hydrogels for cardiac tissue engineering. NPG Asia Mater. 2014, 6, e99. [Google Scholar] [CrossRef]
- McLaughlin, S.; McNeill, B.; Podrebarac, J.; Hosoyama, K.; Sedlakova, V.; Cron, G.; Smyth, D.; Seymour, R.; Goel, K.; Liang, W. Injectable human recombinant collagen matrices limit adverse remodeling and improve cardiac function after myocardial infarction. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Feng, M.; Liu, X.; Hou, X.; Chen, J.; Zhang, H.; Song, S.; Han, X.; Shi, C. Specific angiogenic peptide binding with injectable cardiac ECM collagen gel promotes the recovery of myocardial infarction in rat. J. Biomed. Mater. Res. Part. A 2020, 108, 1881–1889. [Google Scholar] [CrossRef]
- Le, L.V.; Mohindra, P.; Fang, Q.; Sievers, R.E.; Mkrtschjan, M.A.; Solis, C.; Safranek, C.W.; Russell, B.; Lee, R.J.; Desai, T.A. Injectable hyaluronic acid based microrods provide local micromechanical and biochemical cues to attenuate cardiac fibrosis after myocardial infarction. Biomaterials 2018, 169, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Stilhano, R.S.; Madrigal, J.L.; Wong, K.; Williams, P.A.; Martin, P.K.; Yamaguchi, F.S.; Samoto, V.Y.; Han, S.W.; Silva, E.A. Injectable alginate hydrogel for enhanced spatiotemporal control of lentivector delivery in murine skeletal muscle. J. Control. Release 2016, 237, 42–49. [Google Scholar] [CrossRef]
- Fang, R.; Tian, W.; Chen, X. Synthesis of injectable alginate hydrogels with muscle-derived stem cells for potential myocardial infarction repair. Appl. Sci. 2017, 7, 252. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Wu, Y.; Chen, W.; Li, J.; Wang, X.; Chen, Y.; Yu, Y.; Shen, Z.; Zhang, Y. Sustained release of bioactive IGF-1 from a silk fibroin microsphere-based injectable alginate hydrogel for the treatment of myocardial infarction. J. Mater. Chem. B 2020, 8, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, H.H.; Zhou, H.; Weir, M.D.; Chen, Q.; Trotman, C.A. Human umbilical cord stem cell encapsulation in novel macroporous and injectable fibrin for muscle tissue engineering. Acta Biomater. 2013, 9, 4688–4697. [Google Scholar] [CrossRef] [Green Version]
- Melly, L.; Grosso, A.; Stanciu Pop, C.; Yu-Hsuan, C.; Nollevaux, M.C.; Schachtrup, C.; Marsano, A.; Di Maggio, N.; Rondelet, B.; Banfi, A. Fibrin hydrogels promote scar formation and prevent therapeutic angiogenesis in the heart. J. Tissue Eng. Regen. Med. 2020, 14, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Qu, J.; Zhao, X.; Zhang, M. Degradable conductive self-healing hydrogels based on dextran-graft-tetraaniline and N-carboxyethyl chitosan as injectable carriers for myoblast cell therapy and muscle regeneration. Acta Biomater. 2019, 84, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Pollot, B.E.; Rathbone, C.R.; Wenke, J.C.; Guda, T. Natural polymeric hydrogel evaluation for skeletal muscle tissue engineering. J. Biomed. Mater. Res. Part B Appl. Biomater. 2018, 106, 672–679. [Google Scholar] [CrossRef]
- Boso, D.; Maghin, E.; Carraro, E.; Giagante, M.; Pavan, P.; Piccoli, M. Extracellular matrix-derived hydrogels as biomaterial for different skeletal muscle tissue replacements. Materials 2020, 13, 2483. [Google Scholar] [CrossRef]
- Theus, A.S.; Tomov, M.L.; Cetnar, A.; Lima, B.; Nish, J.; McCoy, K.; Mahmoudi, M.; Serpooshan, V. Biomaterial approaches for cardiovascular tissue engineering. Emergent Mater. 2019, 2, 193–207. [Google Scholar] [CrossRef]
- Tashakori-Miyanroudi, M.; Rakhshan, K.; Ramez, M.; Asgarian, S.; Janzadeh, A.; Azizi, Y.; Seifalian, A.; Ramezani, F. Conductive carbon nanofibers incorporated into collagen bio-scaffold assists myocardial injury repair. Int. J. Biol. Macromol. 2020, 163, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Pagliarosi, O.; Picchio, V.; Chimenti, I.; Messina, E.; Gaetani, R. Building an artificial cardiac microenvironment: A focus on the extracellular matrix. Front. Cell Dev. Biol. 2020, 8, 919. [Google Scholar] [CrossRef] [PubMed]
- Mao, M.; He, J.; Li, Z.; Han, K.; Li, D. Multi-directional cellular alignment in 3D guided by electrohydrodynamically-printed microlattices. Acta Biomater. 2020, 101, 141–151. [Google Scholar] [CrossRef]
- Li, M.-T.; Ruehle, M.A.; Stevens, H.Y.; Servies, N.; Willett, N.J.; Karthikeyakannan, S.; Warren, G.L.; Guldberg, R.E.; Krishnan, L. Skeletal myoblast-seeded vascularized tissue scaffolds in the treatment of a large volumetric muscle defect in the rat biceps femoris muscle. Tissue Eng. Part A 2017, 23, 989–1000. [Google Scholar] [CrossRef]
- Gupta, D.; Santoso, J.W.; McCain, M.L. Characterization of Gelatin Hydrogels Cross-Linked with Microbial Transglutaminase as Engineered Skeletal Muscle Substrates. Bioengineering 2021, 8, 6. [Google Scholar] [CrossRef]
- Bettadapur, A.; Suh, G.C.; Geisse, N.A.; Wang, E.R.; Hua, C.; Huber, H.A.; Viscio, A.A.; Kim, J.Y.; Strickland, J.B.; McCain, M.L. Prolonged Culture of Aligned Skeletal Myotubes on Micromolded Gelatin Hydrogels. Sci. Rep. 2016, 6, 28855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wu, Y.; Guo, B.; Ma, P.X. Nanofiber yarn/hydrogel core–shell scaffolds mimicking native skeletal muscle tissue for guiding 3D myoblast alignment, elongation, and differentiation. ACS Nano 2015, 9, 9167–9179. [Google Scholar] [CrossRef] [PubMed]
- Broer, T.; Khodabukus, A.; Bursac, N. Can we mimic skeletal muscles for novel drug discovery? Expert Opin. Drug Discov. 2020, 15, 643–645. [Google Scholar] [CrossRef] [Green Version]
- Marcinczyk, M.; Elmashhady, H.; Talovic, M.; Dunn, A.; Bugis, F.; Garg, K. Laminin-111 enriched fibrin hydrogels for skeletal muscle regeneration. Biomaterials 2017, 141, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Gholobova, D.; Terrie, L.; Gerard, M.; Declercq, H.; Thorrez, L. Vascularization of tissue-engineered skeletal muscle constructs. Biomaterials 2020, 235, 119708. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Forsythe, S.; He, Y.; Liu, Q.; Xiong, G.; Wei, S.; Li, G.; Atala, A.; Skardal, A.; Zhang, Y. Tissue-specific extracellular matrix promotes myogenic differentiation of human muscle progenitor cells on gelatin and heparin conjugated alginate hydrogels. Acta Biomater. 2017, 62, 222–233. [Google Scholar] [CrossRef]
- Majid, Q.A.; Fricker, A.T.; Gregory, D.A.; Davidenko, N.; Hernandez Cruz, O.; Jabbour, R.J.; Owen, T.J.; Basnett, P.; Lukasiewicz, B.; Stevens, M. Natural Biomaterials for Cardiac Tissue Engineering: A Highly Biocompatible Solution. Front. Cardiovasc. Med. 2020, 7, 192. [Google Scholar] [CrossRef]
- Ansari, S.; Chen, C.; Xu, X.; Annabi, N.; Zadeh, H.H.; Wu, B.M.; Khademhosseini, A.; Shi, S.; Moshaverinia, A. Muscle tissue engineering using gingival mesenchymal stem cells encapsulated in alginate hydrogels containing multiple growth factors. Ann. Biomed. Eng. 2016, 44, 1908–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwee, B.J.; Mooney, D.J. Biomaterials for skeletal muscle tissue engineering. Curr. Opin. Biotechnol. 2017, 47, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Boontheekul, T.; Hill, E.E.; Kong, H.-J.; Mooney, D.J. Regulating myoblast phenotype through controlled gel stiffness and degradation. Tissue Eng. 2007, 13, 1431–1442. [Google Scholar] [CrossRef] [Green Version]
- Yeo, M.; Lee, H.; Kim, G.H. Combining a micro/nano-hierarchical scaffold with cell-printing of myoblasts induces cell alignment and differentiation favorable to skeletal muscle tissue regeneration. Biofabrication 2016, 8, 035021. [Google Scholar] [CrossRef] [PubMed]
- Rico, P.; Rodrigo-Navarro, A.; Salmerón-Sánchez, M. Borax-loaded PLLA for promotion of myogenic differentiation. Tissue Eng. Part A 2015, 21, 2662–2672. [Google Scholar] [CrossRef] [PubMed]
- Levenberg, S.; Rouwkema, J.; Macdonald, M.; Garfein, E.S.; Kohane, D.S.; Darland, D.C.; Marini, R.; Van Blitterswijk, C.A.; Mulligan, R.C.; D’Amore, P.A. Engineering vascularized skeletal muscle tissue. Nat. Biotechnol. 2005, 23, 879–884. [Google Scholar] [CrossRef]
- Wolf, M.T.; Dearth, C.L.; Sonnenberg, S.B.; Loboa, E.G.; Badylak, S.F. Naturally derived and synthetic scaffolds for skeletal muscle reconstruction. Adv. Drug Deliv. Rev. 2015, 84, 208–221. [Google Scholar] [CrossRef] [Green Version]
- Ostrovidov, S.; Salehi, S.; Costantini, M.; Suthiwanich, K.; Ebrahimi, M.; Sadeghian, R.B.; Fujie, T.; Shi, X.; Cannata, S.; Gargioli, C. 3D bioprinting in skeletal muscle tissue engineering. Small 2019, 15, 1805530. [Google Scholar] [CrossRef]
- Apsite, I.; Uribe, J.M.; Posada, A.F.; Rosenfeldt, S.; Salehi, S.; Ionov, L. 4D biofabrication of skeletal muscle microtissues. Biofabrication 2019, 12, 015016. [Google Scholar] [CrossRef]
- Kook, Y.-M.; Hwang, S.; Kim, H.; Rhee, K.-J.; Lee, K.; Koh, W.-G. Cardiovascular tissue regeneration system based on multiscale scaffolds comprising double-layered hydrogels and fibers. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Salimath, A.S.; García, A.J. Biofunctional hydrogels for skeletal muscle constructs. J. Tissue Eng. Regen. Med. 2016, 10, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Han, W.M.; Mohiuddin, M.; Anderson, S.E.; García, A.J.; Jang, Y.C. Co-delivery of Wnt7a and muscle stem cells using synthetic bioadhesive hydrogel enhances murine muscle regeneration and cell migration during engraftment. Acta Biomater. 2019, 94, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Han, W.M.; Anderson, S.E.; Mohiuddin, M.; Barros, D.; Nakhai, S.A.; Shin, E.; Amaral, I.F.; Pêgo, A.P.; García, A.J.; Jang, Y.C. Synthetic matrix enhances transplanted satellite cell engraftment in dystrophic and aged skeletal muscle with comorbid trauma. Sci. Adv. 2018, 4, eaar4008. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Wang, L.; Guo, B.; Wang, Z.; Chen, Y.E.; Ma, P.X. Ductile electroactive biodegradable hyperbranched polylactide copolymers enhancing myoblast differentiation. Biomaterials 2015, 71, 158–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riboldi, S.A.; Sampaolesi, M.; Neuenschwander, P.; Cossu, G.; Mantero, S. Electrospun degradable polyesterurethane membranes: Potential scaffolds for skeletal muscle tissue engineering. Biomaterials 2005, 26, 4606–4615. [Google Scholar] [CrossRef] [Green Version]
- Naureen, B.; Haseeb, A.; Basirun, W.; Muhamad, F. Recent advances in tissue engineering scaffolds based on polyurethane and modified polyurethane. Mater. Sci. Eng. C 2021, 118, 111228. [Google Scholar] [CrossRef]
- Ergene, E.; Yagci, B.S.; Gokyer, S.; Eyidogan, A.; Aksoy, E.A.; Huri, P.Y. A novel polyurethane-based biodegradable elastomer as a promising material for skeletal muscle tissue engineering. Biomed. Mater. 2019, 14, 025014. [Google Scholar] [CrossRef]
- Jamadi, E.S.; Ghasemi-Mobarakeh, L.; Morshed, M.; Sadeghi, M.; Prabhakaran, M.P.; Ramakrishna, S. Synthesis of polyester urethane urea and fabrication of elastomeric nanofibrous scaffolds for myocardial regeneration. Mater. Sci. Eng. C 2016, 63, 106–116. [Google Scholar] [CrossRef]
- Vannozzi, L.; Ricotti, L.; Santaniello, T.; Terencio, T.; Oropesa-Nunez, R.; Canale, C.; Borghi, F.; Menciassi, A.; Lenardi, C.; Gerges, I. 3D porous polyurethanes featured by different mechanical properties: Characterization and interaction with skeletal muscle cells. J. Mech. Behav. Biomed. Mater. 2017, 75, 147–159. [Google Scholar] [CrossRef]
- Sun, Y.; Duffy, R.; Lee, A.; Feinberg, A.W. Optimizing the structure and contractility of engineered skeletal muscle thin films. Acta Biomater. 2013, 9, 7885–7894. [Google Scholar] [CrossRef]
- Browe, D.P.; Wood, C.; Sze, M.T.; White, K.A.; Scott, T.; Olabisi, R.M.; Freeman, J.W. Characterization and optimization of actuating poly (ethylene glycol) diacrylate/acrylic acid hydrogels as artificial muscles. Polymer 2017, 117, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Jo, H.; Sim, M.; Kim, S.; Yang, S.; Yoo, Y.; Park, J.-H.; Yoon, T.H.; Kim, M.-G.; Lee, J.Y. Electrically conductive graphene/polyacrylamide hydrogels produced by mild chemical reduction for enhanced myoblast growth and differentiation. Acta Biomater. 2017, 48, 100–109. [Google Scholar] [CrossRef]
- Hosseinzadeh, S.; Rezayat, S.M.; Giaseddin, A.; Aliyan, A.; Soleimani, M. Microfluidic system for synthesis of nanofibrous conductive hydrogel and muscle differentiation. J. Biomater. Appl. 2018, 32, 853–861. [Google Scholar] [CrossRef]
- Jun, I.; Jeong, S.; Shin, H. The stimulation of myoblast differentiation by electrically conductive sub-micron fibers. Biomaterials 2009, 30, 2038–2047. [Google Scholar] [CrossRef]
- Fallahi, A.; Yazdi, I.K.; Serex, L.; Lesha, E.; Faramarzi, N.; Tarlan, F.; Avci, H.; Costa-Almeida, R.; Sharifi, F.; Rinoldi, C. Customizable composite fibers for engineering skeletal muscle models. Acs Biomater. Sci. Eng. 2019, 6, 1112–1123. [Google Scholar] [CrossRef]
- García-Lizarribar, A.; Fernández-Garibay, X.; Velasco-Mallorquí, F.; Castaño, A.G.; Samitier, J.; Ramon-Azcon, J. Composite Biomaterials as Long-Lasting Scaffolds for 3D Bioprinting of Highly Aligned Muscle Tissue. Macromol. Biosci. 2018, 18, 1800167. [Google Scholar] [CrossRef] [PubMed]
- Noh, S.; Gong, H.Y.; Lee, H.J.; Koh, W.-G. Electrically Conductive Micropatterned Polyaniline-Poly (ethylene glycol) Composite Hydrogel. Materials 2021, 14, 308. [Google Scholar] [CrossRef] [PubMed]
- Ku, S.H.; Park, C.B. Myoblast differentiation on graphene oxide. Biomaterials 2013, 34, 2017–2023. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, B.; Mondal, B.; Kumar, S.; Sarkar, S. Myoblast differentiation and protein expression in electrospun graphene oxide (GO)-poly (ε-caprolactone, PCL) composite meshes. Mater. Lett. 2016, 182, 194–197. [Google Scholar] [CrossRef]
- Uehara, T.M.; Paino, I.M.; Santos, F.A.; Scagion, V.P.; Correa, D.S.; Zucolotto, V. Fabrication of random and aligned electrospun nanofibers containing graphene oxide for skeletal muscle cells scaffold. Polym. Adv. Technol. 2020, 31, 1437–1443. [Google Scholar] [CrossRef]
- Velasco-Mallorquí, F.; Fernández-Costa, J.M.; Neves, L.; Ramón-Azcón, J. New volumetric CNT-doped gelatin–cellulose scaffolds for skeletal muscle tissue engineering. Nanoscale Adv. 2020, 2, 2885–2896. [Google Scholar] [CrossRef]
- Patel, A.; Mukundan, S.; Wang, W.; Karumuri, A.; Sant, V.; Mukhopadhyay, S.M.; Sant, S. Carbon-based hierarchical scaffolds for myoblast differentiation: Synergy between nano-functionalization and alignment. Acta Biomater. 2016, 32, 77–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCorry, M.C.; Ohlson, C.; Gunnel, S.; Higginbottom, S.; Billiar, K.; Page, R. Mechanical stimulation device for skeletal muscle tissue engineering. In Proceedings of the 2012 38th Annual Northeast Bioengineering Conference (NEBEC), Philadelphia, PA, USA, 16–18 March 2012; pp. 249–250. [Google Scholar]
- Ten Broek, R.W.; Grefte, S.; Von den Hoff, J.W. Regulatory factors and cell populations involved in skeletal muscle regeneration. J. Cell Physiol. 2010, 224, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Powell, C.A.; Smiley, B.L.; Mills, J.; Vandenburgh, H.H. Mechanical stimulation improves tissue-engineered human skeletal muscle. Am. J. Physiol. Cell Physiol. 2002, 283, C1557–C1565. [Google Scholar] [CrossRef] [PubMed]
- Zanchi, N.E.; Lancha, A.H., Jr. Mechanical stimuli of skeletal muscle: Implications on mTOR/p70s6k and protein synthesis. Eur. J. Appl. Physiol. 2008, 102, 253–263. [Google Scholar] [CrossRef]
- Okano, T.; Matsuda, T. Tissue engineered skeletal muscle: Preparation of highly dense, highly oriented hybrid muscular tissues. Cell Transplant. 1998, 7, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Auluck, A.; Mudera, V.; Hunt, N.P.; Lewis, M.P. A three-dimensional in vitro model system to study the adaptation of craniofacial skeletal muscle following mechanostimulation. Eur. J. Oral. Sci. 2005, 113, 218–224. [Google Scholar] [CrossRef]
- Sakiyama, K.; Abe, S.; Tamatsu, Y.; Ide, Y. Effects of stretching stress on the muscle contraction proteins of skeletal muscle myoblasts. Biomed. Res. 2005, 26, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.J.; Truskey, G.A.; Kraus, W.E. Effect of cyclic stretch on β1D-integrin expression and activation of FAK and RhoA. Am. J. Physiol. Cell Physiol. 2007, 292, C2057–C2069. [Google Scholar] [CrossRef] [Green Version]
- Moon, D.G.; Christ, G.; Stitzel, J.D.; Atala, A.; Yoo, J.J. Cyclic mechanical preconditioning improves engineered muscle contraction. Tissue Eng. Part A 2008, 14, 473–482. [Google Scholar] [CrossRef]
- Egusa, H.; Kobayashi, M.; Matsumoto, T.; Sasaki, J.-I.; Uraguchi, S.; Yatani, H. Application of Cyclic Strain for Accelerated Skeletal Myogenic Differentiation of Mouse Bone Marrow-Derived Mesenchymal Stromal Cells with Cell Alignment. Tissue Eng. Part A 2013, 19, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.G. On the stereotropism of embryonic cells. Science 1911, 34, 279–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, P. Experiments on cell and axon orientation in vitro: The role of colloidal exudates in tissue organization. J. Exp. Zool. 1945, 100, 353–386. [Google Scholar] [CrossRef]
- Li, Y.; Huang, G.; Zhang, X.; Wang, L.; Du, Y.; Lu, T.J.; Xu, F. Engineering cell alignment in vitro. Biotechnol. Adv. 2014, 32, 347–365. [Google Scholar] [CrossRef]
- Metavarayuth, K.; Sitasuwan, P.; Zhao, X.; Lin, Y.; Wang, Q. Engineering. Influence of surface topographical cues on the differentiation of mesenchymal stem cells in vitro. ACS Biomater. Sci. Eng. 2016, 2, 142–151. [Google Scholar] [CrossRef]
- Avci, H.; Ghorbanpoor, H.; Nurbas, M. Preparation of origanum minutiflorum oil-loaded core–shell structured chitosan nanofibers with tunable properties. Polym. Bull. 2018, 75, 4129–4144. [Google Scholar] [CrossRef]
- Avci, H.; Akkulak, E.; Gergeroglu, H.; Ghorbanpoor, H.; Uysal, O.; Eker Sariboyaci, A.; Demir, B.; Soykan, M.N.; Pat, S.; Mohammadigharehbagh, R.; et al. Flexible poly (styrene-ethylene-butadiene-styrene) hybrid nanofibers for bioengineering and water filtration applications. J. Appl. Polym. Sci. 2020, 137, 49184. [Google Scholar] [CrossRef]
- Nemati, S.; Kim, S.-j.; Shin, Y.M.; Shin, H. Current progress in application of polymeric nanofibers to tissue engineering. Nano Converg. 2019, 6, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Hajiali, H.; Contestabile, A.; Mele, E.; Athanassiou, A. Influence of topography of nanofibrous scaffolds on functionality of engineered neural tissue. J. Mater. Chem. B 2018, 6, 930–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, N.F.; Patel, S.; Thakar, R.G.; Wu, J.; Hsiao, B.S.; Chu, B.; Lee, R.J.; Li, S. Myotube assembly on nanofibrous and micropatterned polymers. Nano Lett. 2006, 6, 537–542. [Google Scholar] [CrossRef]
- Woods, I.; Black, A.; Jockenhoevel, S.; Flanagan, T.C. Harnessing topographical & biochemical cues to enhance elastogenesis by paediatric cells for cardiovascular tissue engineering applications. Biochem. Biophys. Res. Commun. 2019, 512, 156–162. [Google Scholar]
- Tsai, S.-W.; Yu, Y.-L.; Hsu, F.-Y. Fabrication of polycaprolactone tubular scaffolds with an orthogonal-bilayer structure for smooth muscle cells. Mater. Sci. Eng. C 2019, 100, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, L.; Guo, B.; Ma, P.X. Interwoven aligned conductive nanofiber yarn/hydrogel composite scaffolds for engineered 3D cardiac anisotropy. ACS Nano 2017, 11, 5646–5659. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Su, C.; Zeng, Z.; Zhang, H.; Feng, R.; Feng, J.; Li, S. Fabrication of multilayer tubular scaffolds with aligned nanofibers to guide the growth of endothelial cells. J. Biomater. Appl. 2020, 35, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.; Park, S.M.; Lim, J.; Kim, D.S. Electrospun random/aligned hybrid nanofiber mat for development of multi-layered cardiac muscle patch. In Proceedings of the 2018 IEEE International Conference on Cyborg and Bionic Systems (CBS), Shenzhen, China, 25–27 October 2018; pp. 177–180. [Google Scholar]
- McKeon-Fischer, K.D.; Flagg, D.H.; Freeman, J.W. Coaxial electrospun poly (ε-caprolactone), multiwalled carbon nanotubes, and polyacrylic acid/polyvinyl alcohol scaffold for skeletal muscle tissue engineering. J. Biomed. Mater. Res. Part A 2011, 99, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, V.; Ahadian, S.; Ostrovidov, S.; Camci-Unal, G.; Chen, S.; Kaji, H.; Ramalingam, M.; Khademhosseini, A. Engineered contractile skeletal muscle tissue on a microgrooved methacrylated gelatin substrate. Tissue Eng. Part A 2012, 18, 2453–2465. [Google Scholar] [CrossRef] [Green Version]
- Jiwlawat, N.; Lynch, E.M.; Napiwocki, B.N.; Stempien, A.; Ashton, R.S.; Kamp, T.J.; Crone, W.C.; Suzuki, M. Micropatterned substrates with physiological stiffness promote cell maturation and Pompe disease phenotype in human induced pluripotent stem cell-derived skeletal myocytes. Biotechnol. Bioeng. 2019, 116, 2377–2392. [Google Scholar] [CrossRef]
- Chen, S.; Nakamoto, T.; Kawazoe, N.; Chen, G. Engineering multi-layered skeletal muscle tissue by using 3D microgrooved collagen scaffolds. Biomaterials 2015, 73, 23–31. [Google Scholar] [CrossRef]
- Capel, A.J.; Rimington, R.P.; Fleming, J.W.; Player, D.J.; Baker, L.A.; Turner, M.C.; Jones, J.M.; Martin, N.R.; Ferguson, R.A.; Mudera, V.C.; et al. Scalable 3D printed molds for human tissue engineered skeletal muscle. Front. Bioeng. Biotechnol. 2019, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Lee, H.; Lee, J.; Atala, A.; Yoo, J.J.; Lee, S.J.; Kim, G.H. Efficient myotube formation in 3D bioprinted tissue construct by biochemical and topographical cues. Biomaterials 2020, 230, 119632. [Google Scholar] [CrossRef]
- Choi, Y.-J.; Jun, Y.-J.; Kim, D.Y.; Yi, H.-G.; Chae, S.-H.; Kang, J.; Lee, J.; Gao, G.; Kong, J.-S.; Jang, J.; et al. A 3D cell printed muscle construct with tissue-derived bioink for the treatment of volumetric muscle loss. Biomaterials 2019, 206, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Distler, T.; Sulistio, A.; Schneidereit, D.; Friedrich, O.; Detsch, R.; Boccaccini, A.R. 3D printed oxidized alginate-gelatin bioink provides guidance for C2C12 muscle precursor cell orientation and differentiation via shear stress during bioprinting. Biofabrication 2020, 12, 045005. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Kim, M.; Kim, G.H. 3D-printed biomimetic scaffold simulating microfibril muscle structure. Adv. Funct. Mater. 2018, 28, 1800405. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alarcin, E.; Bal-Öztürk, A.; Avci, H.; Ghorbanpoor, H.; Dogan Guzel, F.; Akpek, A.; Yesiltas, G.; Canak-Ipek, T.; Avci-Adali, M. Current Strategies for the Regeneration of Skeletal Muscle Tissue. Int. J. Mol. Sci. 2021, 22, 5929. https://doi.org/10.3390/ijms22115929
Alarcin E, Bal-Öztürk A, Avci H, Ghorbanpoor H, Dogan Guzel F, Akpek A, Yesiltas G, Canak-Ipek T, Avci-Adali M. Current Strategies for the Regeneration of Skeletal Muscle Tissue. International Journal of Molecular Sciences. 2021; 22(11):5929. https://doi.org/10.3390/ijms22115929
Chicago/Turabian StyleAlarcin, Emine, Ayca Bal-Öztürk, Hüseyin Avci, Hamed Ghorbanpoor, Fatma Dogan Guzel, Ali Akpek, Gözde Yesiltas, Tuba Canak-Ipek, and Meltem Avci-Adali. 2021. "Current Strategies for the Regeneration of Skeletal Muscle Tissue" International Journal of Molecular Sciences 22, no. 11: 5929. https://doi.org/10.3390/ijms22115929