
Proteome Characterization of BALF Extracellular Vesicles in Idiopathic Pulmonary Fibrosis: Unveiling Undercover Molecular Pathways

 , , , , , , , , ,
, , , , , , , , ,
Abstract
:1. Introduction
2. Results
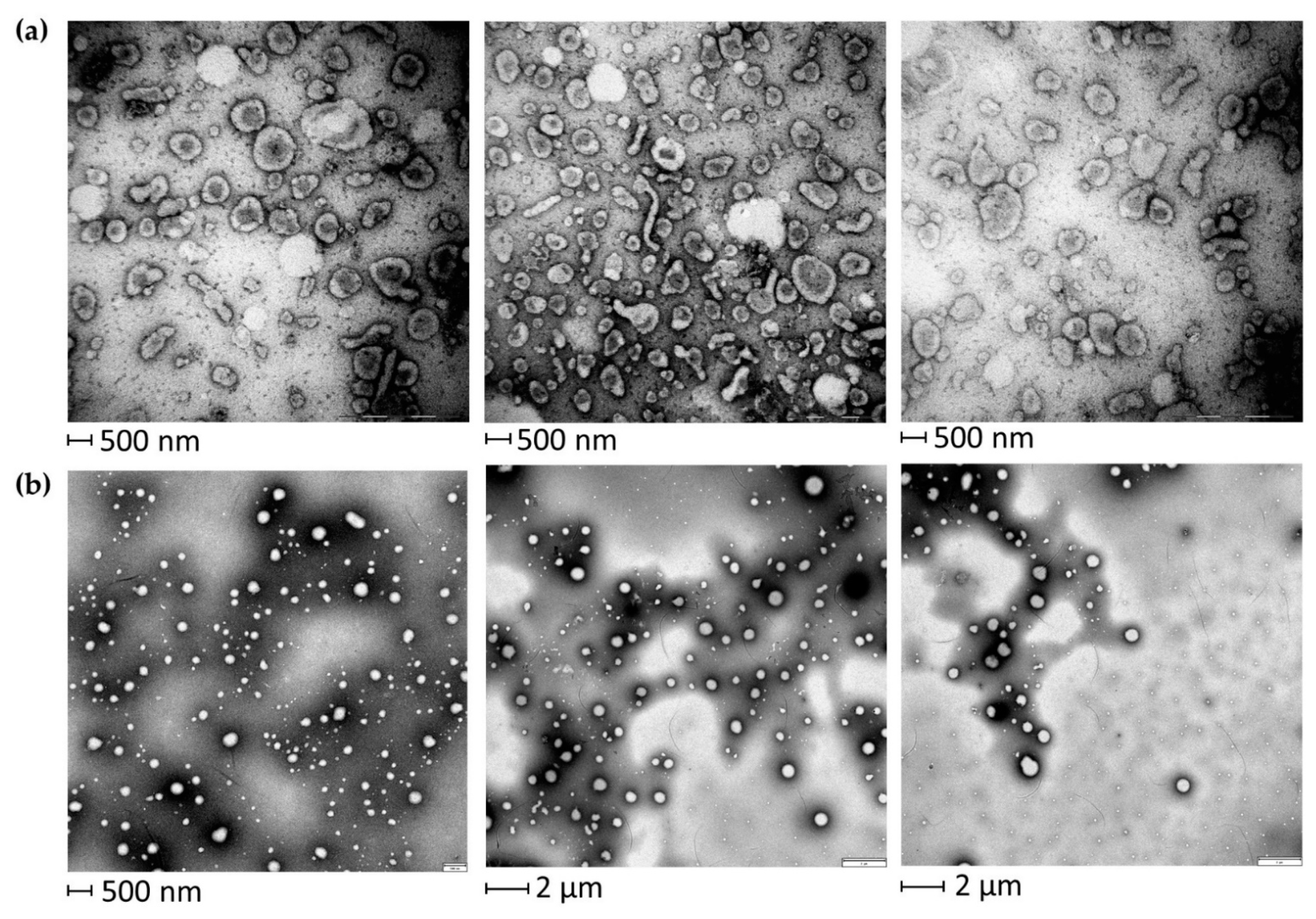
2.1. Quality Control Assessment of EVs from BALF of IPF Patients
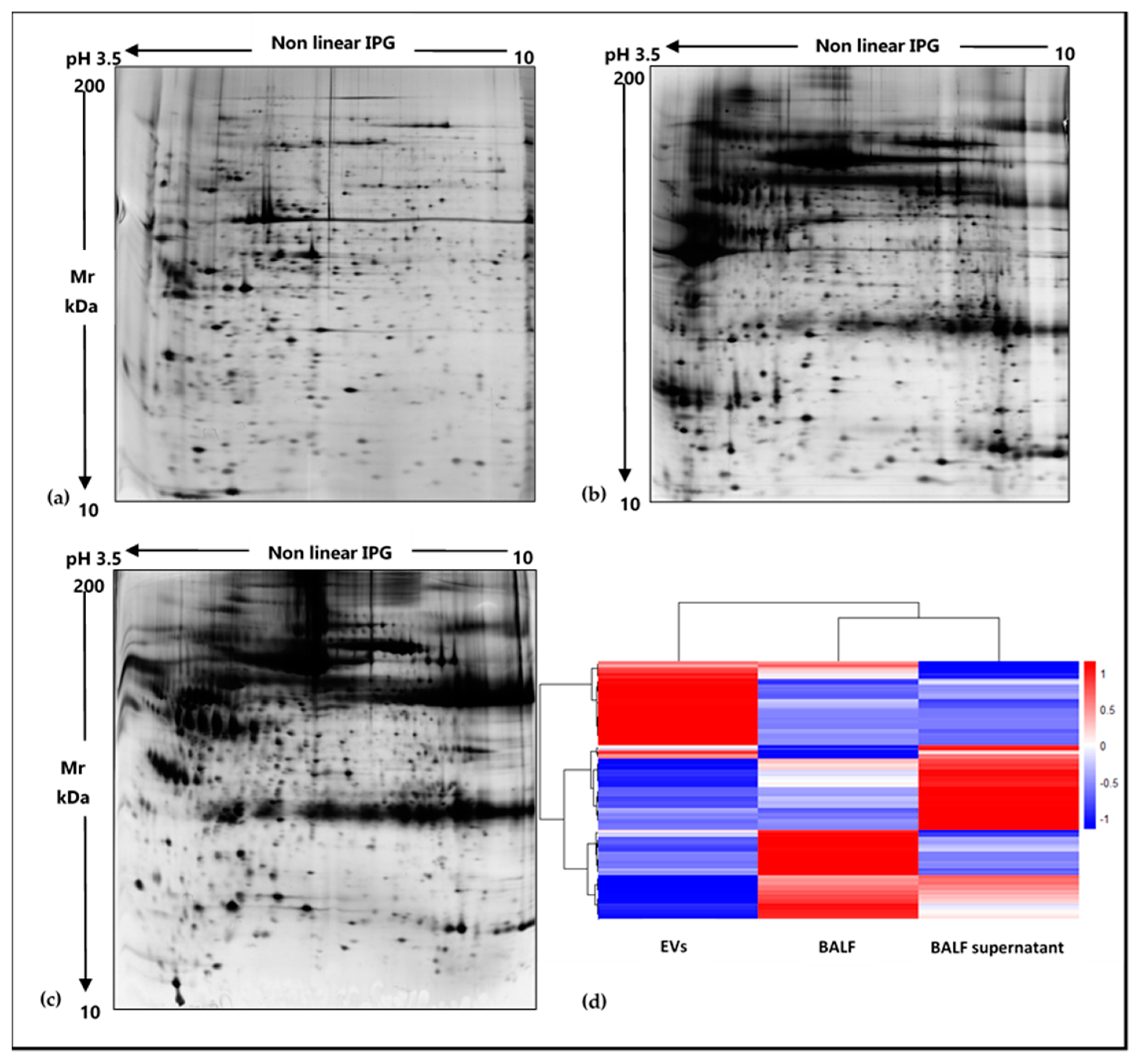
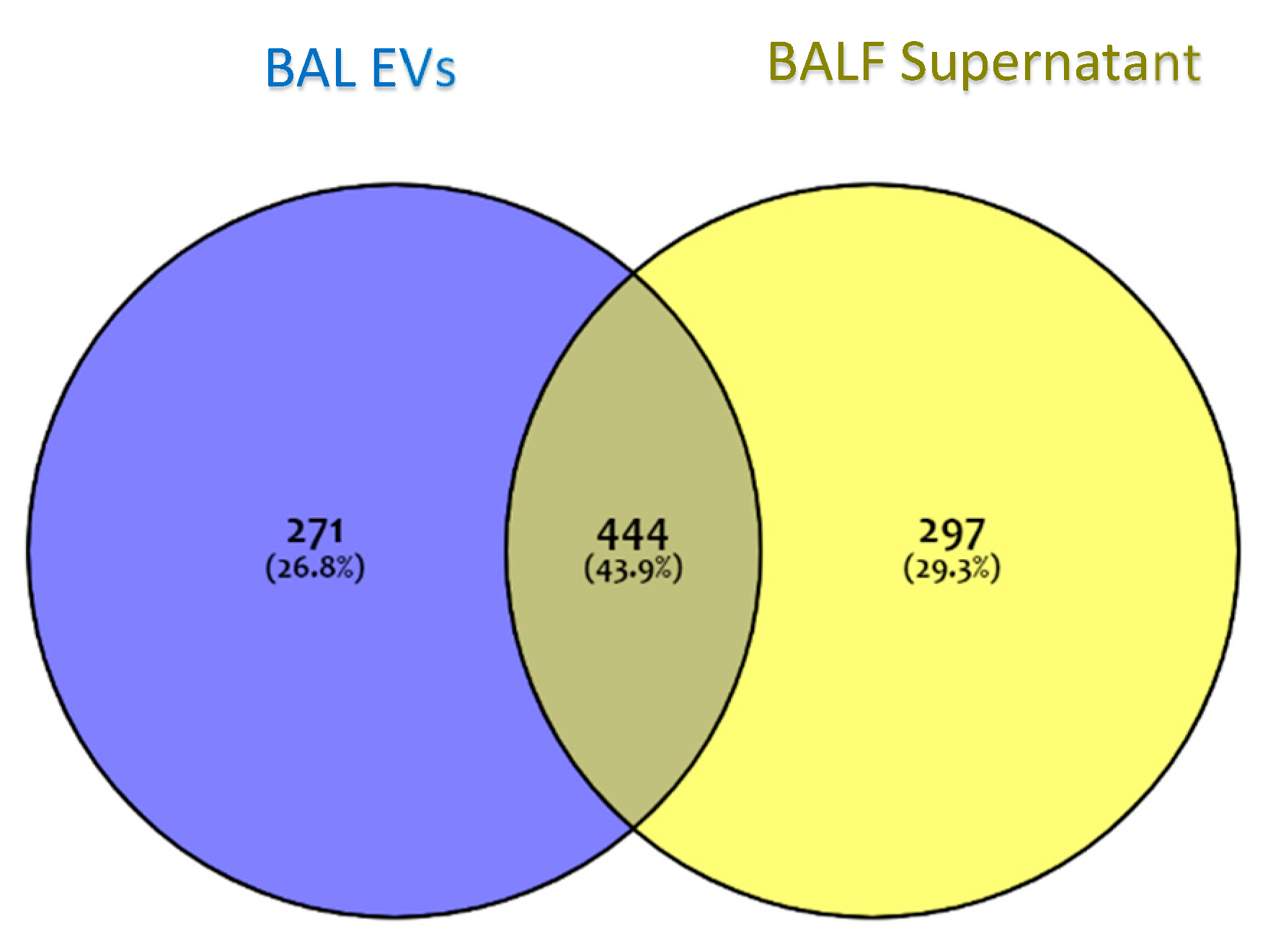
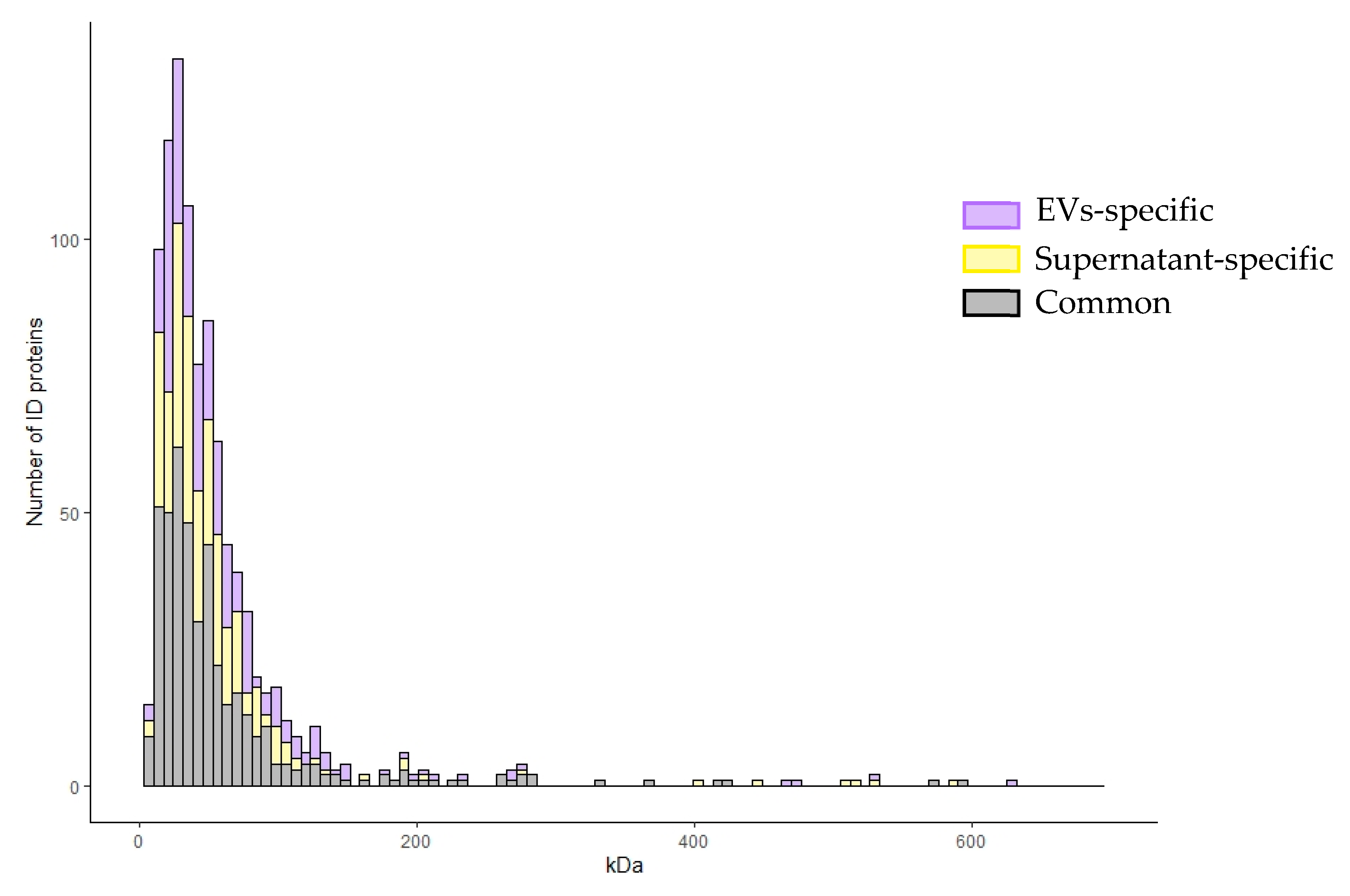
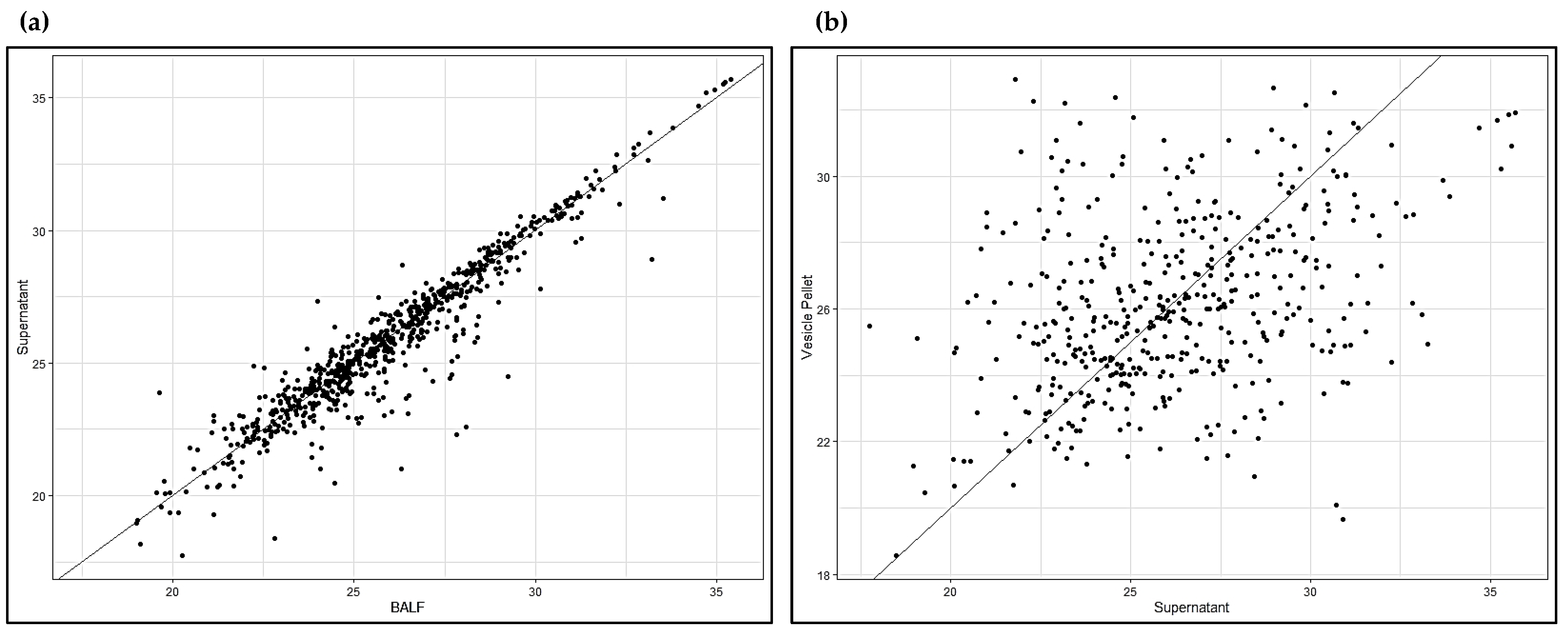
2.2. Shotgun Proteomics of BALF EVs and BALF Fluidic Portion
2.3. MetaCore Enrichment Analysis
3. Discussion
4. Materials and Methods
4.1. Population
4.2. EVs Isolation from BALF
4.3. TEM
4.4. Samples Preparation for 2DE Analysis
4.5. 2D-Electrophoresis
4.6. MS-Preparative SDS-PAGE
4.7. Preparation of Samples for LC-MS/MS
4.8. LC-MS/MS Analysis and Protein Identification
4.9. Enrichment Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reynolds, H.Y. Use of Bronchoalveolar Lavage in Humans—Past Necessity and Future Imperative. Lung 2000, 178, 271–293. [Google Scholar] [CrossRef]
- Govender, P.; Dunn, M.J.; Donnelly, S.C. Proteomics and the lung: Analysis of bronchoalveolar lavage fluid. Proteom. Clin. Appl. 2009, 3, 1044–1051. [Google Scholar] [CrossRef]
- Nguyen, E.V.; Gharib, S.A.; Schnapp, L.M.; Goodlett, D.R. Shotgun MS proteomic analysis of bronchoalveolar lavage fluid in normal subjects. Proteom. Clin. Appl. 2014, 8, 737–747. [Google Scholar] [CrossRef] [Green Version]
- Travis, W.D.; Costabel, U.; Hansell, D.M.; King, T.E.; Lynch, D.A.; Nicholson, A.G.; Ryerson, C.J.; Ryu, J.H.; Selman, M.; Wells, A.U.; et al. An Official American Thoracic Society/European Respiratory Society Statement: Update of the International Multidisciplinary Classification of the Idiopathic Interstitial Pneumonias. Am. J. Respir. Crit. Care Med. 2013, 188, 733–748. [Google Scholar] [CrossRef]
- Landi, C.; Bargagli, E.; Carleo, A.; Bianchi, L.; Gagliardi, A.; Prasse, A.; Perari, M.G.; Refini, R.M.; Bini, L.; Rottoli, P. A system biology study of BALF from patients affected by idiopathic pulmonary fibrosis (IPF) and healthy controls. Proteom. Clin. Appl. 2014, 8, 932–950. [Google Scholar] [CrossRef] [PubMed]
- Carleo, A.; Landi, C.; Prasse, A.; Bergantini, L.; D’Alessandro, M.; Cameli, P.; Janciauskiene, S.; Rottoli, P.; Bini, L.; Bargagli, E. Proteomic characterization of idiopathic pulmonary fibrosis patients: Stable versus acute exacerbation. Monaldi Arch. Chest Dis. 2020, 90. [Google Scholar] [CrossRef] [PubMed]
- Efared, B.; Ebang-Atsame, G.; Rabiou, S.; Diarra, A.S.; Tahiri, L.; Hammas, N.; Smahi, M.; Amara, B.; Benjelloun, M.C.; Serraj, M.; et al. The diagnostic value of the bronchoalveolar lavage in interstitial lung diseases. J. Negat. Results Biomed. 2017, 16, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rottoli, P.; Bargagli, E.; Landi, C.; Magi, B. Proteomic analysis in interstitial lung diseases: A review. Curr. Opin. Pulm. Med. 2009, 15, 470–478. [Google Scholar] [CrossRef]
- Landi, C.; Bargagli, E.; Bianchi, L.; Gagliardi, A.; Carleo, A.; Bennett, D.; Perari, M.G.; Armini, A.; Prasse, A.; Rottoli, P.; et al. Towards a functional proteomics approach to the comprehension of idiopathic pulmonary fibrosis, sarcoidosis, systemic sclerosis and pulmonary Langerhans cell histiocytosis. J. Proteom. 2013, 83, 60–75. [Google Scholar] [CrossRef]
- D’Alessandro, M.; Bergantini, L.; Refini, R.M.; Cameli, P.; Perillo, F.; Landi, C.; Icorne, F.; Perrone, A.; Sestini, P.; Bonella, F.; et al. Adiponectin and leptin levels in idiopathic pulmonary fibrosis: A new method for BAL and serum assessment. Immunobiology 2020, 225, 151997. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Rollet-Cohen, V.; Bourderioux, M.; Lipecka, J.; Chhuon, C.; Jung, V.A.; Mesbahi, M.; Nguyen-Khoa, T.; Guérin-Pfyffer, S.; Schmitt, A.; Edelman, A.; et al. Comparative proteomics of respiratory exosomes in cystic fibrosis, primary ciliary dyskinesia and asthma. J. Proteom. 2018, 185, 1–7. [Google Scholar] [CrossRef]
- Njock, M.-S.; Guiot, J.; Henket, M.A.; Nivelles, O.; Thiry, M.; Dequiedt, F.; Corhay, J.-L.; Louis, R.E.; Struman, I. Sputum exosomes: Promising biomarkers for idiopathic pulmonary fibrosis. Thorax 2018, 74, 309–312. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M. The Roles of MicroRNAs and Extracellular Vesicles in the Pathogeneses of Idiopathic Pulmonary Fibrosis and Acute Respiratory Distress Syndrome. Tohoku J. Exp. Med. 2020, 251, 313–326. [Google Scholar] [CrossRef]
- Lee, H.; Groot, M.; Pinilla-Vera, M.; Fredenburgh, L.E.; Jin, Y. Identification of miRNA-rich vesicles in bronchoalveolar lavage fluid: Insights into the function and heterogeneity of extracellular vesicles. J. Control. Release 2019, 294, 43–52. [Google Scholar] [CrossRef]
- Liu, B.; Jiang, T.; Hu, X.; Liu, Z.; Zhao, L.; Liu, H.; Liu, Z.; Ma, L. Downregulation of microRNA-30a in bronchoalveolar lavage fluid from idiopathic pulmonary fibrosis patients. Mol. Med. Rep. 2018, 18, 5799–5806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Medina, A.; Lehmann, M.; Burgy, O.; Hermann, S.; Baarsma, H.A.; Wagner, D.E.; De Santis, M.M.; Ciolek, F.; Hofer, T.P.; Frankenberger, M.; et al. Increased Extracellular Vesicles Mediate WNT5A Signaling in Idiopathic Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 2018, 198, 1527–1538. [Google Scholar] [CrossRef] [PubMed]
- Wheelock, C.E.; Goss, V.M.; Balgoma, D.; Nicholas, B.; Brandsma, J.; Skipp, P.J.; Snowden, S.; Burg, D.; D’Amico, A.; Horvath, I.; et al. Application of ’omics technologies to biomarker discovery in inflammatory lung diseases. Eur. Respir. J. 2013, 42, 802–825. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.Y.; Choi, Y.R.; Lee, J.H.; Lim, J.M.; Lee, S.H.; Kim, K.P.; Kim, J.Y.; Kim, M. In-Depth Proteomic Analysis of Human Bronchoalveolar Lavage Fluid toward the Biomarker Discovery for Lung Cancers. Proteom. Clin. Appl. 2019, 13, e1900028. [Google Scholar] [CrossRef]
- Matthiesen, R. MS-Based Biomarker Discovery in Bronchoalveolar Lavage Fluid for Lung Cancer. Proteom. Clin. Appl. 2020, 14, e1900077. [Google Scholar] [CrossRef]
- Liessi, N.; Pedemonte, N.; Armirotti, A.; Braccia, C. Proteomics and Metabolomics for Cystic Fibrosis Research. Int. J. Mol. Sci. 2020, 21, 5439. [Google Scholar] [CrossRef]
- Braccia, C.; Tomati, V.; Caci, E.; Pedemonte, N.; Armirotti, A. SWATH label-free proteomics for cystic fibrosis research. J. Cyst. Fibros. 2019, 18, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Landi, C.; Cameli, P.; Vantaggiato, L.; Bergantini, L.; D’Alessandro, M.; Perruzza, M.; Carleo, A.; Shaba, E.; Di Giuseppe, F.; Angelucci, S.; et al. Ceruloplasmin and oxidative stress in severe eosinophilic asthma patients treated with Mepolizumab and Benralizumab. Biochim. et Biophys. Acta (BBA) Proteins Proteom. 2021, 1869, 140563. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, L.; Chen, D.; Feng, M.; Lu, Y.; Chen, R.; Qiu, C.; Li, J. The application of proteomics in the diagnosis and treatment of bronchial asthma. Ann. Transl. Med. 2020, 8, 132. [Google Scholar] [CrossRef]
- Moon, J.-Y.; Filho, F.S.L.; Shahangian, K.; Takiguchi, H.; Sin, D.D. Blood and sputum protein biomarkers for chronic obstructive pulmonary disease (COPD). Expert Rev. Proteom. 2018, 15, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, H.; Li, C.; Ma, C.; Ge, W. Proteome Profiling of Lung Tissues in Chronic Obstructive Pulmonary Disease (COPD): Platelet and Macrophage Dysfunction Contribute to the Pathogenesis of COPD. Int. J. Chronic Obstr. Pulm. Dis. 2020, 15, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Neri, T.; Tavanti, L.; De Magistris, S.; Lombardi, S.; Romei, C.; Falaschi, F.; Paggiaro, P.; Celi, A. Endothelial Cell-Derived Extracellular Vesicles as Potential Biomarkers in Chronic Interstitial Lung Diseases. Ann. Clin. Lab. Sci. 2019, 49, 608–610. [Google Scholar] [PubMed]
- McVey, M.J.; Maishan, M.; Blokland, K.E.C.; Bartlett, N.; Kuebler, W.M. Extracellular vesicles in lung health, disease, and therapy. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2019, 316, L977–L989. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Cizmar, P.; Yuana, Y. Detection and Characterization of Extracellular Vesicles by Transmission and Cryo-Transmission Elec-tron Microscopy. In Extracellular Vesicles; Humana Press: New York, NY, USA, 2017; Volume 1660, pp. 221–232. [Google Scholar] [CrossRef]
- Kugeratski, F.G.; Kalluri, R. Exosomes as mediators of immune regulation and immunotherapy in cancer. FEBS J. 2021, 288, 10–35. [Google Scholar] [CrossRef]
- Khan, T.; Dasgupta, S.; Ghosh, N.; Chaudhury, K. Proteomics in idiopathic pulmonary fibrosis: The quest for biomarkers. Mol. Omics 2021, 17, 43–58. [Google Scholar] [CrossRef]
- Magi, B.; Bini, L.; Perari, M.G.; Fossi, A.; Sanchez, J.-C.; Hochstrasser, D.; Paesano, S.; Raggiaschi, R.; Santucci, A.; Pallini, V.; et al. Bronchoalveolar lavage fluid protein composition in patients with sarcoidosis and idiopathic pulmonary fibrosis: A two-dimensional electrophoretic study. Electrophoresis 2002, 23, 3434–3444. [Google Scholar] [CrossRef]
- Rottoli, P.; Magi, B.; Cianti, R.; Bargagli, E.; Vagaggini, C.; Nikiforakis, N.; Pallini, V.; Bini, L. Carbonylated proteins in bronchoalveolar lavage of patients with sarcoidosis, pulmonary fibrosis associated with systemic sclerosis and idiopathic pulmonary fibrosis. Proteomics 2005, 5, 2612–2618. [Google Scholar] [CrossRef] [PubMed]
- Rottoli, P.; Magi, B.; Perari, M.G.; Liberatori, S.; Nikiforakis, N.; Bargagli, E.; Cianti, R.; Bini, L.; Pallini, V. Cytokine profile and proteome analysis in bronchoalveolar lavage of patients with sarcoidosis, pulmonary fibrosis associated with systemic sclerosis and idiopathic pulmonary fibrosis. Proteomics 2005, 5, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Cameli, P.; Carleo, A.; Bergantini, L.; Landi, C.; Prasse, A.; Bargagli, E. Oxidant/Antioxidant Disequilibrium in Idiopathic Pulmonary Fibrosis Pathogenesis. Inflammation 2020, 43, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bargagli, E.; Olivieri, C.; Nikiforakis, N.; Cintorino, M.; Magi, B.; Perari, M.G.; Vagaggini, C.; Spina, D.; Prasse, A.; Rottoli, P. Analysis of macrophage migration inhibitory factor (MIF) in patients with idiopathic pulmonary fibrosis. Respir. Physiol. Neurobiol. 2009, 167, 261–267. [Google Scholar] [CrossRef]
- Bargagli, E.; Olivieri, C.; Prasse, A.; Bianchi, N.; Magi, B.; Cianti, R.; Bini, L.; Rottoli, P. Calgranulin B (S100A9) Levels in Bronchoalveolar Lavage Fluid of Patients with Interstitial Lung Diseases. Inflammation 2008, 31, 351–354. [Google Scholar] [CrossRef]
- Hara, A.; Sakamoto, N.; Ishimatsu, Y.; Kakugawa, T.; Nakashima, S.; Hara, S.; Adachi, M.; Fujita, H.; Mukae, H.; Kohno, S. S100A9 in BALF is a candidate biomarker of idiopathic pulmonary fibrosis. Respir. Med. 2012, 106, 571–580. [Google Scholar] [CrossRef] [Green Version]
- Carleo, A.; Bargagli, E.; Landi, C.; Bennett, D.; Bianchi, L.; Gagliardi, A.; Carnemolla, C.; Perari, M.G.; Cillis, G.; Armini, A.; et al. Comparative proteomic analysis of bronchoalveolar lavage of familial and sporadic cases of idiopathic pulmonary fibrosis. J. Breath Res. 2016, 10, 026007. [Google Scholar] [CrossRef]
- Foster, M.W.; Morrison, L.D.; Todd, J.L.; Snyder, L.; Thompson, J.W.; Soderblom, E.J.; Plonk, K.; Weinhold, K.J.; Townsend, R.; Minnich, A.; et al. Quantitative Proteomics of Bronchoalveolar Lavage Fluid in Idiopathic Pulmonary Fibrosis. J. Proteome Res. 2015, 14, 1238–1249. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Maleki, M.; Sathyapalan, T.; Sahebkar, A. The effect of C-peptide on diabetic nephropathy: A review of molecular mechanisms. Life Sci. 2019, 237, 116950. [Google Scholar] [CrossRef]
- Venugopal, S.K.; Mowery, M.L.; Jialal, I. C Peptide; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Kitamura, T.; Kimura, K.; Jung, B.D.; Makondo, K.; Okamoto, S.; Cañas, X.; Sakane, N.; Yoshida, T.; Saito, M. Proinsulin C-peptide rapidly stimulates mitogen-activated protein kinases in Swiss 3T3 fibroblasts: Requirement of protein kinase C, phosphoinositide 3-kinase and pertussis toxin-sensitive G-protein. Biochem. J. 2001, 355, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Kwan, J.Y.Y.; Yip, K.; Liu, P.P.; Liu, F.-F. Targeting metabolic dysregulation for fibrosis therapy. Nat. Rev. Drug Discov. 2020, 19, 57–75. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, Y.; Zhang, W.; Chen, Y.; Chen, X.; Wang, C.; Li, Q.; Chen, C.; Jiang, B.; Lu, Y. C-peptide is associated with NAFLD inflammatory and fibrotic progression in type 2 diabetes. Diabetes/Metab. Res. Rev. 2020, 36, e3210. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhong, Y.; Gong, W.; Gao, X.; Qi, H.; Liu, K.; Qi, J. C-peptide prevents SMAD3 binding to alpha promoters to inhibit collagen type IV synthesis. J. Mol. Endocrinol. 2018, 61, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Chima, R.S.; Lamontagne, T.; Piraino, G.; Hake, P.W.; Denenberg, A.; Zingarelli, B. C-peptide, a novel inhibitor of lung inflammation following hemorrhagic shock. Am. J. Physiol. Cell. Mol. Physiol. 2011, 300, L730–L739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargagli, E.; Refini, R.M.; D’Alessandro, M.; Bergantini, L.; Cameli, P.; Vantaggiato, L.; Bini, L.; Landi, C. Metabolic Dysregulation in Idiopathic Pulmonary Fibrosis. Int. J. Mol. Sci. 2020, 21, 5663. [Google Scholar] [CrossRef]
- Costola-De-Souza, C.; Ribeiro, A.; Ferraz-De-Paula, V.; Calefi, A.S.; Aloia, T.P.A.; Junior, J.A.G.; De Almeida, V.I.; Pinheiro, M.L.; Palermo-Neto, J. Monoacylglycerol Lipase (MAGL) Inhibition Attenuates Acute Lung Injury in Mice. PLoS ONE 2013, 8, e77706. [Google Scholar] [CrossRef]
- Habib, A.; Chokr, D.; Wan, J.; Hegde, P.; Mabire, M.; Siebert, M.; Ribeiro-Parenti, L.; Le Gall, M.; Lettéron, P.; Pilard, N.; et al. Inhibition of monoacylglycerol lipase, an anti-inflammatory and antifibrogenic strategy in the liver. Gut 2019, 68, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Stempien-Otero, A.; Kim, D.-H.; Davis, J. Molecular networks underlying myofibroblast fate and fibrosis. J. Mol. Cell. Cardiol. 2016, 97, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Guillotin, D.; Taylor, A.R.; Platé, M.; Mercer, P.F.; Edwards, L.M.; Haggart, R.; Miele, G.; McAnulty, R.J.; Maher, T.M.; E Hynds, R.; et al. Transcriptome analysis of IPF fibroblastic foci identifies key pathways involved in fibrogenesis. Thorax 2021, 76, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Knipe, R.S.; Tager, A.M.; Liao, J.K. The Rho Kinases: Critical Mediators of Multiple Profibrotic Processes and Rational Targets for New Therapies for Pulmonary Fibrosis. Pharmacol. Rev. 2015, 67, 103–117. [Google Scholar] [CrossRef]
- Franco, C.A.; Liebner, S.; Gerhardt, H. Vascular morphogenesis: A Wnt for every vessel? Curr. Opin. Genet. Dev. 2009, 19, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Liu, W.; Ma, L.; Wang, Y.; Ma, J.; Jiang, M.; Deng, X.; Huang, F.; Yang, T.; Chen, M. Profilin-1 contributes to cardiac injury induced by advanced glycation end-products in rats. Mol. Med. Rep. 2017, 16, 6634–6641. [Google Scholar] [CrossRef] [Green Version]
- Shaikh, G.; Cronstein, B. Signaling pathways involving adenosine A2A and A2B receptors in wound healing and fibrosis. Purinergic Signal. 2016, 12, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Philip, K.; Mills, W.T.; Davies, J.; Chen, N.; Karmouty-Quintana, H.; Luo, F.; Molina, J.G.; Amione-Guerra, J.; Sinha, N.; Guha, A.; et al. HIF1A up-regulates the ADORA2B receptor on alternatively activated macrophages and contributes to pulmonary fibrosis. FASEB J. 2017, 31, 4745–4758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collum, S.D.; Molina, J.G.; Hanmandlu, A.; Bi, W.; Pedroza, M.; Mertens, T.C.J.; Wareing, N.; Wei, W.; Wilson, C.; Sun, W.; et al. Adenosine and hyaluronan modulate lung fibrosis and pulmonary hypertension in combined pulmonary fibrosis and emphysema (CPFE). Dis. Model. Mech. 2019, 12, dmm038711. [Google Scholar] [CrossRef] [Green Version]
- Giacomelli, C.; Daniele, S.; Romei, C.; Tavanti, L.; Neri, T.; Piano, I.; Celi, A.; Martini, C.; Trincavelli, M.L. The A2B Adenosine Receptor Modulates the Epithelial– Mesenchymal Transition through the Balance of cAMP/PKA and MAPK/ERK Pathway Activation in Human Epithelial Lung Cells. Front. Pharmacol. 2018, 9, 54. [Google Scholar] [CrossRef] [Green Version]
- Landi, C.; Carleo, A.; Vantaggiato, L.; Bergantini, L.; d’Alessandro, M.; Cameli, P.; Sebastiani, G.; Dotta, F.; Bargagli, E. Common Molecular Pathways Targeted by Nintedanib in Cancer and IPF: A Bioinformatic Study. Pulm. Pharmacol. Ther. 2020, 64, 101941. [Google Scholar] [CrossRef]
- Rassler, B. Role of α- and β-adrenergic Mechanisms in the Pathogenesis of Pulmonary Injuries Characterized by Edema, Inflammation and Fibrosis. Cardiovasc. Hematol. Disord. Targets 2014, 13, 197–207. [Google Scholar] [CrossRef]
- Prasse, A.; Binder, H.; Schupp, J.; Kayser, G.; Bargagli, E.; Jaeger, B.; Hess, M.; Rittinghausen, S.; Vuga, L.; Lynn, H.; et al. BAL Cell Gene Expression Is Indicative of Outcome and Airway Basal Cell Involvement in Idiopathic Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 2019, 199, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, C.; Jäger, B.; Wieczorek, G.; Engelhard, P.; Oliver, S.J.; Bauernfeind, F.-G.; Littlewood-Evans, A.; Welte, T.; Hornung, V.; Prasse, A. The NLRP3 inflammasome pathway is activated in sarcoidosis and involved in granuloma formation. Eur. Respir. J. 2020, 55, 1900119. [Google Scholar] [CrossRef] [PubMed]
- Harlow, E.; Lane, D. Bradford Assay. Cold Spring Harb. Protoc. 2006, 2006. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diagnosis | Gender | Age | Smoking | FVC | DLCO | GAP |
|---|---|---|---|---|---|---|
| IPF | Male | 74 | ex | 62 | 43 | 2 |
| IPF | Male | 63 | ex | 39 | 22 | 3 |
| IPF | Male | 69 | never | 50 | 32 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaba, E.; Landi, C.; Carleo, A.; Vantaggiato, L.; Paccagnini, E.; Gentile, M.; Bianchi, L.; Lupetti, P.; Bargagli, E.; Prasse, A.; et al. Proteome Characterization of BALF Extracellular Vesicles in Idiopathic Pulmonary Fibrosis: Unveiling Undercover Molecular Pathways. Int. J. Mol. Sci. 2021, 22, 5696. https://doi.org/10.3390/ijms22115696
Shaba E, Landi C, Carleo A, Vantaggiato L, Paccagnini E, Gentile M, Bianchi L, Lupetti P, Bargagli E, Prasse A, et al. Proteome Characterization of BALF Extracellular Vesicles in Idiopathic Pulmonary Fibrosis: Unveiling Undercover Molecular Pathways. International Journal of Molecular Sciences. 2021; 22(11):5696. https://doi.org/10.3390/ijms22115696
Chicago/Turabian StyleShaba, Enxhi, Claudia Landi, Alfonso Carleo, Lorenza Vantaggiato, Eugenio Paccagnini, Mariangela Gentile, Laura Bianchi, Pietro Lupetti, Elena Bargagli, Antje Prasse, and et al. 2021. "Proteome Characterization of BALF Extracellular Vesicles in Idiopathic Pulmonary Fibrosis: Unveiling Undercover Molecular Pathways" International Journal of Molecular Sciences 22, no. 11: 5696. https://doi.org/10.3390/ijms22115696