1. Introduction
Colorectal cancer (CRC) is the third most common cancer and the second most frequent cause of cancer-related mortality [
1]. With its high morbidity rate and incidence increasing year by year, CRC has become a global health problem. Chemotherapy is currently regarded as the standard treatment for patients with CRC, but it comes with many limitations, such as systemic toxicity, low selectivity and insufficient concentrations in tumour tissues [
2]. Combined with the ability of tumours to develop resistance to currently used therapy regimes [
3], there is a need not only for alternative therapies that will increase chemotherapy treatment efficacy and reduce side effects, but also for new therapies that are less aggressive and more effective than conventional ones [
2,
4].
The process of identifying new CRC therapies requires the use of systems that are cost-effective, reliable and fast—preferably automated. Traditional approaches have employed high-throughput screening for small molecules that is based primarily in 2D or 3D cell culture, enzymatic assays or receptor binding assays. Such studies in 2D have relied on the use of CRC cell lines such as Colo 205, SW480 or CR4, and assaying for effects on key markers and genes required for growth, migration and invasion using a combination of immunostaining, basic cell morphology, viability and proliferation assays [
5,
6,
7,
8]. 3D studies in spheroids or organoids have provided the opportunity for more in-depth study of the effects of treatments on physical tumour characteristics, with less reliance on biological markers such as E-cadherin or Vimentin [
9,
10]. Increasingly complex manipulations of 3D in vitro techniques have allowed the control of a variety of biochemical and biophysical factors, while providing the capacity for real-time imaging with standard microscopy. While 2D and 3D cell-culture systems have evolved to enable more sophisticated high-throughput therapeutic screenings, the ease of use and fast-turnaround times of such in vitro studies comes at the cost of removing the rich biological environment found in vivo, alongside a lack of targeted therapy delivery and limited physical outputs. Consequently, the majority of positive hits identified through these types of in vitro screens, are later found to be ineffective and/or toxic in subsequent validation experiments in whole-animal models [
11].
Drosophila melanogaster are emerging as a powerful alternative platform for high-throughput drug screens. With a rapid life cycle and low maintenance costs, such whole-organism-based drug screening permits the assessment of drug absorption, distribution, metabolic stability and toxicity, reducing the possibilities of false positives. With nearly 75% of human disease-causing genes believed to have a functional homologue in the fly [
12],
Drosophila are increasingly used as screening platforms for drug discovery and translational research [
13,
14]. There is a striking similarity between mammalian and
Drosophila intestinal tracts, which makes them particularly suited for modelling CRC [
15]. While on a gross anatomical level, the intestinal architecture differs between mammals and
Drosophila, they are composed of similar cell populations. The intestines in each are maintained by proliferative cells that give rise to two types of post-mitotic cell types; absorptive cells called enterocytes with a characteristic brush border and secretory cells. Therefore, the
Drosophila intestinal tract presents a ‘stripped down’ version of mammalian intestines that retains essential features of the stem cell biology and niche, which are among the essential features when modelling CRC.
A genetic powerhouse, the fly’s experimental tractability has identified a number of cancer-initiating mutations and has led to the generation of a large number of
Drosophila models for different cancers [
16,
17]. Combined mutations in the WNT and EGFR pathways are the most frequent initiating events of CRC [
18], and we and others have previously shown that it is possible to model CRC in
Drosophila by inducing clonal activation of the Wnt and Ras signalling pathways in the adult midgut [
19,
20]. Intestinal epithelial ApcRas clones mutant for the negative regulators of the Wnt pathway,
Apc and
Apc2, and overexpressing the oncogenic form of Ras, UAS-Ras
v12, form tumour-like overgrowths. We showed that these overgrowths reproduce many hallmarks of human CRC, including increased proliferation, a block in cell polarity and differentiation, as well as disrupted organ architecture. Furthermore, metabolic behavioural assays showed that these flies suffer a progressive deterioration in intestinal homeostasis [
19].
The behaviour of mutant clones generated in the adult
Drosophila midgut is commonly analysed by dissecting and imaging midguts, and quantifying parameters such as the number of clones, the number of cells in a clone and the clonal area manually using standard imaging analysis software [
19,
21,
22]. As these methods are laborious, they cannot be done on a rapid, large scale. Here, we have developed two approaches to make the analysis of tumour burden in the
Drosophila CRC model high-throughput, which could be easily adapted to other cancer models. First, we have developed custom built macros to be used in conjunction with Fiji, a widely available distribution of ImageJ for Life Science, which allow for the automated analysis of the total area of the midgut taken up with clones, as well as the average clone size. We demonstrate that clone size is the most robust determinant of whether a clone is behaving as a tumour or not. Secondly, having introduced a second reporter into the system, with clones now labelled with both GFP and luciferase, we show that the overall tumour burden can be rapidly and cheaply assessed by assaying whole fly lysates for luciferase. Finally, we show that after feeding drugs to the flies in their food, these assays can be used to assess tumour burden accurately and rapidly, and demonstrate a robust response of the
Drosophila CRC model to conventionally used CRC therapies. This study acts as a proof of principle to show that this model has the capability to be used in a high-throughput manner for future small compounds screens, and generates quantitative measurements of tumour characteristics that allow more in-depth characterisation of the efficacies of compounds.
2. Results
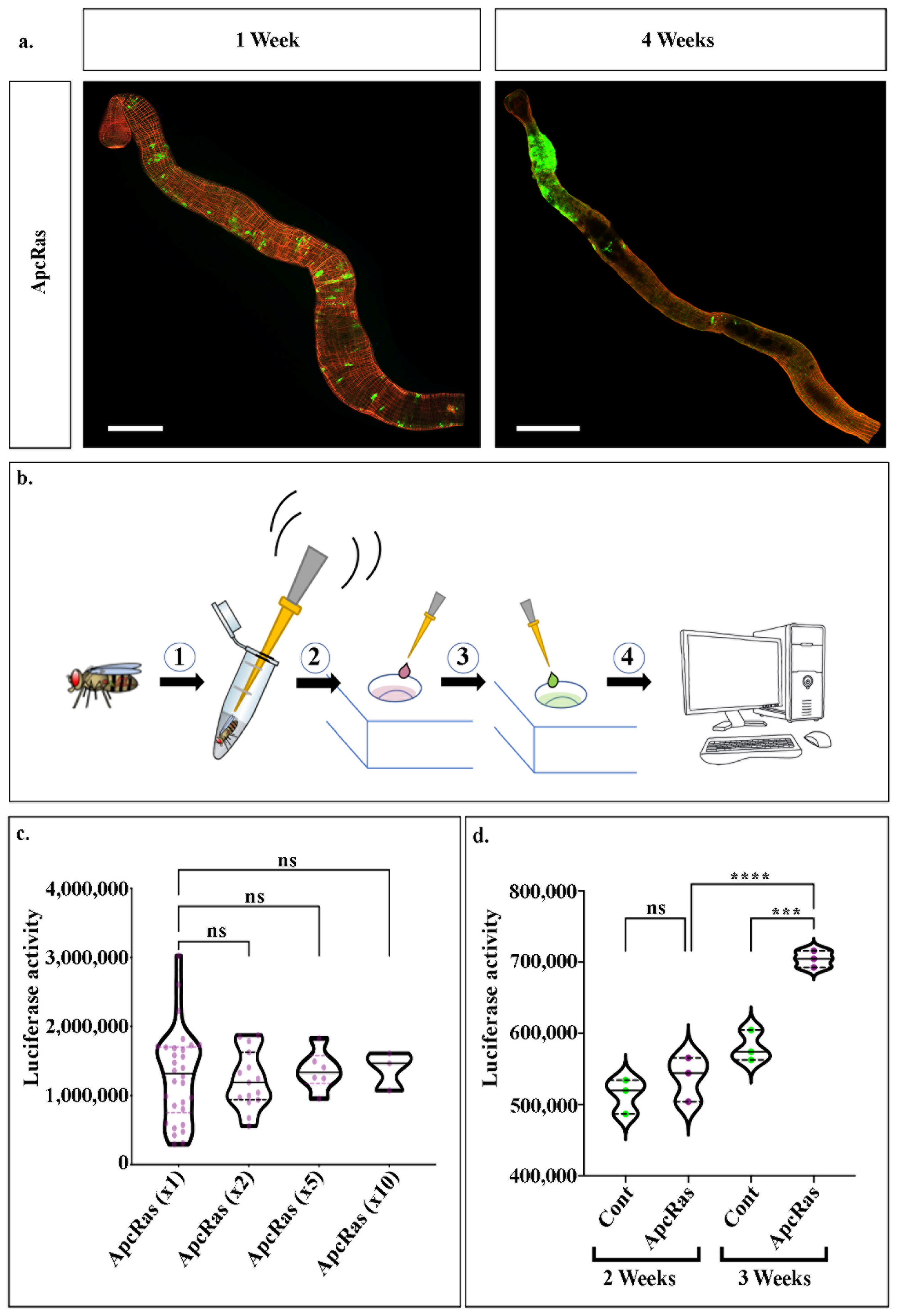
We have previously shown that, by means of the MARCM technique [
23], we can induce GFP-marked Apc-Ras clones in the intestinal stem cells of adult midguts. After 4 weeks, these clones form tumour-like overgrowths that share many similarities with human CRC (
Figure 1a and [
19]), generating an excellent model in adult
Drosophila for CRC [
19]. In order to be able to systematically quantify tumour growth, we recently generated a new CRC model that expresses a UAS-luciferase transgene [
24], in addition to UAS-GFP, in the ApcRas background [
25]. This model allows us to not only visualise cells using GFP, but also to exploit the highly sensitive luciferase-based assay to test for tumour burden, a process which is simple, quick to perform and provides rapid results. First, whole flies are homogenised directly into a lysis buffer, then the luciferin substrate is added, and finally the amount of luciferase activity is read using a luminometer (
Figure 1b) [
26]. As luciferase readings can be performed using a microplate reader, this process can be scaled up considerably, facilitating the analysis of many conditions in parallel.
As a first step towards setting up the conditions for screening whole flies for tumour burden using luciferase, we first separately assayed the luciferase activity in homogenates from 30 individual flies bearing ApcRas clones (
Figure 1b). In agreement with the high variability we described in this CRC model [
19], we observed a high variability in the luciferase readings between each fly, and thus used these measurements to determine the average luciferase activity in a fly with ApcRas clones (
Figure 1c). We next assayed the luciferase activity in homogenates taken from pairs of flies, groups of 5 flies, or groups of 10 (
Figure 1c). We found that increasing the number of flies to be processed together, not only reduces the variability in the luciferase readings, but leads to an almost identical average readout for luciferase activity. Taken together, this suggests that a rapid assay where 3 groups of 10 homogenised flies are put together, should give as reliable results as 30 individual assays. Using these conditions, we compared control flies bearing wildtype clones with flies bearing ApcRas clones, and found that while they gave comparable luciferase readings at 2 weeks, at 3 weeks their luciferase readings were significantly higher, correlating with our previous observations of the timing of tumour development in this model (
Figure 1d compare with [
19]).
We next sought to characterise in more detail how the whole fly luciferase readings relate to the behaviour of GFP positive clones of cells within the gut. As well as providing a more in-depth analysis of cell behaviour in different conditions, we reasoned that this could be used as a secondary screen to validate hits from a luciferase screen. This process, which we established when we developed and characterised the ApcRas model [
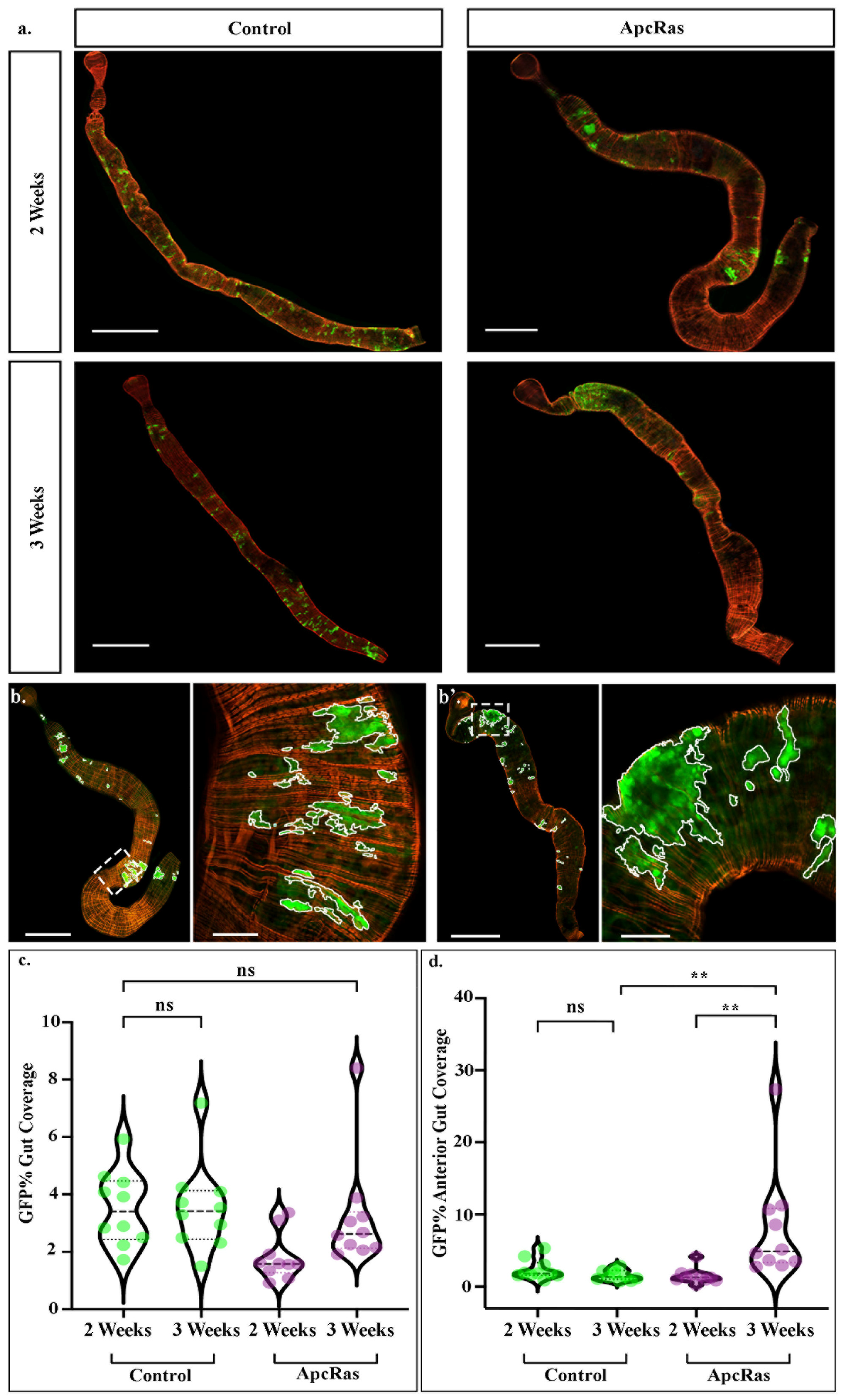
19], involves dissecting guts from mated females at 2 and 3 weeks after clones have been induced, fixing and staining for GFP to visualise the clones, and phalloidin, which labels F-Actin and shows the morphology of the midgut. As shown previously, control clones labelled just with GFP and luciferase, form small clusters dispersed throughout the gut, that do not appear to increase much in size (
Figure 2a). In contrast, ApcRas clones disappear over time, with the few that survive forming tumour-like overgrowths (
Figure 1a and
Figure 2a and [
19]). Until recently, we quantified tumour burden by imaging each midgut, stitching the images together manually (midguts can be up to 6mm long [
27,
28]), and then used standard functions on Fiji to outline and calculate the area of each clone. While this method provides highly accurate parameters to describe the behaviour of clones, it is extremely laborious and can also introduce user bias.
Moving towards a more rapid system for clonal analysis across entire midguts, we started to acquire tile-scans using a laser scanning microscope and image-stitching software, which enables the automated acquisition of a single continuous image of the entire organ. We have developed new tools for the analysis and extraction of reliable quantitative data from these tile scans. We have made these custom scripts publicly available (see
Section 4 for details on how to download and use them). They are for use with ImageJ, an open-source imaging processing and analysis platform, and allow the automated and accurate identification of GFP-positive areas along the entire midgut in seconds, as well as the quantification of various parameters such as area of each individual clone, or sum of the areas of all clones within a midgut (
Figure 2b–d). They also provide an option for a variable threshold which enables images of a range of qualities and microscopes to be analysed in a similar manner, allowing comparisons to be made across a variety of equipment.
As ApcRas clones are confined to the anterior portion of the midgut by 3 weeks after clone induction, we previously quantified tumour burden by calculating the % of the anterior portion of the midgut taken up by GFP positive clones [
19]. We therefore tested our custom macros by measuring the total clone area of wild type or ApcRas clones within the anterior portion of the midgut, along with the area of the anterior portion of the midgut itself, enabling the percentage area of the anterior midgut covered by clones to be calculated automatedly. While this requires manual selection of the anterior portion of the midgut, the use of thresholding and the automated quantification of GFP positive area removes potential user bias and, with proper thresholding, selects GFP positive clones only in the current Z-plane.
While the percentage coverage of the anterior portion of the midgut presents a reliable metric for analysing changes to the tumour burden in the ApcRas model, this may not extend to other models where clones are more evenly distributed throughout the midgut. We therefore also developed a macro to measure the entire midgut area, enabling the calculation of the percentage of the entire midgut covered by clones. In order to further increase the high throughput capabilities of this model of CRC, we also developed a macro that quantified the GFP positive area, along with the whole area of objects in the red channel, of consecutive images. This allows extremely large batches of dissected guts, or even whole experimental conditions, to be processed in minutes. Interestingly, while the calculation of the GFP positive area in the anterior region of midguts is significantly higher in the ApcRas conditions than in controls (
Figure 2d), the percent of the entire midgut covered by GFP positive clones shows no significant difference (
Figure 2c). This suggests that while the percentage coverage of the anterior portion of the midgut may be a good metric for analysing the tumour burden in the ApcRas model versus wildtype, the percentage coverage of the whole midgut is not. Therefore, for a high-throughput screen for drugs altering the tumour burden in the ApcRas model, which may affect the distribution of clones across the midgut, or for measurements in any condition where clones are not restricted to one portion of the midgut, other parameters may be more informative.
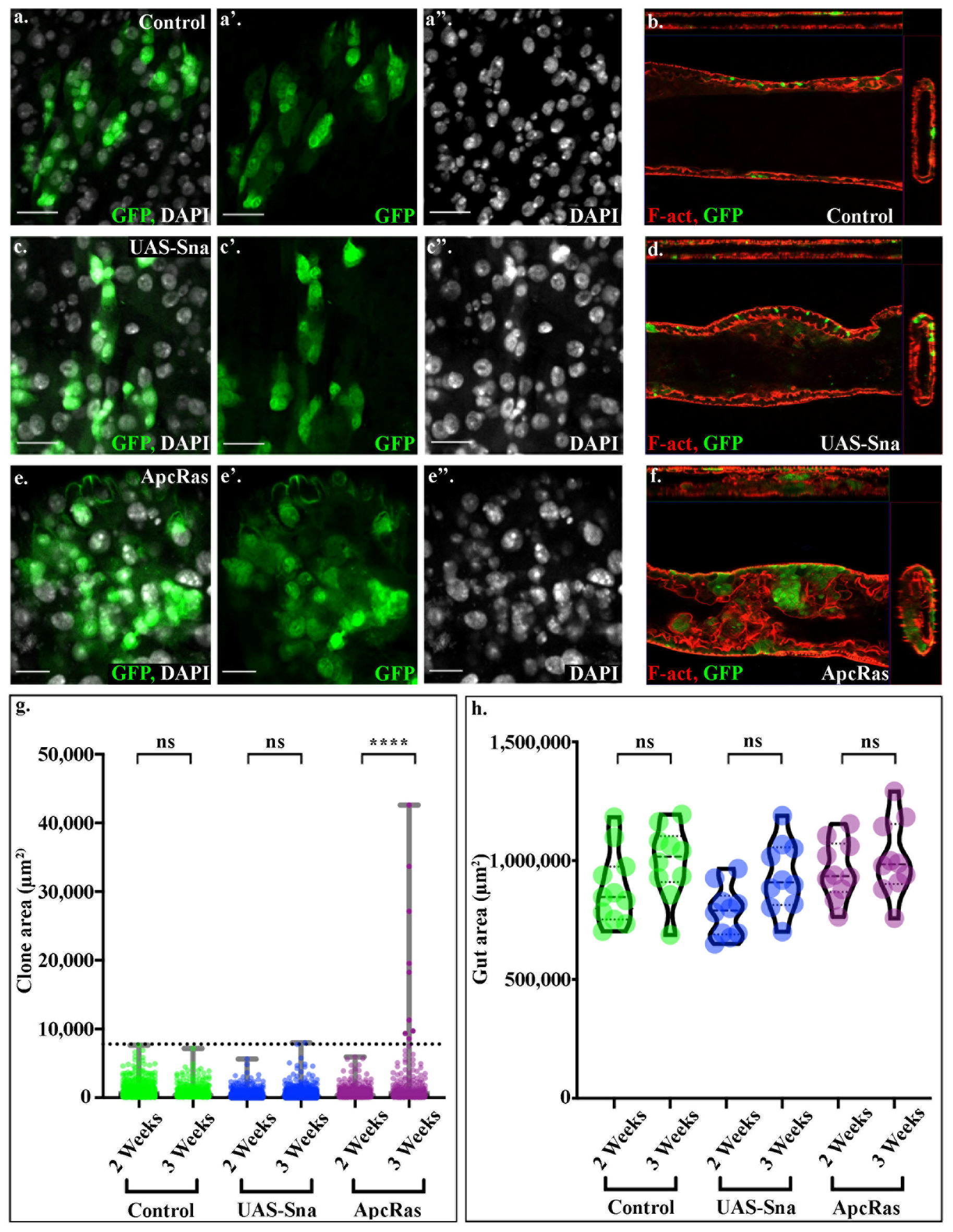
This raises the question as to what quantifiable parameters may determine the difference between a wildtype control clone, a clone that has over-proliferated but not formed a tumour, and a tumour-like clone where cells have lost polarity and are undifferentiated. In previous work, we have shown that the overexpression of Snail in the intestinal stem cells leads to the over-proliferation of cells, but clones do not form tumours [
25]. We therefore decided to look in detail at the cell morphology, and distribution of either control clones, clones over expressing Snail or ApcRas clones (
Figure 3a–f), to identify quantifiable parameters that could be leveraged to distinguish what kind of clone has formed. We found that the tissue architecture, cell organisation and nuclei size are distinct between control clones (
Figure 3a,b), Snail clones (
Figure 3c,d) and ApcRas clones (
Figure 3e,f). The adult
Drosophila intestinal tract shows high levels of cellular heterogeneity, as the epithelium is composed of a range of cell types, as well as cells of varying ages, as the epithelium undergoes continual cell turnover. Regardless of this high heterogeneity, the cell morphology and general tissue architecture in control clones and Snail clones is relatively consistent and well-organised (
Figure 3a–d), while ApcRas clones exhibit poor cell organisation, large variation in cell morphology and shape, increased nuclei density within clones and a huge range of nuclei sizes—characteristics displayed in mammalian epithelial tumours (
Figure 3e,f). A key difference between clones of cells which have overproliferated versus tumour-like clones is an increase in the maximal nuclear size; the maximal nuclear size in control and Snail clones is 420 µm
2, whereas nuclei in ApcRas are on average bigger and can be as large as 980 µm
2 in diameter. While nuclei in ApcRas clones are more packed than in controls (
Figure 3 compare e with a), this is also seen in over proliferating conditions (
Figure 3c). In contrast, while control and Snail clones are always found within a single layer (
Figure 3b,d), ApcRas clones are multi-layered and protrude into the lumen of the midgut (
Figure 3f). However, assessing whether clones have become tumour-like by their maximum nuclear diameter and extent of which they have become multi-layered or protrude into the lumen is very time consuming, as the midgut is a very thick tissue and it requires taking many z-slices in high resolution. Combining this with the need for tile scans to capture the entire length of the midgut, this suggests that while these parameters can be used to assess a few samples, it would not be practical to use them as outputs for a high-throughput screen.
The formation of these large multi-layered tumours, suggested that tumour-like clones could be distinguished from controls and over-proliferating cells by size. Therefore, automated scripts were developed to measure clone area of all clones along the entire midgut. These showed that the large clones over 7800 μm
2, likely formed by the merging of multiple clones, are only seen in ApcRas conditions, and never seen in control or in Snail clones (
Figure 3g). This occurs without any significant changes in the overall gut area (
Figure 3h). Importantly, comparable results were seen whether these were analysed from a tile scan across 3 z-planes of the midgut, or across a z-stack through the entire midgut. Taken together, these results suggest that the clone area is a robust and reliable readout for the presence of tumour-like clones in the midgut and can be assayed and compared using just 3 z-planes.
Once the conditions for screening ApcRas flies for tumour burden had been established, we next wanted to test whether the
Drosophila CRC model responds to drugs currently used to treat human tumours. We decided to test two drugs: Oxaliplatin (Oxal) and 5-Fluoro-5′-Deoxyuridine (5-Fluoro), two chemotherapy drugs that are standard treatments for patients with CRC [
29]. Oxal is a platinum-based DNA crosslinking agent [
30], whereas 5-Fluoro is a Fluorouracil derivative which acts as an antimetabolite by blocking synthesis of thymidine [
31], and both act to block cell division. We reasoned that their efficacy in treating human CRC suggested that they would be good candidates to test our model.
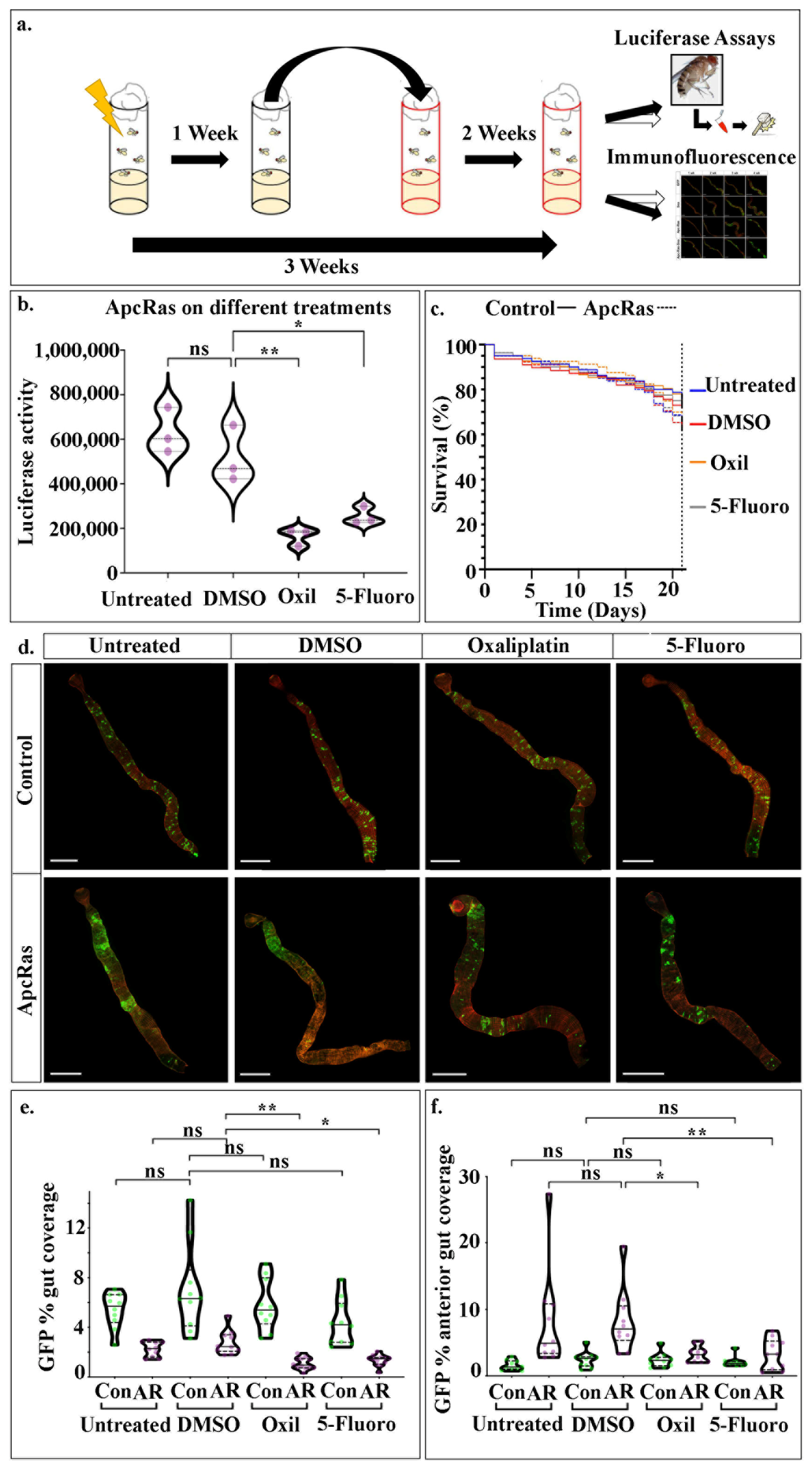
Flies bearing ApcRas clones were induced in adult midguts and allowed to develop for 1 week. The flies were then transferred to new vials containing either food alone (Untreated), food mixed with Oxal or 5-Fluoro or with the additional control of Dimethyl sulfoxide (DMSO) alone, as this was used to dilute the drugs before adding to the food. The drugs were diluted to a final concentration of 100 µM within the food. Flies were left on the food with drugs or control food for 2 weeks, and then assayed for tumour burden by luciferase assays on whole flies (
Figure 4a). These revealed that treatment with either Oxal or 5-Fluoro leads to a large reduction in the overall tumour burden when compared to untreated flies, or flies fed with DMSO alone (
Figure 4b). This demonstrates that
Drosophila CRC flies show a rapid and robust response to treatment with standard human CRC therapeutics. Moreover, this suggests firstly that the
Drosophila CRC model may be a viable screening platform for drugs for treating human CRC. Secondly, as luciferase assays can be performed on whole fly homogenates in 96-well plates, this approach is scalable to high-throughput.
We also followed the % survival of both ApcRas and control flies over the 3 week period as another potential readout for the screen. However, we found only subtle affects with each genotype and treatment: DMSO slightly lowers the survival rate of control flies, and this is partially rescued with feeding on DMSO + Oxal or DMSO+5′Fluoro (
Figure 4c); DMSO also lowers the survival rate of ApcRas flies, and this is also partially rescued by treatment with either Oxal or 5-Fluor, with Oxal bringing the % survival almost as high as control flies fed on DMSO (
Figure 4c). These small changes in survival rates over the 3-week period fits with previous data showing that ApcRas flies display a 50% survival rate at 4 weeks after clone induction [
19], suggesting that flies would need to be monitored for longer than 3 weeks for any effects on survival to be seen. For this to be a robust readout of the screen, it would need to be performed on more flies, over an even longer time course. Taken together, this suggests that luciferase readings on whole flies presents a far more rapid assay to screen for putative CRC treatments in flies.
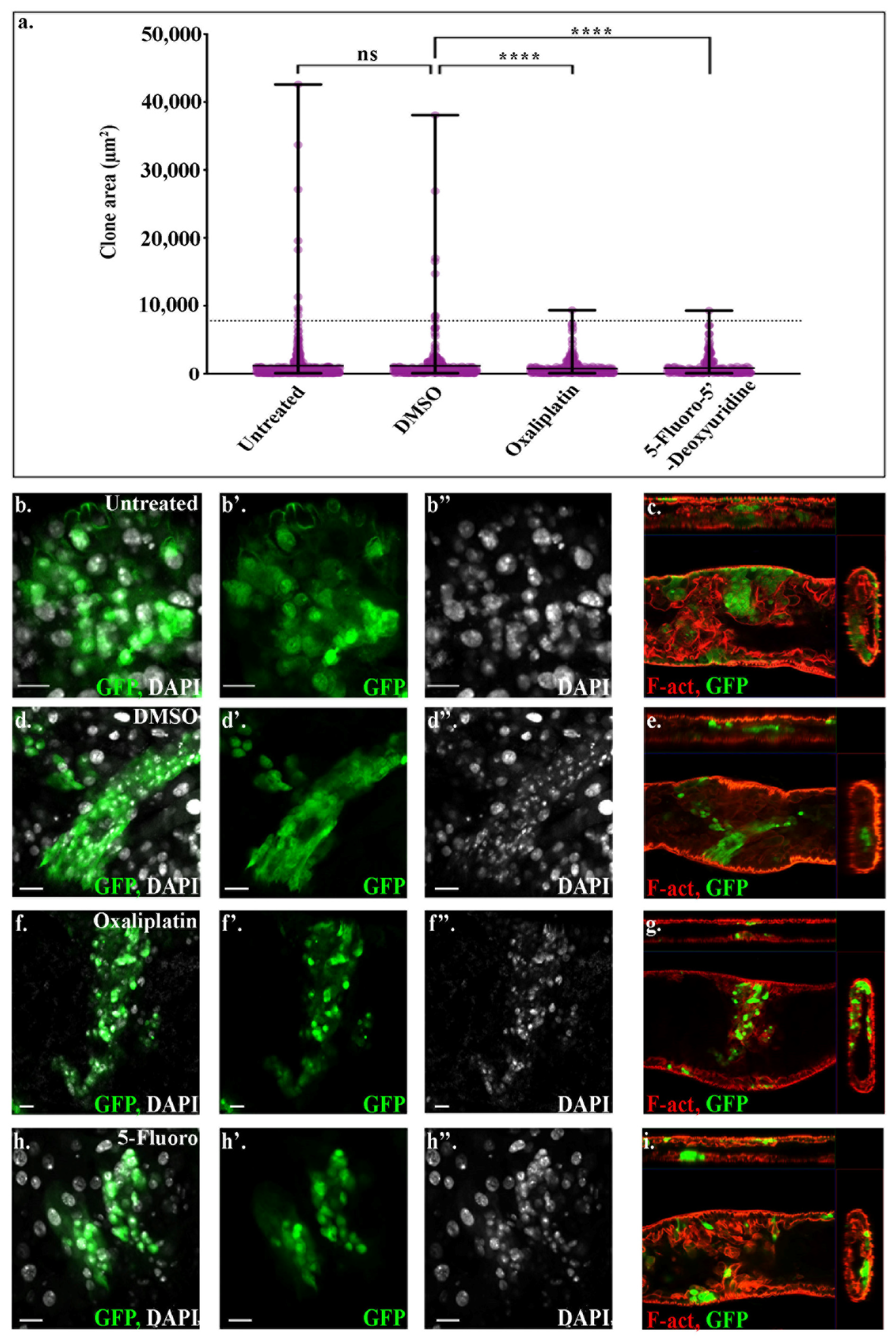
We next sought to characterise in more detail how flies bearing ApcRas clones respond to treatment with Oxal or 5-Fluoro and determine if their effects are specific to the tumour-like ApcRas clones by comparing with control clones. To do this, we studied the behaviour of both control and ApcRas clones in the midguts of flies fed with the different conditions, by dissecting out their midguts after 3 weeks and staining them for GFP and phalloidin, which labels F-actin and can be used to visualise the outline and overall organisation of the midgut. We found that the midgut morphology and size was not affected and the distribution and size of control clones were the same in all conditions (
Figure 4d). In contrast, while the midguts of ApcRas flies raised on untreated and DMSO food had large tumour-like clones in the anterior region of the midguts, the clones in flies fed with Oxal and 5-Fluoro were greatly reduced (
Figure 4d). Thus, the reduction in clone size and distribution appears to be specific to clones with tumour-like properties.
We quantified the behaviour of clones in the different conditions by dissecting, fixing and immunostaining the midguts from these flies, and then analysing the clonal area as a percentage of the anterior region of the midgut or of the whole midgut. Both analyses revealed a reduction in coverage by GFP positive clones by around 50% when the flies were treated with either Oxal or 5-Fluoro (
Figure 4d,e). The fact that no reduction was seen in the coverage by control clones (
Figure 4e,f) suggests that treatment with the standard chemotherapies used to treat human CRC specifically target the tumour-like ApcRas clones, without causing any significant changes to normal wildtype cells.
To look in more detail at the behaviour of ApcRas clones upon treatment with Oxal or 5-Fluoro, we next used our automated scripts to analyse the area of all the clones across the entire midgut in ApcRas conditions on each treatment. We found that many very large clones over 7800 μm
2 in size are found in both untreated ApcRas flies, or ApcRas flies treated with DMSO alone. Strikingly, after ApcRas flies have been treated with either Oxal or 5-Fluoro, almost no flies are found to contain clones over the 7800 μm
2 threshold, further confirming the efficacy of these treatments (
Figure 5a). In support of this, we found that other tumour-like characteristics of ApcRas clones are reduced (
Figure 5b–i); the maximum nuclear size within a clone was reduced from 986 μm
2 to ~680 μm
2, and while cross sections of midguts showed that there was still some multilayering of cells within ApcRas clones, they never caused the complete luminal blockages seen when these tumour-like clones were left to grow untreated (
Figure 5 compare
Figure 5g,i with
Figure 5c,e). Taken together, these results suggest that while treatment with CRC therapeutics did not completely abrogate tumour growth, the tumour burden was dramatically reduced, and this was accompanied by a reduction in the appearance of clones bulging into the lumen of the gut.
3. Discussion
To date, current drug discovery pipelines have had very limited success in delivering drugs identified in vitro, into the clinic for anti-cancer therapy [
32,
33]. With research in
Drosophila providing the first glimpse into the mechanism of action of many human cancer-related proteins, it is now emerging as an important new platform for anti-cancer drug discovery [
34]. Rapid and cost effective, its key advantage over classic in vitro screens is the identification of drugs that can specifically target tumours in vivo. This permits assessment of drug absorption and toxicity, reducing the possibility of false positives, as well as assaying drug effects on tumour cells which are maintaining key interactions occurring during normal disease progression, such as with the tumour microenvironment. Moreover, the fast genetic manipulation of
Drosophila presents the possibility to personalise transgenic cancer models, in which mutations are directly based on individual tumour genome profiles. Such models have been used to analyse a collection of drugs and assess the mechanisms of emerging drug resistance [
35]. Remarkably, the generation of a personalised model of a treatment-resistant metastatic KRAS-mutant colorectal cancer in
Drosophila larvae has led to the identification of a drug combination, trametinib and zoledronate, that has been used as a treatment for the patient [
36], highlighting the potential of
Drosophila as a model for personalised medicine. Here, we have developed sensitive high-throughput assays, to rapidly and accurately assay for tumour burden in a
Drosophila model for benign CRC, that has previously been shown to recapitulate many hallmarks of the human disease [
19]. We show that it responds to treatment with standard chemotherapies used to treat human CRC, suggesting that compounds identified in future screens in this model may be transferable to humans.
Introducing two reporters into the system, firefly luciferase and GFP, makes the Drosophila ApcRas model amenable to two distinct methods of screening. The first, assaying for luciferase activity in whole flies, enables high-throughput batch processing of whole flies, to identify overall changes in tumour burden. The second method relies on the visualisation and quantification of the GFP reporter. It not only enables the verification of changes detected by luciferase assays, but gives a more in-depth analysis of individual clone behaviour in a semi high-throughput capacity. Excitingly, this suggests that screening batches of flies with luciferase assays could work as a primary screen, and positive candidates could be followed up in a second screen where midguts are dissected and analysed using our high-throughput scripts.
Here we show that a Drosophila model for benign CRC rapidly responds to treatment with standard chemotherapy drugs, and that the response can reliably and reproducibly be assessed in a high-throughput manner. This process is highly scalable to the screening of thousands of flies, opening the door for drug screens to test either single compounds, or multiple compounds in combination. As Drosophila are highly genetically tractable, once a drug or drug combination has been identified, this could later be combined with a genetic screen to identify the drugs in vivo targets. Given the current need for new therapies for CRC that have less side effects and are more effective than conventional ones, the ApcRas Drosophila model presents an exciting new avenue, at a fraction of both cost and time of vertebrate models.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




