
Extracellular Vesicles in Synovial Fluid from Rheumatoid Arthritis Patients Contain miRNAs with Capacity to Modulate Inflammation

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
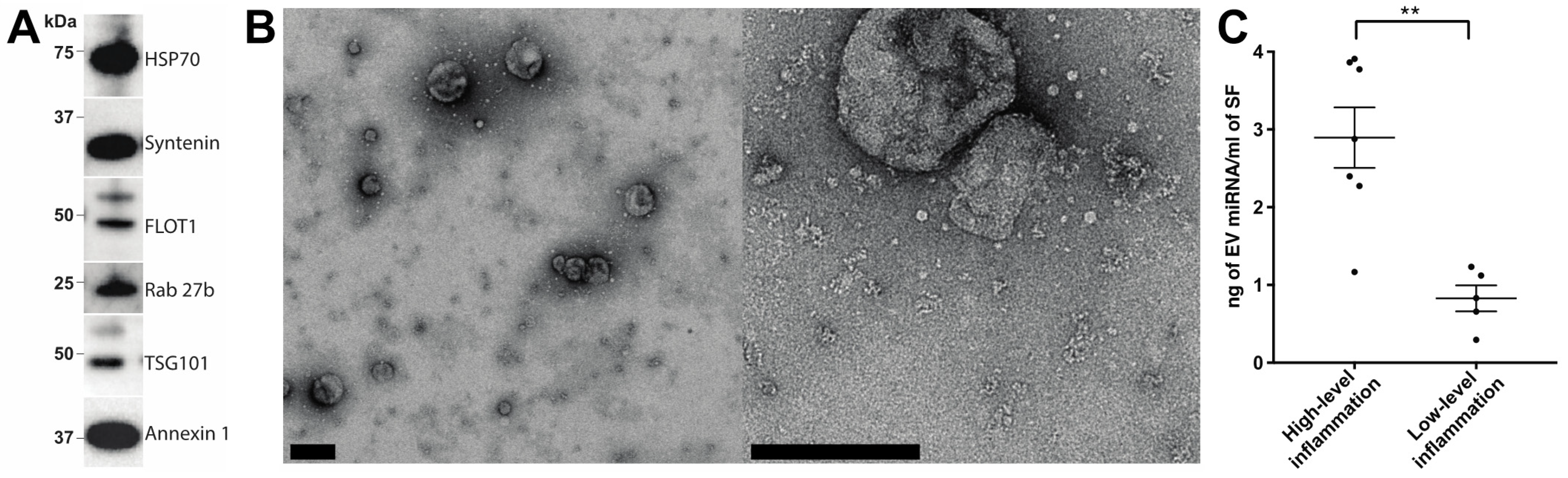
2.1. Characterisation of EV Isolation
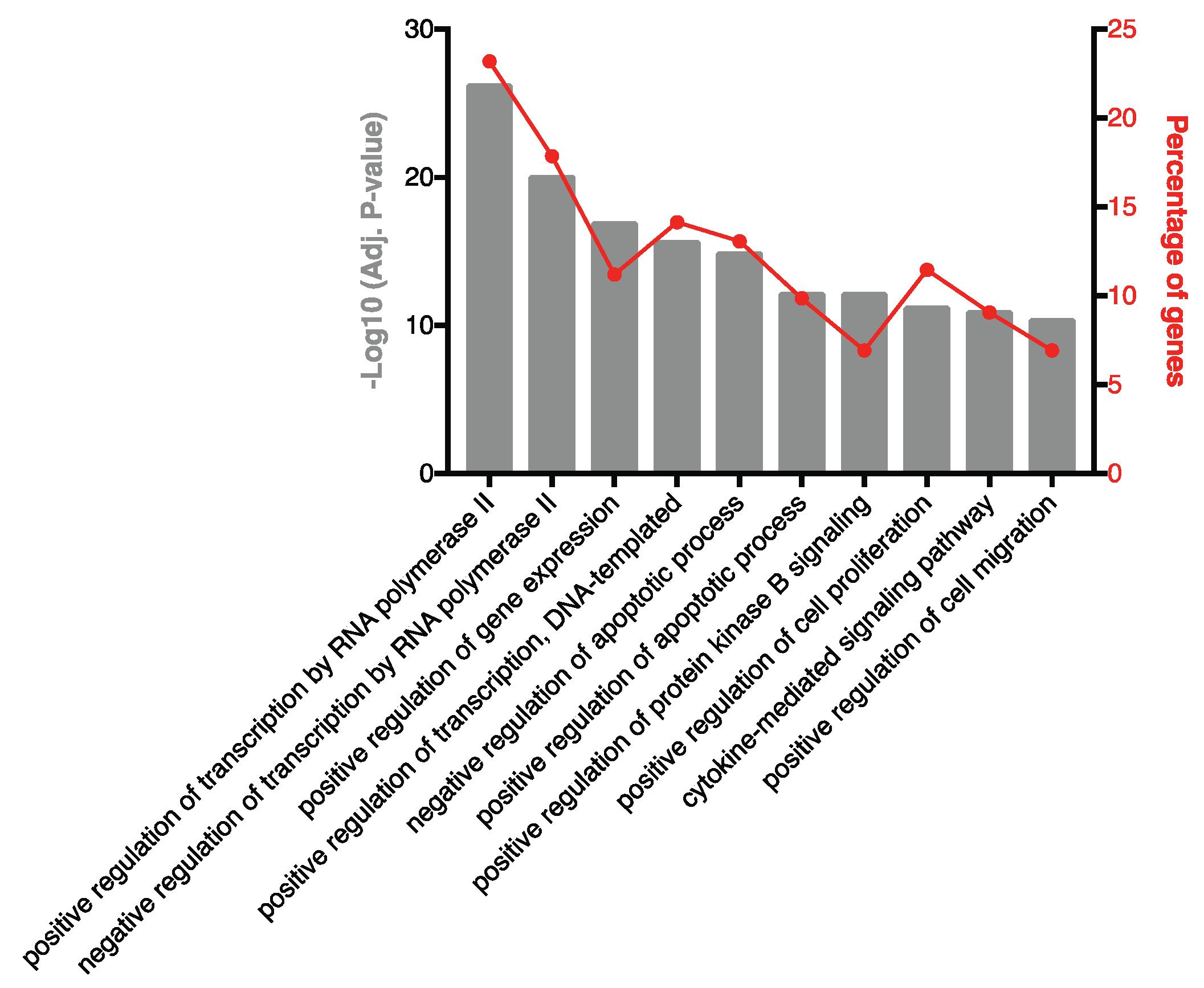
2.2. Highly Ranked SF EV miRNAs Target Immunomodulatory SF EV Proteins
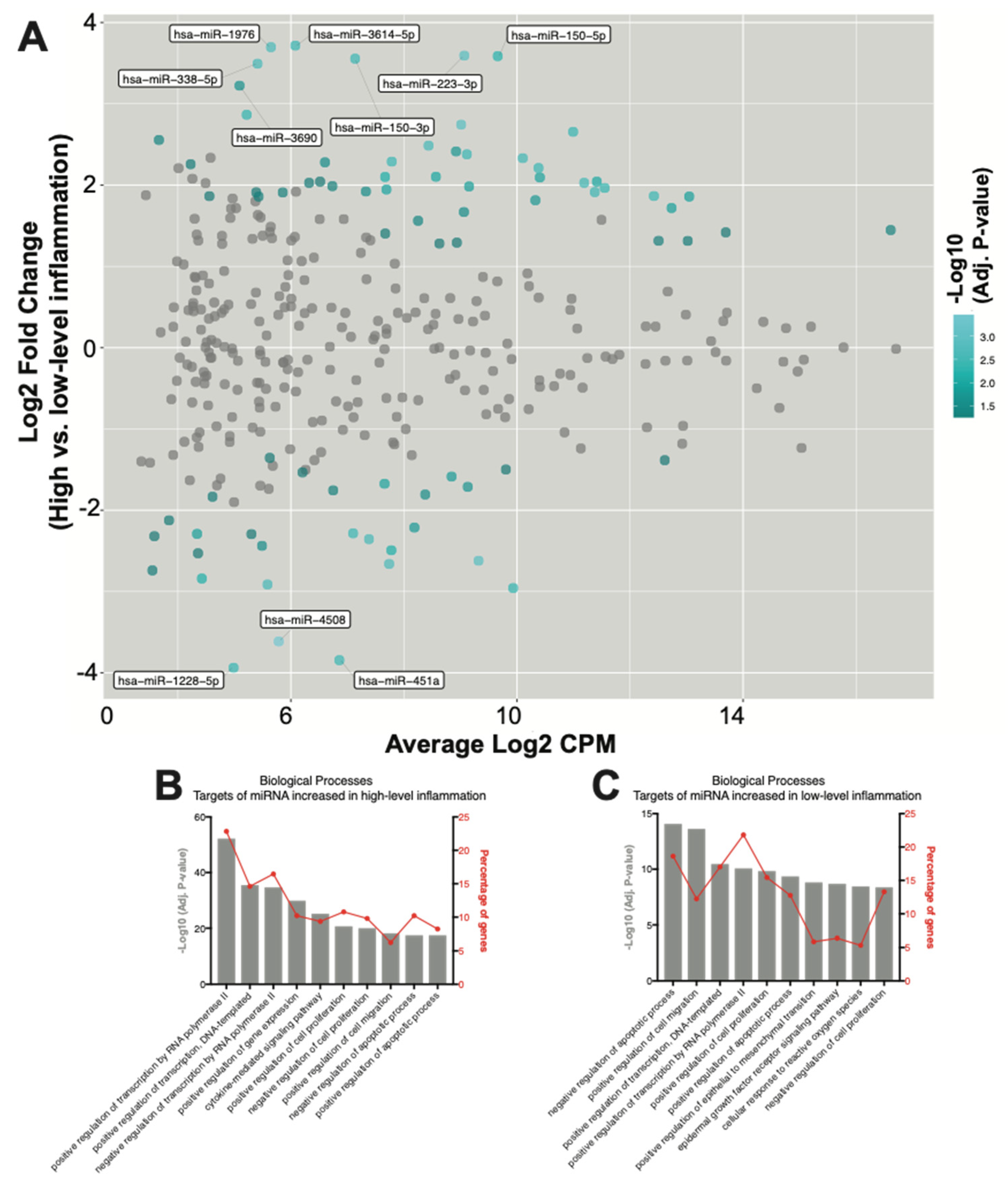
2.3. Seventy-Eight SF EV miRNAs Are Differentially Expressed between RA Patients with High- and Low-Grade Inflammation
3. Discussion
4. Materials and Methods
4.1. Patient Details, Collection and Storage of Human Synovial Fluid
4.2. Sample Preparation and EV Isolation
4.3. Transmission Electron Microscopy
4.4. Western Blot Analysis
4.5. Enrichment and Separation of Small RNA
4.6. miRNA Library Preparation and Sequencing
4.7. Bioinformatic Analysis of Sequencing Data
4.8. Ranking of Prevalent miRNAs
4.9. Pathway Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EV | Extracellular vesicle |
| miRNA | microRNA |
| RA | Rheumatoid arthritis |
| SEC | Size exclusion chromatography |
| SF | Synovial fluid |
References
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Foers, A.D.; Cheng, L.; Hill, A.F.; Wicks, I.P.; Pang, K.C. Review: Extracellular Vesicles in Joint Inflammation. Arthritis Rheumatol. 2017, 69, 1350–1362. [Google Scholar] [CrossRef]
- Malda, J.; Boere, J.; Van De Lest, C.H.A.; Van Weeren, P.R.; Wauben, M.H.M. Extracellular vesicles—New tool for joint repair and regeneration. Nat. Rev. Rheumatol. 2016, 12, 243–249. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, K.; Yoshitomi, H.; Tanida, S.; Ishikawa, M.; Nishitani, K.; Ito, H.; Nakamura, T. Plasma and synovial fluid microRNAs as potential biomarkers of rheumatoid arthritis and osteoarthritis. Arthritis Res. Ther. 2010, 12, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wan, Y.; Guo, Q.; Zou, L.; Zhang, J.; Fang, Y.; Zhang, J.; Zhang, J.; Fu, X.; Liu, H.; et al. Altered microRNA expression profile with miR-146a upregulation in CD4+ T cells from patients with rheumatoid arthritis. Arthritis Res. Ther. 2010, 12, R81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okoye, I.S.; Coomes, S.M.; Pelly, V.S.; Czieso, S.; Papayannopoulos, V.; Tolmachova, T.; Seabra, M.C.; Wilson, M.S. MicroRNA-containing T-regulatory-cell-derived exosomes suppress pathogenic T helper 1 cells. Immunity 2014, 41, 89–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, S.; Young, N.A.; Mignot, G.; Bach, J.-M. Epigenetic Mechanisms in Immune Disease: The Significance of Toll-Like Receptor-Binding Extracellular Vesicle-Encapsulated microRNA. Front. Genet. 2020, 11, 578335. [Google Scholar] [CrossRef] [PubMed]
- Bernard, M.A.; Zhao, H.; Yue, S.C.; Anandaiah, A.; Koziel, H.; Tachado, S.D. Novel HIV-1 MiRNAs Stimulate TNFα Release in Human Macrophages via TLR8 Signaling Pathway. PLoS ONE 2014, 9, e106006. [Google Scholar] [CrossRef] [PubMed]
- Foers, A.D.; Chatfield, S.; Dagley, L.F.; Scicluna, B.J.; Webb, A.I.; Cheng, L.; Hill, A.F.; Wicks, I.P.; Pang, K.C. Enrichment of extracellular vesicles from human synovial fluid using size exclusion chromatography. J. Extracell. Vesicles 2018, 7, 1490145. [Google Scholar] [CrossRef]
- Foers, A.D.; Dagley, L.F.; Chatfield, S.; Webb, A.I.; Cheng, L.; Hill, A.F.; Wicks, I.P.; Pang, K.C. Proteomic analysis of extracellular vesicles reveals an immunogenic cargo in rheumatoid arthritis synovial fluid. Clin. Transl. Immunol. 2020, 9, e1185. [Google Scholar] [CrossRef]
- Erlandsson, M.C.; Silfverswärd, S.T.; Nadali, M.; Turkkila, M.; Svensson, M.N.; Jonsson, I.-M.; Andersson, K.M.; Bokarewa, M.I. IGF-1R signalling contributes to IL-6 production and T cell dependent inflammation in rheumatoid arthritis. Biochim. Biophys. Acta 2017, 1863, 2158–2170. [Google Scholar] [CrossRef] [PubMed]
- Tsushima, H.; Morimoto, S.; Fujishiro, M.; Yoshida, Y.; Hayakawa, K.; Hirai, T.; Miyashita, T.; Ikeda, K.; Yamaji, K.; Takamori, K.; et al. Kinase inhibitors of the IGF-1R as a potential therapeutic agent for rheumatoid arthritis. Autoimmunity 2017, 50, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Oike, T.; Sato, Y.; Kobayashi, T.; Miyamoto, K.; Nakamura, S.; Kaneko, Y.; Kobayashi, S.; Harato, K.; Saya, H.; Matsumoto, M.; et al. Stat3 as a potential therapeutic target for rheumatoid arthritis. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarino, A.V.; Kanno, Y.; O’Shea, A.V.V.Y.K.J.J. Mechanisms and consequences of Jak–STAT signaling in the immune system. Nat. Immunol. 2017, 18, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Minna, E.; Romeo, P.; Dugo, M.; De Cecco, L.; Todoerti, K.; Pilotti, S.; Perrone, F.; Seregni, E.; Agnelli, L.; Neri, A.; et al. miR-451a is underexpressed and targets AKT/mTOR pathway in papillary thyroid carcinoma. Oncotarget 2016, 7, 12731–12747. [Google Scholar] [CrossRef]
- Rosenberger, C.M.; Podyminogin, R.L.; Navarro, G.; Zhao, G.-W.; Askovich, P.S.; Weiss, M.J.; Aderem, A. miR-451 Regulates Dendritic Cell Cytokine Responses to Influenza Infection. J. Immunol. 2012, 189, 5965–5975. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Wang, K.; Li, Y.; Xia, N.; Nie, S.; Lv, B.; Zhang, M.; Tu, X.; Li, Q.; Tang, T.; et al. Down-regulation of microRNA-451a facilitates the activation and proliferation of CD4+ T cells by targeting Myc in patients with dilated cardiomyopathy. J. Biol. Chem. 2017, 292, 6004–6013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugatani, T.; Hruska, K.A. MicroRNA-223 is a key factor in osteoclast differentiation. J. Cell. Biochem. 2007, 101, 996–999. [Google Scholar] [CrossRef]
- Sugatani, T.; Hruska, K.A. Impaired Micro-RNA Pathways Diminish Osteoclast Differentiation and Function. J. Biol. Chem. 2009, 284, 4667–4678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-T.; Chen, S.-Y.; Wang, C.-R.; Liu, M.-F.; Lin, C.-C.; Jou, I.-M.; Shiau, A.-L.; Wu, C.-L. Brief Report: Amelioration of collagen-induced arthritis in mice by lentivirus-mediated silencing of microRNA-223. Arthritis Rheum. 2012, 64, 3240–3245. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, H.; Liu, Y.; Song, Y.; Lai, L.; Han, Q.; Cao, X.; Wang, Q. Inducible MicroRNA-223 Down-Regulation Promotes TLR-Triggered IL-6 and IL-1β Production in Macrophages by Targeting STAT3. PLoS ONE 2012, 7, e42971. [Google Scholar] [CrossRef] [Green Version]
- Johnnidis, J.B.; Harris, M.H.; Wheeler, R.T.; Stehling-Sun, S.; Lam, M.H.; Kirak, O.; Brummelkamp, T.R.; Fleming, M.D.; Camargo, F.D. Regulation of progenitor cell proliferation and granulocyte function by microRNA-223. Nature 2008, 451, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc. Natl. Acad. Sci. USA 2012, 109, E2110–E2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, N.A.; Valiente, G.R.; Hampton, J.M.; Wu, L.-C.; Burd, C.J.; Willis, W.L.; Bruss, M.; Steigelman, H.; Gotsatsenko, M.; Amici, S.A.; et al. Estrogen-regulated STAT1 activation promotes TLR8 expression to facilitate signaling via microRNA-21 in systemic lupus erythematosus. Clin. Immunol. 2017, 176, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.M.; Krüger, C.; Park, B.; Derkow, K.; Rosenberger, K.; Baumgart, J.; Trimbuch, T.; Eom, G.; Hinz, M.; Kaul, D.; et al. An unconventional role for miRNA: Let-7 activates Toll-like receptor 7 and causes neurodegeneration. Nat. Neurosci. 2012, 15, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Chen, Z.; Essani, A.B.; Elshabrawy, H.A.; Volin, M.V.; Volkov, S.; Swedler, W.; Arami, S.; Sweiss, N.; Shahrara, S. Identification of a Novel Toll-like Receptor 7 Endogenous Ligand in Rheumatoid Arthritis Synovial Fluid That Can Provoke Arthritic Joint Inflammation. Arthritis Rheumatol. 2016, 68, 1099–1110. [Google Scholar] [PubMed]
- Feng, Y.; Zou, L.; Yan, F.; Chen, H.; Xu, G.; Jian, W.; Cui, P.; Chao, W. Extracellular MicroRNAs Induce Potent Innate Immune Responses via TLR7/MyD88-Dependent Mechanisms. J. Immunol. 2017, 199, 2106–2117. [Google Scholar] [CrossRef]
- Van Deun, J.; Mestdagh, P.; Agostinis, P.; Akay, Ö.; Anand, S.; Anckaert, J.; Martinez, Z.A.; Baetens, T.; Beghein, E.; Bertier, L.; et al. EV-TRACK: Transparent reporting and centralizing knowledge in extracellular vesicle research. Nat. Methods 2017, 14, 228–232. [Google Scholar] [CrossRef]
- Shmerling, R.H.; Delbanco, T.L.; Tosteson, A.N.; Trentham, D.E. Synovial fluid tests. What should be ordered? JAMA 1990, 264, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. The Subread aligner: Fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 2013, 41, e108. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathan, M.; Keerthikumar, S.; Ang, C.-S.; Gangoda, L.; Quek, C.Y.; Williamson, N.A.; Mouradov, D.; Sieber, O.M.; Simpson, R.J.; Salim, A.; et al. FunRich: An open access standalone functional enrichment and interaction network analysis tool. Proteomics 2015, 15, 2597–2601. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.H.; Shrestha, S.; Yang, C.D.; Chang, N.W.; Lin, Y.L.; Liao, K.W.; Huang, W.C.; Sun, T.H.; Tu, S.J.; Lee, W.H.; et al. miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018, 46, D296–D302. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| RA (High-Grade Inflammation) | RA (Low-Grade Inflammation) | p-Value | |
|---|---|---|---|
| n | 7 | 5 | - |
| Age—mean (s.d.) | 64.4 (11.8) | 67.6 (10.0) | 0.64 |
| Sex—number of females/males | 3/4 | 2/3 | >0.99 |
| White cell count—mean (s.d.) cells µL−1 | 8940 * | 171.8 | <0.001 |
| Anti-citrullinated protein antibody (% positive) | 71% | 20% | 0.24 |
| Rheumatoid factor (% positive) | 86% | 60% | 0.52 |
| Disease Activity Score 28—median (range) | 4.7 (3.31–5.41) | 3.5 (2.74–5.0) | 0.15 |
| C-reactive protein—median (range) mg L−1 | 20 (6–164) | 2 (1.4–2) | 0.16 |
| Rank | miRNA | Average Expression (CPM) |
|---|---|---|
| 1 | hsa-miR-100-5p | 132,441 |
| 2 | hsa-miR-21-5p | 179,929 |
| 3 | hsa-miR-148a-3p | 86,059 |
| 4 | hsa-let-7a-5p | 43,627 |
| 5 | hsa-miR-92a-3p | 45,881 |
| 6 | hsa-let-7b-5p | 31,280 |
| 7 | hsa-miR-10b-5p | 35,499 |
| 8 | hsa-miR-99b-5p | 31,569 |
| 9 | hsa-miR-26a-5p | 34,975 |
| 10 | hsa-miR-99a-5p | 45,879 |
| Target Gene ID | Target Gene Name | miRNA Regulators |
|---|---|---|
| IGF1R | Insulin like growth factor 1 receptor | hsa-let-7b-5p hsa-miR-100-5p hsa-miR-21-5p hsa-miR-99a-5p hsa-miR-99b-5p |
| CCND2 | Cyclin D2 | hsa-let-7a-3p hsa-let-7a-5p hsa-let-7b-5p hsa-miR-26a-5p |
| E2F2 | E2F transcription factor 2 | hsa-let-7a-3p hsa-let-7a-5p hsa-let-7b-5p hsa-miR-26a-5p |
| PTEN | Phosphatase and tensin homolog | hsa-miR-10b-5p hsa-miR-21-5p hsa-miR-26a-5p hsa-miR-92a-3p |
| STAT3 | Signal transducer and activator of transcription 3 | hsa-let-7a-5p hsa-miR-148a-3p hsa-miR-21-5p hsa-miR-92a-3p |
| miRNA | Average Read Count (Log2 CPM) | Log2 Fold Change(High- vs. Low−Grade Inflammation) | Adjusted p-Value |
|---|---|---|---|
| hsa-miR-4508 | 5.78 | −3.61 | 3.94 × 10−4 |
| hsa-miR-223-3p | 9.07 | 3.59 | 6.42 × 10−4 |
| hsa-miR-3529-3p | 9.01 | 2.74 | 6.42 × 10−4 |
| hsa-miR-615-3p | 9.32 | −2.62 | 6.42 × 10−4 |
| hsa-miR-1976 | 5.65 | 3.70 | 1.15 × 10−3 |
| hsa-miR-543 | 7.74 | −2.66 | 1.15 × 10−3 |
| hsa-miR-338-5p | 5.41 | 3.49 | 1.36 × 10−3 |
| hsa-miR-146b-3p | 8.43 | 2.49 | 1.36 × 10−3 |
| hsa-miR-433-3p | 7.38 | −2.36 | 1.36 × 10−3 |
| hsa-miR-485-3p | 5.59 | −2.91 | 1.36 × 10−3 |
| hsa-miR-101-3p | 10.10 | 2.33 | 1.52 × 10−3 |
| hsa-miR-27a-5p | 11.19 | 2.03 | 1.53 × 10−3 |
| hsa-miR-361-3p | 11.38 | 1.91 | 1.76 × 10−3 |
| hsa-miR-3614-5p | 6.08 | 3.72 | 1.86 × 10−3 |
| hsa-miR-150-3p | 7.14 | 3.55 | 1.89 × 10−3 |
| hsa-miR-223-5p | 9.11 | 2.38 | 1.89 × 10−3 |
| hsa-miR-142-5p | 7.78 | 2.29 | 1.89 × 10−3 |
| hsa-miR-106b-3p | 10.38 | 2.21 | 1.89 × 10−3 |
| hsa-miR-28-3p | 12.42 | 1.87 | 1.89 × 10−3 |
| hsa-miR-455-5p | 7.10 | −2.28 | 1.89 × 10−3 |
| hsa-miR-451a | 6.86 | −3.84 | 1.89 × 10−3 |
| hsa-miR-143-3p | 10.99 | 2.66 | 2.04 × 10−3 |
| hsa-miR-1228-5p | 4.99 | −3.94 | 2.11 × 10−3 |
| hsa-miR-30e-3p | 11.55 | 1.96 | 2.25 × 10−3 |
| hsa-miR-486-5p | 9.93 | −2.96 | 2.30 × 10−3 |
| hsa-miR-1273h-3p | 5.22 | 2.87 | 2.56 × 10−3 |
| hsa-miR-150-5p | 9.66 | 3.59 | 3.01 × 10−3 |
| hsa-miR-378c | 7.67 | 2.10 | 3.61 × 10−3 |
| hsa-miR-92b-5p | 7.78 | −2.49 | 3.61 × 10−3 |
| hsa-miR-4448 | 4.42 | −2.84 | 3.61 × 10−3 |
| hsa-miR-103b | 8.57 | 2.10 | 4.76 × 10−3 |
| hsa-miR-941 | 11.41 | 2.04 | 4.76 × 10−3 |
| hsa-miR-103a-3p | 7.69 | 1.95 | 5.24 × 10−3 |
| hsa-miR-1246 | 10.40 | 2.09 | 5.64 × 10−3 |
| hsa-miR-125b-1-3p | 7.66 | −1.67 | 5.64 × 10−3 |
| hsa-miR-769-5p | 9.15 | 1.98 | 6.68 × 10−3 |
| hsa-miR-378a-3p | 13.05 | 1.86 | 6.68 × 10−3 |
| hsa-miR-140-3p | 12.73 | 1.72 | 6.68 × 10−3 |
| hsa-miR-214-5p | 4.34 | −2.29 | 6.68 × 10−3 |
| hsa-miR-574-3p | 8.84 | −1.59 | 7.83 × 10−3 |
| hsa-miR-1180-3p | 8.18 | −2.21 | 8.45 × 10−3 |
| hsa-miR-155-5p | 10.33 | 1.81 | 8.69 × 10−3 |
| hsa-miR-629-5p | 8.92 | 2.41 | 9.41 × 10−3 |
| hsa-miR-328-3p | 9.13 | −1.71 | 1.05 × 10−2 |
| hsa-miR-3690 | 5.09 | 3.22 | 1.18 × 10−2 |
| hsa-miR-7704 | 8.38 | −1.81 | 1.48 × 10−2 |
| hsa-miR-221-5p | 9.06 | 1.67 | 1.51 × 10−2 |
| hsa-miR-486-3p | 5.49 | −2.44 | 1.51 × 10−2 |
| hsa-miR-589-5p | 6.60 | 2.28 | 1.56 × 10−2 |
| hsa-miR-142-3p | 7.33 | 1.92 | 1.56 × 10−2 |
| hsa-miR-192-5p | 8.25 | 1.56 | 1.98 × 10−2 |
| hsa-miR-618 | 3.67 | 2.55 | 2.18 × 10−2 |
| hsa-miR-21-5p | 16.61 | 1.45 | 2.22 × 10−2 |
| hsa-miR-21-3p | 6.73 | 1.99 | 2.57 × 10−2 |
| hsa-miR-345-5p | 5.86 | 1.91 | 2.59 × 10−2 |
| hsa-miR-214-3p | 6.75 | −1.76 | 2.59 × 10−2 |
| hsa-miR-532-5p | 12.50 | 1.31 | 2.70 × 10−2 |
| hsa-miR-378e | 5.43 | 1.86 | 2.86 × 10−2 |
| hsa-miR-185-3p | 6.32 | 2.03 | 3.05 × 10−2 |
| hsa-miR-6787-3p | 3.55 | −2.74 | 3.07 × 10−2 |
| hsa-miR-503-5p | 4.23 | 2.26 | 3.11 × 10−2 |
| hsa-miR-25-3p | 13.02 | 1.31 | 3.11 × 10−2 |
| hsa-miR-3120-5p | 4.61 | −1.83 | 3.24 × 10−2 |
| hsa-miR-203b-5p | 3.84 | −2.12 | 3.35 × 10−2 |
| hsa-miR-500a-3p | 7.67 | 1.40 | 3.42 × 10−2 |
| hsa-miR-7-5p | 4.56 | 1.86 | 3.56 × 10−2 |
| hsa-miR-3622a-5p | 4.36 | −2.53 | 3.56 × 10−2 |
| hsa-miR-365a-5p | 5.30 | −2.29 | 3.76 × 10−2 |
| hsa-miR-23b-3p | 9.80 | −1.50 | 3.85 × 10−2 |
| hsa-miR-424-3p | 6.52 | 2.04 | 4.29 × 10−2 |
| hsa-miR-501-3p | 8.94 | 1.29 | 4.30 × 10−2 |
| hsa-miR-92b-3p | 12.61 | −1.38 | 4.37 × 10−2 |
| hsa-miR-146b-5p | 13.69 | 1.42 | 4.42 × 10−2 |
| hsa-miR-185-5p | 8.63 | 1.28 | 4.48 × 10−2 |
| hsa-miR-23b-5p | 6.21 | −1.53 | 4.58 × 10−2 |
| hsa-miR-378f | 5.39 | 1.91 | 4.73 × 10−2 |
| hsa-miR-27b-5p | 5.63 | −1.36 | 4.76 × 10−2 |
| hsa-miR-1306-5p | 3.58 | −2.32 | 4.98 × 10−2 |
| Target Gene ID | Target Gene Name | miRNA Regulators |
|---|---|---|
| IGF1R | Insulin like growth factor 1 receptor | hsa-miR-143-3p hsa-miR-150-3p hsa-miR-185-5p hsa-miR-21-5p hsa-miR-223-3p hsa-miR-223-5p hsa-miR-378a-3p hsa-miR-503-5p hsa-miR-7-5p |
| PTEN | Phosphatase and tensin homolog | hsa-miR-103a-3p hsa-miR-106b-3p hsa-miR-142-5p hsa-miR-155-5p hsa-miR-21-3p hsa-miR-21-5p hsa-miR-25-3p |
| VEGFA | Vascular endothelial growth factor A | hsa-miR-101-3p hsa-miR-150-5p hsa-miR-185-5p hsa-miR-21-5p hsa-miR-378a-3p hsa-miR-503-5p |
| BCL2 | BCL2, apoptosis regulator | hsa-miR-143-3p hsa-miR-192-5p hsa-miR-21-5p hsa-miR-503-5p hsa-miR-7-5p |
| FBXW7 | F-box and WD repeat domain containing 7 | hsa-miR-155-5p hsa-miR-223-3p hsa-miR-223-5p hsa-miR-25-3p hsa-miR-503-5p |
| MYB | MYB proto-oncogene, transcription factor | hsa-miR-103a-3p hsa-miR-150-3p hsa-miR-150-5p hsa-miR-155-5p hsa-miR-503-5p |
| EGFR | Epidermal growth factor receptor | hsa-miR-146b-5p hsa-miR-21-5p hsa-miR-27a-5p hsa-miR-7-5p |
| RAC1 | Rac family small GTPase 1 | hsa-miR-101-3p hsa-miR-142-3p hsa-miR-142-5p hsa-miR-155-5p |
| TP53 | Tumor protein p53 | hsa-miR-150-3p hsa-miR-150-5p hsa-miR-25-3p hsa-miR-28-3p |
| ZEB1 | Zinc finger E-box binding homeobox 1 | hsa-miR-101-3p hsa-miR-142-5p hsa-miR-150-5p hsa-miR-223-3p |
| Target Gene ID | Target Gene Name | miRNA Regulators |
|---|---|---|
| PTEN | Phosphatase and tensin homolog | hsa-miR-214-3p hsa-miR-23b-3p hsa-miR-486-5p hsa-miR-92b-3p |
| TP53 | Tumor protein p53 | hsa-miR-125b-1-3p hsa-miR-214-3p hsa-miR-214-5p |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foers, A.D.; Garnham, A.L.; Chatfield, S.; Smyth, G.K.; Cheng, L.; Hill, A.F.; Wicks, I.P.; Pang, K.C. Extracellular Vesicles in Synovial Fluid from Rheumatoid Arthritis Patients Contain miRNAs with Capacity to Modulate Inflammation. Int. J. Mol. Sci. 2021, 22, 4910. https://doi.org/10.3390/ijms22094910
Foers AD, Garnham AL, Chatfield S, Smyth GK, Cheng L, Hill AF, Wicks IP, Pang KC. Extracellular Vesicles in Synovial Fluid from Rheumatoid Arthritis Patients Contain miRNAs with Capacity to Modulate Inflammation. International Journal of Molecular Sciences. 2021; 22(9):4910. https://doi.org/10.3390/ijms22094910
Chicago/Turabian StyleFoers, Andrew D., Alexandra L. Garnham, Simon Chatfield, Gordon K. Smyth, Lesley Cheng, Andrew F. Hill, Ian P. Wicks, and Ken C. Pang. 2021. "Extracellular Vesicles in Synovial Fluid from Rheumatoid Arthritis Patients Contain miRNAs with Capacity to Modulate Inflammation" International Journal of Molecular Sciences 22, no. 9: 4910. https://doi.org/10.3390/ijms22094910