
AP Collagen Peptides Prevent Cortisol-Induced Decrease of Collagen Type I in Human Dermal Fibroblasts

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
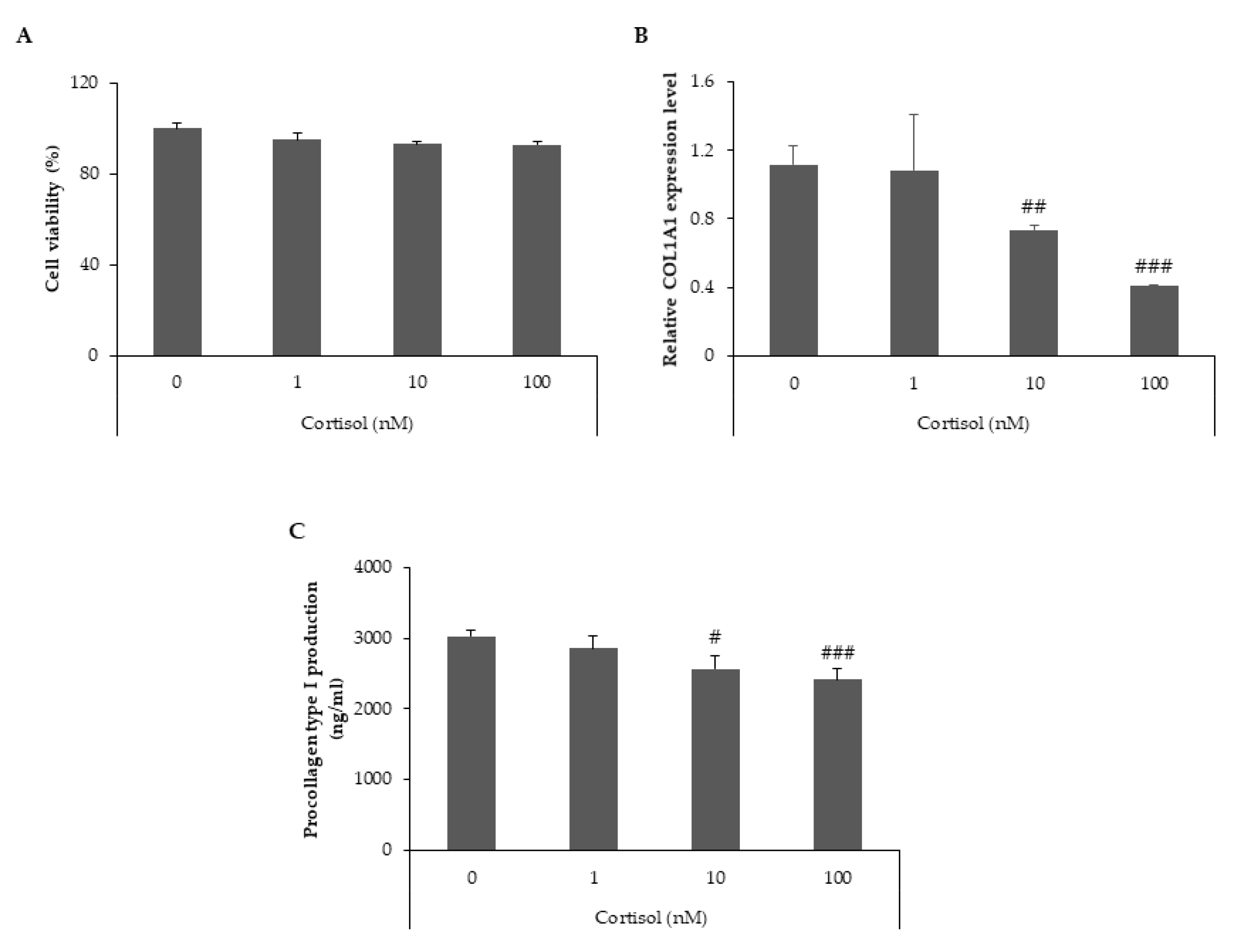
2.1. Cortisol Inhibits Collagen Type I Synthesis in Human Dermal Fibroblasts
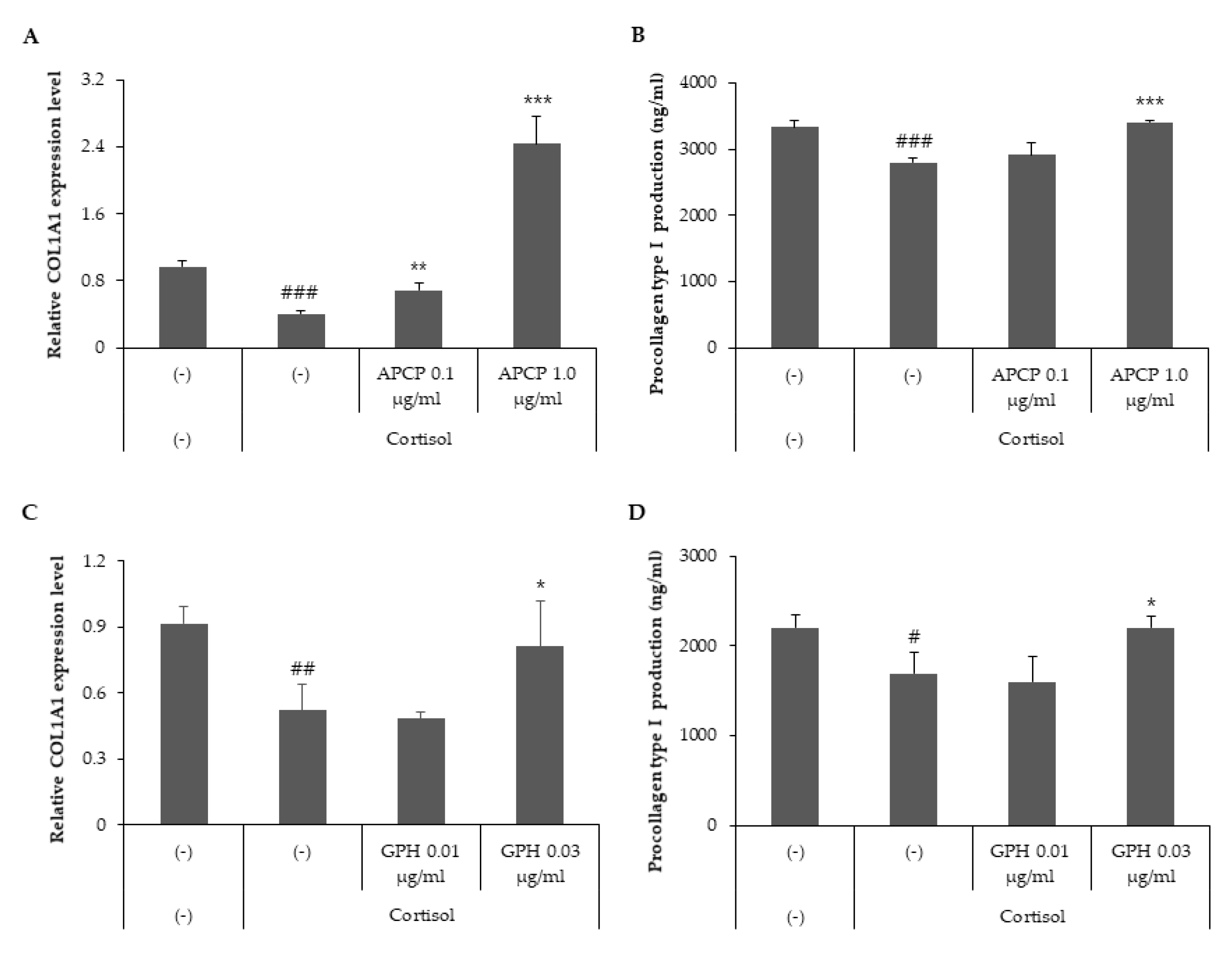
2.2. AP Collagen Peptides and GPH Prevent the Inhibition of Collagen Type I Mediated by Cortisol
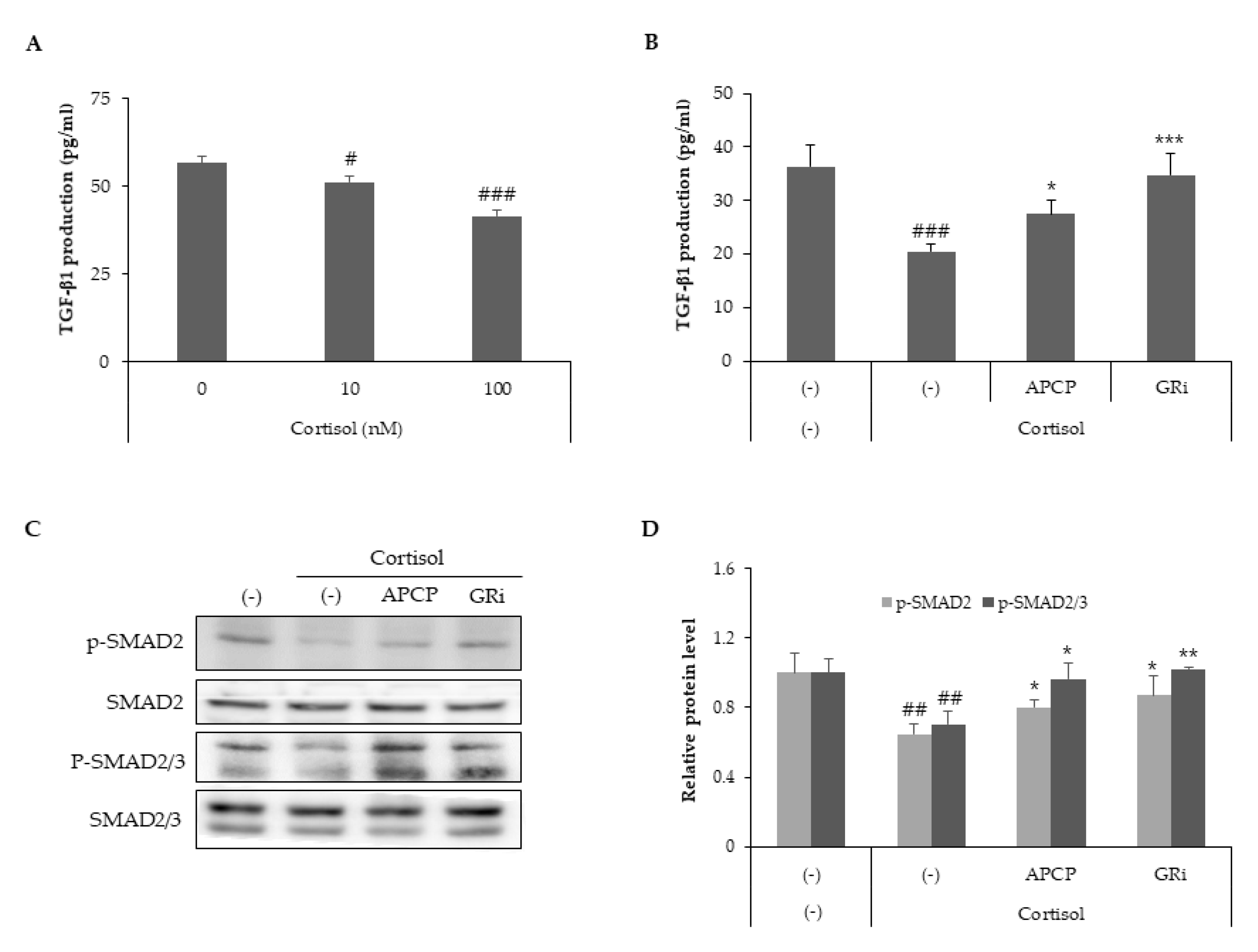
2.3. GR Signaling Is Responsible for Cortisol-Dependent Inhibition of Collagen Type I Synthesis and AP Collagen Peptides Inhibit GR Activation
2.4. AP Collagen Peptides Recover Cortisol-Dependent Inhibition of TGF-β Signaling
2.5. AP Collagen Peptides Alleviate Cortisol-Dependent Inhibition of Collagen Type I in Replicative Senescent HDFs Presenting Increased Cortisol Sensitivity and GR Expression
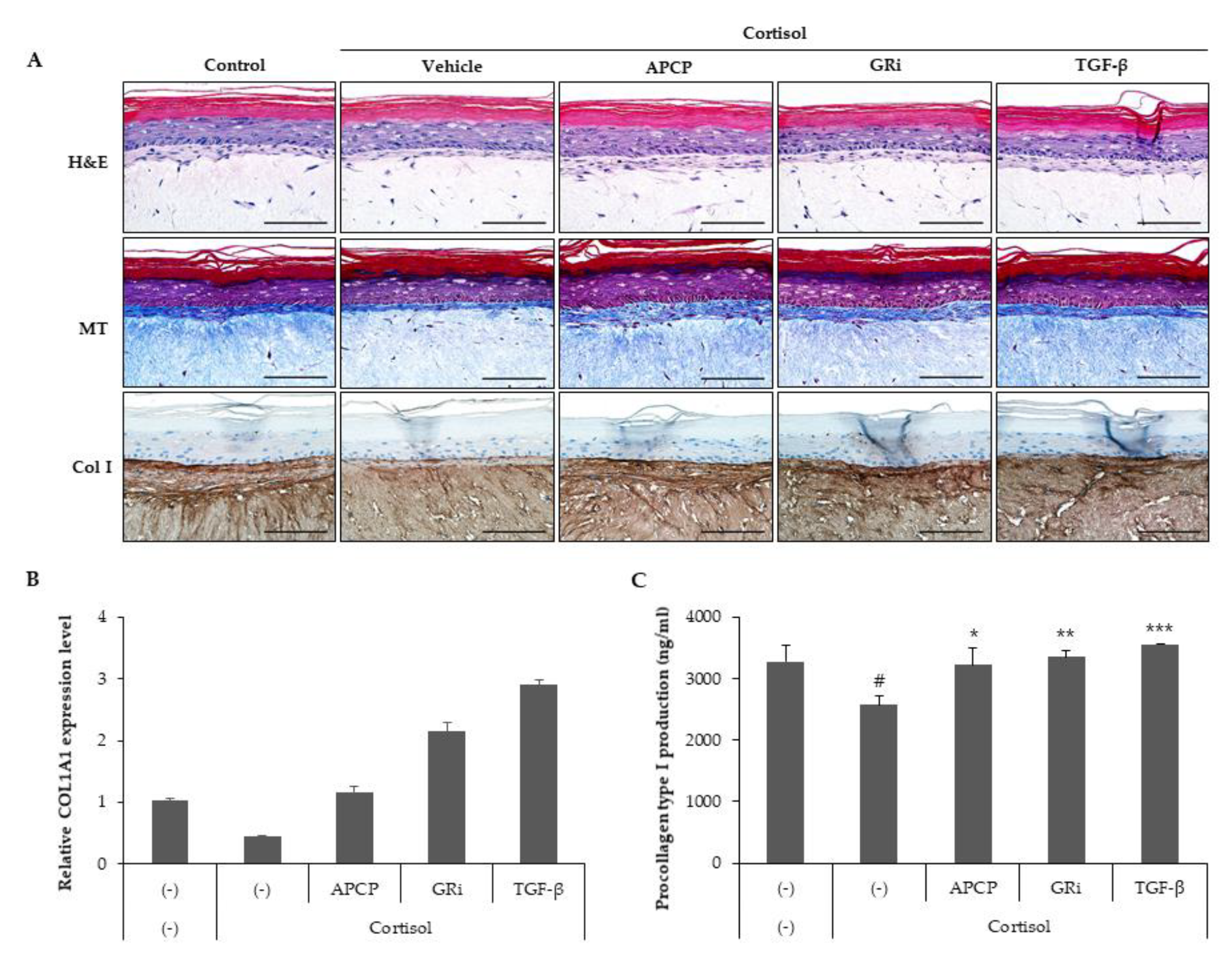
2.6. AP Collagen Peptides Prevent Cortisol-Dependent Inhibition of Collagen Synthesis in a Reconstituted Human Skin Model
3. Discussion
4. Materials and Methods
4.1. Collagen Peptides and Chemicals
4.2. Cell Culture
4.3. Quantitative Real-Time RT-PCR (qRT-PCR)
4.4. Western Blot Analysis
4.5. ELISA
4.6. Senescence Associated Beta-Galactosidase (SA-β-Gal) Assay
4.7. Immunohistochemistry Analysis of a Skin Model
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miller, W.L.; Auchus, R.J. The Molecular Biology, Biochemistry, and Physiology of Human Steroidogenesis and Its Disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [Green Version]
- Farman, N.; Maubec, E.; Poeggeler, B.; Klatte, J.E.; Jaisser, F.; Paus, R. The mineralocorticoid receptor as a novel player in skin biology: Beyond the renal horizon? Exp. Dermatol. 2010, 19, 100–107. [Google Scholar] [CrossRef]
- Pérez, P. Glucocorticoid receptors, epidermal homeostasis and hair follicle differentiation. Dermato-Endocrinology 2011, 3, 166–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichrath, J. Ancient friends, revisited: New aspects on the important role of nuclear receptor signalling for skin physiology and for the treatment of skin diseases. Dermato-Endocrinology 2011, 3, 121–124. [Google Scholar] [CrossRef]
- Schmuth, M.; Watson, R.E.; Deplewski, D.; Dubrac, S.; Zouboulis, C.C.; Griffiths, C.E. Nuclear Hormone Receptors in Human Skin. Horm. Metab. Res. 2007, 39, 96–105. [Google Scholar] [CrossRef]
- Jaisser, F.; Farman, N. Emerging Roles of the Mineralocorticoid Receptor in Pathology: Toward New Paradigms in Clinical Pharmacology. Pharmacol. Rev. 2016, 68, 49–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinerie, L.; Munier, M.; le Menuet, D.; Meduri, G.; Viengchareun, S.; Lombès, M. The mineralocorticoid signaling pathway throughout development: Expression, regulation and pathophysiological implications. Biochim. et Biophys. Acta (BBA) 2013, 95, 148–157. [Google Scholar] [CrossRef]
- Mitts, T.F.; Bunda, S.; Wang, Y.; Hinek, A. Aldosterone and Mineralocorticoid Receptor Antagonists Modulate Elastin and Collagen Deposition in Human Skin. J. Investig. Dermatol. 2010, 130, 2396–2406. [Google Scholar] [CrossRef]
- Pratt, W.B. The role of heat shock proteins in regulating the function, folding, and trafficking of the glucocorticoid receptor. J. Biol. Chem. 1993, 268, 21455–21458. [Google Scholar] [CrossRef]
- Ray, A.; Prefontaine, K.E. Physical association and functional antagonism between the p65 subunit of transcription factor NF-kappa B and the glucocorticoid receptor. Proc. Natl. Acad. Sci. USA 1994, 91, 752–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialog. Clin. Neurosci. 2006, 8, 383–395. [Google Scholar]
- Boumpas, D.T.; Chrousos, G.P.; Wilder, R.L.; Cupps, T.R.; Balow, J.E. Glucocorticoid Therapy for Immune-Mediated Diseases: Basic and Clinical Correlates. Ann. Intern. Med. 1993, 119, 1198–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boscaro, M.; Barzon, L.; Fallo, F.; Sonino, N. Cushing’s syndrome. Lancet 2001, 357, 783–791. [Google Scholar] [CrossRef]
- Boix, J.; Bigas, J.; Sevilla, L.M.; Iacobone, M.; Citton, M.; Torresan, F.; Caroccia, B.; Rossi, G.P.; Pérez, P. Primary aldosteronism patients show skin alterations and abnormal activation of glucocorticoid receptor in keratinocytes. Sci. Rep. 2017, 7, 15806. [Google Scholar] [CrossRef] [Green Version]
- Terao, M.; Katayama, I. Local cortisol/corticosterone activation in skin physiology and pathology. J. Dermatol. Sci. 2016, 84, 11–16. [Google Scholar] [CrossRef]
- Wang, Y.; Azaïs, T.; Robin, M.; Vallée, A.; Catania, C.; Legriel, P.; Pehau-Arnaudet, G.; Babonneau, F.; Giraud-Guille, M.-M.; Nassif, N. The predominant role of collagen in the nucleation, growth, structure and orientation of bone apatite. Nat. Mater. 2012, 11, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Sugihara, F.; Wang, X. Ingestion of bioactive collagen hydrolysates enhance facial skin moisture and elasticity and reduce facial ageing signs in a randomised double-blind placebo-controlled clinical study. J. Sci. Food Agric. 2016, 96, 4077–4081. [Google Scholar] [CrossRef] [PubMed]
- Proksch, E.; Schunck, M.; Zague, V.; Segger, D.; Degwert, J.; Oesser, S. Oral Intake of Specific Bioactive Collagen Peptides Reduces Skin Wrinkles and Increases Dermal Matrix Synthesis. Ski. Pharmacol. Physiol. 2014, 27, 113–119. [Google Scholar] [CrossRef]
- Oba, C.; Ohara, H.; Morifuji, M.; Ito, K.; Ichikawa, S.; Kawahata, K.; Koga, J. Collagen hydrolysate intake improves the loss of epidermal barrier function and skin elasticity induced by UVB irradiation in hairless mice. Photodermatol. Photoimmunol. Photomed. 2013, 29, 204–211. [Google Scholar] [CrossRef]
- Sevilla, L.M.; Pérez, P. Roles of the Glucocorticoid and Mineralocorticoid Receptors in Skin Pathophysiology. Int. J. Mol. Sci. 2018, 19, 1906. [Google Scholar] [CrossRef] [Green Version]
- Reichardt, H.; Tuckermann, J.; Bauer, A.; Schütz, G. Molecular genetic dissection of glucocorticoid receptor function in vivo. Z. Rheumatol. 2000, 59, S1–S5. [Google Scholar] [CrossRef]
- Yamamoto, K.R. Steroid Receptor Regulated Transcription of Specific Genes and Gene Networks. Annu. Rev. Genet. 1985, 19, 209–252. [Google Scholar] [CrossRef]
- Chen, W.; Dang, T.; Blind, R.D.; Wang, Z.; Cavasotto, C.N.; Hittelman, A.B.; Rogatsky, I.; Logan, S.K.; Garabedian, M.J. Glucocorticoid Receptor Phosphorylation Differentially Affects Target Gene Expression. Mol. Endocrinol. 2008, 22, 1754–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Frederick, J.; Garabedian, M.J. Deciphering the Phosphorylation “Code” of the Glucocorticoid Receptor In Vivo. J. Biol. Chem. 2002, 277, 26573–26580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varga, J.; Rosenbloom, J.; A Jimenez, S. Transforming growth factor β (TGFβ) causes a persistent increase in steady-state amounts of type I and type III collagen and fibronectin mRNAs in normal human dermal fibroblasts. Biochem. J. 1987, 247, 597–604. [Google Scholar] [CrossRef]
- Quan, T.; Shao, Y.; He, T.; Voorhees, J.J.; Fisher, G.J. Reduced Expression of Connective Tissue Growth Factor (CTGF/CCN2) Mediates Collagen Loss in Chronologically Aged Human Skin. J. Investig. Dermatol. 2010, 130, 415–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derynck, R.; Zhang, Y.E. Smad-dependent and Smad-independent pathways in TGF-β family signalling. Nat. Cell Biol. 2003, 425, 577–584. [Google Scholar] [CrossRef]
- Massagué, J.; Seoane, J.; Wotton, D. Smad transcription factors. Genes Dev. 2005, 19, 2783–2810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderbilt, J.N.; Miesfeld, R.; Maler, B.A.; Yamamoto, K.R. Intracellular Receptor Concentration Limits Glucocorticoid-Dependent Enhancer Activity. Mol. Endocrinol. 1987, 1, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Bamberger, C.M.; Schulte, H.M.; Chrousos, G.P. Molecular Determinants of Glucocorticoid Receptor Function and Tissue Sensitivity to Glucocorticoids. Endocr. Rev. 1996, 17, 245–261. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Bioactive Peptides. J. AOAC Int. 2008, 91, 914–931. [Google Scholar] [CrossRef] [Green Version]
- Silva-Sánchez, C.; de la Rosa, A.P.B.; León-Galván, M.F.; de Lumen, B.O.; de León-Rodríguez, A.; de Mejía, E.G. Bioactive Peptides in Amaranth (Amaranthus hypochondriacus) Seed. J. Agric. Food Chem. 2008, 56, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, R.W. Role of collagen hydrolysate in bone and joint disease. Semin. Arthritis Rheum. 2000, 30, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Guillerminet, F.; Beaupied, H.; Fabien-Soulé, V.; Tomé, D.; Benhamou, C.-L.; Roux, C.; Blais, A. Hydrolyzed collagen improves bone metabolism and biomechanical parameters in ovariectomized mice: An in vitro and in vivo study. Bone 2010, 46, 827–834. [Google Scholar] [CrossRef]
- Zhang, Y.; Kouguchi, T.; Shimizu, M.; Ohmori, T.; Takahata, Y.; Morimatsu, F. Chicken Collagen Hydrolysate Protects Rats from Hypertension and Cardiovascular Damage. J. Med. Food 2010, 13, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Hayasaka, F.; Deguchi, K.; Okudera, T.; Furusawa, T.; Sakai, Y. Absorption and plasma kinetics of collagen tripeptide after peroral or intraperitoneal administration in rats. Biosci. Biotechnol. Biochem. 2015, 79, 2026–2033. [Google Scholar] [CrossRef] [Green Version]
- Iwai, K.; Hasegawa, T.; Taguchi, Y.; Morimatsu, F.; Sato, K.; Nakamura, Y.; Higashi, A.; Kido, Y.; Nakabo, Y.; Ohtsuki, K. Identification of Food-Derived Collagen Peptides in Human Blood after Oral Ingestion of Gelatin Hydrolysates. J. Agric. Food Chem. 2005, 53, 6531–6536. [Google Scholar] [CrossRef]
- Yazaki, M.; Ito, Y.; Yamada, M.; Goulas, S.; Teramoto, S.; Nakaya, M.-A.; Ohno, S.; Yamaguchi, K. Oral Ingestion of Collagen Hydrolysate Leads to the Transportation of Highly Concentrated Gly-Pro-Hyp and Its Hydrolyzed Form of Pro-Hyp into the Bloodstream and Skin. J. Agric. Food Chem. 2017, 65, 2315–2322. [Google Scholar] [CrossRef] [Green Version]
- Ohara, H.; Matsumoto, H.; Ito, K.; Iwai, K.; Sato, K. Comparison of Quantity and Structures of Hydroxyproline-Containing Peptides in Human Blood after Oral Ingestion of Gelatin Hydrolysates from Different Sources. J. Agric. Food Chem. 2007, 55, 1532–1535. [Google Scholar] [CrossRef]
- Juher, T.F.; Pérez, E.B. An overview of the beneficial effects of hydrolysed collagen intake on joint and bone health and on skin ageing. Nutr. Hosp. 2015, 32, 62–66. [Google Scholar]
- Fleischmajer, R.; Perlish, J.S.; Timpl, R. Collagen Fibrillogenesis in Human Skin. Ann. N. Y. Acad. Sci. 1985, 460, 246–257. [Google Scholar] [CrossRef]
- Fleischmajer, R.; MacDonald, E.D.; Perlish, J.S.; Burgeson, R.E.; Fisher, L.W. Dermal collagen fibrils are hybrids of type I and type III collagen molecules. J. Struct. Biol. 1990, 105, 162–169. [Google Scholar] [CrossRef]
- Varani, J.; Dame, M.K.; Rittie, L.; Fligiel, S.E.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Decreased Collagen Production in Chronologically Aged Skin: Roles of Age-Dependent Alteration in Fibroblast Function and Defective Mechanical Stimulation. Am. J. Pathol. 2006, 168, 1861–1868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovell, C.; Smolenski, K.; Duance, V.; Light, N.; Young, S.; Dyson, M. Type I and III collagen content and fibre distribution in normal human skin during ageing. Br. J. Dermatol. 1987, 117, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J. Introduction to skin aging. J. Tissue Viability 2017, 26, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oikarinen, A. The aging of skin: Chronoaging versus photoaging. Photodermatol. Photoimmunol. Photomed. 1990, 7, 3–4. [Google Scholar]
- Zouboulis, C.C.; Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Elewa, R.; Makrantonaki, E. Aesthetic aspects of skin aging, prevention, and local treatment. Clin. Dermatol. 2019, 37, 365–372. [Google Scholar] [CrossRef]
- Venkatesan, J.; Anil, S.; Kim, S.-K.; Shim, M.S. Marine Fish Proteins and Peptides for Cosmeceuticals: A Review. Mar. Drugs 2017, 15, 143. [Google Scholar] [CrossRef]
- Czajka, A.; Kania, E.M.; Genovese, L.; Corbo, A.; Merone, G.; Luci, C.; Sibilla, S. Daily oral supplementation with collagen peptides combined with vitamins and other bioactive compounds improves skin elasticity and has a beneficial effect on joint and general wellbeing. Nutr. Res. 2018, 57, 97–108. [Google Scholar] [CrossRef]
- Haydont, V.; Bernard, B.A.; Fortunel, N.O. Age-related evolutions of the dermis: Clinical signs, fibroblast and extracellular matrix dynamics. Mech. Ageing Dev. 2019, 177, 150–156. [Google Scholar] [CrossRef]
- Song, H.; Zhang, S.; Zhang, L.; Li, B. Effect of Orally Administered Collagen Peptides from Bovine Bone on Skin Aging in Chronologically Aged Mice. Nutrients 2017, 9, 1209. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Zhuang, Y.; Li, B. Effects of Collagen and Collagen Hydrolysate from Jellyfish Umbrella on Histological and Immunity Changes of Mice Photoaging. Nutrients 2013, 5, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, H.; Li, B.; Zhang, Z.; Xue, C.; Yu, G.; Wang, J.; Bao, Y.; Bu, L.; Sun, J.; Peng, Z.; et al. Moisture absorption and retention properties, and activity in alleviating skin photodamage of collagen polypeptide from marine fish skin. Food Chem. 2012, 135, 1432–1439. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Meng, M.; Cheng, X.; Li, B.; Wang, C. The effect of collagen hydrolysates from silver carp (Hypophthalmichthys molitrix) skin on UV-induced photoaging in mice: Molecular weight affects skin repair. Food Funct. 2017, 8, 1538–1546. [Google Scholar] [CrossRef]
- León-López, A.; Morales-Peñaloza, A.; Martínez-Juárez, V.M.; Vargas-Torres, A.; Zeugolis, D.I.; Aguirre-Álvarez, G. Hydrolyzed Collagen—Sources and Applications. Molecules 2019, 24, 4031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusubata, M.; Koyama, Y.-I.; Tometsuka, C.; Shigemura, Y.; Sato, K. Detection of endogenous and food-derived collagen dipeptide prolylhydroxyproline (Pro-Hyp) in allergic contact dermatitis-affected mouse ear. Biosci. Biotechnol. Biochem. 2015, 79, 1356–1361. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Asai, T.T.; Jimi, S. Collagen-Derived Di-Peptide, Prolylhydroxyproline (Pro-Hyp): A New Low Molecular Weight Growth-Initiating Factor for Specific Fibroblasts Associated with Wound Healing. Front. Cell Dev. Biol. 2020, 8, 548975. [Google Scholar] [CrossRef]
- Chiang, T.M.; Postlethwaite, A.E.; Beachey, E.H.; Seyer, J.M.; Kang, A.H. Binding of Chemotactic Collagen-Derived Peptides to Fibroblasts. J. Clin. Investig. 1978, 62, 916–922. [Google Scholar] [CrossRef] [Green Version]
- E Postlethwaite, A.; Seyer, J.M.; Kang, A.H. Chemotactic attraction of human fibroblasts to type I, II, and III collagens and collagen-derived peptides. Proc. Natl. Acad. Sci. USA 1978, 75, 871–875. [Google Scholar] [CrossRef] [Green Version]
- Ohara, H.; Ichikawa, S.; Matsumoto, H.; Akiyama, M.; Fujimoto, N.; Kobayashi, T.; Tajima, S. Collagen-derived dipeptide, proline-hydroxyproline, stimulates cell proliferation and hyaluronic acid synthesis in cultured human dermal fibroblasts. J. Dermatol. 2010, 37, 330–338. [Google Scholar] [CrossRef]
- Sapolsky, R.M.; Krey, L.C.; McEwen, B.S. The Neuroendocrinology of Stress and Aging: The Glucocorticoid Cascade Hypothesis. Endocr. Rev. 1986, 7, 284–301. [Google Scholar] [CrossRef]
- Chen, Y.; Lyga, J. Brain-skin connection: Stress, inflammation and skin aging. Curr. Drug Targets Inflamm. Allergy 2014, 13, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Terao, M.; Tani, M.; Itoi, S.; Yoshimura, T.; Hamasaki, T.; Murota, H.; Katayama, I. 11β-Hydroxysteroid Dehydrogenase 1 Specific Inhibitor Increased Dermal Collagen Content and Promotes Fibroblast Proliferation. PLoS ONE 2014, 9, e93051. [Google Scholar] [CrossRef] [Green Version]
- Tiganescu, A.; Tahrani, A.A.; Morgan, S.A.; Otranto, M.; Desmoulière, A.; Abrahams, L.; Hassan-Smith, Z.; Walker, E.A.; Rabbitt, E.H.; Cooper, M.S.; et al. 11β-Hydroxysteroid dehydrogenase blockade prevents age-induced skin structure and function defects. J. Clin. Investig. 2013, 123, 3051–3060. [Google Scholar] [CrossRef]
- Charmandari, E.; Kino, T.; Chrousos, G.P. Familial/Sporadic Glucocorticoid Resistance: Clinical Phenotype and Molecular Mechanisms. Ann. N. Y. Acad. Sci. 2004, 1024, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Longui, C.A.; Faria, C.D.C. Evaluation of Glucocorticoid Sensitivity and Its Potential Clinical Applicability. Horm. Res. Paediatr. 2009, 71, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Leiferman, K.M.; Schroeter, A.; Kirschner, M.K.; Spelsberg, T.C. Characterization of the Glucocorticoid Receptor in Human Skin. J. Investig. Dermatol. 1983, 81, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Bigas, J.; Sevilla, L.M.; Carceller, E.; Boix, J.; Pérez, P. Epidermal glucocorticoid and mineralocorticoid receptors act cooperatively to regulate epidermal development and counteract skin inflammation. Cell Death Dis. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Cai, J.; Zheng, T.; Lotz, M.; Zhang, Y.; Masood, R.; Gill, P. Glucocorticoids Induce Kaposi’s Sarcoma Cell Proliferation Through the Regulation of Transforming Growth Factor-β. Blood 1997, 89, 1491–1500. [Google Scholar] [CrossRef] [Green Version]
- Bolkenius, U.; Hahn, D.; Gressner, A.M.; Breitkopf, K.; Dooley, S.; Wickert, L. Glucocorticoids decrease the bioavailability of TGF-β which leads to a reduced TGF-β signaling in hepatic stellate cells. Biochem. Biophys. Res. Commun. 2004, 325, 1264–1270. [Google Scholar] [CrossRef] [PubMed]
- Wen, F.-Q.; Kohyama, T.; Sköld, C.M.; Zhu, Y.K.; Liu, X.; Romberger, D.J.; Stoner, J.; Rennard, S.I. Glucocorticoids Modulate TGF-β Production. Inflammation 2002, 26, 279–290. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chae, M.; Bae, I.-H.; Lim, S.; Jung, K.; Roh, J.; Kim, W. AP Collagen Peptides Prevent Cortisol-Induced Decrease of Collagen Type I in Human Dermal Fibroblasts. Int. J. Mol. Sci. 2021, 22, 4788. https://doi.org/10.3390/ijms22094788
Chae M, Bae I-H, Lim S, Jung K, Roh J, Kim W. AP Collagen Peptides Prevent Cortisol-Induced Decrease of Collagen Type I in Human Dermal Fibroblasts. International Journal of Molecular Sciences. 2021; 22(9):4788. https://doi.org/10.3390/ijms22094788
Chicago/Turabian StyleChae, Minjung, Il-Hong Bae, Sunghwan Lim, Kyoungmi Jung, Jonghwa Roh, and Wangi Kim. 2021. "AP Collagen Peptides Prevent Cortisol-Induced Decrease of Collagen Type I in Human Dermal Fibroblasts" International Journal of Molecular Sciences 22, no. 9: 4788. https://doi.org/10.3390/ijms22094788