
Running and Swimming Differently Adapt the BDNF/TrkB Pathway to a Slow Molecular Pattern at the NMJ

 , , and
, , and
Abstract
:1. Introduction
2. Results
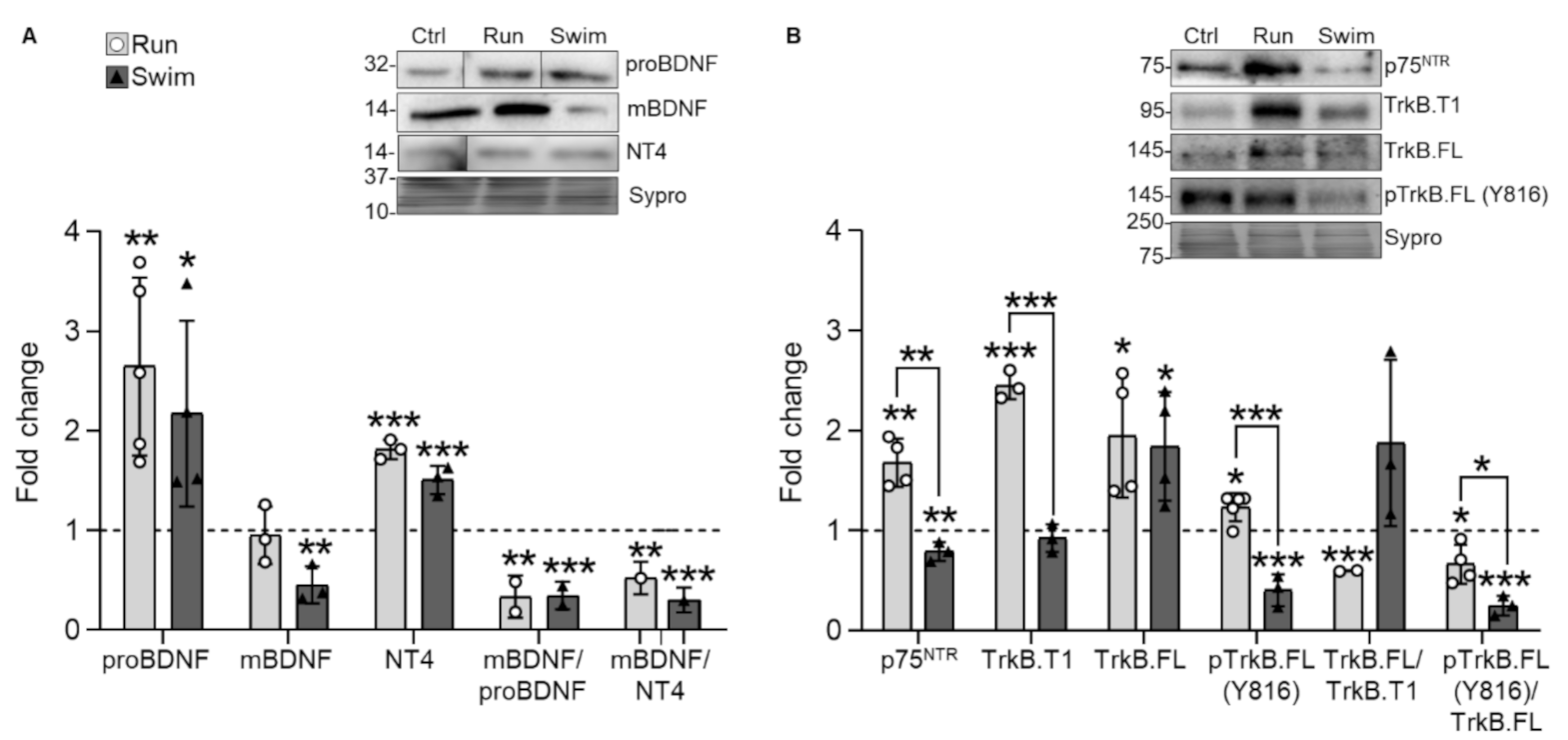
2.1. Neurotrophic Factors and Receptors
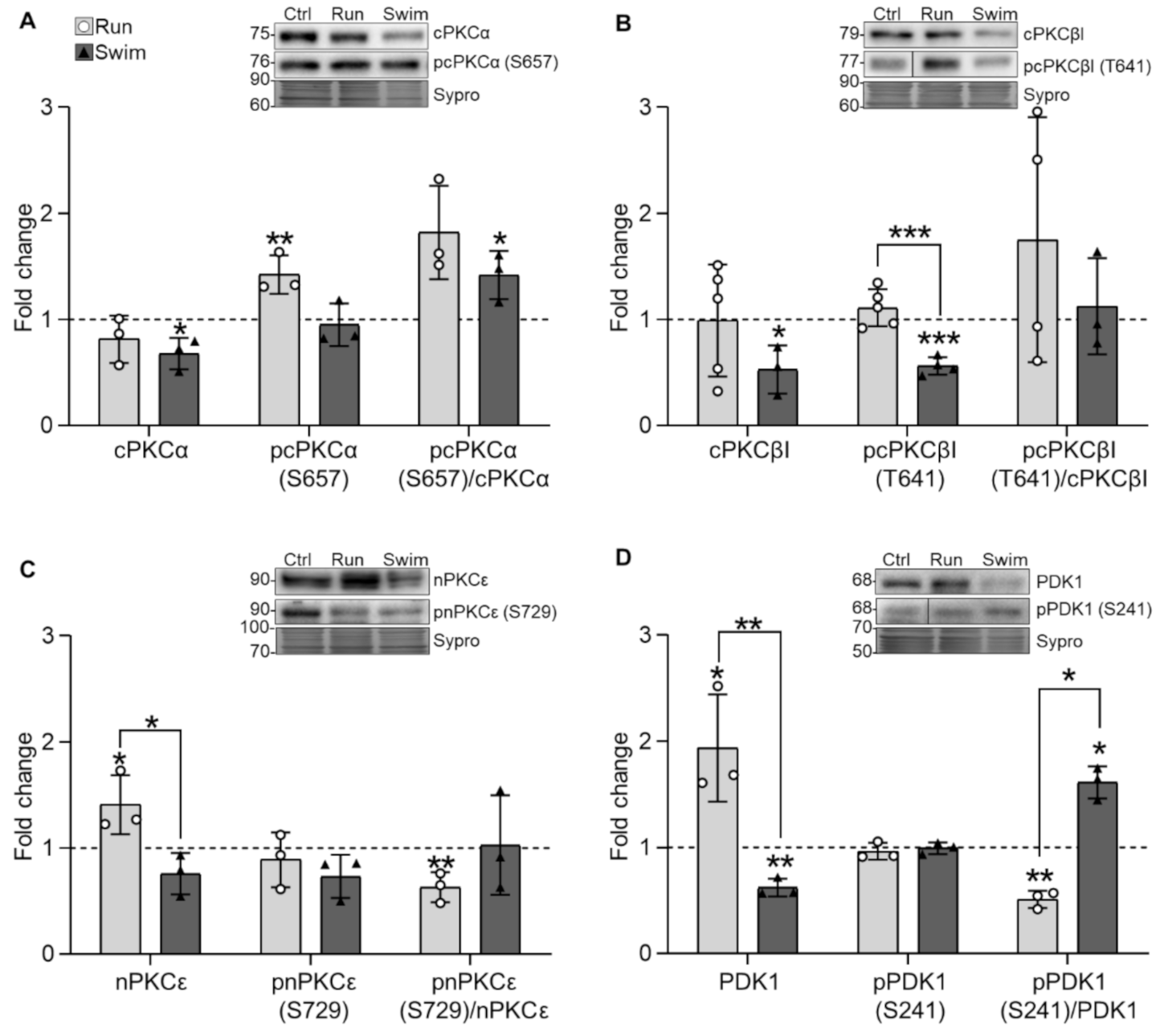
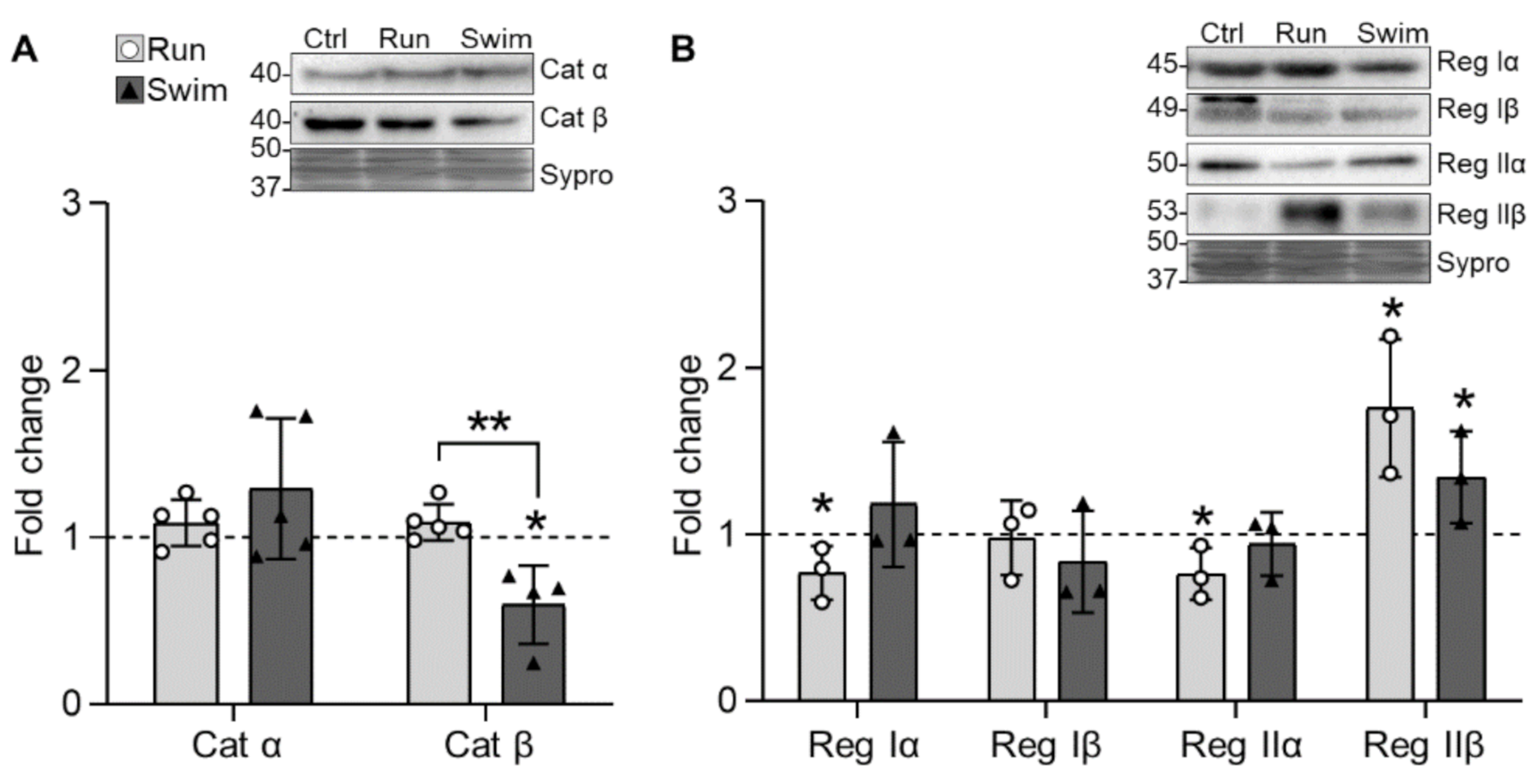
2.2. Serine-Threonine Kinases
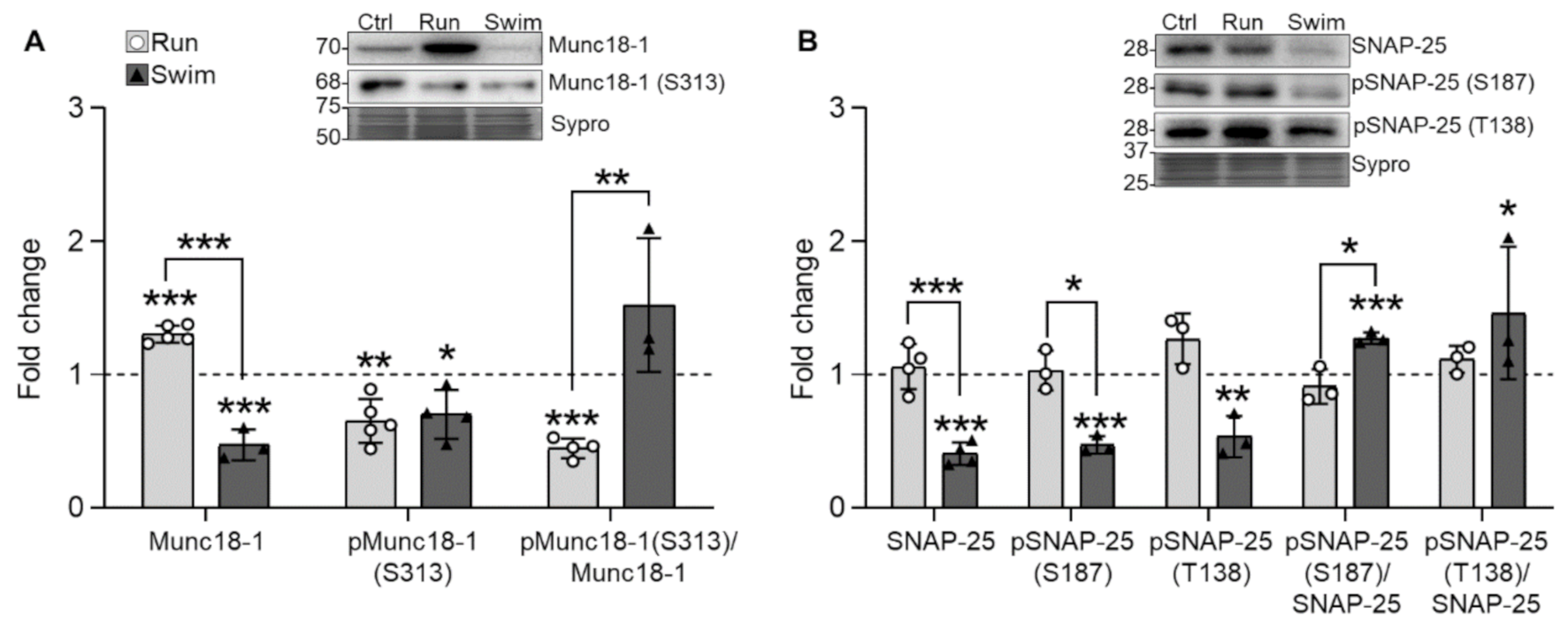
2.3. SNARE/SM Proteins
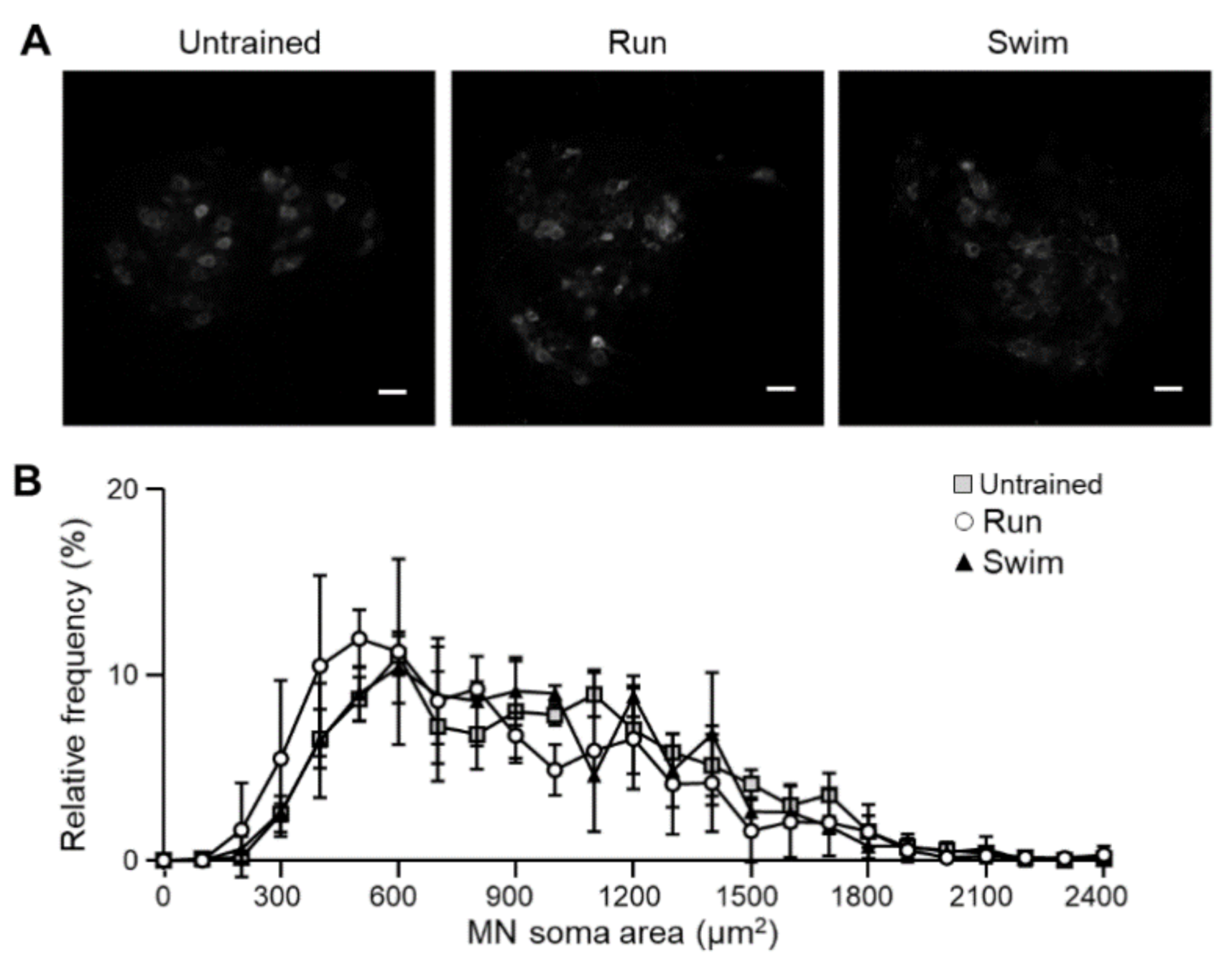
2.4. Motor Neurons
3. Discussion
3.1. Neurotrophic Factors and Receptors
3.2. Serine Threonine Kinases and Their Targets
3.2.1. PDK1 and PKC
3.2.2. PKA
3.3. BDNF/TrkB Impact over the Morphology of the Neuromuscular System
4. Materials and Methods
4.1. Animals
4.2. Training Protocol
4.3. Western Blotting
4.4. Immunohistochemistry
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Baldwin, K.M.; Haddad, F.; Pandorf, C.E.; Roy, R.R.; Edgerton, V.R. Alterations in muscle mass and contractile phenotype in response to unloading models: Role of transcriptional/pretranslational mechanisms. Front. Physiol. 2013, 4, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cisterna, B.A.; Cardozo, C.; Saez, J.C. Neuronal involvement in muscular atrophy. Front. Cell. Neurosci. 2014, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurtado, E.; Cilleros, V.; Nadal, L.; Simó, A.; Obis, T.; Garcia, N.; Santafé, M.M.; Tomàs, M.; Halievski, K.; Jordan, C.L.; et al. Muscle Contraction Regulates BDNF/TrkB Signaling to Modulate Synaptic Function through Presynaptic cPKCα and cPKCβI. Front. Mol. Neurosci. 2017, 10, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Fischbach, G.D.; Robbins, N. Effect of chronic disuse of rat soleus neuromuscular junctions on postsynaptic membrane. J. Neurophysiol. 1971, 34, 562–569. [Google Scholar] [CrossRef]
- Robbins, N.; Fischbach, G.D. Effect of chronic disuse of rat soleus neuromuscular junctions on presynaptic function. J. Neurophysiol. 1971, 34, 570–578. [Google Scholar] [CrossRef]
- Deschenes, M.R. Adaptations of the neuromuscular junction to exercise training. Curr. Opin. Physiol. 2019, 10, 10–16. [Google Scholar] [CrossRef]
- Delezie, J.; Handschin, C. Endocrine Crosstalk Between Skeletal Muscle and the Brain. Front. Neurol. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Wu, C.-W.; Chang, Y.-T.; Yu, L.; Chen, H.; Jen, C.J.; Wu, S.-Y.; Lo, C.-P.; Kuo, Y.-M. Exercise enhances the proliferation of neural stem cells and neurite growth and survival of neuronal progenitor cells in dentate gyrus of middle-aged mice. J. Appl. Physiol. 2008, 105, 1585–1594. [Google Scholar] [CrossRef]
- Gómez-Pinilla, F.; Ying, Z.; Opazo, P.; Roy, R.R.; Edgerton, V.R. Differential regulation by exercise of BDNF and NT-3 in rat spinal cord and skeletal muscle. Eur. J. Neurosci. 2001, 13, 1078–1084. [Google Scholar] [CrossRef]
- Gómez-Pinilla, F.; Ying, Z.; Roy, R.R.; Molteni, R.; Edgerton, V.R. Voluntary Exercise Induces a BDNF-Mediated Mechanism That Promotes Neuroplasticity. J. Neurophysiol. 2002, 88, 2187–2195. [Google Scholar] [CrossRef] [Green Version]
- Asensio-Pinilla, E.; Udina, E.; Jaramillo, J.; Navarro, X. Electrical stimulation combined with exercise increase axonal regeneration after peripheral nerve injury. Exp. Neurol. 2009, 219, 258–265. [Google Scholar] [CrossRef]
- Husain, K.; Somani, S.M. Response of cardiac antioxidant system to alcohol and exercise training in the rat. Alcohol 1997, 14, 301–307. [Google Scholar] [CrossRef]
- Miyazaki, H.; Oh-ishi, S.; Ookawara, T.; Kizaki, T.; Toshinai, K.; Ha, S.; Haga, S.; Ji, L.L.; Ohno, H. Strenuous endurance training in humans reduces oxidative stress following exhausting exercise. Eur. J. Appl. Physiol. 2001, 84, 1–6. [Google Scholar] [CrossRef]
- Molé, P.A.; Oscai, L.B.; Holloszy, J.O. Adaptation of muscle to exercise. Increase in levels of palmityl Coa synthetase, carnitine palmityltransferase, and palmityl Coa dehydrogenase, and in the capacity to oxidize fatty acids. J. Clin. Investig. 1971, 50, 2323–2330. [Google Scholar] [CrossRef] [Green Version]
- Dorlöchter, M.; Irintchev, A.; Brinkers, M.; Wernig, A. Effects of enhanced activity on synaptic transmission in mouse extensor digitorum longus muscle. J. Physiol. 1991, 436, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Tomas, J.; Batlle, J.; Fenoll, M.R.; Santafé, M.; Lanuza, M.A. Activity-dependent plastic changes in the motor nerve terminals of the adult rat. Biol. Cell 1993, 79, 133–137. [Google Scholar] [CrossRef]
- Tomas, J.; Santafé, M.; Lanuza, M.A.; Fenoll-Brunet, M.R. Physiological activity-dependent ultrastructural plasticity in normal adult rat neuromuscular junctions. Biol. Cell 1997, 89, 19–28. [Google Scholar] [CrossRef]
- Andonian, M.H.; Fahim, M.A. Effects of endurance exercise on the morphology of mouse neuromuscular junctions during ageing. J. Neurocytol. 1987, 16, 589–599. [Google Scholar] [CrossRef]
- Deschenes, M.R.; Maresh, C.M.; Crivello, J.F.; Armstrong, L.E.; Kraemer, W.J.; Covault, J. The effects of exercise training of different intensities on neuromuscular junction morphology. J. Neurocytol. 1993, 22, 603–615. [Google Scholar] [CrossRef]
- Deschenes, M.R.; Sherman, E.G.; Roby, M.A.; Glass, E.K.; Harris, M.B. Effect of resistance training on neuromuscular junctions of young and aged muscles featuring different recruitment patterns. J. Neurosci. Res. 2015, 93, 504–513. [Google Scholar] [CrossRef] [Green Version]
- Nishimune, H.; Stanford, J.A.; Mori, Y. Role of exercise in maintaining the integrity of the neuromuscular junction. Muscle Nerve 2014, 49, 315–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdez, G.; Tapia, J.C.; Kang, H.; Clemenson, G.D.; Gage, F.H.; Lichtman, J.W.; Sanes, J.R. Attenuation of age-related changes in mouse neuromuscular synapses by caloric restriction and exercise. Proc. Natl. Acad. Sci. USA 2010, 107, 14863–14868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gür, H.; Gransberg, L.; van Dyke, D.; Knutsson, E.; Larsson, L. Relationship between in vivo muscle force at different speeds of isokinetic movements and myosin isoform expression in men and women. Eur. J. Appl. Physiol. 2003, 88, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Mackey, A.L.; Karlsen, A.; Couppé, C.; Mikkelsen, U.R.; Nielsen, R.H.; Magnusson, S.P.; Kjaer, M. Differential satellite cell density of type I and II fibres with lifelong endurance running in old men. Acta Physiol. 2014, 210, 612–627. [Google Scholar] [CrossRef]
- Cobley, J.N.; Moult, P.R.; Burniston, J.G.; Morton, J.P.; Close, G.L. Exercise improves mitochondrial and redox-regulated stress responses in the elderly: Better late than never! Biogerontology 2015, 16, 249–264. [Google Scholar] [CrossRef]
- Laye, M.J.; Nielsen, M.B.; Hansen, L.S.; Knudsen, T.; Pedersen, B.K. Physical Activity Enhances Metabolic Fitness Independently of Cardiorespiratory Fitness in Marathon Runners. Dis. Mark. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cocks, M.; Wagenmakers, A.J.M. The effect of different training modes on skeletal muscle microvascular density and endothelial enzymes controlling NO availability. J. Physiol. 2016, 594, 2245–2257. [Google Scholar] [CrossRef] [Green Version]
- Pette, D.; Staron, R.S. Myosin isoforms, muscle fiber types, and transitions. Microsc. Res. Tech. 2000, 50, 500–509. [Google Scholar] [CrossRef]
- Peuker, H.; Conjard, A.; Pette, D. α-cardiac-like myosin heavy chain as an intermediate between MHCIIa and MHCIβ in transforming rabbit muscle. Am. J. Physiol. Cell Physiol. 1998, 274, 595–602. [Google Scholar] [CrossRef]
- Sakamoto, K.; Aschenbach, W.G.; Hirshman, M.F.; Goodyear, L.J. Akt signaling in skeletal muscle: Regulation by exercise and passive stretch. Am. J. Physiol. Endocrinol. Metab. 2003, 285, 1081–1088. [Google Scholar] [CrossRef] [Green Version]
- Parkington, J.D.; Siebert, A.P.; LeBrasseur, N.K.; Fielding, R.A. Differential activation of mTOR signaling by contractile activity in skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, 1086–1090. [Google Scholar] [CrossRef] [Green Version]
- Just-Borràs, L.; Hurtado, E.; Cilleros-Mañé, V.; Biondi, O.; Charbonnier, F.; Tomàs, M.; Garcia, N.; Lanuza, M.A.; Tomàs, J. Overview of Impaired BDNF Signaling, Their Coupled Downstream Serine-Threonine Kinases and SNARE/SM Complex in the Neuromuscular Junction of the Amyotrophic Lateral Sclerosis Model SOD1-G93A Mice. Mol. Neurobiol. 2019, 56, 6856–6872. [Google Scholar] [CrossRef]
- Lu, B. BDNF and Activity-Dependent Synaptic Modulation. Learn. Mem. 2003, 10, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Mantilla, C.B.; Stowe, J.M.; Sieck, D.C.; Ermilov, L.G.; Greising, S.M.; Zhang, C.; Shokat, K.M.; Sieck, G.C. TrkB kinase activity maintains synaptic function and structural integrity at adult neuromuscular junctions. J. Appl. Physiol. 2014, 117, 910–920. [Google Scholar] [CrossRef]
- Garcia, N.; Santafe, M.M.; Tomàs, M.; Lanuza, M.A.; Besalduch, N.; Tomàs, J. Involvement of brain-derived neurotrophic factor (BDNF) in the functional elimination of synaptic contacts at polyinnervated neuromuscular synapses during development. J. Neurosci. Res. 2010, 88, 1406–1419. [Google Scholar] [CrossRef]
- Mantilla, C.B.; Zhan, W.-Z.; Sieck, G.C. Neurotrophins improve neuromuscular transmission in the adult rat diaphragm. Muscle Nerve 2004, 29, 381–386. [Google Scholar] [CrossRef]
- Santafé, M.M.; Garcia, N.; Tomàs, M.; Obis, T.; Lanuza, M.A.; Besalduch, N.; Tomàs, J. The interaction between tropomyosin-related kinase B receptors and serine kinases modulates acetylcholine release in adult neuromuscular junctions. Neurosci. Lett. 2014, 561, 171–175. [Google Scholar] [CrossRef]
- Matthews, V.B.; Åström, M.-B.; Chan, M.H.S.; Bruce, C.R.; Krabbe, K.S.; Prelovsek, O.; Åkerström, T.; Yfanti, C.; Broholm, C.; Mortensen, O.H.; et al. Brain-derived neurotrophic factor is produced by skeletal muscle cells in response to contraction and enhances fat oxidation via activation of AMP-activated protein kinase. Diabetologia 2009, 52, 1409–1418. [Google Scholar] [CrossRef] [Green Version]
- Je, H.S.; Yang, F.; Ji, Y.; Potluri, S.; Fu, X.-Q.; Luo, Z.-G.; Nagappan, G.; Chan, J.P.; Hempstead, B.; Son, Y.-J.; et al. ProBDNF and mature BDNF as punishment and reward signals for synapse elimination at mouse neuromuscular junctions. J. Neurosci. 2013, 33, 9957–9962. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Je, H.-S.; Ji, Y.; Nagappan, G.; Hempstead, B.; Lu, B. ProBDNF induced synaptic depression and retraction at developing neuromuscular synapses. J. Cell Biol. 2009, 185, 727–741. [Google Scholar] [CrossRef]
- Hempstead, B.L. Dissecting the diverse actions of pro- and mature neurotrophins. Curr. Alzheimer Res. 2006, 3, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middlemas, D.S.; Lindberg, R.A.; Hunter, T. trkB, a neural receptor protein-tyrosine kinase: Evidence for a full-length and two truncated receptors. Mol. Cell. Biol. 1991, 11, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Eide, F.F.; Vining, E.R.; Eide, B.L.; Zang, K.; Wang, X.Y.; Reichardt, L.F. Naturally occurring truncated trkB receptors have dominant inhibitory effects on brain-derived neurotrophic factor signaling. J. Neurosci. 1996, 16, 3123–3129. [Google Scholar] [CrossRef]
- Haapasalo, A.; Koponen, E.; Hoppe, E.; Wong, G.; Castrén, E. Truncated trkB.T1 Is Dominant Negative Inhibitor of trkB.TK+-Mediated Cell Survival. Biochem. Biophys. Res. Commun. 2001, 280, 1352–1358. [Google Scholar] [CrossRef]
- Gonzalez, M.; Ruggiero, F.P.; Chang, Q.; Shi, Y.J.; Rich, M.M.; Kraner, S.; Balice-Gordon, R.J. Disruption of Trkb-mediated signaling induces disassembly of postsynaptic receptor clusters at neuromuscular junctions. Neuron 1999, 24, 567–583. [Google Scholar] [CrossRef] [Green Version]
- Hurtado, E.; Cilleros, V.; Just, L.; Simó, A.; Nadal, L.; Tomàs, M.; Garcia, N.; Lanuza, M.A.; Tomàs, J. Synaptic Activity and Muscle Contraction Increases PDK1 and PKCβI Phosphorylation in the Presynaptic Membrane of the Neuromuscular Junction. Front. Mol. Neurosci. 2017, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Obis, T.; Hurtado, E.; Nadal, L.; Tomàs, M.; Priego, M.; Simon, A.; Garcia, N.; Santafe, M.M.; Lanuza, M.A.; Tomàs, J. The novel protein kinase C epsilon isoform modulates acetylcholine release in the rat neuromuscular junction. Mol. Brain 2015, 8, 80. [Google Scholar] [CrossRef] [Green Version]
- Besalduch, N.; Tomàs, M.; Santafé, M.M.; Garcia, N.; Tomàs, J.; Lanuza, M.A. Synaptic activity-related classical protein kinase C isoform localization in the adult rat neuromuscular synapse. J. Comp. Neurol. 2010, 518, 211–228. [Google Scholar] [CrossRef]
- Simó, A.; Cilleros-Mañé, V.; Just-Borràs, L.; Hurtado, E.; Nadal, L.; Tomàs, M.; Garcia, N.; Lanuza, M.A.; Tomàs, J. nPKCε Mediates SNAP-25 Phosphorylation of Ser-187 in Basal Conditions and After Synaptic Activity at the Neuromuscular Junction. Mol. Neurobiol. 2019, 56, 5346–5364. [Google Scholar] [CrossRef]
- Simó, A.; Just-Borràs, L.; Cilleros-Mañé, V.; Hurtado, E.; Nadal, L.; Tomàs, M.; Garcia, N.; Lanuza, M.A.; Tomàs, J. BDNF-TrkB Signaling Coupled to nPKCε and cPKCβI Modulate the Phosphorylation of the Exocytotic Protein Munc18-1 During Synaptic Activity at the Neuromuscular Junction. Front. Mol. Neurosci. 2018, 11, 207–227. [Google Scholar] [CrossRef] [Green Version]
- Cilleros-Mañé, V.; Just-Borràs, L.; Tomàs, M.; Garcia, N.; Tomàs, J.M.; Lanuza, M.A. The M 2 muscarinic receptor, in association to M 1, regulates the neuromuscular PKA molecular dynamics. FASEB J. 2020, 34, 4934–4955. [Google Scholar] [CrossRef] [Green Version]
- Garcia, N.; Tomàs, M.; Santafé, M.M.; Besalduch, N.; Lanuza, M.A.; Tomàs, J. The interaction between tropomyosin-related kinase B receptors and presynaptic muscarinic receptors modulates transmitter release in adult rodent motor nerve terminals. J. Neurosci. 2010, 30, 16514–16522. [Google Scholar] [CrossRef] [Green Version]
- Just-Borràs, L.; Hurtado, E.; Cilleros-Mañé, V.; Biondi, O.; Charbonnier, F.; Tomàs, M.; Garcia, N.; Tomàs, J.; Lanuza, M.A. Running and swimming prevent the deregulation of the BDNF/TrkB neurotrophic signalling at the neuromuscular junction in mice with amyotrophic lateral sclerosis. Cell. Mol. Life Sci. 2020, 77, 3027–3040. [Google Scholar] [CrossRef]
- Leenders, A.G.M.; Sheng, Z.-H. Modulation of neurotransmitter release by the second messenger-activated protein kinases: Implications for presynaptic plasticity. Pharmacol. Ther. 2005, 105, 69–84. [Google Scholar] [CrossRef] [Green Version]
- Kanning, K.C.; Kaplan, A.; Henderson, C.E. Motor Neuron Diversity in Development and Disease. Annu. Rev. Neurosci. 2010, 33, 409–440. [Google Scholar] [CrossRef]
- Shneider, N.A.; Brown, M.N.; Smith, C.A.; Pickel, J.; Alvarez, F.J. Gamma motor neurons express distinct genetic markers at birth and require muscle spindle-derived GDNF for postnatal survival. Neural Dev. 2009, 4, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Greising, S.M.; Ermilov, L.G.; Sieck, G.C.; Mantilla, C.B. Ageing and neurotrophic signalling effects on diaphragm neuromuscular function. J. Physiol. 2015, 593, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Moloney, E.B.; de Winter, F.; Verhaagen, J. ALS as a distal axonopathy: Molecular mechanisms affecting neuromuscular junction stability in the presymptomatic stages of the disease. Front. Neurosci. 2014, 8, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Ogborn, D.I.; Gardiner, P.F. Effects of exercise and muscle type on BDNF, NT-4/5, and TrKB expression in skeletal muscle. Muscle Nerve 2010, 41, 385–391. [Google Scholar] [CrossRef]
- Cuppini, R.; Sartini, S.; Agostini, D.; Guescini, M.; Ambrogini, P.; Betti, M.; Bertini, L.; Vallasciani, M.; Stocchi, V. Bdnf expression in rat skeletal muscle after acute or repeated exercise. Arch. Ital. Biol. 2007, 145, 99–110. [Google Scholar]
- Lee, H.W.; Ahmad, M.; Wang, H.W.; Leenen, F.H.H. Effects of exercise training on brain-derived neurotrophic factor in skeletal muscle and heart of rats post myocardial infarction. Exp. Physiol. 2017, 102, 314–328. [Google Scholar] [CrossRef] [PubMed]
- Zhan, G.; Huang, N.; Li, S.; Hua, D.; Zhang, J.; Fang, X.; Yang, N.; Luo, A.; Yang, C. PGC-1α-FNDC5-BDNF signaling pathway in skeletal muscle confers resilience to stress in mice subjected to chronic social defeat. Psychopharmacology 2018, 235, 3351–3358. [Google Scholar] [CrossRef]
- Wernig, A.; Salvini, T.F.; Irintchev, A. Axonal sprouting and changes in fibre types after running-induced muscle damage. J. Neurocytol. 1991, 20, 903–913. [Google Scholar] [CrossRef]
- Connor, E.A.; Smith, M.A. Retrograde signaling in the formation and maintenance of the neuromuscular junction. J. Neurobiol. 1994, 25, 722–739. [Google Scholar] [CrossRef]
- Czéh, G.; Gallego, R.; Kudo, N.; Kuno, M. Evidence for the maintenance of motoneurone properties by msucel activity. J. Physiol. 1978, 281, 239–252. [Google Scholar] [CrossRef]
- Funakoshi, H.; Belluardo, N.; Arenas, E.; Yamamoto, Y.; Casabona, A.; Persson, H.; Ibanez, C. Muscle-derived neurotrophin-4 as an activity-dependent trophic signal for adult motor neurons. Science (80-) 1995, 268, 1495–1499. [Google Scholar] [CrossRef] [Green Version]
- Proenca, C.C.; Song, M.; Lee, F.S. Differential effects of BDNF and neurotrophin 4 (NT4) on endocytic sorting of TrkB receptors. J. Neurochem. 2016, 138, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Garcia, N.; Tomàs, M.; Santafe, M.M.; Lanuza, M.A.; Besalduch, N.; Tomàs, J. Blocking p75NTR receptors alters polyinnervationz of neuromuscular synapses during development. J. Neurosci. Res. 2011, 89, 1331–1341. [Google Scholar] [CrossRef]
- Grondard, C.; Biondi, O.; Pariset, C.; Lopes, P.; Deforges, S.; Lécolle, S.; Della Gaspera, B.; Gallien, C.-L.; Chanoine, C.; Charbonnier, F. Exercise-Induced Modulation of Calcineurin Activity Parallels theTime Course of MyofibreTransitions. J. Cell. Physiol. 2008, 2008, 126–135. [Google Scholar] [CrossRef]
- Meeker, R.; Williams, K. Dynamic Nature of the p75 Neurotrophin Receptor in Response to Injury and Disease. J. Neuroimmune Pharmacol. 2014, 9, 615–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, G.T.; Radeke, M.J.; Kuo, R.C.; Makrides, V.; Hinkle, B.; Hoang, R.; Medina-Selby, A.; Coit, D.; Valenzuela, P.; Feinstein, S.C. Signal transduction mediated by the truncated trkB receptor isoforms, trkB.T1 and trkB.T2. J. Neurosci. 1997, 17, 2683–2690. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, S.G.; Lovering, R.M.; Renn, C.L.; Leitch, C.C.; Liu, X.; Tallon, L.J.; Sadzewicz, L.D.; Pratap, A.; Ott, S.; Sengamalay, N.; et al. Genetic deletion of trkB.T1 increases neuromuscular function. Am. J. Physiol. Physiol. 2012, 302, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Rose, C.R.; Blum, R.; Pichler, B.; Lepier, A.; Kafitz, K.W.; Konnerth, A. Truncated TrkB-T1 mediates neurotrophin-evoked calcium signalling in glia cells. Nature 2003, 426, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.; Garner, B. Evidence that truncated TrkB isoform, TrkB-Shc can regulate phosphorylated TrkB protein levels. Biochem. Biophys. Res. Commun. 2012, 420, 331–335. [Google Scholar] [CrossRef]
- Skup, M.; Dwornik, A.; Macias, M.; Sulejczak, D.; Wiater, M.; Czarkowska-Bauch, J. Long-term locomotor training up-regulates TrkB(FL) receptor-like proteins, brain-derived neurotrophic factor, and neurotrophin 4 with different topographies of expression in oligodendroglia and neurons in the spinal cord. Exp. Neurol. 2002, 176, 289–307. [Google Scholar] [CrossRef]
- Kim, S.-E.; Ko, I.; Ji, E.; Jin, J.; Hwang, L.; Kim, S.-H.; Cho, S.; Kim, C.-J.; Kim, K.; Kim, K.H. Treadmill Exercise Alleviates Circadian Rhythm Disruption-Induced Memory Deficits by Activation of Glucocorticoid Receptor and Brain-Derived Neurotrophic Factor-Dependent Pathway. Int. Neurourol. J. 2019, 23, S40–S49. [Google Scholar] [CrossRef]
- Lin, T.W.; Shih, Y.H.; Chen, S.J.; Lien, C.H.; Chang, C.Y.; Huang, T.Y.; Chen, S.H.; Jen, C.J.; Kuo, Y.M. Running exercise delays neurodegeneration in amygdala and hippocampus of Alzheimer’s disease (APP/PS1) transgenic mice. Neurobiol. Learn. Mem. 2015, 118, 189–197. [Google Scholar] [CrossRef]
- Bellingham, M.C. Pharmacological Dissection of G Protein-Mediated Second Messenger Cascades in Neurons. In Stimulation and Inhibition of Neurons; Humana Press: Totowa, NJ, USA, 2013; pp. 61–106. [Google Scholar]
- Obis, T.; Besalduch, N.; Hurtado, E.; Nadal, L.; Santafe, M.M.; Garcia, N.; Tomàs, M.; Priego, M.; Lanuza, M.A.; Tomàs, J. The novel protein kinase C epsilon isoform at the adult neuromuscular synapse: Location, regulation by synaptic activity-dependent muscle contraction through TrkB signaling and coupling to ACh release. Mol. Brain 2015, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Grondard, C.; Biondi, O.; Armand, A.-S.; Lécolle, S.; Della Gaspera, B.; Pariset, C.; Li, H.; Gallien, C.-L.; Vidal, P.-P.; Chanoine, C.; et al. Regular exercise prolongs survival in a type 2 spinal muscular atrophy model mouse. J. Neurosci. 2005, 25, 7615–7622. [Google Scholar] [CrossRef] [Green Version]
- Gundersen, K. Excitation-transcription coupling in skeletal muscle: The molecular pathways of exercise. Biol. Rev. 2011, 86, 564–600. [Google Scholar] [CrossRef] [Green Version]
- Rose, A.J.; Michell, B.J.; Kemp, B.E.; Hargreaves, M. Effect of exercise on protein kinase C activity and localization in human skeletal muscle. J. Physiol. 2004, 561, 861–870. [Google Scholar] [CrossRef]
- DiMario, J.X. Protein kinase C signaling controls skeletal muscle fiber types. Exp. Cell Res. 2001, 263, 23–32. [Google Scholar] [CrossRef]
- DiMario, J.X.; Funk, P.E. Protein kinase C activity regulates slow myosin heavy chain 2 gene expression in slow lineage skeletal muscle fibers. Dev. Dyn. 1999, 216, 177–189. [Google Scholar] [CrossRef]
- Jordan, T.; Jiang, H.; Li, H.; DiMario, J.X. Inhibition of ryanodine receptor 1 in fast skeletal muscle fibers induces a fast-to-slow muscle fiber type transition. J. Cell Sci. 2004, 117, 6175–6183. [Google Scholar] [CrossRef] [Green Version]
- Rykx, A.; De Kimpe, L.; Mikhalap, S.; Vantus, T.; Seufferlein, T.; Vandenheede, J.R.; Van Lint, J. Protein kinase D: A family affair. FEBS Lett. 2003, 546, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Liu, Y.; Tan, T.; Guha, S.; Gukovsky, I.; Gukovskaya, A.; Pandol, S.J. Protein Kinase D Regulates Cell Death Pathways in Experimental Pancreatitis. Front. Physiol. 2012, 3, 60. [Google Scholar] [CrossRef] [Green Version]
- Nagy, G.; Reim, K.; Matti, U.; Brose, N.; Binz, T.; Rettig, J.; Neher, E.; Sørensen, J.B. Regulation of releasable vesicle pool sizes by protein kinase A-dependent phosphorylation of SNAP-25. Neuron 2004, 41, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.J.; Slater, C.R. The contribution of postsynaptic folds to the safety factor for neuromuscular transmission in rat fast- and slow-twitch muscles. J. Physiol. 1997, 500, 165–176. [Google Scholar] [CrossRef]
- Vinay, L.; Brocard, F.; Clarac, F. Differential maturation of motoneurons innervating ankle flexor and extensor muscles in the neonatal rat. Eur. J. Neurosci. 2000, 12, 4562–4566. [Google Scholar] [CrossRef]
- Brocard, F.; Vinay, L.; Clarac, F. Development of hindlimb postural control during the first postnatal week in the rat. Dev. Brain Res. 1999, 117, 81–89. [Google Scholar] [CrossRef]
- Santana, L.F.; Chase, E.G.; Votaw, V.S.; Nelson, M.T.; Greven, R. Functional coupling of calcineurin and protein kinase A in mouse ventricular myocytes. J. Physiol. 2002, 544, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Chin, E.R.; Olson, E.N.; Richardson, J.A.; Yang, Q.; Humphries, C.; Shelton, J.M.; Wu, H.; Zhu, W.; Bassel-Duby, R.; Williams, R.S. A calcineurin-dependent transcriptional pathway controls skeletal muscle fiber type. Genes Dev. 1998, 12, 2499–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, Z.A.; Gundersen, K.; Buonanno, A. Activity-dependent repression of muscle genes by NFAT. Proc. Natl. Acad. Sci. USA 2008, 105, 5921–5926. [Google Scholar] [CrossRef] [Green Version]
- Olson, E.N.; Williams, R.S. Remodeling muscles with calcineurin. BioEssays 2000, 22, 510–519. [Google Scholar] [CrossRef]
- Seburn, K.; Coicou, C.; Gardiner, P. Effects of altered muscle activation on oxidative enzyme activity in rat α-motoneurons. J. Appl. Physiol. 1994, 77, 2269–2274. [Google Scholar] [CrossRef]
- Beaumont, E.; Gardiner, P. Effects of daily spontaneous running on the electrophysiological properties of hindlimb motoneurones in rats. J. Physiol. 2002, 540, 129–138. [Google Scholar] [CrossRef]
- Ishihara, A.; Kawano, F.; Ishioka, N.; Oishi, H.; Higashibata, A.; Shimazu, T.; Ohira, Y. Effects of running exercise during recovery from hindlimb unloading on soleus muscle fibers and their spinal motoneurons in rats. Neurosci. Res. 2004, 48, 119–127. [Google Scholar] [CrossRef]
- Gazula, V.R.; Roberts, M.; Luzzio, C.; Jawad, A.F.; Kalb, R.G. Effects of limb exercise after spinal cord injury on motor neuron dendrite stucture. J. Comp. Neurol. 2004, 476, 130–145. [Google Scholar] [CrossRef]
- Gardiner, P.; Dai, Y.; Heckman, C.J. Effects of exercise training on α-motoneurons. J. Appl. Physiol. 2006, 101, 1228–1236. [Google Scholar] [CrossRef]
- Petrie, M.A.; Sharma, A.; Taylor, E.B.; Suneja, M.; Shields, R.K. Impact of short- and long-term electrically induced muscle exercise on gene signaling pathways, gene expression, and PGC1a methylation in men with spinal cord injury. Physiol. Genom. 2020, 52, 71–80. [Google Scholar] [CrossRef]
- Gerchman, L.R.B.; Edgerton, V.R.; Carrow, R.E. Effects of physical training on the histochemistry and morphology of ventral motor neurons. Exp. Neurol. 1975, 49, 790–801. [Google Scholar] [CrossRef]
- Edström, J.-. E Effects of increased motor activity on the dimensions and the staining properties of the neuron soma. J. Comp. Neurol. 1957, 107, 295–304. [Google Scholar] [CrossRef]
- Dahlström, A.; Heiwall, P.-O.; Bööj, S.; Dahllöf, A.-G. The influence of supraspinal impulse activity on the intra-axonal transport of acetylcholine, choline acetyltransferase and acetylcholinesterase in rat motor neurons. Acta Physiol. Scand. 1978, 103, 308–319. [Google Scholar] [CrossRef]
- Jasmin, B.J.; Lavoie, P.A.; Gardiner, P.F. Fast axonal transport of acetylcholinesterase in rat sciatic motoneurons is enhanced following prolonged daily running, but not following swimming. Neurosci. Lett. 1987, 78, 156–160. [Google Scholar] [CrossRef]
- Jasmin, B.J.; Lavoie, P.A.; Gardiner, P.F. Fast axonal transport of labeled proteins in motoneurons of exercise-trained rats. Am. J. Physiol. Cell Physiol. 1988, 255, C731–C736. [Google Scholar] [CrossRef]
- Kang, C.M.; Lavoie, P.A.; Gardiner, P.F. Chronic exercise increases SNAP-25 abundance in fast-transported proteins of rat motoneurones. Neuroreport 1995, 6, 549–553. [Google Scholar] [CrossRef]
- Yan, Z.; Okutsu, M.; Akhtar, Y.N.; Lira, V.A. Regulation of exercise-induced fiber type transformation, mitochondrial biogenesis, and angiogenesis in skeletal muscle. J. Appl. Physiol. 2011, 110, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Hood, D.A. Mechanisms of exercise-induced mitochondrial biogenesis in skeletal muscle. Appl. Physiol. Nutr. Metab. 2009, 34, 465–472. [Google Scholar] [CrossRef]
- Skulachev, V.P. Mitochondrial filaments and clusters as intracellular power-transmitting cables. Trends Biochem. Sci. 2001, 26, 23–29. [Google Scholar] [CrossRef]
- Ljubicic, V.; Joseph, A.M.; Saleem, A.; Uguccioni, G.; Collu-Marchese, M.; Lai, R.Y.J.; Nguyen, L.M.D.; Hood, D.A. Transcriptional and post-transcriptional regulation of mitochondrial biogenesis in skeletal muscle: Effects of exercise and aging. Biochim. Biophys. Acta Gen. Subj. 2010, 1800, 223–234. [Google Scholar] [CrossRef]
- Gnaiger, E. Capacity of oxidative phosphorylation in human skeletal muscle. New perspectives of mitochondrial physiology. Int. J. Biochem. Cell Biol. 2009, 41, 1837–1845. [Google Scholar] [CrossRef]
- Baar, K.; Wende, A.R.; Jones, T.E.; Marison, M.; Nolte, L.A.; Chen, M.; Kelly, D.P.; Holloszy, J.O. Adaptations of skeletal muscle to exercise: Rapid increase in the transcriptional coactivator PGC-1. FASEB J. 2002, 16, 1879–1886. [Google Scholar] [CrossRef]
- Deldicque, L.; Atherton, P.; Patel, R.; Theisen, D.; Nielens, H.; Rennie, M.J.; Francaux, M. Effects of resistance exercise with and without creatine supplementation on gene expression and cell signaling in human skeletal muscle. J. Appl. Physiol. 2008, 104, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Roves, P.M.; Huss, J.; Holloszy, J.O. Role of calcineurin in exercise-induced mitochondrial biogenesis. Am. J. Physiol. Endocrinol. Metab. 2006, 290, 1172–1179. [Google Scholar] [CrossRef] [Green Version]
- Jäer, S.; Handschin, C.; St-Pierre, J.; Spiegelman, B.M. AMP-activated protein kinase (AMPK) action in skeletal muscle via direct phosphorylation of PGC-1α. Proc. Natl. Acad. Sci. USA 2007, 104, 12017–12022. [Google Scholar]
- Arnold, A.-S.; Gill, J.; Christe, M.; Ruiz, R.; McGuirk, S.; St-Pierre, J.; Tabares, L.; Handschin, C. Morphological and functional remodelling of the neuromuscular junction by skeletal muscle PGC-1α. Nat. Commun. 2014, 5, 3569. [Google Scholar] [CrossRef] [Green Version]
- Mech, A.M.; Brown, A.; Schiavo, G.; Sleigh, J.N. Morphological variability is greater at developing than mature mouse neuromuscular junctions. J. Anat. 2020, 237, 603–617. [Google Scholar] [CrossRef]
- Argaw, A.; Desaulniers, P.; Gardiner, P.F. Enhanced neuromuscular transmission efficacy in overloaded rat plantaris muscle. Muscle Nerve 2004, 29, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Fahim, M.A. Endurance exercise modulates neuromuscular junction of C57BL/6NNia aging mice. J. Appl. Physiol. 1997, 83, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Deschenes, M.R.; Roby, M.A.; Glass, E.K. Aging influences adaptations of the neuromuscular junction to endurance training. Neuroscience 2011, 190, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neto, W.; Ciena, A.P.; Anaruma, C.A.; Rodrigues De Souza, R.; Gama, E.F. Effects of exercise on neuromuscular junction components across age: Systematic review of animal experimental studies Neuroscience. BMC Res. Notes 2015, 8, 1–15. [Google Scholar]
- Gisiger, V.; Bélisle, M.; Gardiner, P.F. Acetylcholinesterase Adaptation to Voluntary Wheel Running is Proportional to the Volume of Activity in Fast, but not Slow, Rat Hindlimb Muscles. Eur. J. Neurosci. 1994, 6, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Desaulniers, P.; Lavoie, P.A.; Gardiner, P.F. Endurance training increases acetylcholine receptor quantity at neuromuscular junctions of adult rat skeletal muscle. Neuroreport 1998, 9, 3549–3552. [Google Scholar] [CrossRef]
- Acsadi, G.; Anguelov, R.A.; Yang, H.; Toth, G.; Thomas, R.; Jani, A.; Wang, Y.; Ianakova, E.; Mohammad, S.; Lewis, R.A.; et al. Increased Survival and Function of SOD1 Mice After Glial Cell-Derived Neurotrophic Factor Gene Therapy. Hum. Gene Ther. 2002, 13, 1047–1059. [Google Scholar] [CrossRef]
- Manabe, Y.; Nagano, I.; Gazi, M.S.A.; Murakami, T.; Shiote, M.; Shoji, M.; Kitagawa, H.; Setoguchi, Y.; Abe, K. Adenovirus-mediated gene transfer of glial cell line-derived neurotrophic factor prevents motor neuron loss of transgenic model mice for amyotrophic lateral sclerosis. Apoptosis 2002, 7, 329–334. [Google Scholar] [CrossRef]
- Sun, W.; Funakoshi, H.; Nakamura, T. Overexpression of HGF retards disease progression and prolongs life span in a transgenic mouse model of ALS. J. Neurosci. 2002, 22, 6537–6548. [Google Scholar] [CrossRef]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Greising, S.M.; Stowe, J.M.; Sieck, G.C.; Mantilla, C.B. Role of TrkB kinase activity in aging diaphragm neuromuscular junctions. Exp. Gerontol. 2015, 72, 184–191. [Google Scholar] [CrossRef]
- Nadal, L.; Garcia, N.; Hurtado, E.; Simó, A.; Tomàs, M.; Lanuza, M.A.; Santafé, M.; Tomàs, J. Presynaptic muscarinic acetylcholine autoreceptors (M1, M2 and M4 subtypes), adenosine receptors (A1 and A2A ) and tropomyosin-related kinase B receptor (TrkB) modulate the developmental synapse elimination process at the neuromuscular junction. Mol. Brain 2016, 9, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Delezie, J.; Weihrauch, M.; Maier, G.; Tejero, R.; Ham, D.J.; Gill, J.F.; Karrer-Cardel, B.; Rüegg, M.A.; Tabares, L.; Handschin, C. BDNF is a mediator of glycolytic fiber-type specification in mouse skeletal muscle. Proc. Natl. Acad. Sci. USA 2019, 116, 16111–16120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrann, C.D.; White, J.P.; Salogiannnis, J.; Laznik-Bogoslavski, D.; Wu, J.; Ma, D.; Lin, J.D.; Greenberg, M.E.; Spiegelman, B.M. Exercise induces hippocampal BDNF through a PGC-1α/FNDC5 pathway. Cell Metab. 2013, 18, 649–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dell, R.B.; Holleran, S.; Ramakrishnan, R. Sample Size Determination. ILAR J. 2002, 43, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods Applied to Experiments in Agriculture and Biology, 8th ed.; Iowa State University Press: Ames, IA, USA, 1989. [Google Scholar]
- Aldridge, G.M.; Podrebarac, D.M.; Greenough, W.T.; Weiler, I.J. The use of total protein stains as loading controls: An alternative to high-abundance single-protein controls in semi-quantitative immunoblotting. J. Neurosci. Methods 2008, 172, 250–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | SOL WT | PLA WT Run | PLA WT Swim | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NTFs and receptors | proBDNF | 1.05 | ± | 0.31 | 0.921 | 2.65 | ± | 0.89 | 0.008 | ** | 2.17 | ± | 0.93 | 0.045 | * | |
| mBDNF | 0.57 | ± | 0.27 | 0.021 | * | 0.95 | ± | 0.29 | 0.736 | 0.45 | ± | 0.18 | 0.002 | ** | ||
| NT4 | 1.89 | ± | 0.03 | 0.000 | *** | 1.81 | ± | 0.10 | 0.000 | *** | 1.51 | ± | 0.14 | 0.000 | *** | |
| p75NTR | 1.11 | ± | 0.03 | 0.393 | 1.68 | ± | 0.24 | 0.001 | ** | 0.79 | ± | 0.10 | 0.006 | ** | ||
| TrkB.T1 | 0.92 | ± | 0.06 | 0.619 | 2.45 | ± | 0.14 | 0.000 | *** | 0.92 | ± | 0.14 | 0.305 | |||
| TrkB.FL | 0.63 | ± | 0.04 | 0.012 | ** | 1.95 | ± | 0.62 | 0.022 | * | 1.84 | ± | 0.54 | 0.021 | * | |
| pTrkB.FL (Y816) | 0.19 | ± | 0.01 | 0.000 | *** | 1.23 | ± | 0.14 | 0.014 | * | 0.40 | ± | 0.16 | 0.000 | *** | |
| Serine-threonine kinases | PDK1 | 1.35 | ± | 0.09 | 0.089 | 1.94 | ± | 0.51 | 0.033 | * | 0.62 | ± | 0.09 | 0.002 | ** | |
| pPDK1 (S241) | 0.99 | ± | 0.26 | 0.998 | 0.96 | ± | 0.08 | 0.452 | 0.99 | ± | 0.05 | 0.824 | ||||
| cPKCα | 1.19 | ± | 0.01 | 0.258 | 0.81 | ± | 0.22 | 0.222 | 0.68 | ± | 0.15 | 0.020 | * | |||
| pcPKCα (S657) | 0.61 | ± | 0.03 | 0.014 | ** | 1.42 | ± | 0.18 | 0.008 | ** | 0.95 | ± | 0.20 | 0.688 | ||
| cPKCβI | 0.15 | ± | 0.11 | 0.000 | *** | 0.99 | ± | 0.53 | 0.976 | 0.53 | ± | 0.23 | 0.023 | * | ||
| pcPKCβI (T621) | 0.51 | ± | 0.22 | 0.000 | *** | 1.11 | ± | 0.18 | 0.328 | 0.56 | ± | 0.08 | 0.000 | *** | ||
| nPKCε | 1.65 | ± | 0.32 | 0.000 | *** | 1.41 | ± | 0.28 | 0.017 | * | 0.76 | ± | 0.20 | 0.099 | ||
| pnPKCε (S729) | 0.77 | ± | 0.06 | 0.140 | 0.89 | ± | 0.26 | 0.508 | 0.73 | ± | 0.20 | 0.085 | ||||
| PKA Cα | 2.04 | ± | 0.07 | 0.000 | *** | 1.09 | ± | 0.14 | 0.270 | 1.29 | ± | 0.42 | 0.215 | |||
| PKA Cβ | 1.13 | ± | 0.43 | 0.648 | 1.09 | ± | 0.11 | 0.147 | 0.60 | ± | 0.24 | 0.014 | * | |||
| PKA RIα | 2.4 | ± | 0.56 | 0.000 | *** | 0.77 | ± | 0.16 | 0.032 | * | 1.18 | ± | 0.37 | 0.364 | ||
| PKA RIβ | 1.15 | ± | 0.01 | 0.776 | 0.98 | ± | 0.22 | 0.846 | 0.83 | ± | 0.31 | 0.312 | ||||
| PKA RIIα | 0.79 | ± | 0.09 | 0.583 | 0.76 | ± | 0.16 | 0.027 | * | 0.94 | ± | 0.19 | 0.553 | |||
| PKA RIIβ | 1.98 | ± | 0.07 | 0.006 | ** | 1.76 | ± | 0.41 | 0.013 | * | 1.34 | ± | 0.28 | 0.049 | * | |
| Exocytosis machinery | Munc18-1 | 0.75 | ± | 0.12 | 0.081 | 1.30 | ± | 0.06 | 0.000 | *** | 0.47 | ± | 0.12 | 0.000 | *** | |
| pMunc18-1 (S313) | 0.54 | ± | 0.11 | 0.000 | *** | 0.65 | ± | 0.17 | 0.004 | ** | 0.70 | ± | 0.18 | 0.017 | * | |
| SNAP-25 | 0.75 | ± | 0.17 | 0.050 | 1.06 | ± | 0.17 | 0.515 | 0.40 | ± | 0.08 | 0.000 | *** | |||
| pSNAP-25 (S187) | 2.31 | ± | 0.06 | 0.000 | *** | 1.03 | ± | 0.15 | 0.700 | 0.47 | ± | 0.07 | 0.000 | *** | ||
| pSNAP-25 (T138) | 0.35 | ± | 0.21 | 0.000 | *** | 1.27 | ± | 0.19 | 0.158 | 0.53 | ± | 0.16 | 0.002 | ** | ||
| B |  | |||||||||||||||
| Target | Source | Reference | Dilution | Target | Source | Reference | Dilution |
|---|---|---|---|---|---|---|---|
| BDNF | Rb pAb | Sc-20981 | 1/500 | PKA Cα | Rb pAb | Sc-903 | 1/1000 |
| NT4 | Rb pAb | Sc-545 | 1/500 | PKA Cβ | Rb pAb | Sc-904 | 1/1000 |
| p75NTR | Rb pAb | 07-476 | 1/800 | PKA RIα | Ms mAb | Sc-136231 | 1/1000 |
| TrkB | Ms mAb | Sc-377218 | 1/1000 | PKA RIβ | Rb pAb | Sc-907 | 1/1000 |
| pTrkB (Y816) | Rb pAb | ABN1381 | 1/1000 | PKA RIIα | Rb pAb | Sc-909 | 1/1000 |
| PDK1 | Ms mAb | Sc-17765 | 1/1000 | PKA RIIβ | Ms mAb | Sc-376778 | 1/1000 |
| pDPK1 (S241) | Rb pAb | #3061 | 1/1000 | Munc18-1 | Rb mAb | 13414 | 1/1000 |
| cPKCα | Rb pAb | Sc-208 | 1/800 | pMunc18-1 (S313) | Rb pAb | Ab138687 | 1/1000 |
| pcPKCα (S657) | Rb pAb | 06-822 | 1/1000 | SNAP-25 | Rb mAb | #5309 | 1/1000 |
| cPKCβI | Rb pAb | Sc-209 | 1/1000 | pSNAP-25 (S187) | Rb pAb | Ab169871 | 1/1000 |
| pcPKCβI (T641) | Rb pAb | Ab75657 | 1/1000 | pSNAP-25 (T138) | Rb pAb | Orb163730 | 1/1000 |
| nPKCε | Rb pAb | Sc-214 | 1/1000 | HRP-conjugated | Dk a-Rb pAb | 711-035-152 | 1/10.000 |
| pnPKCε (S729) | Rb pAb | Sc-12355 | 1/1000 | HRP-conjugated | Rb a-Ms pAb | A9044 | 1/10.000 |
| ChAT | Gt pAb | AB144P | 1/400 | Cy3 | Dk a-Gt pAb | 705-165-003 | 1/400 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Just-Borràs, L.; Cilleros-Mañé, V.; Hurtado, E.; Biondi, O.; Charbonnier, F.; Tomàs, M.; Garcia, N.; Tomàs, J.; Lanuza, M.A. Running and Swimming Differently Adapt the BDNF/TrkB Pathway to a Slow Molecular Pattern at the NMJ. Int. J. Mol. Sci. 2021, 22, 4577. https://doi.org/10.3390/ijms22094577
Just-Borràs L, Cilleros-Mañé V, Hurtado E, Biondi O, Charbonnier F, Tomàs M, Garcia N, Tomàs J, Lanuza MA. Running and Swimming Differently Adapt the BDNF/TrkB Pathway to a Slow Molecular Pattern at the NMJ. International Journal of Molecular Sciences. 2021; 22(9):4577. https://doi.org/10.3390/ijms22094577
Chicago/Turabian StyleJust-Borràs, Laia, Víctor Cilleros-Mañé, Erica Hurtado, Olivier Biondi, Frédéric Charbonnier, Marta Tomàs, Neus Garcia, Josep Tomàs, and Maria A. Lanuza. 2021. "Running and Swimming Differently Adapt the BDNF/TrkB Pathway to a Slow Molecular Pattern at the NMJ" International Journal of Molecular Sciences 22, no. 9: 4577. https://doi.org/10.3390/ijms22094577