
Live Cell FRET Imaging Reveals Amyloid β-Peptide Oligomerization in Hippocampal Neurons

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
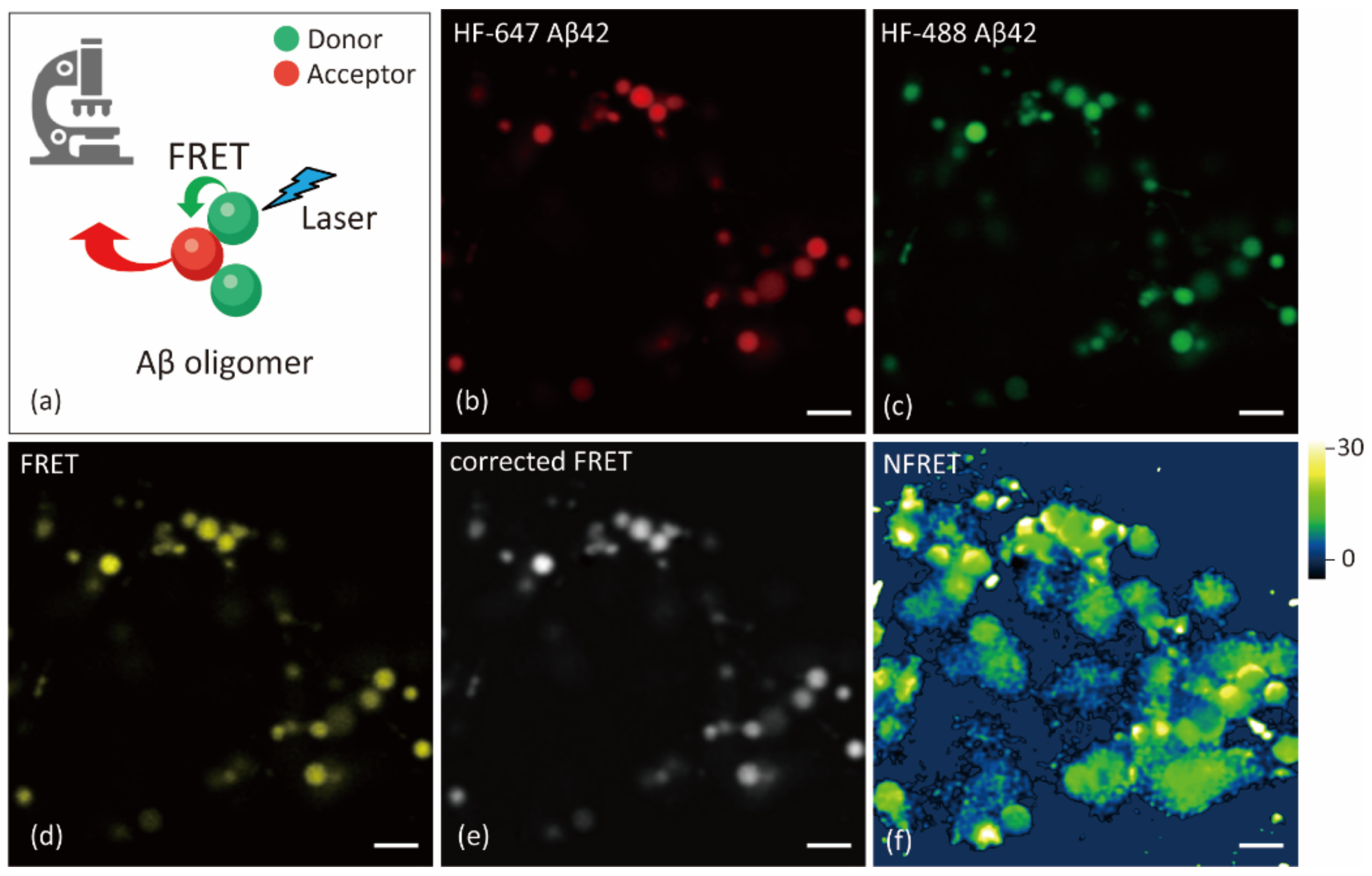
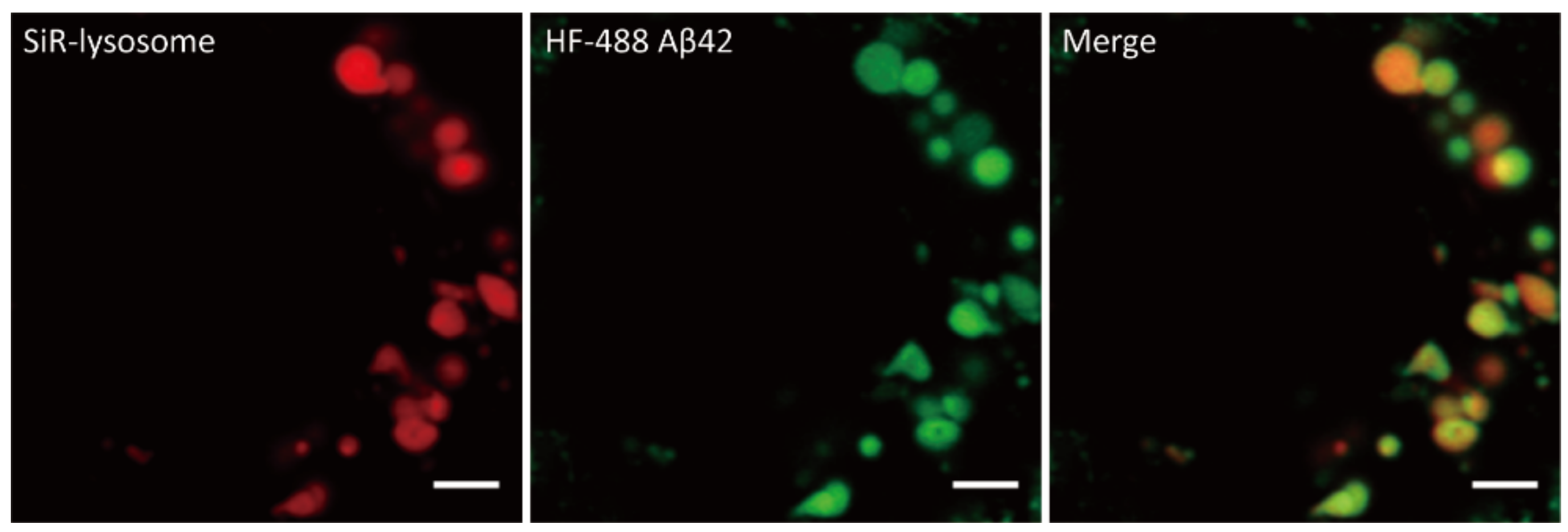
2.1. Internalized Aβ Oligomerizes in Lysosomes/Late Endosomes
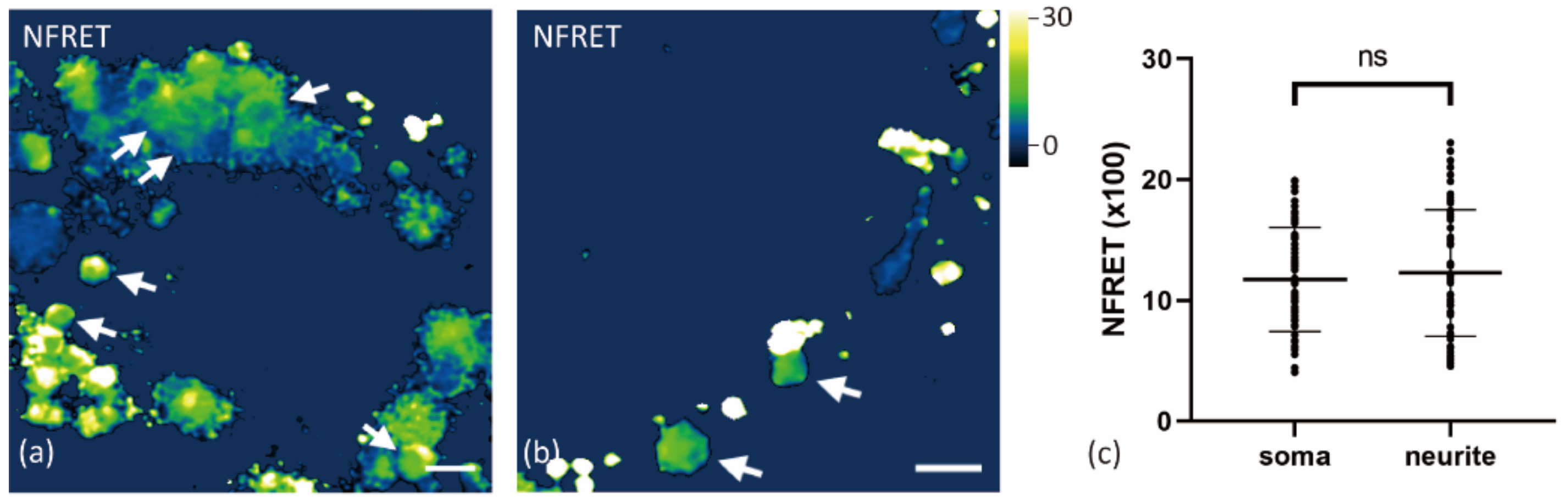
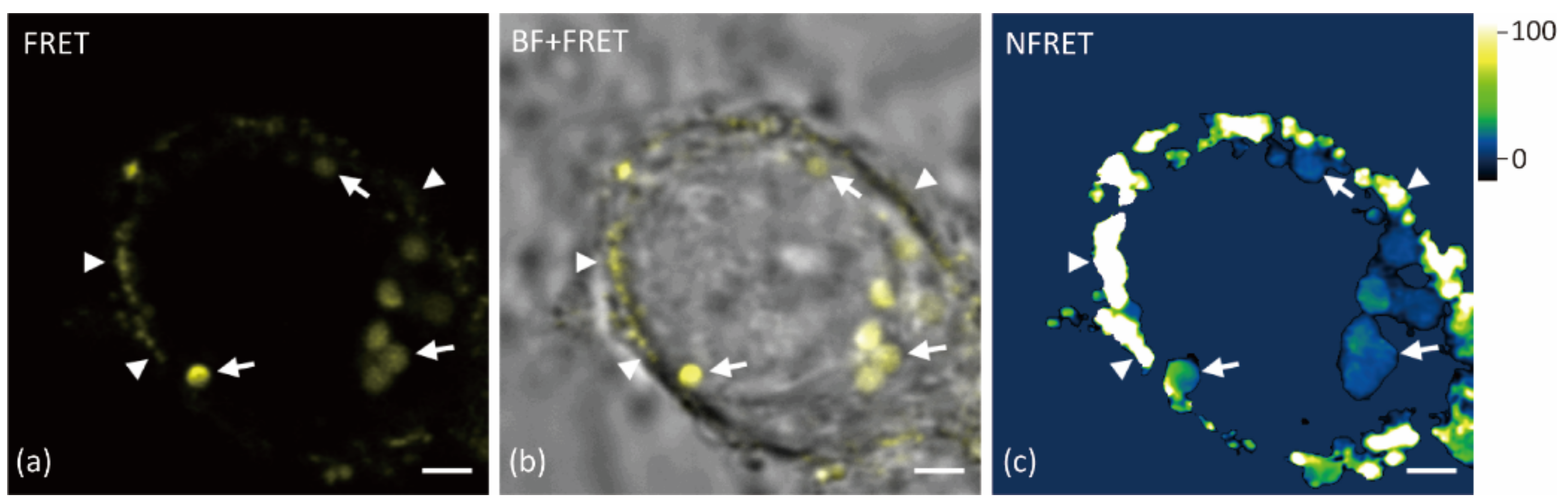
2.2. Aβ42 Oligomerization Occurs at Both Surface and Vesicles of Neurons
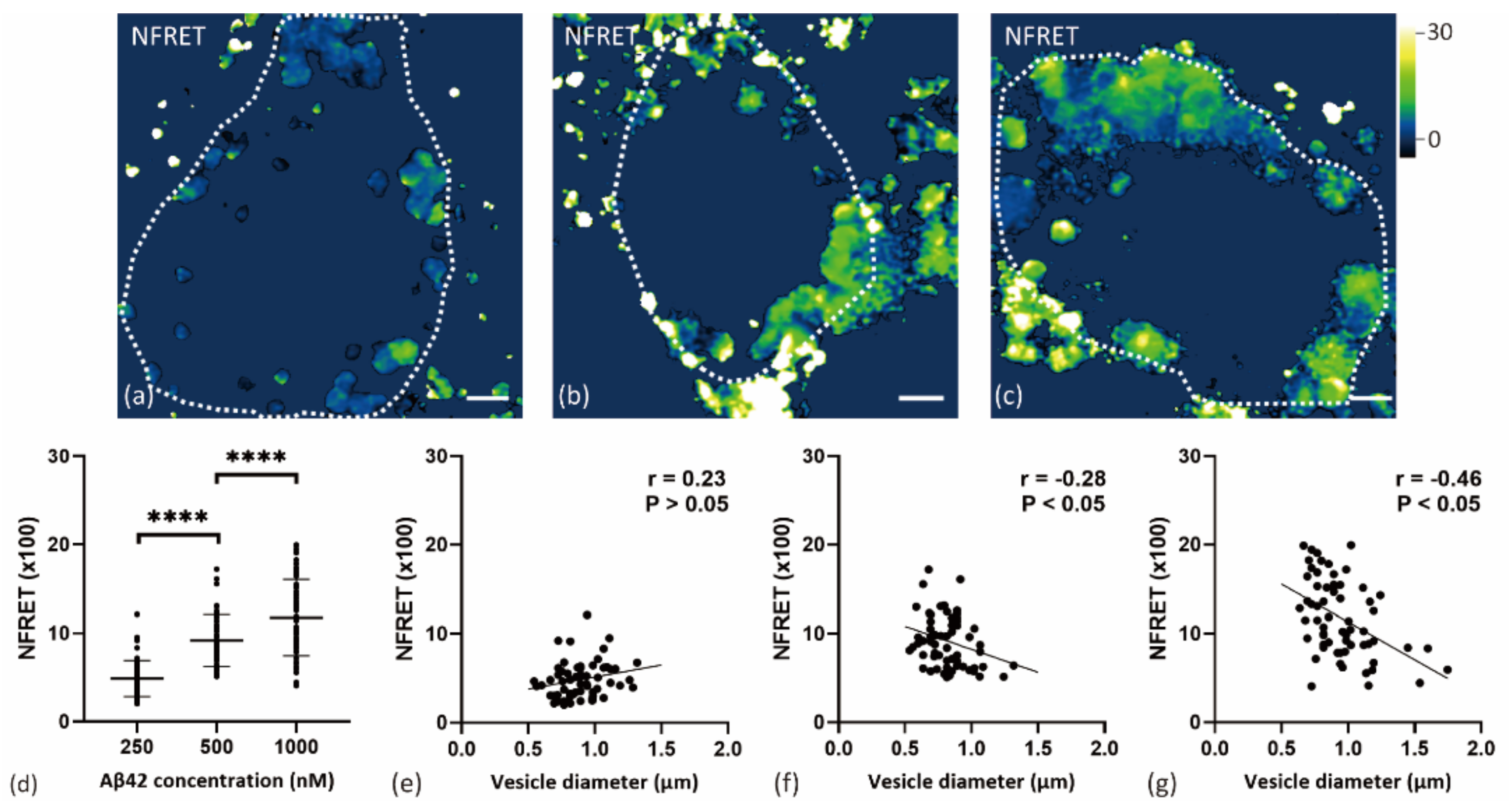
2.3. Internalized Aβ42 Oligomerization in Soma Is Concentration-Dependent
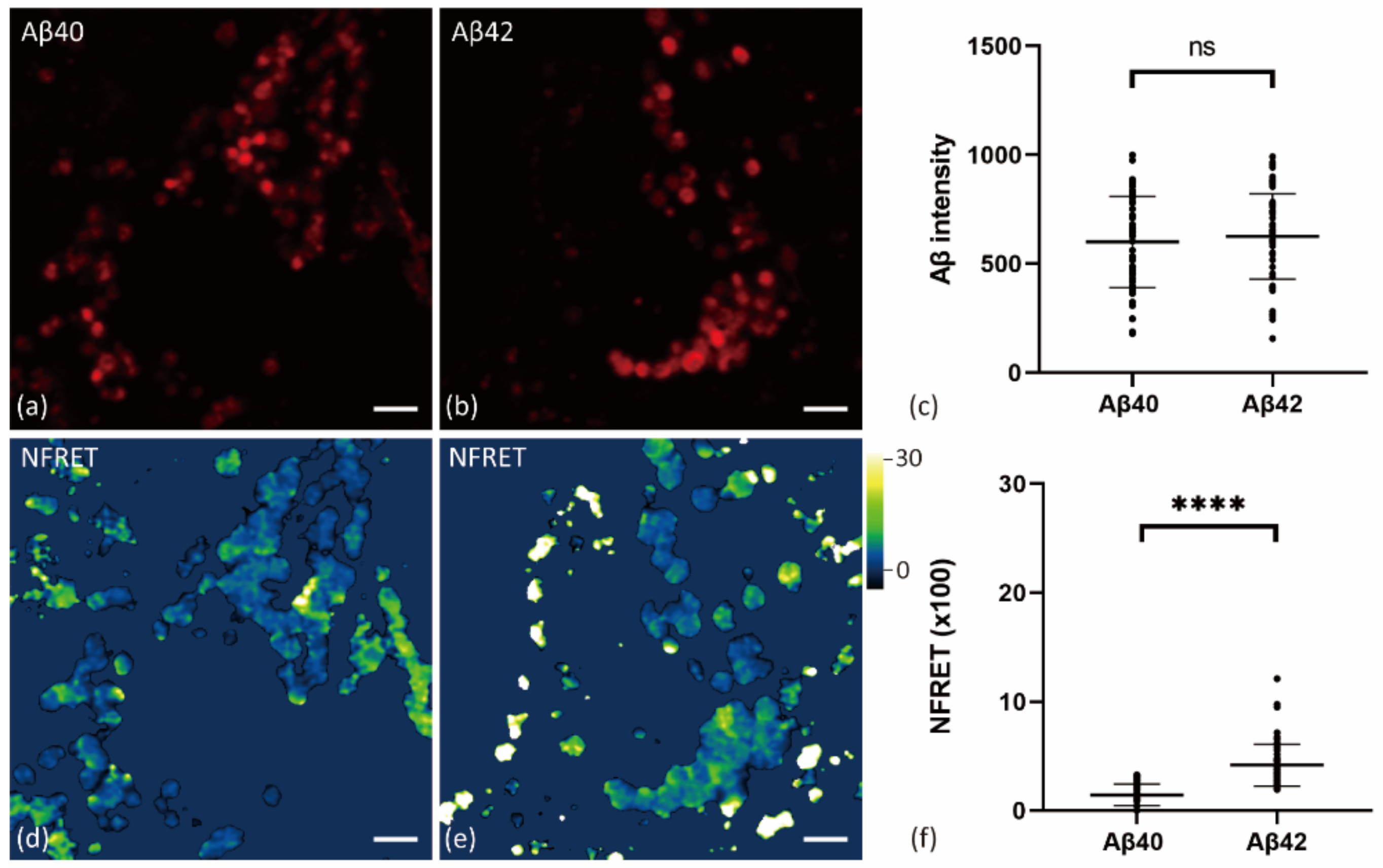
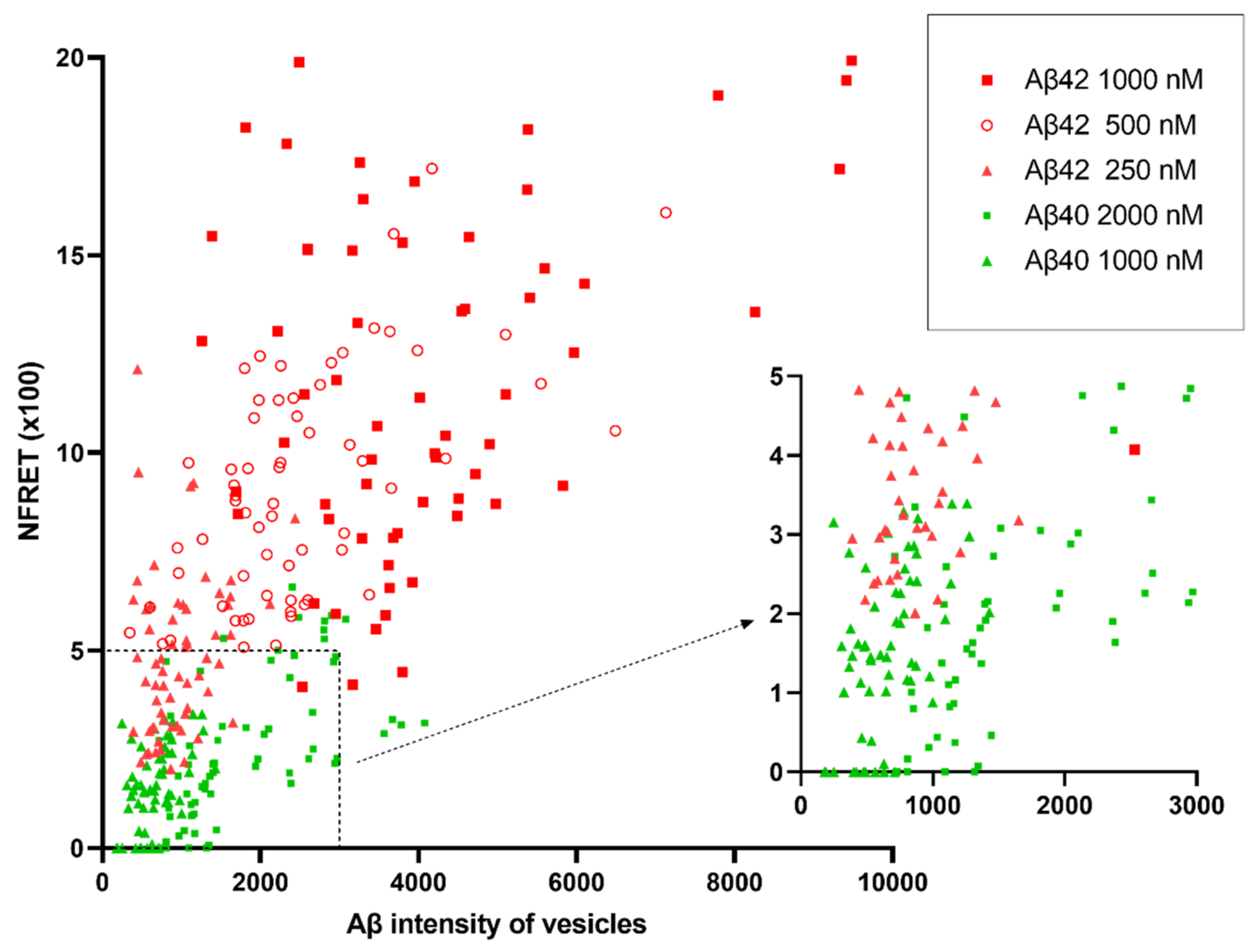
2.4. Internalized Aβ42 Oligomerizes in Neurons With Higher Efficiency Than Aβ40
3. Discussion
4. Materials and Methods
4.1. Synthetic Human Aβ Peptide and Preparation
4.2. Primary Neuron Culture
4.3. Lysosomes/Late Endosomes Labeling in Live Cell Imaging
4.4. Confocal Microscope and FRET Measurements
4.5. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballard, C.; Gauthier, S.; Corbett, A.; Brayne, C.; Aarsland, D.; Jones, E. Alzheimer’s Disease. Lancet 2011, 377, 1019–1031. [Google Scholar] [CrossRef]
- 2020 Alzheimer’s Disease Facts and Figures. Alzheimers Dement. 2020, 16, 391–460. [CrossRef] [PubMed]
- Viola, K.L.; Klein, W.L. Amyloid β Oligomers in Alzheimer’s Disease Pathogenesis, Treatment, and Diagnosis. Acta Neuropathol. 2015, 129, 183–206. [Google Scholar] [CrossRef]
- LaFerla, F.M.; Green, K.N.; Oddo, S. Intracellular Amyloid-β in Alzheimer’s Disease. Nat. Rev. Neurosci. 2007, 8, 499–509. [Google Scholar] [CrossRef]
- Turner, R.S.; Suzuki, N.; Chyung, A.S.; Younkin, S.G.; Lee, V.M.-Y. Amyloids Beta40 and Beta42 Are Generated Intracellularly in Cultured Human Neurons and Their Secretion Increases with Maturation. J. Biol. Chem. 1996, 271, 8966–8970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tienari, P.J.; Ida, N.; Ikonen, E.; Simons, M.; Weidemann, A.; Multhaup, G.; Masters, C.L.; Dotti, C.G.; Beyreuther, K. Intracellular and Secreted Alzheimer Beta-Amyloid Species Are Generated by Distinct Mechanisms in Cultured Hippocampal Neurons. Proc. Natl. Acad. Sci. USA 1997, 94, 4125–4130. [Google Scholar] [CrossRef] [Green Version]
- Walsh, D.M.; Selkoe, D.J. Aβ Oligomers—A Decade of Discovery. J. Neurochem. 2007, 101, 1172–1184. [Google Scholar] [CrossRef] [PubMed]
- Karisetty, B.C.; Bhatnagar, A.; Armour, E.M.; Beaver, M.; Zhang, H.; Elefant, F. Amyloid-β Peptide Impact on Synaptic Function and Neuroepigenetic Gene Control Reveal New Therapeutic Strategies for Alzheimer’s Disease. Front. Mol. Neurosci. 2020, 13. [Google Scholar] [CrossRef]
- Hashimoto, M.; Bogdanovic, N.; Volkmann, I.; Aoki, M.; Winblad, B.; Tjernberg, L.O. Analysis of Microdissected Human Neurons by a Sensitive ELISA Reveals a Correlation between Elevated Intracellular Concentrations of Abeta42 and Alzheimer’s Disease Neuropathology. Acta Neuropathol. 2010, 119, 543–554. [Google Scholar] [CrossRef]
- Aoki, M.; Volkmann, I.; Tjernberg, L.O.; Winblad, B.; Bogdanovic, N. Amyloid β-Peptide Levels in Laser Capture Microdissected Cornu Ammonis 1 Pyramidal Neurons of Alzheimer’s Brain. NeuroReport 2008, 19, 1085–1089. [Google Scholar] [CrossRef]
- Nag, S.; Sarkar, B.; Bandyopadhyay, A.; Sahoo, B.; Sreenivasan, V.K.A.; Kombrabail, M.; Muralidharan, C.; Maiti, S. Nature of the Amyloid-β Monomer and the Monomer-Oligomer Equilibrium. J. Biol. Chem. 2011, 286, 13827–13833. [Google Scholar] [CrossRef] [Green Version]
- Hayden, E.Y.; Teplow, D.B. Amyloid β-Protein Oligomers and Alzheimer’s Disease. Alzheimers Res. Ther. 2013, 5, 60. [Google Scholar] [CrossRef] [Green Version]
- Bruggink, K.A.; Müller, M.; Kuiperij, H.B.; Verbeek, M.M. Methods for Analysis of Amyloid-β Aggregates. J. Alzheimers Dis. 2012, 28, 735–758. [Google Scholar] [CrossRef]
- Algar, W.R.; Hildebrandt, N.; Vogel, S.S.; Medintz, I.L. FRET as a Biomolecular Research Tool—Understanding Its Potential While Avoiding Pitfalls. Nat. Methods 2019, 16, 815–829. [Google Scholar] [CrossRef]
- Wesén, E.; Gallud, A.; Paul, A.; Lindberg, D.J.; Malmberg, P.; Esbjörner, E.K. Cell Surface Proteoglycan-Mediated Uptake and Accumulation of the Alzheimer’s Disease Peptide Aβ(1–42). Biochim. Biophys. Acta BBA Biomembr. 2018, 1860, 2204–2214. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Crick, S.L.; Bu, G.; Frieden, C.; Pappu, R.V.; Lee, J.-M. Amyloid Seeds Formed by Cellular Uptake, Concentration, and Aggregation of the Amyloid-Beta Peptide. Proc. Natl. Acad. Sci. USA 2009, 106, 20324–20329. [Google Scholar] [CrossRef] [Green Version]
- Wennmalm, S.; Chmyrov, V.; Widengren, J.; Tjernberg, L. Highly Sensitive FRET-FCS Detects Amyloid β-Peptide Oligomers in Solution at Physiological Concentrations. Anal. Chem. 2015, 87, 11700–11705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mroczko, B.; Groblewska, M.; Litman-Zawadzka, A.; Kornhuber, J.; Lewczuk, P. Amyloid β Oligomers (AβOs) in Alzheimer’s Disease. J. Neural Transm. 2018, 125, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Burdick, D.; Soreghan, B.; Kwon, M.; Kosmoski, J.; Knauer, M.; Henschen, A.; Yates, J.; Cotman, C.; Glabe, C. Assembly and Aggregation Properties of Synthetic Alzheimer’s A4/Beta Amyloid Peptide Analogs. J. Biol. Chem. 1992, 267, 546–554. [Google Scholar] [CrossRef]
- Kulkarni, V.V.; Maday, S. Neuronal Endosomes to Lysosomes: A Journey to the Soma. J. Cell Biol. 2018, 217, 2977–2979. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-I.; Yi, J.-S.; Ko, Y.-G. Amyloid β Oligomerization Is Induced by Brain Lipid Rafts. J. Cell. Biochem. 2006, 99, 878–889. [Google Scholar] [CrossRef]
- Xia, Z.; Liu, Y. Reliable and Global Measurement of Fluorescence Resonance Energy Transfer Using Fluorescence Microscopes. Biophys. J. 2001, 81, 2395–2402. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Chang, P.-T. Acidic PH Promotes the Formation of Toxic Fibrils from β-Amyloid Peptide. Brain Res. 2001, 893, 287–291. [Google Scholar] [CrossRef]
- Jarrett, J.T.; Berger, E.P.; Lansbury, P.T. The Carboxy Terminus of the.Beta. Amyloid Protein Is Critical for the Seeding of Amyloid Formation: Implications for the Pathogenesis of Alzheimer’s Disease. Biochemistry 1993, 32, 4693–4697. [Google Scholar] [CrossRef]
- Asami-Odaka, A.; Ishibashi, Y.; Kikuchi, T.; Kitada, C.; Suzuki, N. Long Amyloid Beta-Protein Secreted from Wild-Type Human Neuroblastoma IMR-32 Cells. Biochemistry 1995, 34, 10272–10278. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Jans, D.C.; Winblad, B.; Tjernberg, L.O.; Schedin-Weiss, S. Neuronal Aβ42 Is Enriched in Small Vesicles at the Presynaptic Side of Synapses. Life Sci. Alliance 2018, 1, e201800028. [Google Scholar] [CrossRef] [PubMed]
- Feige, J.N.; Sage, D.; Wahli, W.; Desvergne, B.; Gelman, L. PixFRET, an ImageJ Plug-in for FRET Calculation That Can Accommodate Variations in Spectral Bleed-Throughs. Microsc. Res. Tech. 2005, 68, 51–58. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Wennmalm, S.; Winblad, B.; Schedin-Weiss, S.; Tjernberg, L.O. Live Cell FRET Imaging Reveals Amyloid β-Peptide Oligomerization in Hippocampal Neurons. Int. J. Mol. Sci. 2021, 22, 4530. https://doi.org/10.3390/ijms22094530
Gao Y, Wennmalm S, Winblad B, Schedin-Weiss S, Tjernberg LO. Live Cell FRET Imaging Reveals Amyloid β-Peptide Oligomerization in Hippocampal Neurons. International Journal of Molecular Sciences. 2021; 22(9):4530. https://doi.org/10.3390/ijms22094530
Chicago/Turabian StyleGao, Yang, Stefan Wennmalm, Bengt Winblad, Sophia Schedin-Weiss, and Lars O. Tjernberg. 2021. "Live Cell FRET Imaging Reveals Amyloid β-Peptide Oligomerization in Hippocampal Neurons" International Journal of Molecular Sciences 22, no. 9: 4530. https://doi.org/10.3390/ijms22094530