
The Enigma of the Adrenarche: Identifying the Early Life Mechanisms and Possible Role in Postnatal Brain Development

 , , ,
, , , 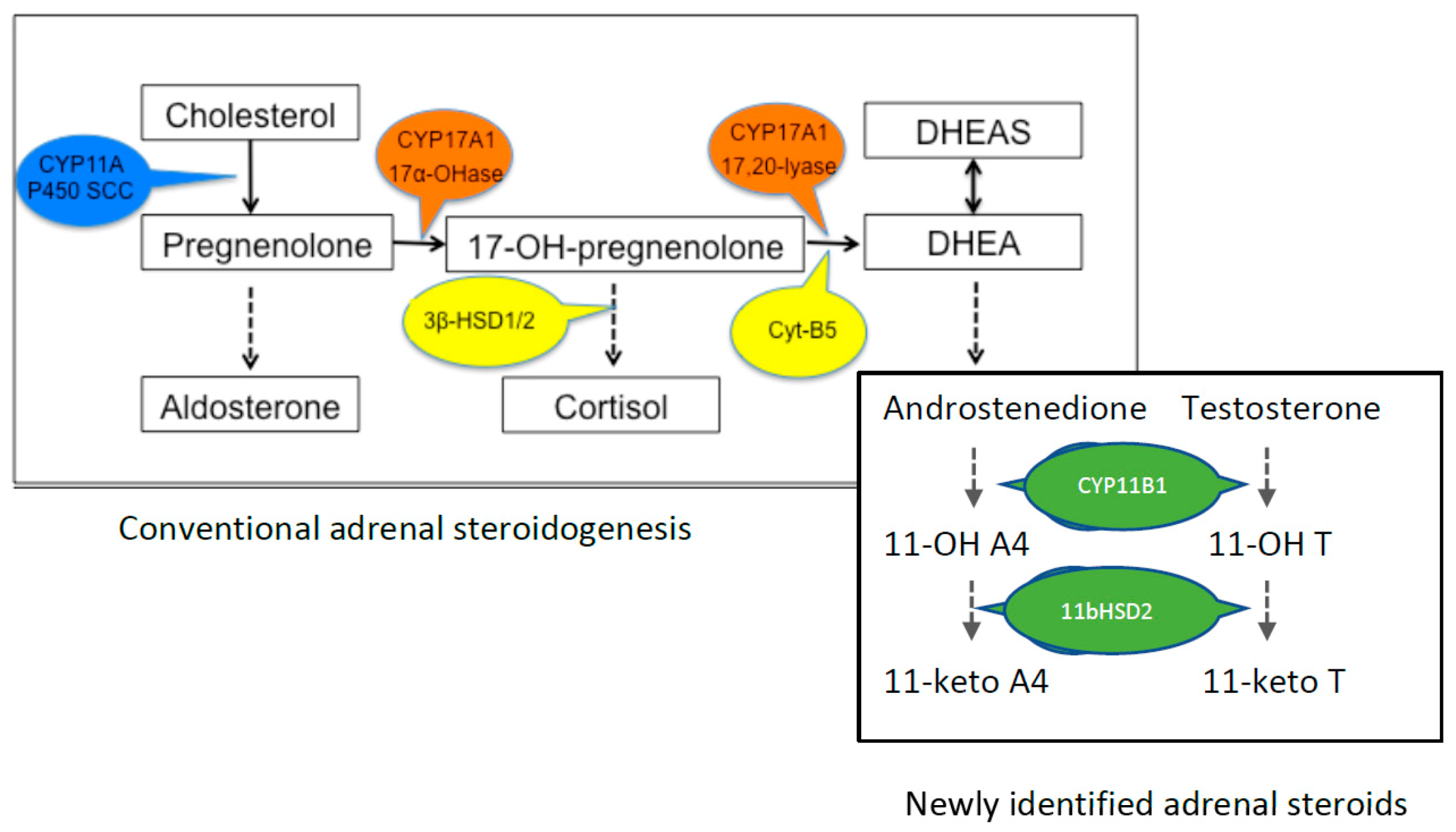{kind=link}
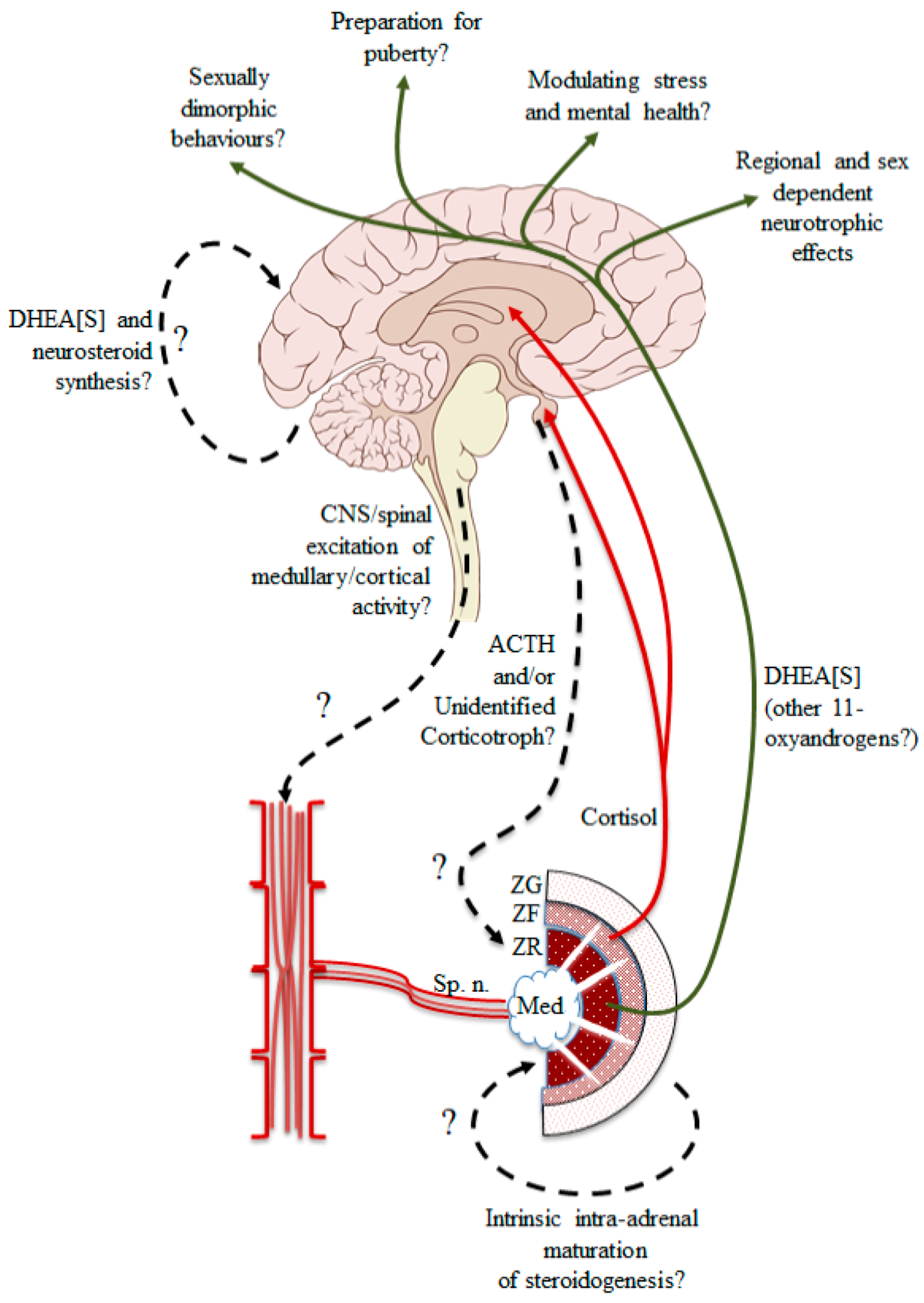{kind=link}
Abstract
:1. Introduction
- (1)
- What are the fundamental mechanisms that lead to the dramatic remodeling of the adrenal gland in early childhood?
- (2)
- Are they intrinsic to the development of the adrenal gland, or dependent on coordinated changes in the brain?
- (3)
- What are the functional consequences of adrenarche for brain development and the impacts on behavior in adolescence and early adulthood?
2. Production of DHEA
3. Why Is This Important?
4. Relationship of DHEA[S] with Psychiatric Disorders and the Potential Developmental Origins of Abnormal Adrenarche
5. A New Approach to an Old Problem
6. Conclusions: What Needs to Be Done Now?
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Campbell, B. DHEAS and Human Development: An Evolutionary Perspective. Front. Endocrinol. 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenfield, R.L. Normal and Premature Adrenarche. Endocr. Rev. 2021. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M.L.; Whittle, S.; Vijayakumar, N.; Dennison, M.; Simmons, J.G.; Allen, N.B. A systematic review of adrenarche as a sensitive period in neurobiological development and mental health. Dev. Cogn. Neurosci. 2017, 25, 12–28. [Google Scholar] [CrossRef]
- Barendse, M.E.A.; Simmons, J.G.; Byrne, M.L.; Patton, G.; Mundy, L.; Olsson, C.A.; Seal, M.L.; Allen, N.B.; Whittle, S. Associations between adrenarcheal hormones, amygdala functional connectivity and anxiety symptoms in children. Psychoneuroendocrinology 2018, 97, 156–163. [Google Scholar] [CrossRef]
- Liimatta, J.; Jääskeläinen, J.; Karvonen, A.M.; Remes, S.; Voutilainen, R.; Pekkanen, J. Tracking of serum DHEAS concentrations from age 1 to 6 years: A prospective cohort study. J. Endocr. Soc. 2020, 4, bvaa012. [Google Scholar] [CrossRef]
- Palmert, M.R.; Hayden, D.L.; Mansfield, M.J.; Crigler, J.F., Jr.; Crowley, W.F., Jr.; Chandler, D.W.; Boepple, P.A. The Longitudinal Study of Adrenal Maturation during Gonadal Suppression: Evidence that Adrenarche Is a Gradual Process. J. Clin. Endocrinol. Metab. 2001, 86, 4536–4542. [Google Scholar] [CrossRef]
- Remer, T.; Boye, K.R.; Hartmann, M.F.; Wudy, S.A. Urinary markers of adrenarche: Reference values in healthy subjects, aged 3–18 years. J. Clin. Endocrinol. Metab. 2005, 90, 2015–2021. [Google Scholar] [CrossRef]
- Francois, I.; De Zegher, F. Adrenarche and fetal growth. Pediatr. Res. 1997, 41, 440–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulte, S.; Schreiner, F.; Plamper, M.; Kasner, C.; Gruenewald, M.; Bartmann, P.; Fimmers, R.; Hartmann, M.F.; Wudy, S.A.; Stoffel-Wagner, B.; et al. Influence of Prenatal Environment on Androgen Steroid Metabolism in Monozygotic Twins with Birthweight Differences. J. Clin. Endocrinol. Metab. 2020, 105, e3672–e3687. [Google Scholar] [CrossRef]
- Nguyen, T.V. Developmental effects of androgens in the human brain. J. Neuroendocrinol. 2018, 30. [Google Scholar] [CrossRef] [PubMed]
- Conley, A.J.; Bernstein, R.M.; Nguyen, A.D. Adrenarche in nonhuman primates: The evidence for it and the need to redefine it. J. Endocrinol. 2012, 214, 121–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumontet, T.; Sahut-Barnola, I.; Septier, A.; Montanier, N.; Plotton, I.; Roucher-Boulez, F.; Ducros, V.; Lefrançois-Martinez, A.-M.; Pointud, J.-C.; Zubair, M. Adrenocortical development: Lessons from mouse models. Ann. Endocrinol. 2018, 79, 95–97. [Google Scholar] [CrossRef]
- Lincoln, D.W.; Fraser, H.M.; Lincoln, G.A.; Martin, G.B.; McNeilly, A.S. Hypothalamic pulse generators. Recent Prog. Horm. Res. 1985, 41, 369–419. [Google Scholar] [CrossRef]
- van Weerden, W.M.; Bierings, H.G.; van Steenbrugge, G.J.; de Jong, F.H.; Schroder, F.H. Adrenal glands of mouse and rat do not synthesize androgens. Life Sci. 1992, 50, 857–861. [Google Scholar] [CrossRef]
- Keegan, C.E.; Hammer, G.D. Recent insights into organogenesis of the adrenal cortex. Trends Endocrinol. Metab. 2002, 13, 200–208. [Google Scholar] [CrossRef]
- Holmes, P.V.; Dickson, A.D. X-zone degeneration in the adrenal glands of adult and immature female mice. J. Anat. 1971, 108, 159–168. [Google Scholar] [PubMed]
- Bornstein, S.R.; Chrousos, G.P. Adrenocorticotropin (ACTH)- and Non-ACTH-Mediated Regulation of the Adrenal Cortex: Neural and Immune Inputs. J. Clin. Endocrinol. Metab. 1999, 84, 1729–1736. [Google Scholar] [CrossRef] [PubMed]
- L’Allemand, D.; Biason-Lauber, A. Intra-adrenal regulation of androgen synthesis. Eur. J. Clin. Investig. 2000, 30 (Suppl. S3), 28–33. [Google Scholar] [CrossRef] [PubMed]
- Ishimoto, H.; Jaffe, R.B. Development and function of the human fetal adrenal cortex: A key component in the feto-placental unit. Endocr. Rev. 2011, 32, 317–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.-C.J.; Kang, Y. The transient cortical zone in the adrenal gland: The mystery of the adrenal X-zone. J. Endocrinol. 2019, 241, R51–R63. [Google Scholar] [CrossRef]
- Rege, J.; Garber, S.; Conley, A.J.; Elsey, R.M.; Turcu, A.F.; Auchus, R.J.; Rainey, W.E. Circulating 11-oxygenated androgens across species. J. Steroid Biochem. Mol. Biol. 2019, 190, 242–249. [Google Scholar] [CrossRef]
- Rege, J.; Turcu, A.F.; Kasa-Vubu, J.Z.; Lerario, A.M.; Auchus, G.C.; Auchus, R.J.; Smith, J.M.; White, P.C.; Rainey, W.E. 11-Ketotestosterone Is the Dominant Circulating Bioactive Androgen during Normal and Premature Adrenarche. J. Clin. Endocrinol. Metab. 2018, 103, 4589–4598. [Google Scholar] [CrossRef] [Green Version]
- Bélanger, B.; Fiet, J.; Bé’elanger, A. Effects of adrenocorticotropin on adrenal and plasma 11β-hydroxyandrostenedione in the guinea pig and determination of its relative androgen potency. Steroids 1993, 58, 29–34. [Google Scholar] [CrossRef]
- Quinn, T.A.; Ratnayake, U.; Dickinson, H.; Castillo-Melendez, M.; Walker, D.W. Ontogenetic Change in the Regional Distribution of Dehydroepiandrosterone-Synthesizing Enzyme and the Glucocorticoid Receptor in the Brain of the Spiny Mouse (Acomys cahirinus). Dev. Neurosci. 2016, 38, 54–73. [Google Scholar] [CrossRef]
- Cutler, G.B., Jr.; Glenn, M.; Bush, M.; Hodgen, G.D.; Graham, C.E.; Loriaux, D.L. Adrenarche: A Survey of Rodents, Domestic Animals, and Primates. Endocrinology 1978, 103, 2112–2118. [Google Scholar] [CrossRef] [Green Version]
- Guillemette, C.; Hum, D.W.; Belanger, A. Levels of plasma C19 steroids and 5 alpha-reduced C19 steroid glucuronides in primates, rodents, and domestic animals. Am. J. Physiol.-Endocrinol. Metab. 1996, 271, E348–E353. [Google Scholar] [CrossRef] [PubMed]
- Quinn, T.A.; Ratnayake, U.; Dickinson, H.; Nguyen, T.H.; McIntosh, M.; Castillo-Melendez, M.; Conley, A.J.; Walker, D.W. Ontogeny of the adrenal gland in the spiny mouse, with particular reference to production of the steroids cortisol and dehydroepiandrosterone. Endocrinology 2013, 154, 1190–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auchus, R.J. Overview of dehydroepiandrosterone biosynthesis. Semin. Reprod. Med. 2004, 22, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Grube, M.; Hagen, P.; Jedlitschky, G. Neurosteroid transport in the brain: Role of ABC and SLC transporters. Front. Pharmacol. 2018, 9, 354. [Google Scholar] [CrossRef] [PubMed]
- Maninger, N.; Wolkowitz, O.M.; Reus, V.I.; Epel, E.S.; Mellon, S.H. Neurobiological and neuropsychiatric effects of dehydroepiandrosterone (DHEA) and DHEA sulfate (DHEAS). Front. Neuroendocrinol. 2009, 30, 65–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compagnone, N.A.; Mellon, S.H. Dehydroepiandrosterone: A potential signalling molecule for neocortical organization during development. Proc. Natl. Acad. Sci. USA 1998, 95, 4678–4683. [Google Scholar] [CrossRef] [Green Version]
- Hajszan, T.; Milner, T.A.; Leranth, C. Sex steroids and the dentate gyrus. Prog. Brain Res. 2007, 163, 399–816. [Google Scholar]
- Hajszan, T.; MacLusky, N.J.; Leranth, C. Dehydroepiandrosterone increases hippocampal spine synapse density in ovariectomized female rats. Endocrinology 2004, 145, 1042–1045. [Google Scholar] [CrossRef] [Green Version]
- Majewska, M.D. Neuronal actions of dehydroepiandrosterone possible roles in brain development, aging, memory, and affect. Ann. N. Y. Acad. Sci. 1995, 774, 111–120. [Google Scholar] [CrossRef]
- Starka, L.; Duskova, M.; Hill, M. Dehydroepiandrosterone: A neuroactive steroid. J. Steroid Biochem. Mol. Biol. 2015, 145, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Cardounel, A.; Gursoy, E.; Anderson, P.; Kalimi, M. Anti-stress effects of dehydroepiandrosterone: Protection of rats against repeated immobilization stress-induced weight loss, glucocorticoid receptor production, and lipid peroxidation. Biochem. Pharmacol. 2000, 59, 753–762. [Google Scholar] [CrossRef]
- Kimonides, V.G.; Spillantini, M.G.; Sofroniew, M.V.; Fawcett, J.W.; Herbert, J. Dehydroepiandrosterone antagonizes the neurotoxic effects of corticosterone and translocation of stress-activated protein kinase 3 in hippocampal primary cultures. Neuroscience 1999, 89, 429–436. [Google Scholar] [CrossRef]
- Bastianetto, S.; Ramassamy, C.; Poirier, J.; Quirion, R. Dehydroepiandrosterone (DHEA) protects hippocampal cells from oxidative stress-induced damage. Brain Res. Mol. Brain Res. 1999, 66, 35–41. [Google Scholar] [CrossRef]
- Cardounel, A.; Regelson, W.; Kalimi, M. Dehydroepiandrosterone protects hippocampal neurons against neurotoxin-induced cell death: Mechanism of action. Proc. Soc. Exp. Biol. Med. 1999, 222, 145–149. [Google Scholar] [CrossRef]
- Kurata, K.; Takebayashi, M.; Morinobu, S.; Yamawaki, S. beta-estradiol, dehydroepiandrosterone, and dehydroepiandrosterone sulfate protect against N-methyl-D-aspartate-induced neurotoxicity in rat hippocampal neurons by different mechanisms. J. Pharmacol. Exp. Ther. 2004, 311, 237–245. [Google Scholar] [CrossRef]
- Maurice, T.; Grégoire, C.; Espallergues, J. Neuro (active) steroids actions at the neuromodulatory sigma1 (σ1) receptor: Biochemical and physiological evidences, consequences in neuroprotection. Pharmacol. Biochem. Behav. 2006, 84, 581–597. [Google Scholar] [CrossRef]
- Kaasik, A.; Safiulina, D.; Kalda, A.; Zharkovsky, A. Dehydroepiandrosterone with other neurosteroids preserve neuronal mitochondria from calcium overload. J. Steroid Biochem. Mol. Biol. 2003, 87, 97–103. [Google Scholar] [CrossRef]
- Schverer, M.; Lanfumey, L.; Baulieu, E.-E.; Froger, N.; Villey, I. Neurosteroids: Non-genomic pathways in neuroplasticity and involvement in neurological diseases. Pharmacol. Ther. 2018, 191, 190–206. [Google Scholar] [CrossRef]
- Aldred, S.; Mecocci, P. Decreased dehydroepiandrosterone (DHEA) and dehydroepiandrosterone sulfate (DHEAS) concentrations in plasma of Alzheimer’s disease (AD) patients. Arch. Gerontol. Geriatr. 2010, 51, e16–e18. [Google Scholar] [CrossRef]
- Rasmuson, S.; Näsman, B.; Olsson, T. Increased serum levels of dehydroepiandrosterone (DHEA) and interleukin-6 (IL-6) in women with mild to moderate Alzheimer’s disease. Int. Psychogeriatr. 2011, 23, 1386–1392. [Google Scholar] [CrossRef] [PubMed]
- Yanase, T.; Fukahori, M.; Taniguchi, S.; Nishi, Y.; Sakai, Y.; Takayanagi, R.; Haji, M.; Nawata, H. Serum dehydroepiandrosterone (DHEA) and DHEA-sulfate (DHEA-S) in Alzheimer’s disease and in cerebrovascular dementia. Endocr. J. 1996, 43, 119–123. [Google Scholar] [CrossRef] [Green Version]
- Tourney, G.; Hatfield, L. Plasma androgens in male schizophrenics. Arch. Gen. Psychiatry 1972, 27, 753–755. [Google Scholar] [CrossRef]
- Harris, D.S.; Wolkowitz, O.M.; Reus, V.I. Movement disorder, memory, psychiatric symptoms and serum DHEA levels in schizophrenic and schizoaffective patients. World J. Biol. Psychiatry 2001, 2, 99–102. [Google Scholar] [CrossRef]
- Ritsner, M.; Maayan, R.; Gibel, A.; Weizman, A. Differences in blood pregnenolone and dehydroepiandrosterone levels between schizophrenia patients and healthy subjects. Eur. Neuropsychopharmacol. 2007, 17, 358–365. [Google Scholar] [CrossRef]
- Bremner, D.; Vermetten, E.; Kelley, M.E. Cortisol, dehydroepiandrosterone, and estradiol measured over 24 hours in women with childhood sexual abuse-related posttraumatic stress disorder. J. Nerv. Mental Dis. 2007, 195, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, R.; Brand, S.R.; Golier, J.A.; Yang, R.-K. Clinical correlates of DHEA associated with post-traumatic stress disorder. Acta Psychiatr. Scand. 2006, 114, 187–193. [Google Scholar] [CrossRef]
- Spivak, B.; Maayan, R.; Kotler, M.; Mester, R.; Gil-Ad, I.; Shtaif, B.; Weizman, A. Elevated circulatory level of GABA A–antagonistic neurosteroids in patients with combat-related post-traumatic stress disorder. Psychol. Med. 2000, 30, 1227–1231. [Google Scholar] [CrossRef]
- Gill, J.; Vythilingam, M.; Page, G.G. Low cortisol, high DHEA, and high levels of stimulated TNF-α, and IL-6 in women with PTSD. J. Trauma. Stress 2008, 21, 530–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabian, T.J.; Dew, M.A.; Pollock, B.G.; Reynolds III, C.F.; Mulsant, B.H.; Butters, M.A.; Zmuda, M.D.; Linares, A.M.; Trottini, M.; Kroboth, P.D. Endogenous concentrations of DHEA and DHEA-S decrease with remission of depression in older adults. Biol. Psychiatry 2001, 50, 767–774. [Google Scholar] [CrossRef]
- Hough, C.M.; Lindqvist, D.; Epel, E.S.; Denis, M.S.; Reus, V.I.; Bersani, F.S.; Rosser, R.; Mahan, L.; Burke, H.M.; Wolkowitz, O.M.; et al. Higher serum DHEA concentrations before and after SSRI treatment are associated with remission of major depression. Psychoneuroendocrinology 2017, 77, 122–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voutilainen, R.; Jääskeläinen, J. Premature adrenarche: Etiology, clinical findings, and consequences. J. Steroid Biochem. Mol. Biol. 2015, 145, 226–236. [Google Scholar] [CrossRef]
- Dorn, L.D.; Rose, S.R.; Rotenstein, D.; Susman, E.J.; Huang, B.; Loucks, T.L.; Berga, S.L. Differences in endocrine parameters and psychopathology in girls with premature adrenarche versus on-time adrenarche. J. Pediatr. Endocrinol. Metab. JPEM 2008, 21, 439–448. [Google Scholar] [CrossRef] [Green Version]
- Marakaki, C.; Pervanidou, P.; Papassotiriou, I.; Mastorakos, G.; Hochberg, Z.e.; Chrousos, G.; Papadimitriou, A. Increased symptoms of anxiety and depression in prepubertal girls, but not boys, with premature adrenarche: Associations with serum DHEAS and daily salivary cortisol concentrations. Stress 2018, 21, 564–568. [Google Scholar] [CrossRef]
- Sontag-Padilla, L.M.; Dorn, L.D.; Tissot, A.; Susman, E.J.; Beers, S.R.; Rose, S.R. Executive functioning, cortisol reactivity, and symptoms of psychopathology in girls with premature adrenarche. Dev. Psychopathol. 2012, 24, 211–223. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez, L.; DiMartino-Nardi, J.; Potau, N.; Saenger, P. Premature Adrenarche—Normal Variant or Forerunner of Adult Disease? Endocr. Rev. 2000, 21, 671–696. [Google Scholar] [CrossRef]
- Maliqueo, M.; Sir-Petermann, T.; Pérez, V.; Echiburú, B.; Ladrón de Guevara, A.; Gálvez, C.; Crisosto, N.; Azziz, R. Adrenal Function during Childhood and Puberty in Daughters of Women with Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2009, 94, 3282–3288. [Google Scholar] [CrossRef] [Green Version]
- Chaudhari, A.P.; Mazumdar, K.; Mehta, P.D. Anxiety, Depression, and Quality of Life in Women with Polycystic Ovarian Syndrome. Indian J. Psychol. Med. 2018, 40, 239–246. [Google Scholar] [CrossRef]
- Himelein, M.J.; Thatcher, S.S. Polycystic Ovary Syndrome and Mental Health: A Review. Obstet. Gynecol. Surv. 2006, 61, 723–732. [Google Scholar] [CrossRef]
- Kerchner, A.; Lester, W.; Stuart, S.P.; Dokras, A. Risk of depression and other mental health disorders in women with polycystic ovary syndrome: A longitudinal study. Fertil. Steril. 2009, 91, 207–212. [Google Scholar] [CrossRef]
- Çelik, N.; Alp, H.; Çamtosun, E.; Alp, E.; Çelik, S.; Berk, E. The Association between Premature Adrenarche and Cardiovascular Risk May Be Greater than Expected. Horm. Res. Paediatr. 2017, 87, 7–14. [Google Scholar] [CrossRef]
- Utriainen, P.; Laakso, S.; Liimatta, J.; Jääskeläinen, J.; Voutilainen, R. Premature Adrenarche—A Common Condition with Variable Presentation. Horm. Res. Paediatr. 2015, 83, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.N.; Barker, D.J. Type 2 (non-insulin-dependent) diabetes mellitus: The thrifty phenotype hypothesis. Diabetologia 1992, 35, 595–601. [Google Scholar] [CrossRef]
- O’Donnell, K.J.; Meaney, M.J. Fetal Origins of Mental Health: The Developmental Origins of Health and Disease Hypothesis. Am. J. Psychiatry 2017, 174, 319–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawn, J.E.; Cousens, S.; Zupan, J. 4 million neonatal deaths: When? Where? Why? Lancet 2005, 365, 891–900. [Google Scholar] [CrossRef]
- Kramer, M.S. Determinants of low birth weight: Methodological assessment and meta-analysis. Bull. World Health Organ. 1987, 65, 663–737. [Google Scholar]
- Gagnon, R.; Johnston, L.; Murotsuki, J. Fetal placental embolization in the late-gestation ovine fetus: Alterations in umbilical blood flow and fetal heart rate patterns. Am. J. Obstet. Gynecol. 1996, 175, 63–72. [Google Scholar] [CrossRef]
- Go, K.S.; Lingas, R.; Wheeler, M.B.; Irwin, D.M.; Matthews, S.G. Decreased CRH mRNA expression in the fetal guinea pig hypothalamus following maternal nutrient restriction. Brain Res. 2001, 896, 179–182. [Google Scholar] [CrossRef]
- Iwata, S.; Kinoshita, M.; Okamura, H.; Tsuda, K.; Saikusa, M.; Harada, E.; Saitoh, S.; Iwata, O. Intrauterine growth and the maturation process of adrenal function. PeerJ 2019, 7, e6368. [Google Scholar] [CrossRef]
- Lingas, R.; Dean, F.; Matthews, S.G. Maternal nutrient restriction (48 h) modifies brain corticosteroid receptor expression and endocrine function in the fetal guinea pig. Brain Res. 1999, 846, 236–242. [Google Scholar] [CrossRef]
- Schmelter, V.; Antonaci, L.; Fröhlich, S.; Herebian, D.; Verde, P.; Heil, M.; Hoehn, T. Quantification of fetal steroids in nails of neonates to quantify prenatal stress and growth restriction. Biol. Psychol. 2019, 140, 81–85. [Google Scholar] [CrossRef]
- Turnipseed, M.R.; Bentley, K.; Reynolds, J.W. Serum Dehydroepiandrosterone Sulfate in Premature Infants and Infants with Intrauterine Growth Retardation. J. Clin. Endocrinol. Metab. 1976, 43, 1219–1225. [Google Scholar] [CrossRef] [PubMed]
- Farzad Mohajeri, Z.; Aalipour, S.; Sheikh, M.; Shafaat, M.; Hantoushzadeh, S.; Borna, S.; Khazardoost, S. Ultrasound measurement of fetal adrenal gland in fetuses with intrauterine growth restriction, an early predictive method for adverse outcomes. J. Matern.-Fetal Neonatal Med. 2019, 32, 1485–1491. [Google Scholar] [CrossRef]
- Poudel, R.; McMillen, I.C.; Dunn, S.L.; Zhang, S.; Morrison, J.L. Impact of chronic hypoxemia on blood flow to the brain, heart, and adrenal gland in the late-gestation IUGR sheep fetus. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2015, 308, R151–R162. [Google Scholar] [CrossRef] [Green Version]
- Simonetta, G.; Rourke, A.K.; Owens, J.A.; Robinson, J.S.; McMillen, I.C. Impact of Placental Restriction on the Development of the Sympathoadrenal System. Pediatr. Res. 1997, 42, 805–811. [Google Scholar] [CrossRef] [Green Version]
- Parker, C.R.; Buchina, E.S.; Barefoot, T.K. Abnormal Adrenal Steroidogenesis in Growth-Retarded Newborn Infants. Pediatr. Res. 1994, 35, 633–636. [Google Scholar] [CrossRef] [Green Version]
- Edwards, A.V.; Jones, C.T. Autonomic control of adrenal function. J. Anat. 1993, 183, 291–307. [Google Scholar]
- Bornstein, S.R.; Ehrhart-Bornstein, M.; Usadel, H.; Böckmann, M.; Scherbaum, W.A. Morphological evidence for a close interaction of chromaffin cells with cortical cells within the adrenal gland. Cell Tissue Res. 1991, 265, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.T.; Edwards, A.V. Release of adrenocorticotrophin from the adrenal gland in the conscious calf. J. Physiol. 1990, 426, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Ehrhart-Bornstein, M.; Hinson, J.P.; Bornstein, S.R.; Scherbaum, W.A.; Vinson, G.P. Intraadrenal Interactions in the Regulation of Adrenocortical Steroidogenesis. Endocr. Rev. 1998, 19, 101–143. [Google Scholar] [CrossRef]
- Lynch, P.J.; Jaffe, C.C. Multimedia Teaching Projects; Section, B.H.S., Ed.; Center for Advanced Instructional Media, Yale University School of Medicine, Yale University: New Haven, CT, USA, 2000. [Google Scholar]
- Kalimi, M.; Shafagoj, Y.; Loria, R.; Padgett, D.; Regelson, W. Anti-glucocorticoid effects of dehydroepiandrosterone (DHEA). Mol. Cell. Biochem. 1994, 131, 99–104. [Google Scholar] [CrossRef]
- Pryce, C.R. Postnatal ontogeny of expression of the corticosteroid receptor genes in mammalian brains: Inter-species and intra-species differences. Brain Res. Rev. 2008, 57, 596–605. [Google Scholar] [CrossRef]
- Sanchez, M.M.; Young, L.J.; Plotsky, P.M.; Insel, T.R. Distribution of corticosteroid receptors in the rhesus brain: Relative absence of glucocorticoid receptors in the hippocampal formation. J. Neurosci. 2000, 20, 4657–4668. [Google Scholar] [CrossRef] [Green Version]
- Galuska, C.E.; Hartmann, M.F.; Sanchez-Guijo, A.; Bakhaus, K.; Geyer, J.; Schuler, G.; Zimmer, K.P.; Wudy, S.A. Profiling intact steroid sulfates and unconjugated steroids in biological fluids by liquid chromatography-tandem mass spectrometry (LC-MS-MS). Analyst 2013, 138, 3792–3801. [Google Scholar] [CrossRef]
- Greaves, R.F.; Jolly, L.; Hartmann, M.F.; Ho, C.S.; Kam, R.K.; Joseph, J.; Boyder, C.; Wudy, S.A. Harmonisation of serum dihydrotestosterone analysis: Establishment of an external quality assurance program. Clin. Chem. Lab. Med. 2017, 55, 522–529. [Google Scholar] [CrossRef]
- Sanchez-Guijo, A.; Oji, V.; Hartmann, M.F.; Traupe, H.; Wudy, S.A. Simultaneous quantification of cholesterol sulfate, androgen sulfates, and progestagen sulfates in human serum by LC-MS/MS. J. Lipid Res. 2015, 56, 1843–1851. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cumberland, A.L.; Hirst, J.J.; Badoer, E.; Wudy, S.A.; Greaves, R.F.; Zacharin, M.; Walker, D.W. The Enigma of the Adrenarche: Identifying the Early Life Mechanisms and Possible Role in Postnatal Brain Development. Int. J. Mol. Sci. 2021, 22, 4296. https://doi.org/10.3390/ijms22094296
Cumberland AL, Hirst JJ, Badoer E, Wudy SA, Greaves RF, Zacharin M, Walker DW. The Enigma of the Adrenarche: Identifying the Early Life Mechanisms and Possible Role in Postnatal Brain Development. International Journal of Molecular Sciences. 2021; 22(9):4296. https://doi.org/10.3390/ijms22094296
Chicago/Turabian StyleCumberland, Angela L., Jonathan J. Hirst, Emilio Badoer, Stefan A. Wudy, Ronda F. Greaves, Margaret Zacharin, and David W. Walker. 2021. "The Enigma of the Adrenarche: Identifying the Early Life Mechanisms and Possible Role in Postnatal Brain Development" International Journal of Molecular Sciences 22, no. 9: 4296. https://doi.org/10.3390/ijms22094296