
Genome-Wide Identification and Analysis of CC-NBS-LRR Family in Response to Downy Mildew and Black Rot in Chinese Cabbage

 and
and
Abstract
:1. Introduction
2. Results
2.1. Identifcation of the CC–NBS–LRR Family in Brassica rapa
2.2. Location and Distribution of CC–NBS–LRR Family Genes on Brassica rapa Chromosome
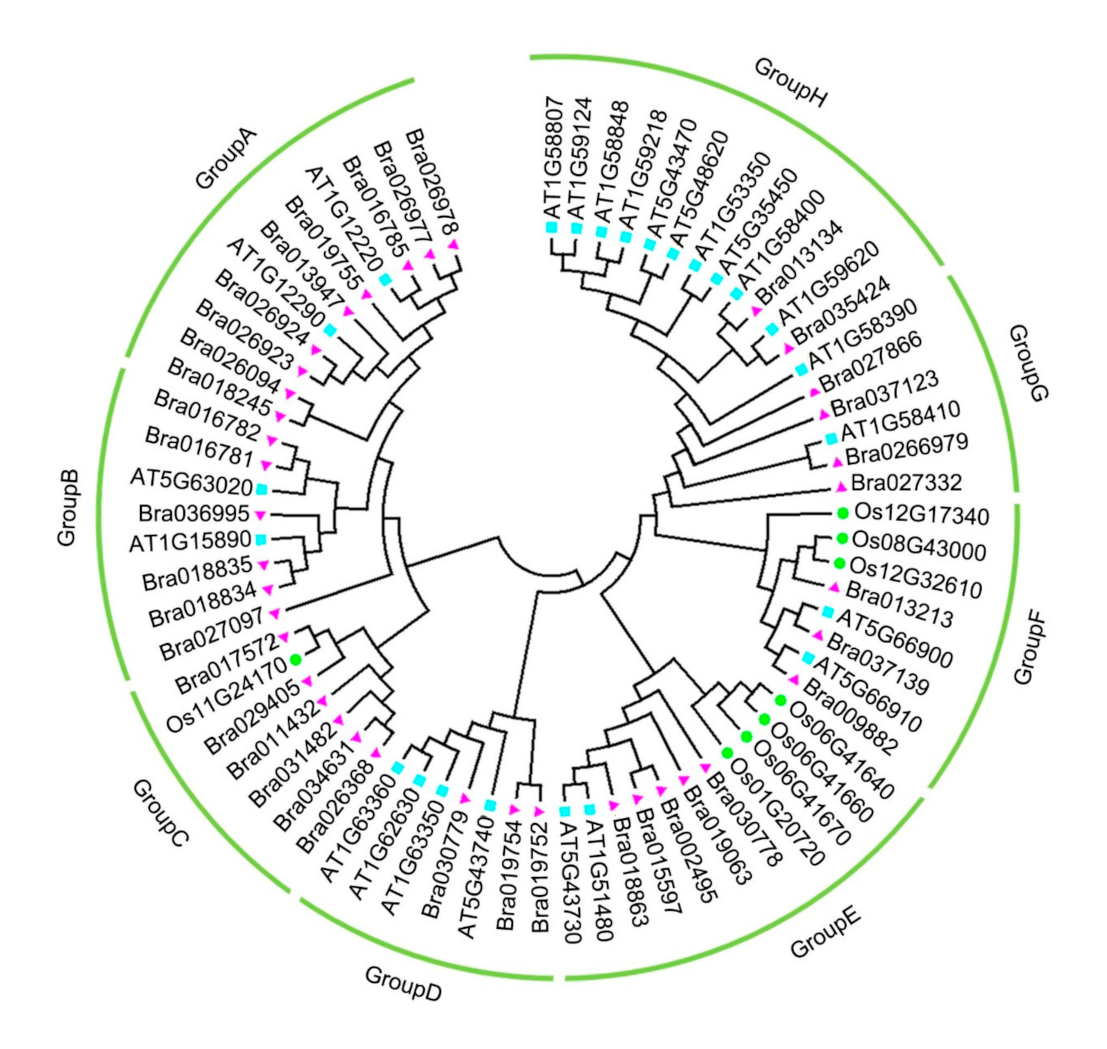
2.3. Phylogenetic Analysis of the CC–NBS–LRR Family in Brassica rapa, Arabidopsis thaliana, and Oryza sativa
2.4. Gene Structures and Protein Function Domains of CC–NBS–LRR Family in Chinese Cabbage
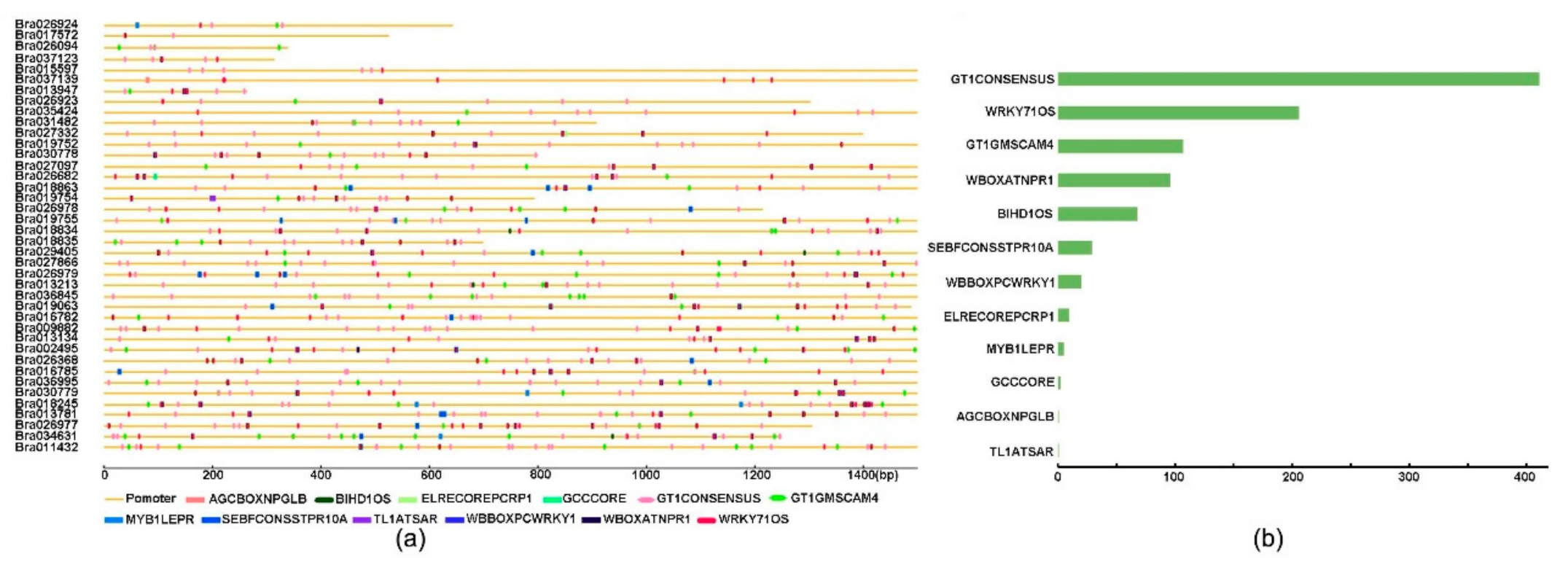
2.5. Cis-Acting Elements Analysis of the CC–NBS–LRR Family Promoter in Brassica rapa
2.6. Tissue Expression Patterns of CC-NBS-LRR Genes in Chinese Cabbage
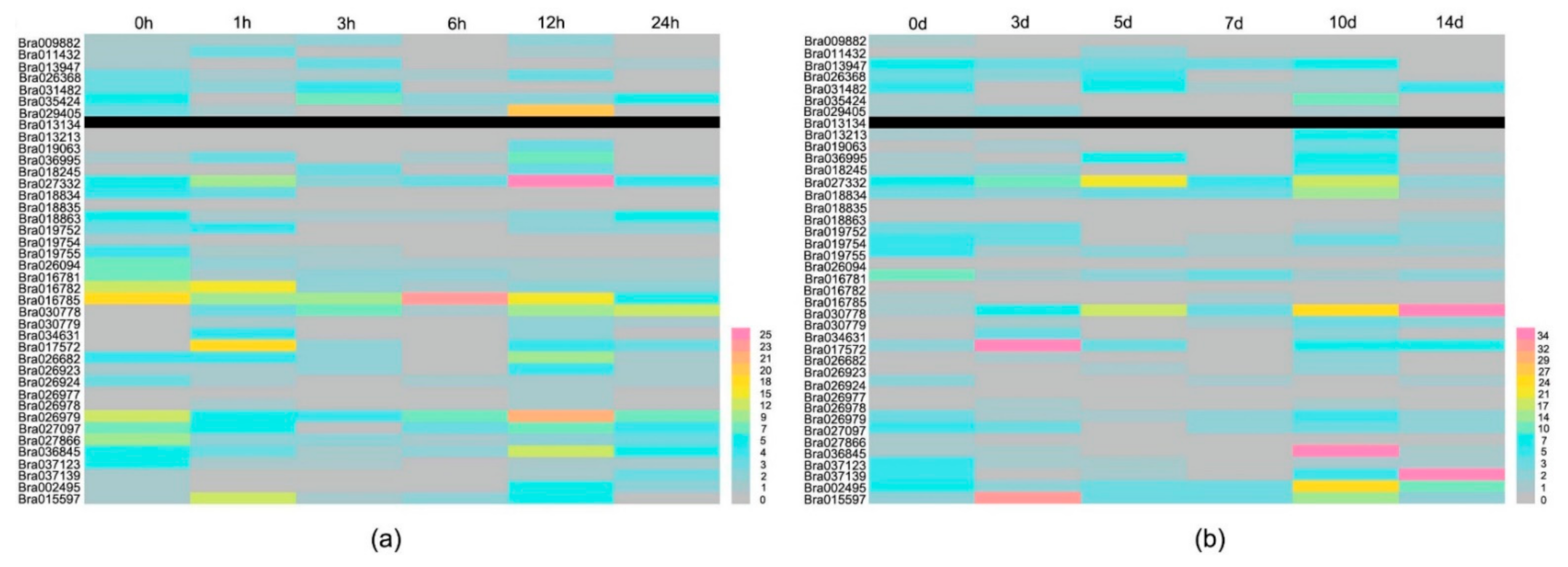
2.7. Expression Profiles of BrCC–NBS–LRR Genes in Response to Phytopathogens
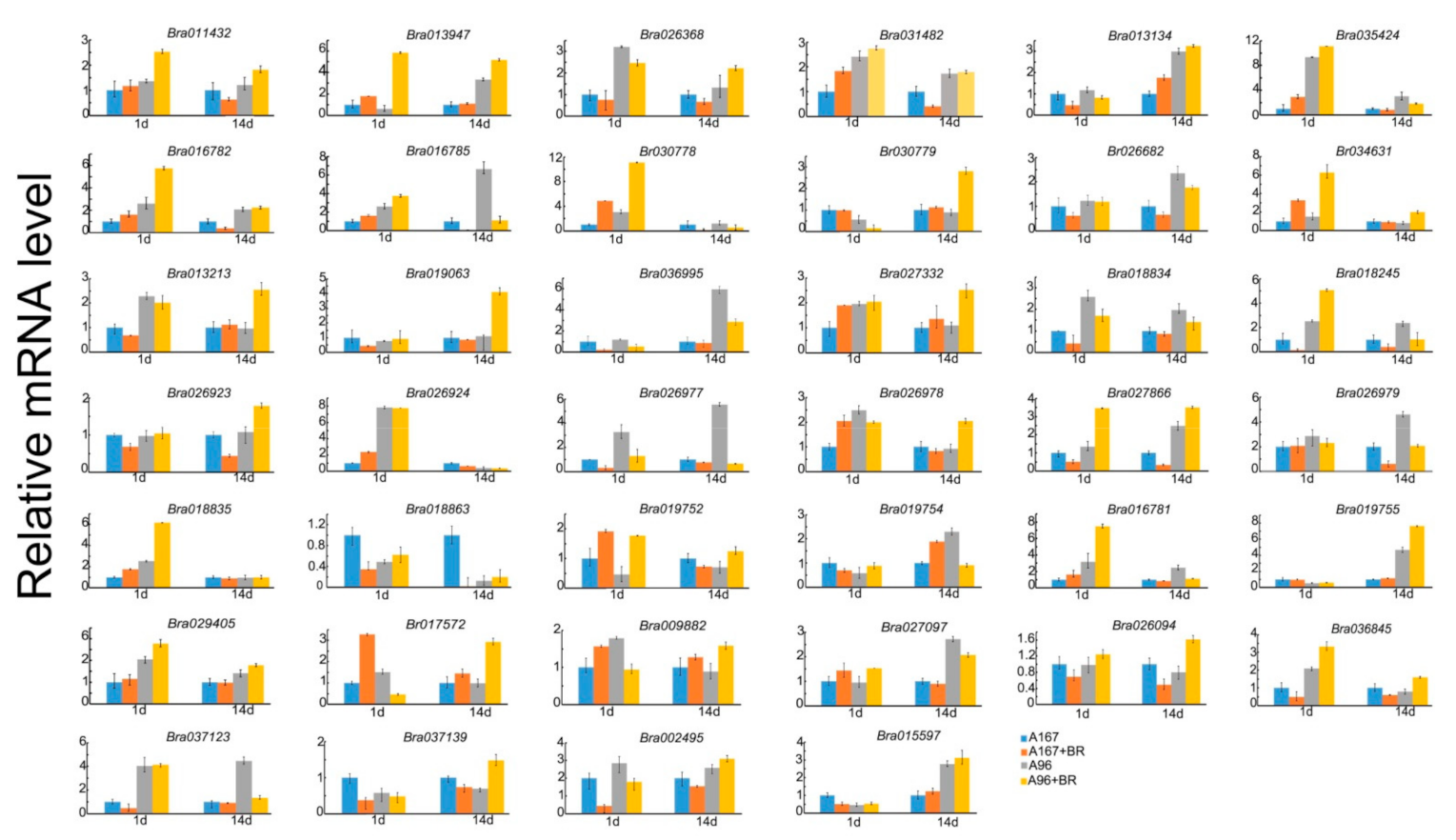
2.8. Associated Expression of BrCC-NBS-LRR in Different Inbred Lines of Chinese Cabbage in Response to Phytopathogen
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Inoculation
4.2. Identification and Analysis of CC–NBS–LRR in Chinese Cabbage
4.3. The Cis-Acting Elements in Promoters, Chromosomal Locations, and Gene Structures
4.4. Phylogenetics and Conserved Motif Analysis of CC–NBS–LRR Protein
4.5. Total RNA Extraction and Real-Time PCR Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
| BR | black rot |
| CC–NBS–LRR | coil-coil nucleotide-binding site leucine-rich repeat |
| DM | Downy mildew |
| ETI | Effector triggered immunity |
| NB-ARC | nucleotide-binding adaptor shared by APAF-1, R proteins and CED-4 |
| PI | isoelectric point |
| SA | salicin |
| TIR | toll/interleukin 1 receptor. |
References
- Ausubel, F.M. Are innate immune signaling pathways in plants and animals conserved? Nat. Immunol. 2005, 6, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Haak, D.C.; Fukao, T.; Grene, R.; Hua, Z.H.; Ivanov, R.; Perrella, G.; Li, S. Multilevel Regulation of Abiotic Stress Responses in Plants. Front. Plant Sci. 2017, 8, 1564. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.M.; Chitrakar, R.; Obulareddy, N.; Panchal, S.; Williams, P.; Melotto, M. Molecular battles between plant and pathogenic bacteria in the phyllosphere. Braz. J. Med. Biol. Res. 2010, 43, 698–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belkhadir, Y.; Subramaniam, R.; Dangl, J.L. Plant disease resistance protein signaling: NBS-LRR proteins and their partners. Curr. Opin. Plant Biol. 2004, 7, 391–399. [Google Scholar] [CrossRef]
- Tameling, W.I.; Elzinga, S.D.; Darmin, P.S.; Vossen, J.H.; Takken, F.L.; Haring, M.A.; Cornelissen, B.J. The tomato R gene products I-2 and MI-1 are functional ATP binding proteins with ATPase activity. Plant Cell 2002, 14, 2929–2939. [Google Scholar] [CrossRef]
- Matsushima, N.; Miyashita, H. Leucine-Rich Repeat (LRR) Domains Containing Intervening Motifs in Plants. Biomolecules 2012, 2, 288–311. [Google Scholar] [CrossRef] [Green Version]
- Wan, H.; Yuan, W.; Ye, Q.; Wang, R.; Ruan, M.; Li, Z.; Zhou, G.; Yao, Z.; Zhao, J.; Liu, S.; et al. Analysis of TIR- and non-TIR-NBS-LRR disease resistance gene analogous in pepper: Characterization, genetic variation, functional divergence and expression patterns. BMC Genom. 2012, 13, 502. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.; Tummala, N.R.; Aziz, S.G.; Risko, C.; Bredas, J.L. Influence of Molecular Shape on Solid-State Packing in Disordered PC61BM and PC71BM Fullerenes. J. Phys. Chem. Lett. 2014, 5, 3427–3433. [Google Scholar] [CrossRef]
- Tarr, D.E.; Alexander, H.M. TIR-NBS-LRR genes are rare in monocots: Evidence from diverse monocot orders. BMC Res. Notes 2009, 2, 197. [Google Scholar] [CrossRef] [Green Version]
- Sukarta, O.C.A.; Slootweg, E.J.; Goverse, A. Structure-informed insights for NLR functioning in plant immunity. Semin. Cell Dev. Biol. 2016, 56, 134–149. [Google Scholar] [CrossRef]
- Song, W.; Wang, B.; Li, X.; Wei, J.; Chen, L.; Zhang, D.; Zhang, W.; Li, R. Identification of Immune Related LRR-Containing Genes in Maize (Zea mays L.) by Genome-Wide Sequence Analysis. Int. J. Genom. 2015, 2015, 231358. [Google Scholar]
- Jia, Y.; McAdams, S.A.; Bryan, G.T.; Hershey, H.P.; Valent, B. Direct interaction of resistance gene and avirulence gene products confers rice blast resistance. EMBO J. 2000, 19, 4004–4014. [Google Scholar] [CrossRef]
- Deslandes, L.; Olivier, J.; Peeters, N.; Feng, D.X.; Khounlotham, M.; Boucher, C.; Somssich, I.; Genin, S.; Marco, Y. Physical interaction between RRS1-R, a protein conferring resistance to bacterial wilt, and PopP2, a type III effector targeted to the plant nucleus. Proc. Natl. Acad. Sci. USA 2003, 100, 8024–8029. [Google Scholar] [CrossRef] [Green Version]
- DeYoung, B.J.; Innes, R.W. Plant NBS-LRR proteins in pathogen sensing and host defense. Nat. Immunol. 2006, 7, 1243–1249. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Upadhyaya, C.P.; Nookaraju, A.; Pandey, S.K.; Park, S.W. Plant disease resistance genes: Current status and future directions. Physiol. Mol. Plant P 2012, 78, 51–65. [Google Scholar] [CrossRef]
- Hu, Y.; Li, Y.; Hou, F.; Wan, D.; Cheng, Y.; Han, Y.; Gao, Y.; Liu, J.; Guo, Y.; Xiao, S.; et al. Ectopic expression of Arabidopsis broad-spectrum resistance gene RPW8.2 improves the resistance to powdery mildew in grapevine (Vitis vinifera). Plant Sci. 2018, 267, 20–31. [Google Scholar] [CrossRef]
- Liu, Y.; Guan, X.Y.; Liu, S.N.; Yang, M.; Ren, J.H.; Guo, M.; Huang, Z.H.; Zhang, Y.W. Genome-Wide Identification and Analysis of TCP Transcription Factors Involved in the Formation of Leafy Head in Chinese cabbage. Int. J. Mol. Sci. 2018, 19, 847. [Google Scholar] [CrossRef] [Green Version]
- Hashiba, T.; Narisawa, K. The development and endophytic nature of the fungus Heteroconium chaetospira. Fems Microbiol Lett 2005, 252, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Zeilmaker, T.; Ludwig, N.R.; Elberse, J.; Seidl, M.F.; Berke, L.; Van Doorn, A.; Schuurink, R.C.; Snel, B.; Van den Ackerveken, G. DOWNY MILDEW RESISTANT 6 and DMR6-LIKE OXYGENASE 1 are partially redundant but distinct suppressors of immunity in Arabidopsis. Plant J. 2015, 81, 210–222. [Google Scholar] [CrossRef]
- Zhang, B.; Li, P.; Su, T.B.; Li, P.R.; Xin, X.Y.; Wang, W.H.; Zhao, X.Y.; Yu, Y.J.; Zhang, D.S.; Yu, S.C.; et al. BrRLP48, Encoding a Receptor-Like Protein, Involved in Downy Mildew Resistance in Brassica rapa. Front. Plant Sci. 2018, 9, 1708. [Google Scholar] [CrossRef]
- Mun, J.H.; Yu, H.J.; Park, S.; Park, B.S. Genome-wide identification of NBS-encoding resistance genes in Brassica rapa. Mol. Genet. Genom. 2009, 282, 617–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, S.W.; Changwei, Z.; Tang, J.; Li, Y.X.; Wang, Z.; Jiang, D.H.; Hou, X.L. Genome-wide analysis and identification of TIR-NBS-LRR genes in Chinese cabbage (Brassica rapa ssp pekinensis) reveal expression patterns to TuMV infection. Physiol Mol Plant P 2015, 90, 89–97. [Google Scholar] [CrossRef]
- Buchel, A.S.; Brederode, F.T.; Bol, J.F.; Linthorst, H.J. Mutation of GT-1 binding sites in the Pr-1A promoter influences the level of inducible gene expression in vivo. Plant Mol. Biol. 1999, 40, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Zhang, Z.L.; Zou, X.; Huang, J.; Ruas, P.; Thompson, D.; Shen, Q.J. Annotations and functional analyses of the rice WRKY gene superfamily reveal positive and negative regulators of abscisic acid signaling in aleurone cells. Plant Physiol. 2005, 137, 176–189. [Google Scholar] [CrossRef] [Green Version]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Luo, H.; Song, F.; Goodman, R.M.; Zheng, Z. Up-regulation of OsBIHD1, a rice gene encoding BELL homeodomain transcriptional factor, in disease resistance responses. Plant Biol. 2005, 7, 459–468. [Google Scholar] [CrossRef]
- Chakravarthy, S.; Tuori, R.P.; D’Ascenzo, M.D.; Fobert, P.R.; Despres, C.; Martin, G.B. The tomato transcription factor Pti4 regulates defense-related gene expression via GCC box and non-GCC box cis elements. Plant Cell 2003, 15, 3033–3050. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.P.; Wang, J.P. Genome-Wide Analysis of NBS-LRR Genes in Sorghum Genome Revealed Several Events Contributing to NBS-LRR Gene Evolution in Grass Species. Evol. Bioinform. 2016, 12, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Meyers, B.C.; Kozik, A.; Griego, A.; Kuang, H.H.; Michelmore, R.W. Genome-wide analysis of NBS-LRR-encoding genes in Arabidopsis. Plant Cell 2003, 15, 809–834. [Google Scholar] [CrossRef] [Green Version]
- Shao, Z.Q.; Zhang, Y.M.; Hang, Y.Y.; Xue, J.Y.; Zhou, G.C.; Wu, P.; Wu, X.Y.; Wu, X.Z.; Wang, Q.; Wang, B.; et al. Long-term evolution of nucleotide-binding site-leucine-rich repeat genes: Understanding gained from and beyond the legume family. Plant Physiol. 2014, 166, 217–234. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.Y.; Tehrim, S.; Zhang, F.Q.; Tong, C.B.; Huang, J.Y.; Cheng, X.H.; Dong, C.H.; Zhou, Y.Q.; Qin, R.; Hua, W.; et al. Genome-wide comparative analysis of NBS-encoding genes between Brassica species and Arabidopsis thaliana. BMC Genomics 2014, 15, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Sekhwal, M.K.; Li, P.C.; Lam, I.; Wang, X.E.; Cloutier, S.; You, F.M. Disease Resistance Gene Analogs (RGAs) in Plants. Int. J. Mol. Sci. 2015, 16, 19248–19290. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Forderer, A.; Yu, D.; Chai, J. Structural biology of plant defence. New Phytol. 2021, 229, 692–711. [Google Scholar] [CrossRef]
- Mackey, D.; Holt, B.F., 3rd; Wiig, A.; Dangl, J.L. RIN4 interacts with Pseudomonas syringae type III effector molecules and is required for RPM1-mediated resistance in Arabidopsis. Cell 2002, 108, 743–754. [Google Scholar] [CrossRef] [Green Version]
- Ade, J.; DeYoung, B.J.; Golstein, C.; Innes, R.W. Indirect activation of a plant nucleotide binding site-leucine-rich repeat protein by a bacterial protease. Proc. Natl. Acad. Sci. USA 2007, 104, 2531–2536. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhu, J.F.; Wang, L.F.; Wang, S.M. Genome-Wide Association Study Identifies NBS-LRR-Encoding Genes Related with Anthracnose and Common Bacterial Blight in the Common Bean. Front. Plant Sci. 2017, 8, 1398. [Google Scholar] [CrossRef] [Green Version]
- Rushton, P.J.; Reinstadler, A.; Lipka, V.; Lippok, B.; Somssich, I.E. Synthetic plant promoters containing defined regulatory elements provide novel insights into pathogen- and wound-induced signaling. Plant Cell 2002, 14, 749–762. [Google Scholar] [CrossRef] [Green Version]
- Gilmartin, P.M.; Chua, N.H. Spacing between GT-1 binding sites within a light-responsive element is critical for transcriptional activity. Plant Cell 1990, 2, 447–455. [Google Scholar]
- Villain, P.; Mache, R.; Zhou, D.X. The mechanism of GT element-mediated cell type-specific transcriptional control. J. Biol. Chem. 1996, 271, 32593–32598. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.L.; Xie, Z.; Zou, X.L.; Casaretto, J.; Ho, T.H.D.; Shen, Q.X.J. A rice WRKY gene encodes a transcriptional repressor of the gibberellin signaling pathway in aleurone cells. Plant Physiol. 2004, 134, 1500–1513. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Chen, C.; Chen, Z. Evidence for an important role of WRKY DNA binding proteins in the regulation of NPR1 gene expression. Plant Cell 2001, 13, 1527–1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rushton, P.J.; Torres, J.T.; Parniske, M.; Wernert, P.; Hahlbrock, K.; Somssich, I.E. Interaction of elicitor-induced DNA-binding proteins with elicitor response elements in the promoters of parsley PR1 genes. EMBO J. 1996, 15, 5690–5700. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Shao, Z.Q.; Wang, Q.; Hang, Y.Y.; Xue, J.Y.; Wang, B.; Chen, J.Q. Uncovering the dynamic evolution of nucleotide-binding site-leucine-rich repeat (NBS-LRR) genes in Brassicaceae. J. Integr. Plant Biol. 2016, 58, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Dangl, J.L.; Horvath, D.M.; Staskawicz, B.J. Pivoting the plant immune system from dissection to deployment. Science 2013, 341, 746–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loutre, C.; Wicker, T.; Travella, S.; Galli, P.; Scofield, S.; Fahima, T.; Feuillet, C.; Keller, B. Two different CC-NBS-LRR genes are required for Lr10-mediated leaf rust resistance in tetraploid and hexaploid wheat. Plant J. 2009, 60, 1043–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Frick, M.; Huel, R.; Nykiforuk, C.L.; Wang, X.M.; Gaudet, D.A.; Eudes, F.; Conner, R.L.; Kuzyk, A.; Chen, Q.; et al. The Stripe Rust Resistance Gene Yr10 Encodes an Evolutionary-Conserved and Unique CC-NBS-LRR Sequence in Wheat. Mol. Plant 2014, 7, 1740–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.H.; Tian, W.; Tao, F.; Wang, J.J.; Shang, H.S.; Chen, X.M.; Xu, X.M.; Hu, X.P. TaRPM1 Positively Regulates Wheat High-Temperature Seedling-Plant Resistance to Puccinia striiformis f. sp. tritici. Front. Plant Sci. 2020, 10, 1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, L.; Hu, P.; Liu, J.; Witek, K.; Zhou, S.; Xu, J.; Zhou, W.; Gao, L.; Huang, Z.; Zhang, R.; et al. Pm21 from Haynaldia villosa Encodes a CC-NBS-LRR Protein Conferring Powdery Mildew Resistance in Wheat. Mol. Plant 2018, 11, 874–878. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, N.; Inoue, H.; Kato, T.; Funao, T.; Shirota, M.; Shimizu, T.; Kanamori, H.; Yamane, H.; Hayano-Saito, Y.; Matsumoto, T.; et al. Durable panicle blast-resistance gene Pb1 encodes an atypical CC-NBS-LRR protein and was generated by acquiring a promoter through local genome duplication. Plant J. Cell Mol. Biol. 2010, 64, 498–510. [Google Scholar] [CrossRef]
- Zimmermann, L.; Stephens, A.; Nam, S.Z.; Rau, D.; Kubler, J.; Lozajic, M.; Gabler, F.; Soding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. In Proceedings of the International Conference on Intelligent Systems for Molecular Biology, Stanford, CA, USA, 15–17 August 1994; Association for the Advancement of Artificial Intelligence: Menlo Park, CA, USA, 1994; Volume 2, pp. 28–36. [Google Scholar]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Zhang, Y.W.; Jin, D.; Xu, C.; Zhang, L.; Guo, M.H.; Fang, Z.Y. Regulation of bolting and identification of the alpha-tubulin gene family in Brassica rapa L. ssp pekinensis. Genet. Mol. Res. 2016, 15, 1–13. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Genome Position | CDS Length(bp) | Exon | Protein Length(aa) | Molecular Weight (kDa) | Isoelectric Point |
|---|---|---|---|---|---|---|
| Bra011432 | A01:2,191,374...2,196,276 | 2424 | 5 | 807 | 91.25906 | 5.96 |
| Bra013947 | A01:8,643,759...8,646,482 | 2724 | 1 | 907 | 103.78083 | 5.79 |
| Bra026368 | A01:9,720,079...9,723,012 | 2934 | 1 | 977 | 110.51354 | 6.56 |
| Bra031482 | A01:17,037,929...17,040,208 | 2145 | 2 | 714 | 81.85303 | 8.46 |
| Bra035424 | A01:16,404,803...16,407,820 | 2703 | 3 | 900 | 104.13992 | 6.56 |
| Bra029405 | A02:25352667...25357391 | 3489 | 4 | 1162 | 135.16142 | 6.22 |
| Bra013134 | A03:20,277,197...20,280,164 | 2736 | 3 | 911 | 104.87017 | 8.30 |
| Bra013213 | A03:19,840,894...19,844,167 | 2916 | 3 | 971 | 111.04147 | 7.81 |
| Bra019063 | A03:26,487,398...26,490,427 | 3030 | 1 | 1009 | 114.15502 | 6.30 |
| Bra036995 | A03:29,029,474...29,032,056 | 2583 | 1 | 860 | 97.34071 | 6.87 |
| Bra018245 | A05:6,909,119...6,911,701 | 2583 | 1 | 860 | 98.19411 | 6.50 |
| Bra027332 | A05:20,364,432...20,367,560 | 3129 | 1 | 1042 | 119.33079 | 8.42 |
| Bra009882 | A06:17,846,911...17,849,759 | 2466 | 5 | 821 | 93.52592 | 6.33 |
| Bra018834 | A06:1,698,860...1,701,415 | 2556 | 1 | 851 | 96.87193 | 5.85 |
| Bra018835 | A06:1,694,120...1,696,651 | 2532 | 1 | 843 | 95.76674 | 6.08 |
| Bra018863 | A06:1,554,378...1,556,936 | 2559 | 1 | 852 | 96.60849 | 5.99 |
| Bra019752 | A06:4,608,614...4,611,267 | 2583 | 2 | 860 | 98.23598 | 8.12 |
| Bra019754 | A06:4,599,425...4,602,097 | 2673 | 1 | 890 | 100.98419 | 6.63 |
| Bra019755 | A06:4,595,072...4,597,754 | 2586 | 2 | 861 | 98.09356 | 6.05 |
| Bra026094 | A06:6,170,240...6,176,552 | 4842 | 12 | 1613 | 182.73632 | 6.22 |
| Bra016781 | A08:19,944,556...19,947,218 | 2589 | 2 | 862 | 98.25993 | 5.64 |
| Bra016782 | A08:19,949,435...19,952,182 | 2748 | 1 | 915 | 104.28085 | 5.44 |
| Bra016785 | A08:19,969,925...19,972,603 | 2679 | 1 | 892 | 101.87473 | 6.10 |
| Bra030778 | A08:20,402,328...20,404,925 | 2598 | 1 | 865 | 98.73556 | 5.14 |
| Bra030779 | A08:20,398,567...20,401,178 | 2517 | 2 | 838 | 95.04727 | 5.82 |
| Bra034631 | A08:12,381,249...12,384,097 | 2655 | 3 | 884 | 99.63007 | 6.46 |
| Bra017572 | A09:16,599,373...16,602,112 | 2001 | 5 | 666 | 76.24774 | 5.48 |
| Bra026682 | A09:33,197,405...33,200,004 | 2514 | 2 | 837 | 95.01493 | 6.02 |
| Bra026923 | A09:34,338,460...34,341,119 | 2559 | 2 | 852 | 97.16891 | 6.09 |
| Bra026924 | A09:34,342,578...34,345,222 | 2544 | 2 | 847 | 96.53494 | 5.73 |
| Bra026977 | A09:34,580,053...34,582,713 | 2661 | 1 | 886 | 101.12124 | 5.72 |
| Bra026978 | A09:34,584,346...34,587,018 | 2673 | 1 | 890 | 101.55526 | 6.14 |
| Bra026979 | A09:34,589,539...34,592,283 | 2745 | 1 | 914 | 103.82301 | 6.74 |
| Bra027097 | A09:8,445,238...8,447,974 | 1992 | 3 | 663 | 75.57687 | 6.33 |
| Bra027866 | A09:9,368,958...9,371,944 | 2790 | 3 | 929 | 106.98517 | 6.51 |
| Bra036845 | A09:25,444,116...25,448,953 | 3876 | 5 | 1291 | 146.91249 | 6.22 |
| Bra037123 | A09:4,366,953...4,369,453 | 2415 | 2 | 804 | 93.69916 | 8.98 |
| Bra037139 | A09:4,297,244...4,300,267 | 2454 | 4 | 817 | 93.03288 | 5.42 |
| Bra002495 | A10:9,233,853...9,236,492 | 2640 | 1 | 879 | 99.47706 | 8.46 |
| Bra015597 | A10:747,361...750,005 | 2571 | 2 | 856 | 97.89404 | 7.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Li, D.; Yang, N.; Zhu, X.; Han, K.; Gu, R.; Bai, J.; Wang, A.; Zhang, Y. Genome-Wide Identification and Analysis of CC-NBS-LRR Family in Response to Downy Mildew and Black Rot in Chinese Cabbage. Int. J. Mol. Sci. 2021, 22, 4266. https://doi.org/10.3390/ijms22084266
Liu Y, Li D, Yang N, Zhu X, Han K, Gu R, Bai J, Wang A, Zhang Y. Genome-Wide Identification and Analysis of CC-NBS-LRR Family in Response to Downy Mildew and Black Rot in Chinese Cabbage. International Journal of Molecular Sciences. 2021; 22(8):4266. https://doi.org/10.3390/ijms22084266
Chicago/Turabian StyleLiu, Yan, Dalong Li, Na Yang, Xiaolong Zhu, Kexin Han, Ran Gu, Junyu Bai, Aoxue Wang, and Yaowei Zhang. 2021. "Genome-Wide Identification and Analysis of CC-NBS-LRR Family in Response to Downy Mildew and Black Rot in Chinese Cabbage" International Journal of Molecular Sciences 22, no. 8: 4266. https://doi.org/10.3390/ijms22084266