
Dose-Dependent Physiological and Transcriptomic Responses of Lettuce (Lactuca sativa L.) to Copper Oxide Nanoparticles—Insights into the Phytotoxicity Mechanisms


Abstract
:1. Introduction
2. Results and Discussion
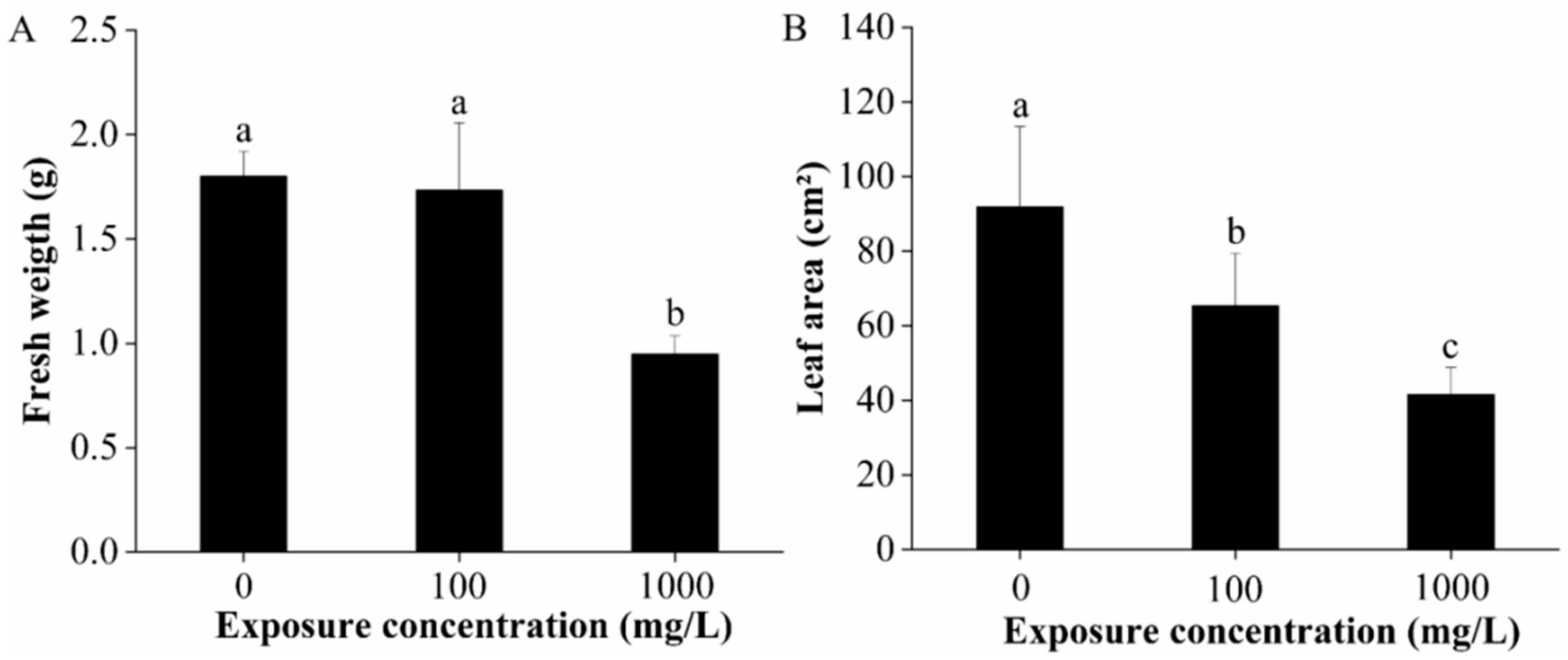
2.1. Biomass and Leaf Area
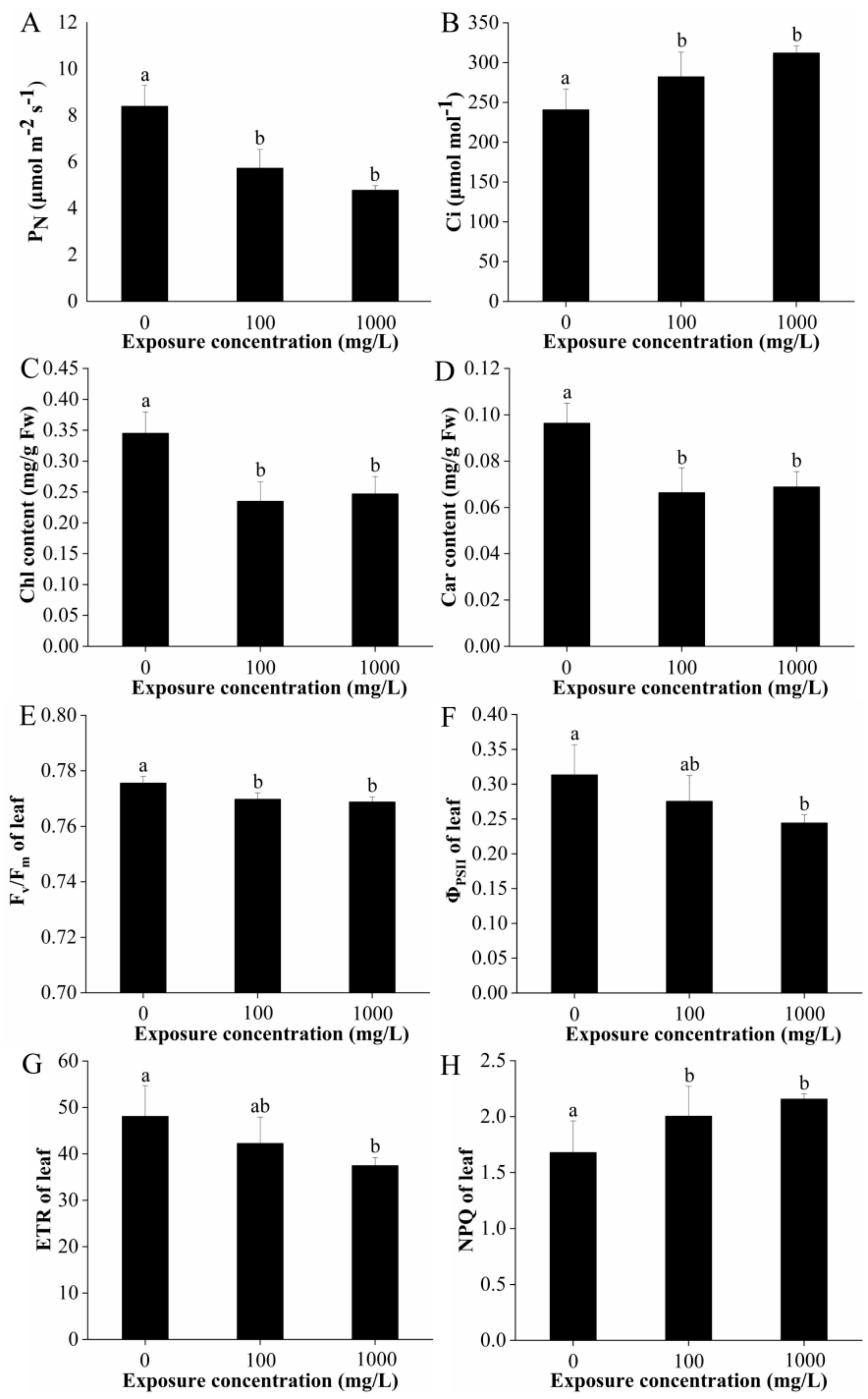
2.2. Gas Exchange, Photosynthetic Pigments Concentration, and Chlorophyll Fluorescence
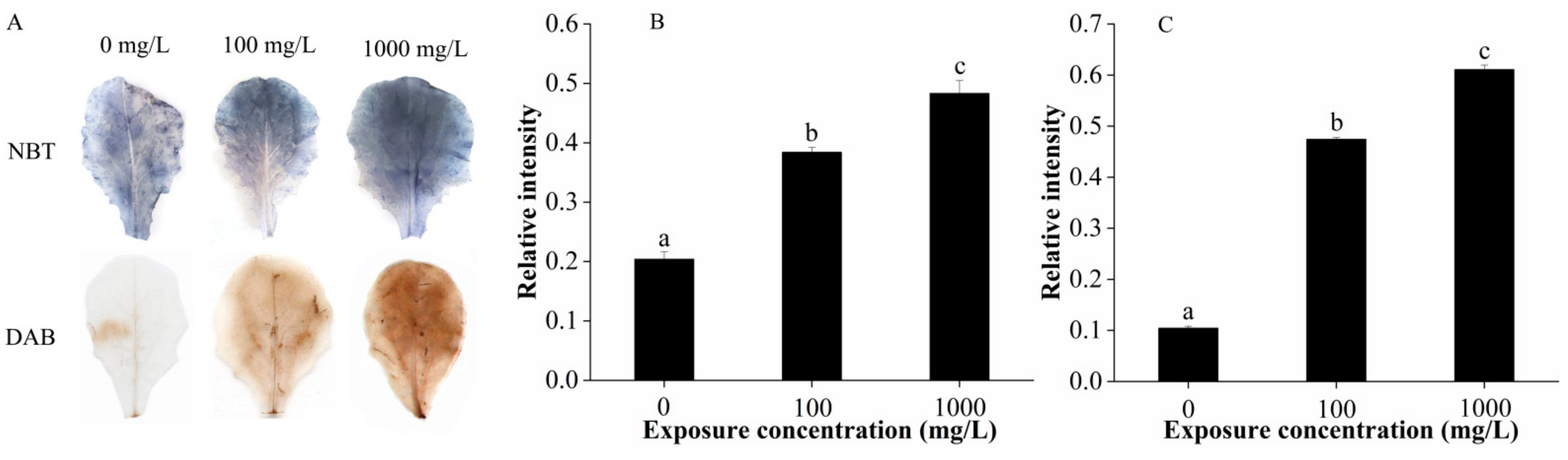
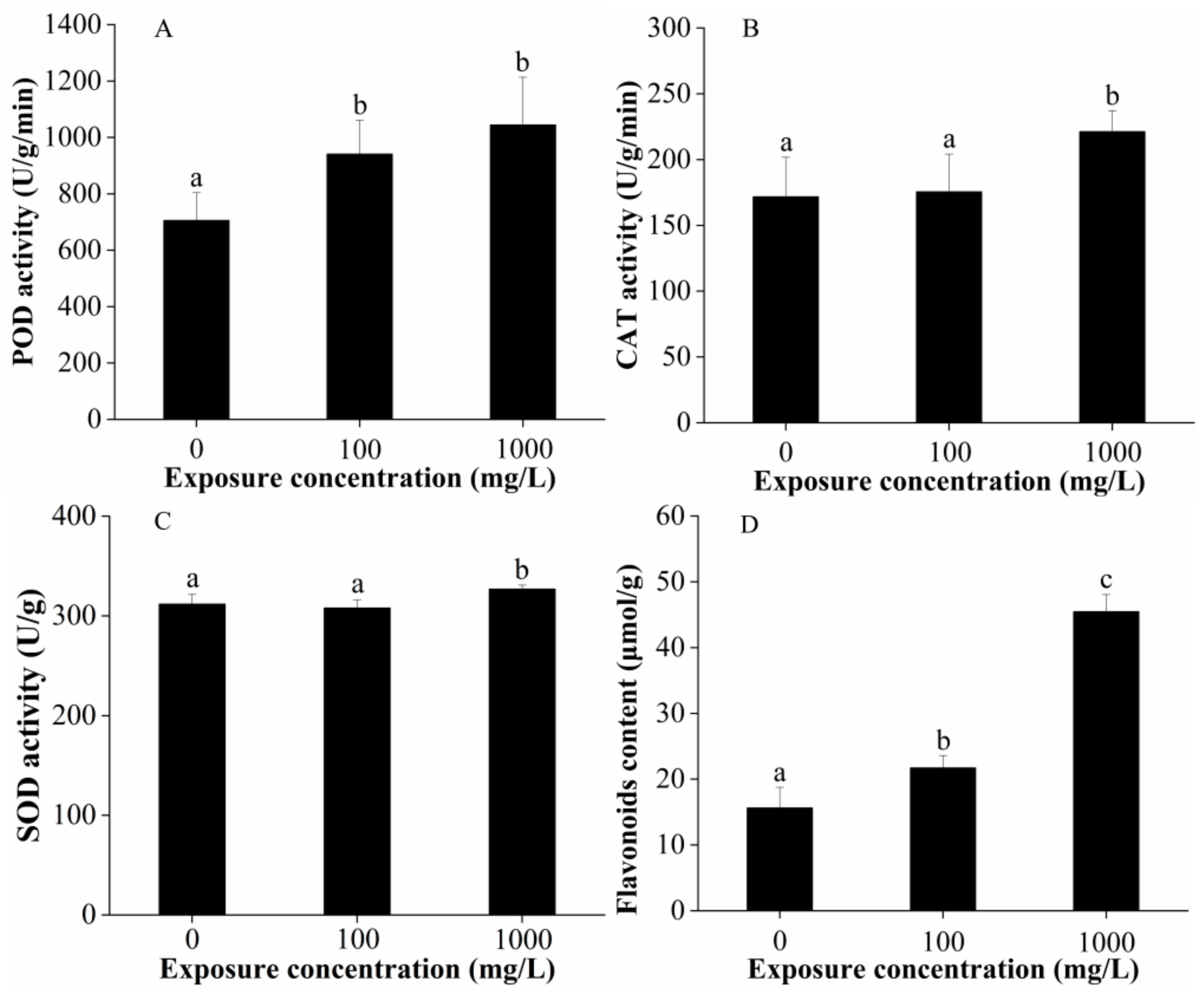
2.3. Reactive Oxygen Species (ROS) Determination and Antioxidant Activities in Plant Leaves
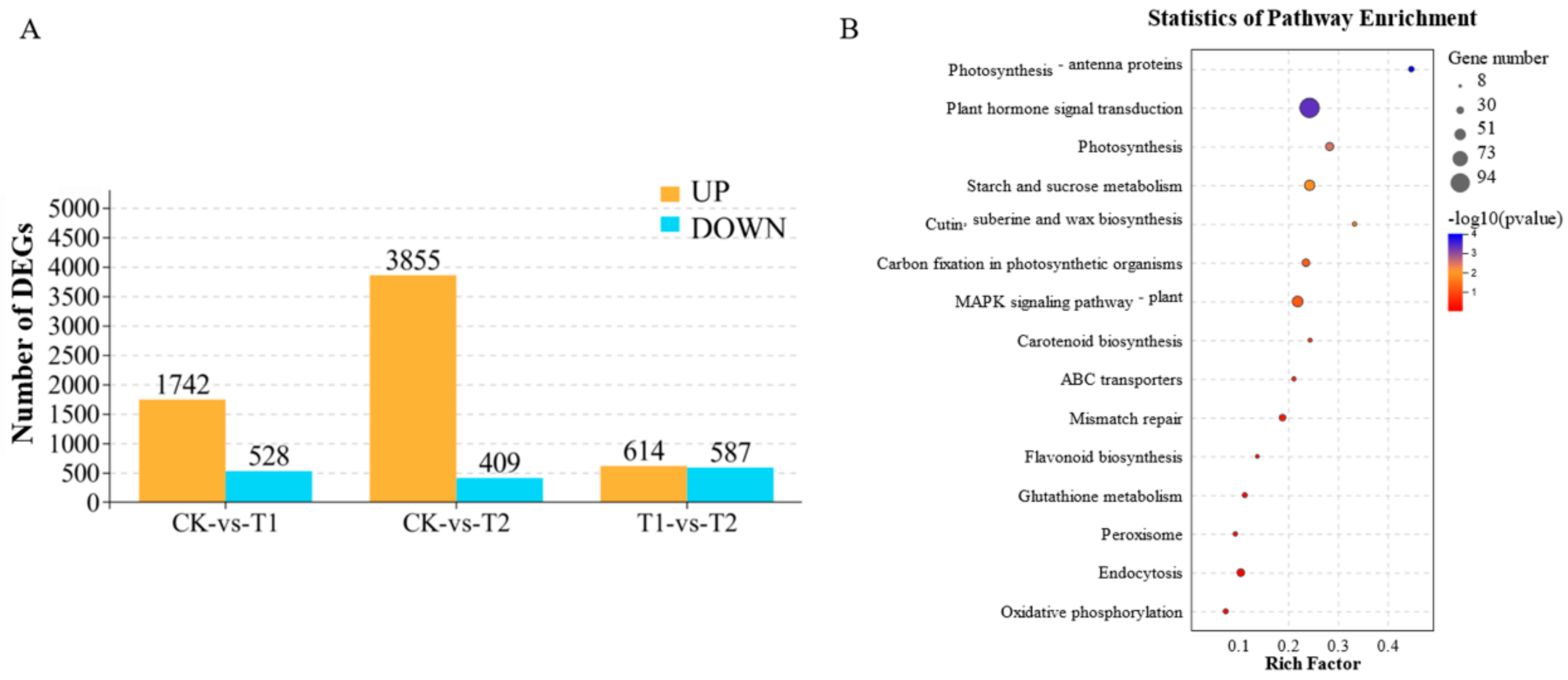
2.4. Overview of Transcriptome Sequencing Analysis
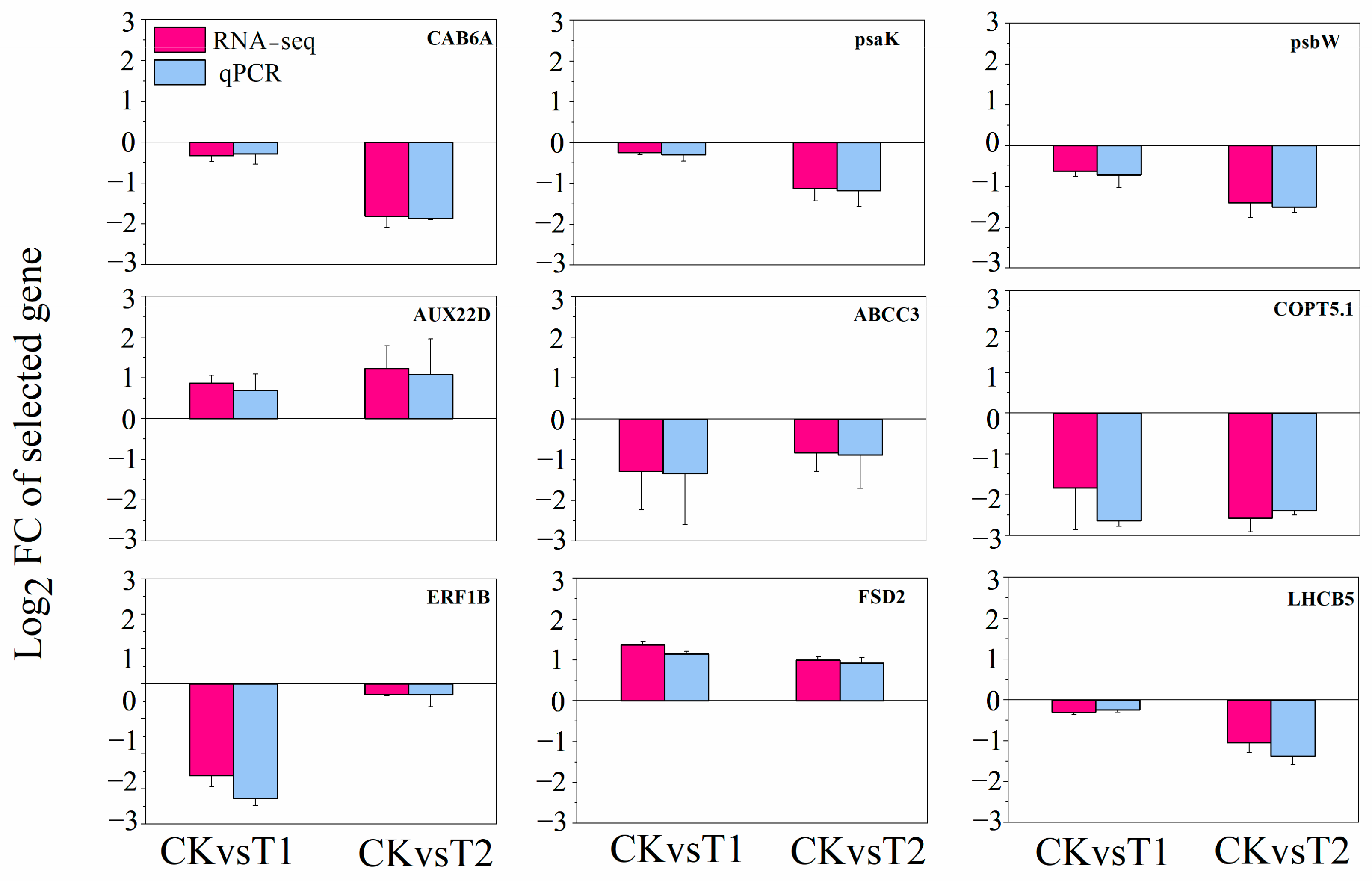
2.5. RNA Sequencing Validation by qPCR
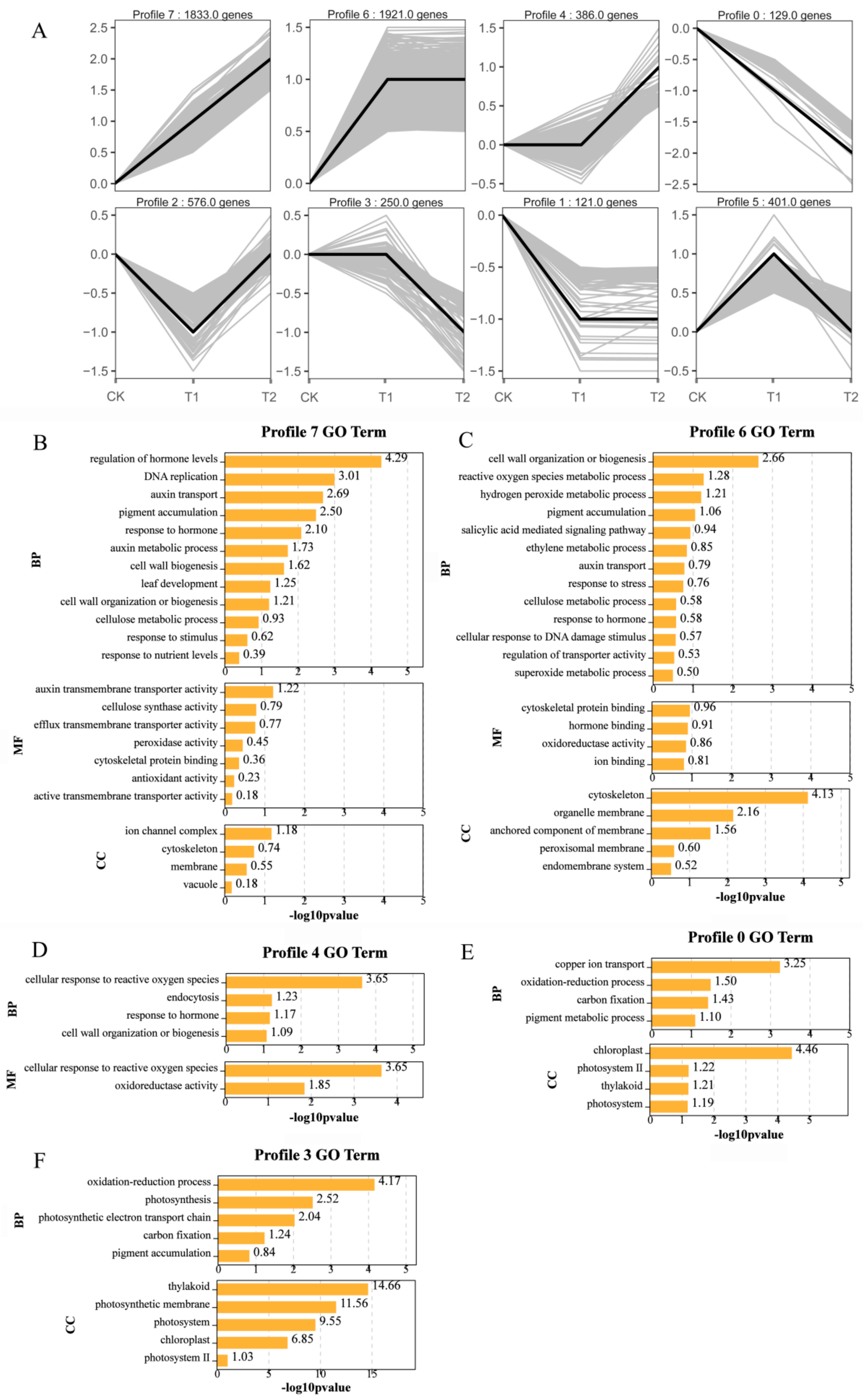
2.6. Gene Expression Pattern Analysis, Clustering, and Functional Enrichment of DEGs
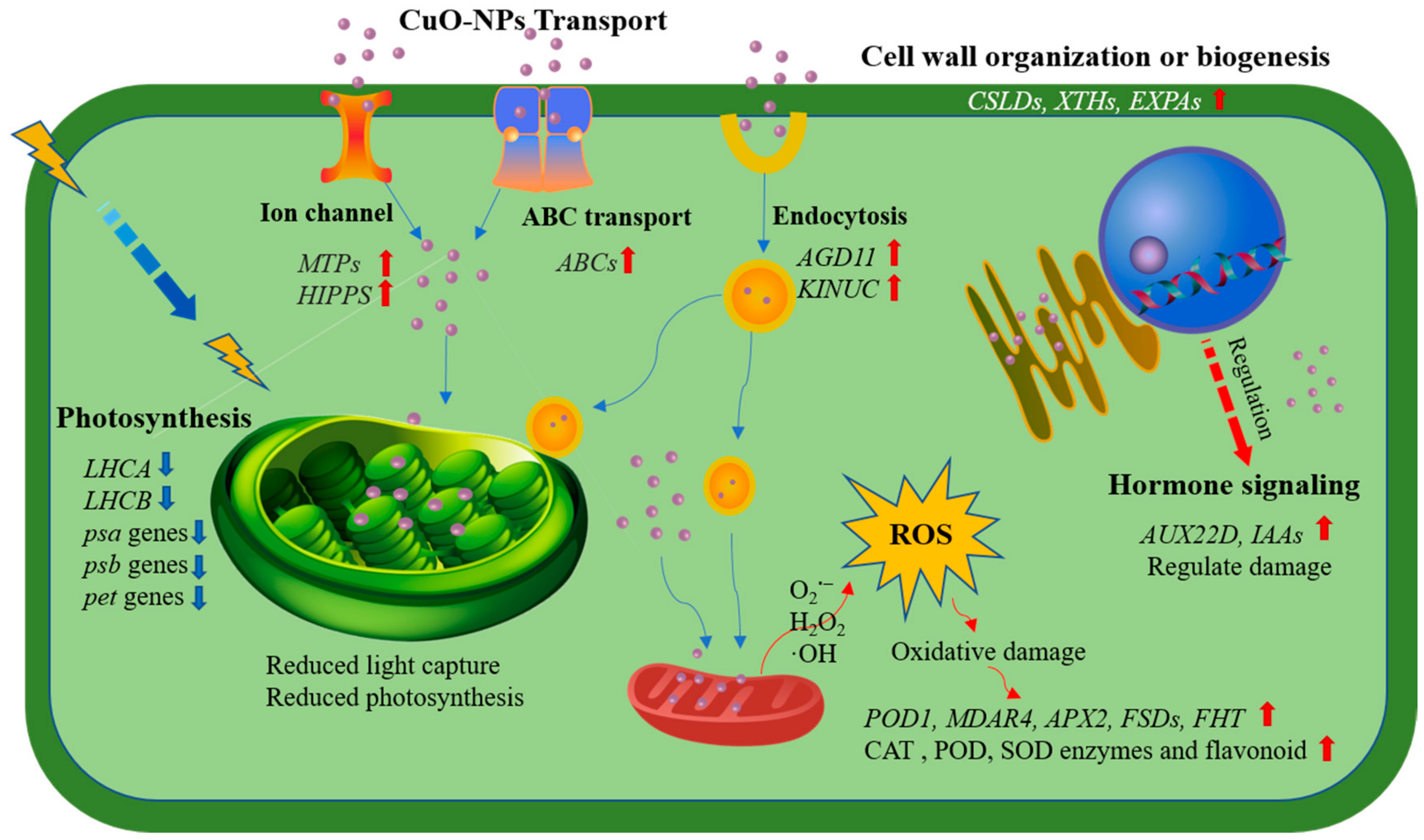
2.7. Significant DEGs
2.7.1. Changed Cell Wall Organization or Biogenesis
2.7.2. Decreased Photosynthesis
2.7.3. Altered Oxidation-Reduction Processes and Antioxidant Activity
2.7.4. CuO-NPs Transport
2.7.5. Hormone Signal Transduction
3. Materials and Methods
3.1. Exposure of Lettuce Leaves to CuO-NPs
3.2. Gas Exchange Parameters, Photosynthetic Pigments Concentration, and Chlorophyll Fluorescence Parameters
3.3. ROS Determination and Antioxidant Enzyme Activities in Plant Leaves
3.4. RNA Extraction, Library Construction, and Sequencing
3.5. Bioinformatics Analysis
3.6. Real-Time Quantitative PCR
3.7. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Z.; Ke, M.; Qu, Q.; Peijnenburg, W.J.G.M.; Lu, T.; Zhang, Q.; Ye, Y.; Xu, P.; Du, B.; Sun, L.; et al. Impact of copper nanoparticles and ionic copper exposure on wheat (Triticum aestivum L.) root morphology and antioxidant response. Environ. Pollut. 2018, 239, 689–697. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.; Sushkova, S.; Mandzhieva, S.; Fedorenko, A.; Lysenko, V.; Bederska-Błaszczyk, M.; Olchowik, J.; Tsitsuashvili, V.; Chaplygin, V. Structural and Ultrastructural Changes in Nanoparticle Exposed Plants. In Nanoscience for Sustainable Agriculture; Springer Nature Switzerland AG: Cham, Switzerland, 2019; pp. 281–295. ISBN 9783319978529. [Google Scholar]
- Rajput, V.D.; Minkina, T.; Suskova, S.; Mandzhieva, S.; Tsitsuashvili, V.; Chapligin, V.; Fedorenko, A. Effects of Copper Nanoparticles (CuO NPs) on Crop Plants: A Mini Review. Bionanoscience 2018, 8, 36–42. [Google Scholar] [CrossRef]
- Soares, C.; Pereira, R.; Fidalgo, F. Metal-based nanomaterials and oxidative stress in plants: Current aspects and overview. In Phytotoxicity of Nanoparticles; Springer: Cham, Switzerland, 2018; ISBN 9783319767086. [Google Scholar]
- Tripathi, D.K.; Shweta; Singh, S.; Singh, S.; Pandey, R.; Singh, V.P.; Sharma, N.C.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. An overview on manufactured nanoparticles in plants: Uptake, translocation, accumulation and phytotoxicity. Plant Physiol. Biochem. 2017, 110, 2–12. [Google Scholar] [CrossRef]
- Wu, J.; Wang, G.; Vijver, M.G.; Bosker, T.; Peijnenburg, W.J.G.M. Foliar versus root exposure of AgNPs to lettuce: Phytotoxicity, antioxidant responses and internal translocation. Environ. Pollut. 2020, 261, 114117. [Google Scholar] [CrossRef]
- Hong, J.; Peralta-Videa, J.R.; Rico, C.; Sahi, S.; Viveros, M.N.; Bartonjo, J.; Zhao, L.; Gardea-Torresdey, J.L. Evidence of translocation and physiological impacts of foliar applied CeO2 nanoparticles on cucumber (Cucumis sativus) plants. Environ. Sci. Technol. 2014, 48, 4376–4385. [Google Scholar] [CrossRef]
- Xiong, T.; Dumat, C.; Dappe, V.; Vezin, H.; Schreck, E.; Shahid, M.; Pierart, A.; Sobanska, S. Copper Oxide Nanoparticle Foliar Uptake, Phytotoxicity, and Consequences for Sustainable Urban Agriculture. Environ. Sci. Technol. 2017, 51, 5242–5251. [Google Scholar] [CrossRef]
- Gogos, A.; Knauer, K.; Bucheli, T.D. Nanomaterials in Plant Protection and Fertilization: Current State, Foreseen Applications, and Research Priorities Foreseen Applications, and Research Priorities. J. Agric. Food Chem. 2012, 60, 9781–9792. [Google Scholar] [CrossRef]
- Trujillo-Reyes, J.; Majumdar, S.; Botez, C.E.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Exposure studies of core-shell Fe/Fe3O4 and Cu/CuO NPs to lettuce (Lactuca sativa) plants: Are they a potential physiological and nutritional hazard? J. Hazard. Mater. 2014, 267, 255–263. [Google Scholar] [CrossRef]
- Zhao, L.; Ortiz, C.; Adeleye, A.S.; Hu, Q.; Zhou, H.; Huang, Y.; Keller, A.A. Metabolomics to Detect Response of Lettuce (Lactuca sativa) to Cu(OH)2 Nano-pesticides: Oxidative Stress Response and Detoxification Mechanisms. Environ. Sci. Technol. 2016, 50, 9697–9707. [Google Scholar] [CrossRef]
- Keller, A.A.; Huang, Y.; Nelson, J. Detection of nanoparticles in edible plant tissues exposed to nano-copper using sin-gle-particle ICP-MS. J. Nanoparticle Res. 2018, 20, 101. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Ali, S.; Rizwan, M.; Zia ur Rehman, M.; Javed, M.R.; Imran, M.; Chatha, S.A.S.; Nazir, R. Zinc oxide nanoparticles alter the wheat physiological response and reduce the cadmium uptake by plants. Environ. Pollut. 2018, 242, 1518–1526. [Google Scholar] [CrossRef]
- Rossi, L.; Fedenia, L.N.; Sharifan, H.; Ma, X.; Lombardini, L. Effects of foliar application of zinc sulfate and zinc nanoparticles in coffee (Coffea arabica L.) plants. Plant Physiol. Biochem. 2019, 135, 160–166. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Qayyum, M.F.; Ok, Y.S.; Adrees, M.; Ibrahim, M.; Zia-ur-Rehman, M.; Farid, M.; Abbas, F. Effect of metal and metal oxide nanoparticles on growth and physiology of globally important food crops: A critical review. J. Hazard. Mater. 2017, 322, 2–16. [Google Scholar] [CrossRef]
- Gkanatsiou, C.; Karamanoli, K.; Menkissoglu-Spiroudi, U.; Dendrinou-Samara, C. Composition effect of Cu-based nanoparticles on phytopathogenic bacteria. Antibacterial studies and phytotoxicity evaluation. Polyhedron 2019, 170, 395–403. [Google Scholar] [CrossRef]
- Adhikari, T.; Sarkar, D.; Mashayekhi, H.; Xing, B. Growth and enzymatic activity of maize (Zea mays L.) plant: Solution culture test for copper dioxide nano particles. J. Plant Nutr. 2016, 39, 99–115. [Google Scholar] [CrossRef]
- Hong, J.; Rico, C.M.; Zhao, L.; Adeleye, A.S.; Keller, A.A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Toxic effects of cop-per-based nanoparticles or compounds to lettuce (Lactuca sativa) and alfalfa (Medicago sativa). Environ. Sci. Process. Impacts 2015, 17, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Shaw, A.K.; Ghosh, S.; Kalaji, H.M.; Bosa, K.; Brestic, M.; Zivcak, M.; Hossain, Z. Nano-CuO stress induced modulation of antioxidative defense and photosynthetic performance of Syrian barley (Hordeum vulgare L.). Environ. Exp. Bot. 2014, 102, 37–47. [Google Scholar] [CrossRef]
- Shaw, A.K.; Hossain, Z. Impact of nano-CuO stress on rice (Oryza sativa L.) seedlings. Chemosphere 2013, 93, 906–915. [Google Scholar] [CrossRef]
- Wang, S.; Liu, H.; Zhang, Y.; Xin, H. The effect of CuO NPs on reactive oxygen species and cell cycle gene expression in roots of rice. Environ. Toxicol. Chem. 2015, 34, 554–561. [Google Scholar] [CrossRef]
- Wu, S.G.; Huang, L.; Head, J.; Ball, M.; Tang, Y.J.; Chen, D. Electrospray Facilitates the Germination of Plant Seeds Elec-trospray Facilitates the Germination of Plant Seeds. Aerosol Air Qual. Res. 2014, 14, 632–641. [Google Scholar] [CrossRef]
- Hong, J.; Wang, L.; Sun, Y.; Zhao, L.; Niu, G.; Tan, W.; Rico, C.M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Foliar applied nanoscale and microscale CeO2 and CuO alter cucumber (Cucumis sativus) fruit quality. Sci. Total Environ. 2015, 563–564, 904–911. [Google Scholar] [CrossRef] [Green Version]
- Atha, D.H.; Wang, H.; Petersen, E.J.; Cleveland, D.; Holbrook, R.D.; Jaruga, P.; Dizdaroglu, M.; Xing, B.; Nelson, B.C. Copper oxide nanoparticle mediated DNA damage in terrestrial plant models. Environ. Sci. Technol. 2012, 46, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, M.V.J.; Sharma, P.K. Effect of copper oxide nanoparticles on growth, morphology, photosynthesis, and antioxidant response in Oryza sativa. Photosynthetica 2016, 54, 110–119. [Google Scholar] [CrossRef]
- Zuverza-Mena, N.; Martínez-Fernández, D.; Du, W.; Hernandez-Viezcas, J.A.; Bonilla-Bird, N.; López-Moreno, M.L.; Komárek, M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Exposure of engineered nanomaterials to plants: Insights into the physiological and biochemical responses-A review. Plant Physiol. Biochem. 2016, 110, 236–264. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.L.; Jiang, H.S.; Gu, S.P.; Zhou, X.H.; Lu, Z.W.; Kang, X.H.; Yin, L.; Huang, J. Combination analysis of the physiology and transcriptome provides insights into the mechanism of silver nanoparticles phytotoxicity. Environ. Pollut. 2019, 252, 1539–1549. [Google Scholar] [CrossRef]
- Chen, Z.; Gao, S.; Jin, M.; Sun, S.; Lu, J.; Yang, P.; Bond, P.L.; Yuan, Z.; Guo, J. Physiological and transcriptomic analyses reveal CuO nanoparticle inhibition of anabolic and catabolic activities of sulfate-reducing bacterium. Environ. Int. 2019, 125, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gong, H.; Liao, W.; Wang, Z. Accumulation of particles on the surface of leaves during leaf expansion. Sci. Total Environ. 2015, 532, 420–434. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Zhao, J.; Yu, X.; Lv, K.; Wang, Z.; Xing, B. Interaction of CuO nanoparticles with duckweed (Lemna minor. L): Uptake, distribution and ROS production sites. Environ. Pollut. 2018, 243, 543–552. [Google Scholar] [CrossRef]
- Agathokleous, E.; Feng, Z.Z.; Peñuelas, J. Chlorophyll hormesis: Are chlorophylls major components of stress biology in higher plants? Sci. Total Environ. 2020, 726, 138637. [Google Scholar] [CrossRef]
- Nekrasova, G.F.; Ushakova, O.S.; Ermakov, A.E.; Uimin, M.A.; Byzov, I. V Effects of copper(II) ions and copper oxide nano-particles on Elodea densa Planch. Russ. J. Ecol. 2011, 42, 458–463. [Google Scholar] [CrossRef]
- Burzyński, M.; Kłobus, G. Changes of photosynthetic parameters in cucumber leaves under Cu, Cd, and Pb stress. Photosynthetica 2004, 42, 505–510. [Google Scholar] [CrossRef]
- Tighe-Neira, R.; Carmora, E.; Recio, G.; Nunes-Nesi, A.; Reyes-Diaz, M.; Alberdi, M.; Rengel, Z.; Inostroza-Blancheteau, C. Metallic nanoparticles influence the structure and function of the photosynthetic apparatus in plants. Plant Physiol. Biochem. 2018, 130, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Lalau, C.M.; Mohedano, R.D.A.; Schmidt, É.C.; Bouzon, Z.L.; Ouriques, L.C.; Dos Santos, R.W.; Da Costa, C.H.; Vicentini, D.S.; Matias, W.G. Toxicological effects of copper oxide nanoparticles on the growth rate, photosynthetic pigment content, and cell morphology of the duckweed Landoltia punctata. Protoplasma 2014, 252, 221–229. [Google Scholar] [CrossRef]
- Gopalakrishnan Nair, P.M.; Kim, S.H.; Chung, I.M. Copper oxide nanoparticle toxicity in mung bean (Vigna radiata L.) seedlings: Physiological and molecular level responses of in vitro grown plants. Acta Physiol. Plant. 2014, 36, 2947–2958. [Google Scholar] [CrossRef]
- El-Kassas, H.Y.; Okbah, M.A.E.A. Phytotoxic effects of seaweed mediated copper nanoparticles against the harmful alga: Lyngbya majuscula. J. Genet. Eng. Biotechnol. 2017, 15, 41–48. [Google Scholar] [CrossRef]
- Natasha; Shahid, M.; Farooq, A.B.U.; Rabbani, F.; Khalid, S.; Dumat, C. Risk assessment and biophysiochemical responses of spinach to foliar application of lead oxide nanoparticles: A multivariate analysis. Chemosphere 2020, 245, 125605. [Google Scholar] [CrossRef]
- Dappe, V.; Dumez, S.; Bernard, F.; Hanoune, B.; Cuny, D.; Dumat, C.; Sobanska, S. The role of epicuticular waxes on foliar metal transfer and phytotoxicity in edible vegetables: Case of Brassica oleracea species exposed to manufactured particles. Environ. Sci. Pollut. Res. 2019, 26, 20092–20106. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, N.B.; Hussain, I.; Singh, H. Effect of biologically synthesized copper oxide nanoparticles on metabolism and antioxidant activity to the crop plants Solanum lycopersicum and Brassica oleracea var. botrytis. J. Biotechnol. 2017, 262, 11–27. [Google Scholar] [CrossRef]
- Cassana, F.F.; Falqueto, A.R.; Braga, E.J.B.; Peters, J.A.; Bacarin, M.A. Chlorophyll a fluorescence of sweet potato plants cultivated in vitro and during ex vitro acclimatization. Brazil. J. Plant Physiol. 2010, 22, 167–170. [Google Scholar] [CrossRef] [Green Version]
- Perreault, F.; Oukarroum, A.; Pirastru, L.; Sirois, L.; Gerson Matias, W.; Popovic, R. Evaluation of Copper Oxide Nanoparticles Toxicity Using Chlorophyll a Fluorescence Imaging in Lemna gibba. J. Bot. 2010, 2010, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Redox sensing and signalling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiol. Plant. 2003, 119, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Fan, X.; Li, X.; Zhang, Z.; Sun, L.; Fu, Z.; Lavoie, M.; Pan, X.; Qian, H. Distinct physiological and molecular responses in Arabidopsis thaliana exposed to aluminum oxide nanoparticles and ionic aluminum. Environ. Pollut. 2017, 228, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Hernández, I.; Alegre, L.; Van Breusegem, F.; Munné-Bosch, S. How relevant are flavonoids as antioxidants in plants? Trends Plant Sci. 2009, 14, 125–132. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Soares, C.; Carvalho, M.E.A.; Azevedo, R.A.; Fidalgo, F. Plants facing oxidative challenges—A little help from the antioxidant networks. Environ. Exp. Bot. 2019, 161, 4–25. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Fryzova, R.; Pohanka, M.; Martinkova, P.; Cihlarova, H.; Brtnicky, M.; Hladky, J.; Kynicky, J. Oxidative Stress and Heavy Metals in Plants. Rev. Environ. Contam. Toxicol. 2017, 245, 129–156. [Google Scholar] [CrossRef]
- Bela, K.; Horváth, E.; Gallé, Á.; Szabados, L.; Tari, I.; Csiszár, J. Plant glutathione peroxidases: Emerging role of the antioxidant enzymes in plant development and stress responses. J. Plant Physiol. 2015, 176, 192–201. [Google Scholar] [CrossRef]
- Jonapá-Hernández, F.; Gutiérrez-Miceli, F.; Santos-Espinosa, A.; Ruíz-Lau, N.; Ruíz-Valdiviezo, V.; Valdez-Salas, B.; Gonzá-lez-Mendoza, D. Foliar application of green nanoparticles in Annona muricata L. plants and their effects in physiological and biochemical parameters. Biocatal. Agric. Biotechnol. 2020, 28, 101751. [Google Scholar] [CrossRef]
- Quiterio-Gutiérrez, T.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Hernández-Fuentes, A.D.; Sandoval-Rangel, A.; Be-navides-Mendoza, A.; Cabrera-De La Fuente, M.; Juárez-Maldonado, A. The application of selenium and copper nanoparticles modifies the biochemical responses of tomato plants under stress by Alternaria Solani. Int. J. Mol. Sci. 2019, 20, 1950. [Google Scholar] [CrossRef] [Green Version]
- Dimkpa, C.O.; Singh, U.; Bindraban, P.S.; Adisa, I.O.; Elmer, W.H.; Gardea-torresdey, J.L.; White, J.C. Addition-omission of zinc, copper, and boron nano and bulk oxide particles demonstrate element and size-specific response of soybean to mi-cronutrients exposure. Sci. Total Environ. 2019, 665, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Ogunkunle, C.O.; Jimoh, M.A.; Asogwa, N.T.; Viswanathan, K.; Vishwakarma, V.; Fatoba, P.O. Effects of manufactured nano-copper on copper uptake, bioaccumulation and enzyme activities in cowpea grown on soil substrate. Ecotoxicol. Environ. Saf. 2018, 155, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Landa, P.; Vankova, R.; Andrlova, J.; Hodek, J.; Marsik, P.; Storchova, H.; White, J.C.; Vanek, T. Nanoparticle-specific changes in Arabidopsis thaliana gene expression after exposure to ZnO, TiO2, and fullerene soot. J. Hazard. Mater. 2012, 241–242, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Tumburu, L.; Andersen, C.P.; Rygiewicz, P.T.; Reichman, J.R. Phenotypic and genomic responses to titanium dioxide and cerium oxide nanoparticles in Arabidopsis germinants. Environ. Toxicol. Chem. 2015, 34, 70–83. [Google Scholar] [CrossRef]
- Simon, D.F.; Domingos, R.F.; Hauser, C.; Hutchins, C.M.; Zerges, W.; Wilkinson, K.J. Transcriptome sequencing (RNA-seq) analysis of the effects of metal nanoparticle exposure on the transcriptome of Chlamydomonas reinhardtii. Appl. Environ. Microbiol. 2013, 79, 4774–4785. [Google Scholar] [CrossRef] [Green Version]
- Beauvais-Flück, R.; Slaveykova, V.I.; Cosio, C. Comparative study of Cu uptake and early transcriptome responses in the green microalga Chlamydomonas reinhardtii and the macrophyte Elodea nuttallii. Environ. Pollut. 2019, 250, 331–337. [Google Scholar] [CrossRef]
- Xiong, T.; Zhang, T.; Xian, Y.; Kang, Z.; Zhang, S.; Dumat, C.; Shahid, M.; Li, S. Foliar uptake, biotransformation, and impact of CuO nanoparticles in Lactuca sativa L. var. ramosa Hort. Environ. Geochem. Health 2020, 43, 423–439. [Google Scholar] [CrossRef]
- Manusadžianas, L.; Gylytė, B.; Grigutytė, R.; Karitonas, R.; Sadauskas, K.; Vitkus, R.; Šiliauskas, L.; Vaičiūnienė, J. Accumulation of copper in the cell compartments of charophyte Nitellopsis obtusa after its exposure to copper oxide nanoparticle suspension. Environ. Sci. Pollut. Res. 2017, 24, 27653–27661. [Google Scholar] [CrossRef]
- Wu, X.; Song, H.; Guan, C.; Zhang, Z. Boron alleviates cadmium toxicity in Brassica napus by promoting the chelation of cadmium onto the root cell wall components. Sci. Total Environ. 2020, 728, 138833. [Google Scholar] [CrossRef]
- Maris, A.; Kaewthai, N.; Eklöf, J.M.; Miller, J.G.; Brumer, H.; Fry, S.C.; Verbelen, J.P.; Vissenberg, K. Differences in enzymic properties of five recombinant xyloglucan endotransglucosylase/hydrolase (XTH) proteins of Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, C.; Branco-Neves, S.; De Sousa, A.; Azenha, M.; Cunha, A.; Pereira, R.; Fidalgo, F. SiO2 nanomaterial as a tool to improve Hordeum vulgare L. tolerance to nano-NiO stress. Sci. Total Environ. 2018, 622–623, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.; Soares, C.; Pinto, A.S.; Fidalgo, F. Phytotoxic effects of bulk and nano-sized Ni on Lycium barbarum L. grown in vitro–Oxidative damage and antioxidant response. Chemosphere 2019, 218, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Küpper, H.; Küpper, F.; Spiller, M. Environmental relevance of heavy metal-substituted chlorophylls using the example of water plants. J. Exp. Bot. 1996, 47, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Thomas, G.; Stärk, H.-J.; Wellenreuther, G.; Dickinson, B.C.; Küpper, H. Effects of nanomolar copper on water plants—Comparison of biochemical and biophysical mechanisms of deficiency and sublethal toxicity under environmentally relevant conditions. Aquat. Toxicol. 2013, 140–141, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Bashri, G.; Parihar, P.; Singh, R.; Patel, A.; Prasad, S.M. Plant and Nanoparticle Interface at the Molecular Level: An Integrated Overview. In Nanomaterials in Plants, Algae, and Microorganisms; Academic Press: Cambridge, MA, USA, 2018; pp. 325–344. [Google Scholar] [CrossRef]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 182–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkens, S. Structure and mechanism of ABC transporters. F1000Prime Rep. 2015, 7–14. [Google Scholar] [CrossRef]
- Hwang, J.U.; Song, W.Y.; Hong, D.; Ko, D.; Yamaoka, Y.; Jang, S.; Yim, S.; Lee, E.; Khare, D.; Kim, K.; et al. Plant ABC Transporters Enable Many Unique Aspects of a Terrestrial Plant’s Lifestyle. Mol. Plant 2016, 9, 338–355. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, P.; Zanella, L.; De Paolis, A.; Di Litta, D.; Cecchetti, V.; Falasca, G.; Barbieri, M.; Altamura, M.M.; Costantino, P.; Cardarelli, M. Cadmium-inducible expression of the ABC-type transporter AtABCC3 increases phytochelatin-mediated cadmium tolerance in Arabidopsis. J. Exp. Bot. 2015, 66, 3815–3829. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, M.; Krishnamurthy, S.; Shukla, D.; Kiiskila, J.; Jain, A. Comparative transcriptome and proteome analysis to reveal the biosynthesis of gold nanoparticles in Arabidopsis. Sci. Rep. 2016, 6, 21733. [Google Scholar] [CrossRef] [Green Version]
- Xia, T.; Kovochich, M.; Brant, J.; Hotze, M.; Sempf, J.; Oberley, T.; Sioutas, C.; Yeh, J.I.; Wiesner, M.R.; Nel, A.E. Comparison of the abilities of ambient and manufactured nanoparticles to induce cellular toxicity according to an oxidative stress paradigm. Nano Lett. 2006, 6, 1794–1807. [Google Scholar] [CrossRef] [PubMed]
- Iversen, T.G.; Skotland, T.; Sandvig, K. Endocytosis and intracellular transport of nanoparticles: Present knowledge and need for future studies. Nano Today 2011, 6, 176–185. [Google Scholar] [CrossRef]
- Samaj, J.; Baluska, F.; Voigt, B.; Schlicht, M.; Volkmann, D.; Menzel, D. Endocytosis, actin cytoskeleton, and signaling. Plant Physiol. 2004, 135, 1150–1161. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.U.; Rono, J.K.; Zhang, B.Q.; Liu, X.S.; Wang, M.Q.; Wang, L.L.; Wu, X.C.; Chen, X.; Cao, H.W.; Yang, Z.M. Identification of novel rice (Oryza sativa) HPP and HIPP genes tolerant to heavy metal toxicity. Ecotoxicol. Environ. Saf. 2019, 175, 8–18. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, X.; Liu, J.; Niu, Y.; Chen, Y.; Hao, Y.; Zhao, J.; Sun, L.; Wang, H.; Xiao, J.; et al. Characterization of the Heavy-Metal-Associated Isoprenylated Plant Protein (HIPP) Gene Family from Triticeae Species. Int. J. Mol. Sci. 2020, 21, 6191. [Google Scholar] [CrossRef] [PubMed]
- De Abreu-Neto, J.B.; Turchetto-Zolet, A.C.; De Oliveira, L.F.V.; Bodanese Zanettini, M.H.; Margis-Pinheiro, M. Heavy met-al-associated isoprenylated plant protein (HIPP): Characterization of a family of proteins exclusive to plants. FEBS J. 2013, 280, 1604–1616. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Xiao, S.; Li, H.Y.; Tsao, S.W.; Chye, M.L. Arabidopsis thaliana acyl-CoA-binding protein ACBP2 interacts with heavy-metal-binding farnesylated protein AtFP6. New Phytol. 2009, 181, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Yamaguchi, Y.; Koizumi, N.; Sano, H. Functional characterization of a heavy metal binding protein Cdl19 from Arabidopsis. Plant J. 2002, 32, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Shirazi, Z.; Abedi, A.; Kordrostami, M.; Burritt, D.J.; Hossain, M.A. Genome-wide identification and characterization of the metal tolerance protein (MTP) family in grape (Vitis vinifera L.). Biotech 2019, 9, 1–17. [Google Scholar] [CrossRef]
- Fu, X.Z.; Tong, Y.H.; Zhou, X.; Ling, L.L.; Chun, C.P.; Cao, L.; Zeng, M.; Peng, L.Z. Genome-wide identification of sweet orange (Citrus sinensis) metal tolerance proteins and analysis of their expression patterns under zinc, manganese, copper, and cadmium toxicity. Gene 2017, 629, 1–8. [Google Scholar] [CrossRef]
- Dai, J.; Wang, N.; Xiong, H.; Qiu, W.; Nakanishi, H.; Kobayashi, T.; Nishizawa, N.K.; Zuo, Y. The yellow stripe-like (YSL) gene functions in internal copper transport in peanut. Genes 2018, 9, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaumann, S.; Nickolaus, S.D.; Fürst, S.H.; Starck, S.; Schneider, S.; Ekkehard Neuhaus, H.; Trentmann, O. The tonoplast copper transporter COPT5 acts as an exporter and is required for interorgan allocation of copper in Arabidopsis thaliana. New Phytol. 2011, 192, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Molina, A.; Andrés-Colás, N.; Perea-García, A.; Del Valle-Tascõn, S.; Peñarrubia, L.; Puig, S. The intracellular arabidopsis COPT5 transport protein is required for photosynthetic electron transport under severe copper deficiency. Plant J. 2011, 65, 848–860. [Google Scholar] [CrossRef]
- Sancenón, V.; Puig, S.; Mira, H.; Thiele, D.J.; Peñarrubia, L. Identification of a copper transporter family in Arabidopsis thaliana. Plant Mol. Biol. 2003, 51, 577–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustin, J.L.; Loureiro, M.E.; Kim, D.; Na, G.; Tikhonova, M.; Salt, D.E. MTP1-dependent Zn sequestration into shoot vacuoles suggests dual roles in Zn tolerance and accumulation in Zn-hyperaccumulating plants. Plant J. 2009, 57, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Lanquar, V.; Ramos, M.S.; Lelièvre, F.; Barbier-Brygoo, H.; Krieger-Liszkay, A.; Krämer, U.; Thomine, S. Export of vacuolar manganese by AtNRAMP3 and AtNRAMP4 is required for optimal photosynthesis and growth under manganese deficiency. Plant Physiol. 2010, 152, 1986–1999. [Google Scholar] [CrossRef] [Green Version]
- Rutschow, H.L.; Baskin, T.I.; Kramer, E.M. The carrier AUXIN RESISTANT (AUX1) dominates auxin flux into Arabidopsis protoplasts. New Phytol. 2014, 204, 536–544. [Google Scholar] [CrossRef]
- Zazímalová, E.; Murphy, A.S.; Yang, H.; Hoyerová, K.; Hosek, P. Auxin transporters--why so many? Cold Spring Harb. Perspect. Biol. 2010, 2, a001552. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Li, J.; Chong, K.; Harter, K.; Lee, Y.; Leung, J.; Martinoia, E.; Matsuoka, M.; Offringa, R.; Qu, L.; et al. Toward a Molecular Understanding of Plant Hormone Actions. Mol. Plant 2016, 9, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Zhang, Y.; Wang, J.; Yan, X.; Wang, C.; Xu, J.; Pan, J. Clathrin-Mediated Auxin Efflux and Maxima Regulate Hypocotyl Hook Formation and Light-Stimulated Hook Opening in Arabidopsis. Mol. Plant 2016, 9, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Zhang, P.; Zhang, Z.; He, X.; Li, Y.; Zhang, J.; Zheng, L.; Chu, S. Origin of the different phytotoxicity and biotransformation of cerium and lanthanum oxide nanoparticles in cucumber. Nanotoxicology 2015, 9, 262–270. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Academic Press: New York, NY, USA, 1984; Volume 105, pp. 121–126. ISBN 0076-6879. [Google Scholar]
- Hemeda, H.M.; Klein, B.P. Effects of Naturally Occurring Antioxidants on Peroxidase Activity of Vegetable Extracts. J. Food Sci. 1990, 55, 184–185. [Google Scholar] [CrossRef]
- Mishra, A.; Patel, M.K.; Jha, B. Non-targeted metabolomics and scavenging activity of reactive oxygen species reveal the potential of Salicornia brachiata as a functional food. J. Funct. Foods 2015, 13, 21–31. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Zhang, S.F.; Zhang, K.; Cheng, H.M.; Lin, L.; Wang, D.Z. Comparative transcriptomics reveals colony formation mechanism of a harmful algal bloom species Phaeocystis globosa. Sci. Total Environ. 2020, 719, 137454. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure Concentration (mg/L) | Sample | Total Clean Reads | Unique Mapped (%) | Multiple Mapped (%) | Total Mapped (%) | Q30 (%) | GC (%) |
|---|---|---|---|---|---|---|---|
| 0 | CK-1 | 48,984,912 | 43,707,254 (89.23%) | 2,323,237 (4.74%) | 46,030,491 (93.97%) | 90.54% | 45.85% |
| CK-2 | 56,584,572 | 46,144,248 (81.55%) | 4,497,480 (7.95%) | 50,641,728 (89.50%) | 90.20% | 47.17% | |
| CK-3 | 51,003,984 | 44,795,180 (87.83%) | 2,680,293 (5.26%) | 47,475,473 (93.08%) | 90.26% | 45.94% | |
| 100 | T1–1 | 84,539,086 | 75,290,760 (89.06%) | 4,295,130 (5.08%) | 79,585,890 (94.14%) | 95.63% | 46.16% |
| T1–2 | 70,293,714 | 59,781,276 (85.04%) | 4,518,475 (6.43%) | 64,299,751 (91.47%) | 95.99% | 47.32% | |
| T1–3 | 58,663,934 | 52,706,698 (89.85%) | 2,639,439 (4.50%) | 55,346,137 (94.34%) | 95.29% | 45.87% | |
| 1000 | T2–1 | 63,165,908 | 56,834,597 (89.98%) | 2,696,569 (4.27%) | 59,531,166 (94.25%) | 95.25% | 45.42% |
| T2–2 | 75,557,582 | 68,347,699 (90.46%) | 3,315,323 (4.39%) | 71,663,022 (94.85%) | 95.84% | 45.68% | |
| T2–3 | 52,794,452 | 46,679,090 (88.42%) | 2,503,238 (4.74%) | 49,182,328 (93.16%) | 95.05% | 45.87% | |
| Average total | 62,398,683 | 54,920,756 (87.94%) | 3,274,354 (5.26%) | 58,195,110 (93.20%) |
| Gene ID | log2 FC | Blast Annotation | Blasted Species | |
| T1 | T2 | |||
| Cell wall organization or biogenesis | ||||
| ncbi_111881311 | − | 1.47 | Cellulose synthase A catalytic subunit 4 (CESA4) | Daucus carota |
| ncbi_111907519 | 2.04 | 3.09 | Cellulose synthase-like protein D5 (CSLD5) | Cynara cardunculus |
| ncbi_111878486 | 1.42 | 1.95 | FASCICLIN-like arabinogalactan protein 8 (FLA8) | Cynara cardunculus |
| ncbi_111903865 | 2.32 | 3.41 | Barwin-like endoglucanase (EXPA1) | Cynara cardunculus |
| ncbi_111912962 | − | 1.11 | Xyloglucan endotransglucosylase/hydrolase 7 (XTH7) | Cynara cardunculus |
| ncbi_111891696 | 1.39 | 1.56 | Xyloglucan endotransglucosylase/hydrolase 8 (XTH8) | Daucus carota |
| ncbi_111905755 | 1.62 | 2.06 | Barwin-like endoglucanase (EXPA6) | Cynara cardunculus |
| ncbi_111916523 | 3.60 | 4.59 | Barwin-like endoglucanase (EXPA6) | Cynara cardunculus |
| ncbi_111894312 | − | 1.88 | Carbohydrate-binding domain CBM49 (At1g64390) | Cynara cardunculus |
| ncbi_111918933 | 1.57 | 1.24 | Cellulose synthase-like protein D3 (CSLD3) | Ziziphus jujuba |
| ncbi_111902216 | − | 1.86 | Barwin-like endoglucanase (EXPA4) | Cynara cardunculus |
| ncbi_111903657 | − | 3.70 | Barwin-like endoglucanase (EXPA4) | Cynara cardunculus |
| ncbi_111876521 | − | 4.81 | Barwin-like endoglucanase (EXPA10) | Cynara cardunculus |
| ncbi_111917522 | − | 8.02 | Barwin-like endoglucanase (EXPA10) | Cynara cardunculus |
| Photosynthesis | ||||
| ncbi_111915071 | − | −1.31 | Light-harvesting chlorophyll a/b-binding protein 5 (LHCB5) | Citrus sinensis |
| ncbi_111914553 | − | −1.38 | Photosystem I chlorophyll a/b-binding protein 3 (LHCA3) | Brassica rapa |
| ncbi_111877237 | − | −1.83 | Chlorophyll a/b-binding protein 6A (CAB6A) | Cynara cardunculus |
| ncbi_111914739 | − | −2.38 | Chlorophyll a/b-binding protein 3C (CAB3C) | Cynara cardunculus |
| MSTRG.16095 | −8.70 | −13.0 | Photosystem I PsaG/PsaK domain-containing protein (psaK) | Cynara cardunculus |
| ncbi_111880682 | − | −1.14 | Photosystem I reaction center subunit psaK (psaK) | Daucus carota |
| ncbi_111898767 | − | −1.74 | Photosystem I reaction center subunit psaD (psaD) | Glycine max |
| ncbi_111894261 | − | −1.01 | Photosystem I reaction center subu-nit psaD (psaD) | Malus domestica |
| ncbi_111918291 | − | −1.57 | Photosystem I reaction center subunit psaL (psaL) | Erythranthe guttata |
| MSTRG.8984 | − | −1.20 | Photosystem II CP47 reaction center protein (psbB) | Arachis duranensis |
| MSTRG.12634 | − | −1.18 | Photosystem II 47 kDa protein (psbB) | Lactuca sativa |
| ncbi_3772835 | − | −1.17 | Photosystem II 47 kDa protein (psbB) | Lactuca sativa |
| ncbi_3772836 | − | −1.39 | Photosystem II reaction center protein Z (psbZ) | Lactuca sativa |
| ncbi_111881262 | − | −1.43 | Photosystem II reaction center protein W (psbW) | Cynara cardunculus |
| ncbi_111895461 | − | −1.06 | Cytochrome b6-f complex Fe-S subunit (petC) | Cynara cardunculus |
| MSTRG.22911 | − | −5.27 | Cytochrome b6-f complex Fe-S subunit (petC) | Cynara cardunculus |
| ncbi_3772843 | − | −1.06 | Photosystem II reaction center protein I (psbI) | Phoenix dactylifera |
| ncbi_3772900 | − | −1.14 | Cytochrome f (petA) | Lactuca sativa |
| Oxidation-reduction process, antioxidant activity | ||||
| ncbi_111914667 | 1.00 | 2.00 | Peroxidase activity protein (POD1) | Populus tomentosa |
| ncbi_111907781 | − | 1.16 | Monodehydroascorbate reductase 4, peroxisome (MDAR4) | Vitis vinifera |
| ncbi_111882573 | − | 4.07 | Ascorbate peroxidase (APX2) | Ipomoea trifida |
| ncbi_111905866 | 1.02 | 1.08 | Manganese/iron superoxide dismutase (FSD2) | Cynara cardunculus |
| ncbi_111884404 | 1.11 | 1.47 | Manganese/iron superoxide dismutase (FSD3) | Cynara cardunculus |
| ncbi_111897600 | 1.84 | − | Flavanone 3-hydroxylase (FHT) | Erigeron breviscapus |
| ncbi_111905867 | − | −3.32 | Iron superoxide dismutase isoform 2 (SODB) | Solanum lycopersicum |
| ncbi_111900015 | − | −1.58 | Carotenoid oxygenase (CCD4) | Lactuca sativa |
| Transport | ||||
| ncbi_111921599 | − | 1.90 | ABC transporter B family member 19 (ABCB19) | Cynara cardunculus |
| ncbi_111890853 | − | 2.33 | ABC transporter C family member 10 (ABCC10) | Populus euphratica |
| ncbi_111879294 | 2.44 | 4.38 | ABC transporter G family members 5 (ABCG5) | Cynara cardunculus |
| ncbi_111907936 | 1.47 | 2.07 | ABC transporter G family member 36 (ABCG36) | Beta vulgaris |
| ncbi_111910825 | − | 1.03 | ABC transporter C family member 12 (ABCC12) | Vitis vinifera |
| ncbi_111888188 | −1.23 | − | ABC transporter C family member 3 (ABCC3) | Juglans regia |
| ncbi_111885307 | 4.42 | 5.35 | Alpha-tubulin (TUBA) | Cynara cardunculus |
| ncbi_111901730 | 1.56 | 1.83 | Tubulin beta−1 chain (TUBB1) | Sesamum indicum |
| ncbi_111900393 | 1.92 | 2.42 | Tubulin beta-2 chain(TUBB2) | Nelumbo nucifera |
| ncbi_111882692 | 2.97 | 3.73 | Beta-tubulin (TUBB8) | Cynara cardunculus |
| ncbi_111906603 | − | 1.09 | ADP-ribosylation factor GTPase-activating protein AGD11 isoform X1 (AGD11) | Cynara cardunculus |
| ncbi_111914610 | − | 5.29 | ADP-ribosylation factor GTPase-activating protein AGD11 isoform X1 (AGD11) | Capsicum annuum |
| ncbi_111878707 | 4.65 | 5.39 | Heat shock protein 70 family (Hsc70) | Corchorus olitorius; |
| ncbi_111881479 | 3.67 | 4.98 | Heat shock cognate 70 kDa protein 2 (Hsc70) | Ricinus communis |
| ncbi_111913208 | − | 1.08 | Kinesin motor family protein isoform 1 (KINUC) | Theobroma cacao |
| ncbi_111921158 | − | 2.64 | Heavy metal-associated isoprenylated plant protein 37 (HIPP37) | Cephalotus follicularis |
| ncbi_111882047 | − | 4.22 | Heavy metal-associated isoprenylated plant protein 37 (HIPP37) | Cephalotus follicularis |
| ncbi_111886124 | − | 5.99 | Heavy metal-associated isoprenylated plant protein 3 (HIPP03) | Daucus carota |
| ncbi_111894055 | 3.67 | 3.92 | Heavy metal-associated isoprenylated plant protein (HIPP01) | Cynara cardunculus |
| ncbi_111894603 | 1.18 | 1.28 | Heavy metal-associated isoprenylated plant protein (HIPP39) | Cynara cardunculus |
| ncbi_111895123 | − | 1.05 | Heavy metal-associated isoprenylated plant protein (HIPP32) | Cynara cardunculus |
| ncbi_111895347 | 2.12 | 2.91 | Heavy metal-associated isoprenylated plant protein (HIPP07) | Daucus carota |
| ncbi_111896151 | 1.35 | 1.38 | Heavy metal-associated isoprenylated plant protein (HIPP36) | Cynara cardunculus |
| ncbi_111900275 | 1.06 | − | Heavy metal-associated isoprenylated plant protein (HIPP21) | Cynara cardunculus |
| ncbi_111900793 | 2.52 | 3.62 | Heavy metal-associated isoprenylated plant protein (HIPP31) | Nicotiana sylvestris |
| ncbi_111901657 | − | 1.44 | Copper transport protein ATX1 (HIPP31) | Nelumbo nucifera |
| ncbi_111903392 | 4.70 | 4.92 | Heavy metal-associated isoprenylated plant protein (HIPP09) | Cynara cardunculus |
| ncbi_111916369 | 1.13 | 1.43 | Heavy metal-associated isoprenylated plant protein (HIPP05) | Cynara cardunculus |
| ncbi_111919298 | 1.95 | 1.74 | Heavy metal-associated isoprenylated plant protein (HIPP26) | Cynara cardunculus |
| ncbi_111900175 | − | 1.59 | Cu-transporting ATPase responsive-to-antagonist1 (RAN1) | Juglans regia |
| ncbi_111913891 | 1.58 | 1.08 | Monocopper oxidase-like protein SKU5 (SKU5) | Erythranthe guttata |
| ncbi_111903866 | 2.35 | 2.94 | Monocopper oxidase-like protein SKU5 (SKU5) | Erythranthe guttata |
| ncbi_111913012 | − | 2.38 | Metal tolerance protein 4-like isoform X2 (MTP4) | Nicotiana attenuata |
| ncbi_111888942 | − | 7.82 | Cation efflux protein (MTP11) | Cynara cardunculus |
| ncbi_111892195 | 1.57 | 2.02 | Natural resistance-associated macrophage proteins family metal transporter 6 (NRAMP6) | Chengiopanax sciadophylloides |
| ncbi_111893192 | −1.68 | −2.59 | Copper transporter 5.1-like (COPT5.1) | Cynara cardunculus |
| Hormone signal transduction | ||||
| ncbi_111883505 | − | 1.19 | Auxin-induced protein 22D (AUX22D) | Daucus carota |
| ncbi_111886094 | − | 1.41 | Auxin responsive SAUR protein (SAUR50) | Cynara cardunculus |
| ncbi_111896245 | − | 1.46 | Auxin-responsive protein IAA9 like (IAA9) | Zinnia violacea |
| ncbi_111909089 | − | 1.45 | Auxin-responsive protein IAA9 like (IAA27) | Prunus mume |
| ncbi_111911429 | − | 1.03 | Basic-leucine zipper domain-containing protein (ABF2) | Cynara cardunculus |
| ncbi_111898981 | 1.35 | 2.12 | Auxin transporter-like protein 2 (LAX2) | Dorcoceras hygrometricum |
| ncbi_111881996 | 8.32 | 8.87 | Auxin influx carrier protein (LAX2) | Zinnia violacea |
| ncbi_111901327 | − | 1.07 | Auxin influx carrier protein (LAX2) | Zinnia violacea |
| ncbi_111895220 | − | 2.00 | CheY-like superfamily (ARR2) | Cynara cardunculus |
| ncbi_111877939 | 1.98 | 2.04 | CheY-like superfamily (ARR6) | Cynara cardunculus |
| ncbi_111921291 | 1.04 | 1.06 | Signal transduction histidine kinase, phosphotransfer (Hpt) domain-containing protein (AHP1) | Cynara cardunculus |
| ncbi_111878847 | 2.44 | 2.89 | Glycoside hydrolase, catalytic domain-containing protein (At3g13560) | Cynara cardunculus |
| ncbi_111902332 | − | 7.44 | Auxin efflux carrier component 2 (PIN2) | Cynara cardunculus |
| ncbi_111881242 | − | 1.20 | Auxin-responsive protein IAA12-like (IAA12) | Daucus carota |
| ncbi_111894991 | 1.12 | 1.74 | Auxin response factor 3 isoform X1 (ARF3) | Erythranthe guttata |
| ncbi_111898759 | − | 1.22 | Auxin response factor (ARF9) | Cynara cardunculus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, T.; Zhang, S.; Kang, Z.; Zhang, T.; Li, S. Dose-Dependent Physiological and Transcriptomic Responses of Lettuce (Lactuca sativa L.) to Copper Oxide Nanoparticles—Insights into the Phytotoxicity Mechanisms. Int. J. Mol. Sci. 2021, 22, 3688. https://doi.org/10.3390/ijms22073688
Xiong T, Zhang S, Kang Z, Zhang T, Li S. Dose-Dependent Physiological and Transcriptomic Responses of Lettuce (Lactuca sativa L.) to Copper Oxide Nanoparticles—Insights into the Phytotoxicity Mechanisms. International Journal of Molecular Sciences. 2021; 22(7):3688. https://doi.org/10.3390/ijms22073688
Chicago/Turabian StyleXiong, Tiantian, Shasha Zhang, Zhuangzhuang Kang, Ting Zhang, and Shaoshan Li. 2021. "Dose-Dependent Physiological and Transcriptomic Responses of Lettuce (Lactuca sativa L.) to Copper Oxide Nanoparticles—Insights into the Phytotoxicity Mechanisms" International Journal of Molecular Sciences 22, no. 7: 3688. https://doi.org/10.3390/ijms22073688