
High Diversity of β-Glucosidase-Producing Bacteria and Their Genes Associated with Scleractinian Corals

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Diversity of β-Glucosidase-Producing Bacteria Associated with Scleractinian Corals
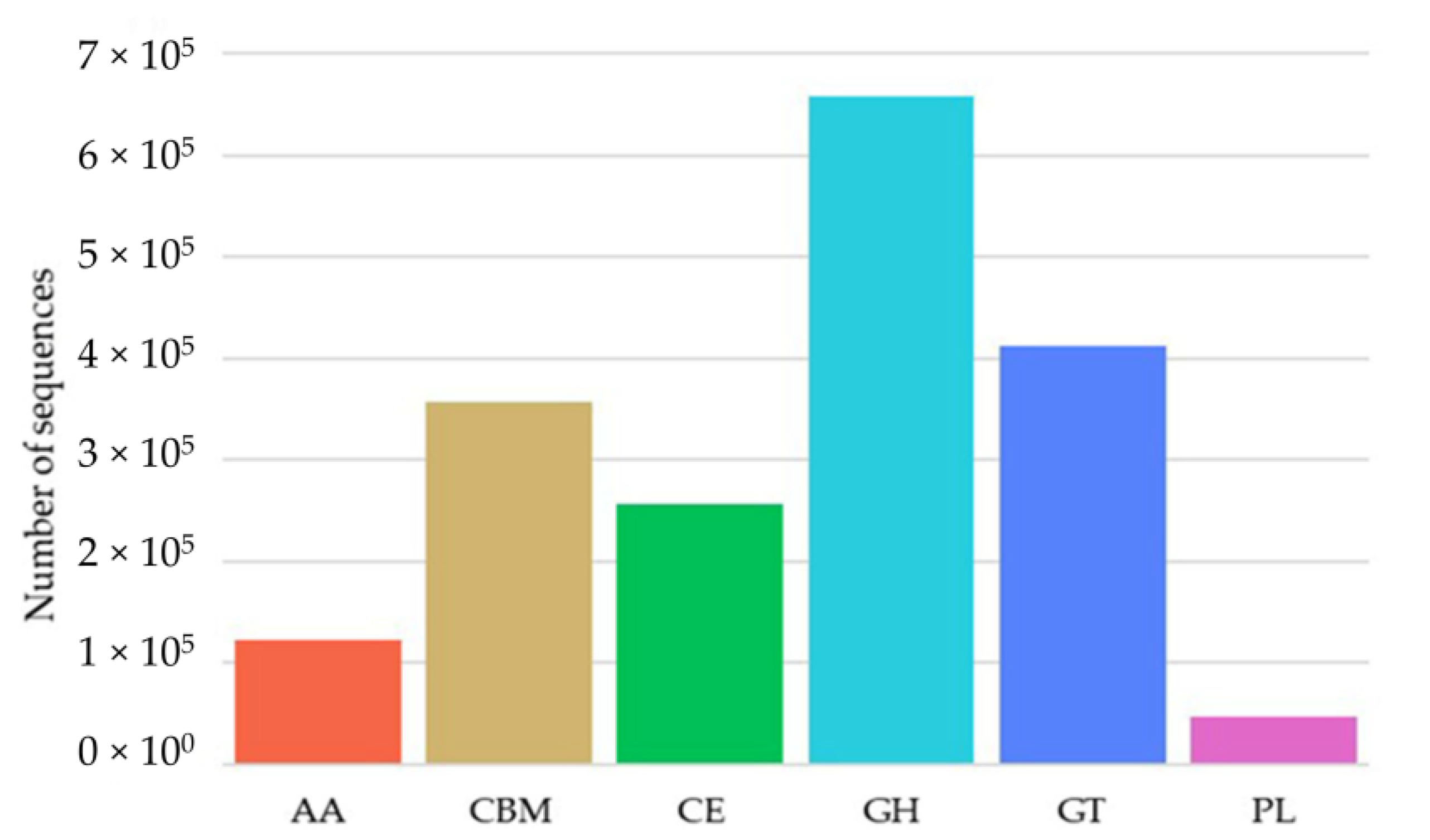
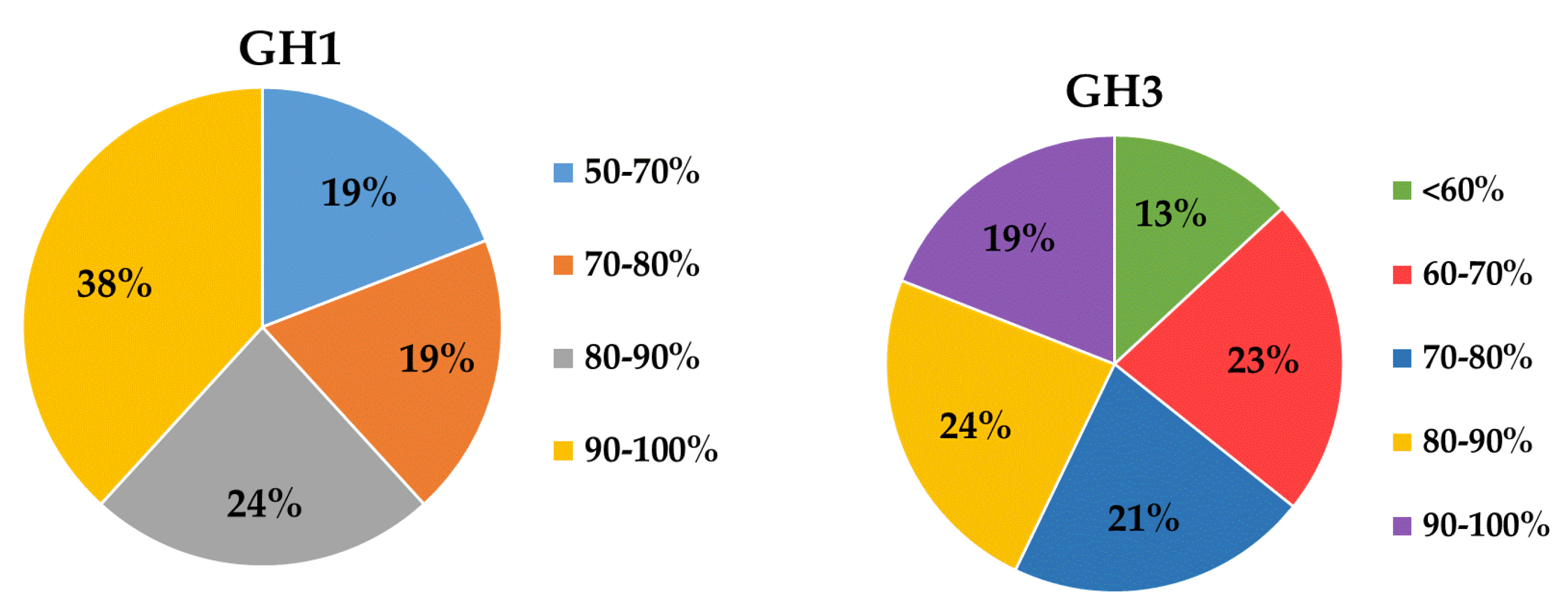
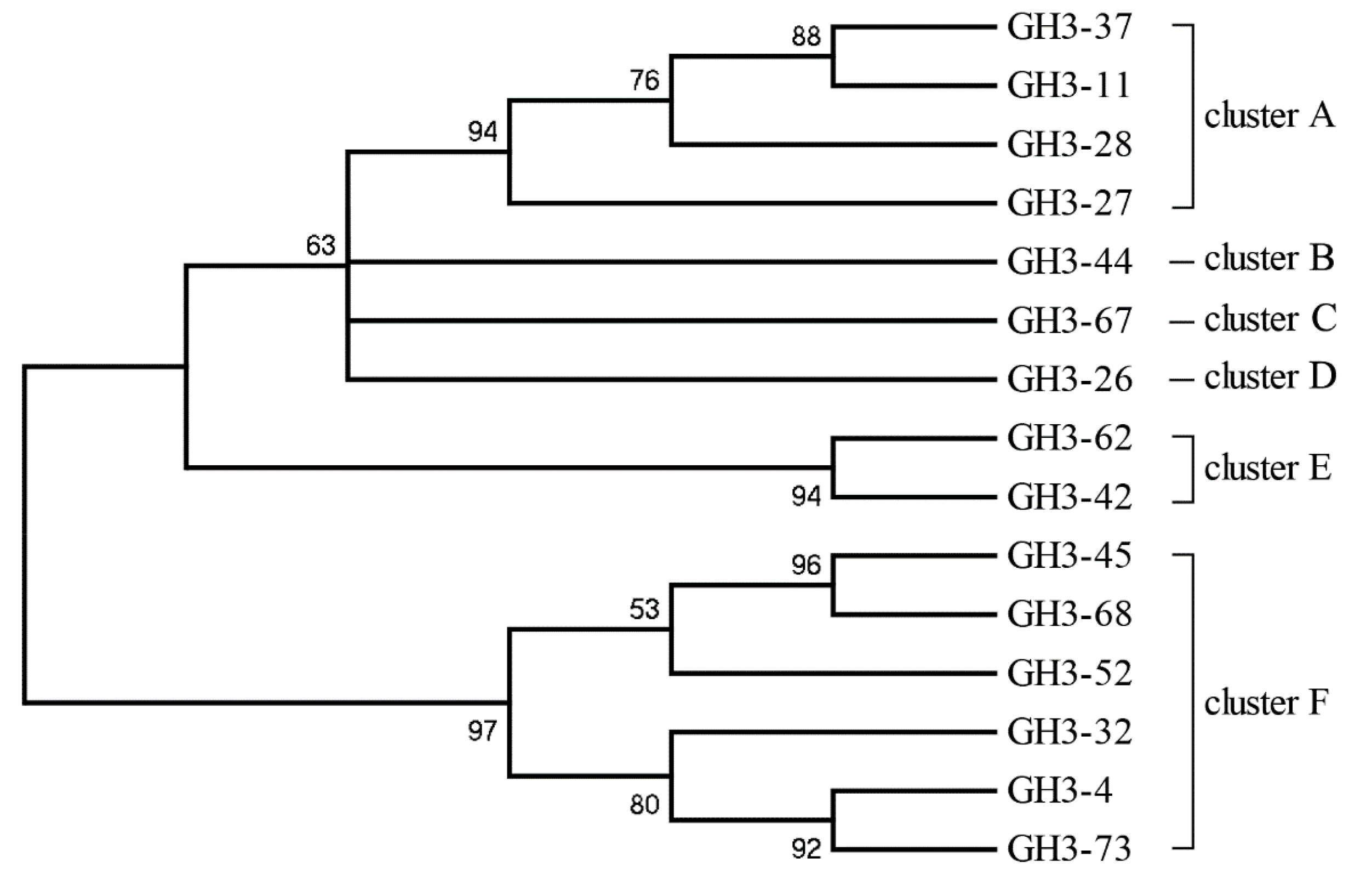
2.2. Diversity of the β-Glucosidase Gene
2.3. Expression and Purification of Mg9373
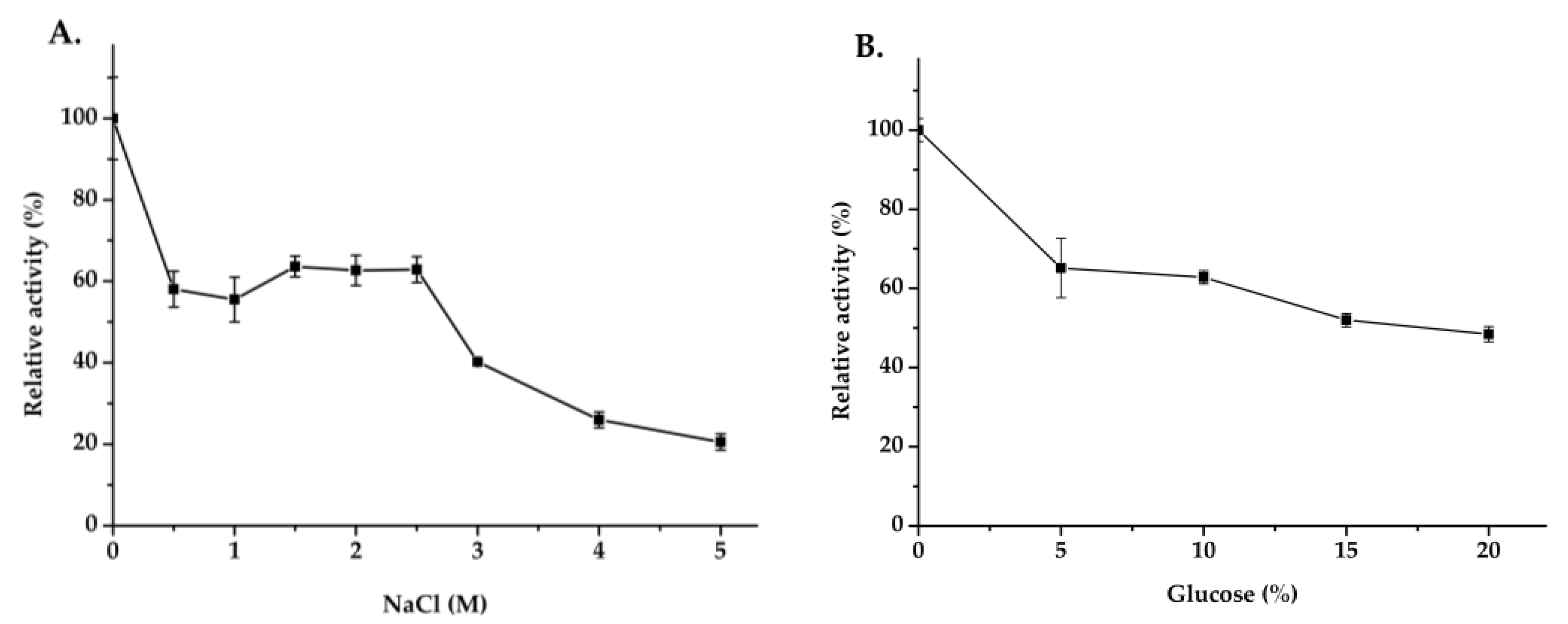
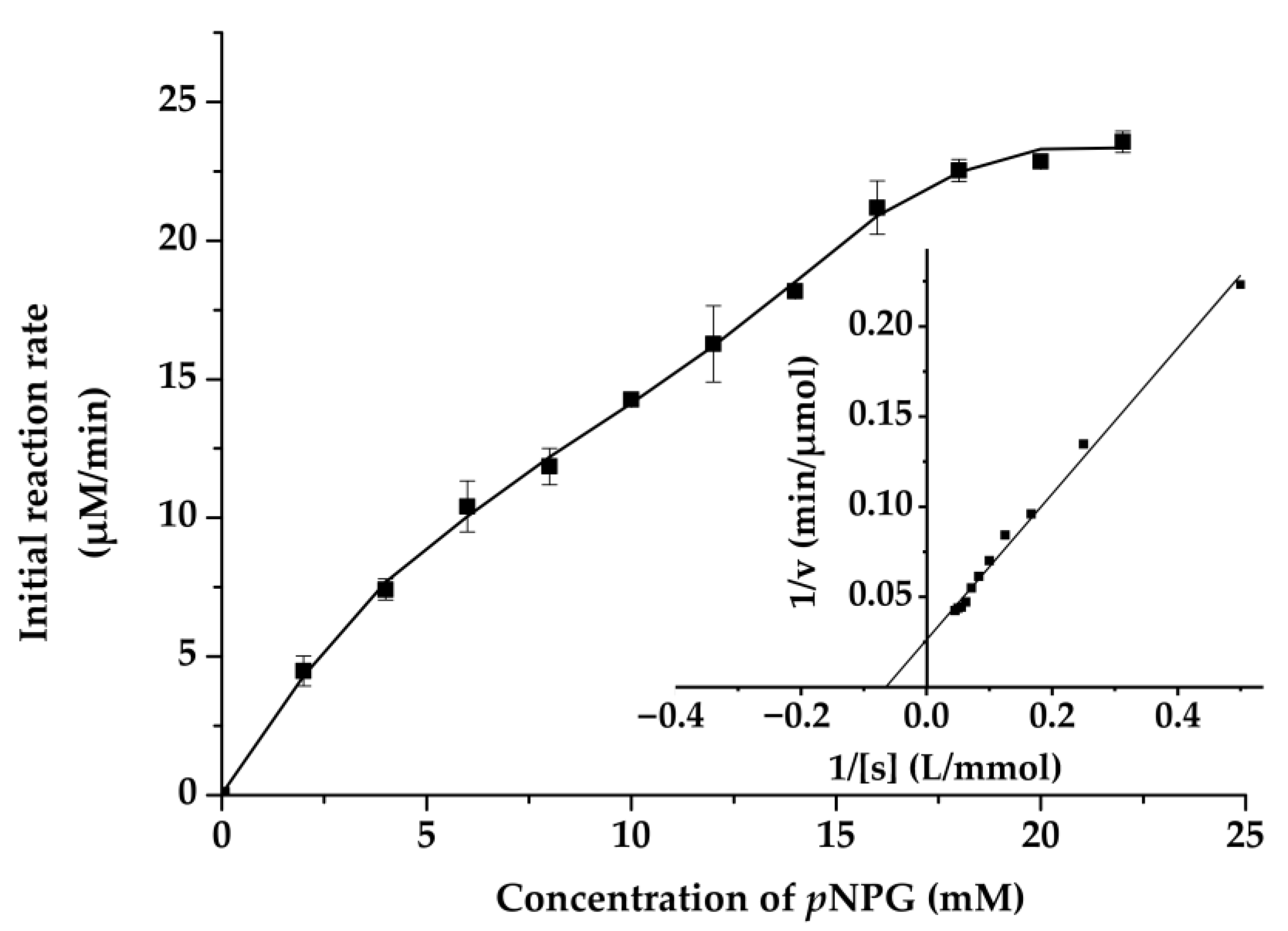
2.4. Biochemical Characterization of Mg9373
3. Discussion
3.1. Diversity Analysis of β-Glucosidase-Producing Bacteria Associated with Scleractinian Corals
3.2. Diversity Analysis of β-Glucosidase Gene
3.3. Expression and Purification of Mg9373
3.4. Biochemical Characterization of Mg9373
4. Materials and Methods
4.1. Coral Sample Collection
4.2. Isolation and Purification of β-Glucosidase-Producing Bacteria
4.3. Molecular Identification of the Bacterial Strains
4.4. Diversity of the β-Glucosidase Gene
4.5. Cloning and Expression of the β-Glucosidase Gene
4.6. Purification and Identification of Mg9373
4.7. Effect of pH, Temperature, and Metal Ions on the Activity and Stability of Mg9373
4.8. Enzyme Activity and Kinetic Assay of Mg9373
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| IPTG | isopropylthio-β-galactoside |
| SDS-PAGE | sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| pNPG | pNP-β-D-glucopyranoside |
References
- Forest, R.; Victor, S.; Farooq, A.; Nancy, K. Diversity and distribution of coral-associated bacteria. Mar. Ecol. Prog. Ser. 2002, 243, 1–10. [Google Scholar]
- Pernice, M.; Raina, J.B.; Radecker, N.; Cardenas, A.; Pogoreutz, C.; Voolstra, C.R. Down to the bone: The role of overlooked endolithic microbiomes in reef coral health. ISME J. 2020, 14, 325–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eugene, R. The role of microorganisms in coral health, disease and evolution. Nat. Rev. Microbiol. 2007, 5, 355–362. [Google Scholar]
- Garren, M.; Azam, F. New method for counting bacteria associated with coral mucus. Appl. Environ. Microbiol. 2010, 76, 6128–6133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garren, M.; Azam, F. New directions in coral reef microbial ecology. Environ. Microbiol. 2012, 14, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Rohwer, F.; Breitbart, M.; Jara, J.; Azam, F.; Knowlton, N. Diversity of bacteria associated with the caribbean coral Montastraea franksi. Coral Reefs 2001, 20, 85–91. [Google Scholar]
- Rosenberg, E. Coral microbiology. Microb. Biotechnol. 2009, 2, 147. [Google Scholar] [CrossRef] [Green Version]
- Meron, D.; Atias, E.; Kruh, L.I.; Elifantz, H.; Minz, D.; Fine, M.; Banin, E. The impact of reduced ph on the microbial community of the coral Acropora eurystoma. ISME J. 2011, 5, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chen, Q.; Long, L.-J.; Dong, J.-D.; Yang, J.; Zhang, S. Bacterial dynamics within the mucus, tissue and skeleton of the coral porites lutea during different seasons. Sci. Rep. 2014, 4, 7320. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, T.D.; Krause, L.; Bridge, T.; Torda, G.; Raina, J.B.; Zakrzewski, M.; Gates, R.D.; Padilla-Gamiño, J.L.; Spalding, H.L.; Smith, C.; et al. The coral core microbiome identifies rare bacterial taxa as ubiquitous endosymbionts. ISME J. 2015, 9, 2261–2274. [Google Scholar] [CrossRef]
- Chen, C.P.; Tseng, C.H.; Chen, C.A.; Tang, S.L. The dynamics of microbial partnerships in the coral Isopora palifera. ISME J. 2011, 5, 728–740. [Google Scholar] [CrossRef] [Green Version]
- Hong, M.J.; Yu, Y.T.; Chen, C.A.; Chiang, P.W.; Tang, S.L. Influence of species specificity and other factors on bacteria associated with the coral Stylophora pistillata in taiwan. Appl. Environ. Microbiol. 2009, 75, 7797–7806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Yu, K.; Wang, Y.; Huang, X.; Huang, W.; Qin, Z.; Pan, Z.; Yao, Q.; Wang, W.; Wu, Z. Distinct bacterial communities associated with massive and branching scleractinian corals and potential linkages to coral susceptibility to thermal or cold stress. Front. Microbiol. 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Rajasabapathy, R.; Ramasamy, K.P.; Manikandan, B.; Mohandass, C.; James, R.A. Bacterial communities associated with healthy and diseased (skeletal growth anomaly) reef coral Acropora cytherea from palk bay, India. Front. Mar. Sci. 2020, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Yu, K.; Liang, Y.; Chen, B.; Huang, X. Latitudinal variation in reef coral tissue thickness in the south china sea: Potential linkage with coral tolerance to environmental stress. Sci. Total Environ. 2020, 711, 134610. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Xiao, Z.; Yu, K.; Huang, Q.; Wang, G.; Wang, Y.; Liang, J.; Huang, W.; Huang, X.; Wei, F.; et al. Diversity of cultivable protease-producing bacteria and their extracellular proteases associated to scleractinian corals. PeerJ 2020, 8, e9055. [Google Scholar] [CrossRef] [PubMed]
- Reshef, L.; Koren, O.; Loya, Y.; Zilber-Rosenberg, I.; Rosenberg, E. The coral probiotic hypothesis. Environ. Microbiol. 2006, 8, 2068–2073. [Google Scholar] [CrossRef] [Green Version]
- Carlson, C.; Giovannoni, S.; Hansell, D.; Goldberg, S.; Parsons, R.; Otero, M.; Vergin, K.; Wheeler, B. Effect of nutrient amendments on bacterioplankton production, community structure, and doc utilization in the northwestern Sargasso Sea. Aquat. Microb. Ecol. 2002, 30, 19–36. [Google Scholar] [CrossRef]
- Kline, D.I.; Kuntz, N.M.; Breitbart, M.; Knowlton, N.; Rohwer, F. Role of elevated organic carbon levels and microbial activity in coral mortality. Mar. Ecol. Prog. Ser. 2006, 314, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Wegley, L.; Edwards, R.; Rodriguez-Brito, B.; Liu, H.; Rohwer, F. Metagenomic analysis of the microbial community associated with the coral Porites astreoides. Environ. Microbiol. 2007, 9, 2707–2719. [Google Scholar] [CrossRef]
- Kimes, N.E.; Van Nostrand, J.D.; Weil, E.; Zhou, J.; Morris, P.J. Microbial functional structure of montastraea faveolata, an important caribbean reef-building coral, differs between healthy and yellow-band diseased colonies. Environ. Microbiol. 2010, 12, 541–556. [Google Scholar] [CrossRef]
- Rodrigues, L.J.; Grottoli, A.G. Energy reserves and metabolism as indicators of coral recovery from bleaching. Limnol. Oceanogr. 2007, 52, 1874–1882. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Leletkin, V.A.; Tsukahara, J.; van Woesik, R.; Yamazato, K. Degradation of zooxanthellae and regulation of their density in hermatypic corals. Mar. Ecol. Prog. Ser. 1996, 139, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, Y.; Tanabe, T.; Iguchi, A. The presence of genes encoding enzymes that digest carbohydrates in coral genomes and analysis of their activities. Peerj 2017, 5, e4087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krediet, C.J.; Ritchie, K.B.; Teplitski, M. Catabolite regulation of enzymatic activities in a white pox pathogen and commensal bacteria during growth on mucus polymers from the coral Acropora palmata. Dis. Aquat. Org. 2009, 87, 57–66. [Google Scholar] [CrossRef]
- Akhtar, N.; Aanchal Goyal, D.; Goyal, A. Biodiversity of cellulase producing bacteria and their applications. Cell. Chem. Technol. 2016, 50, 9–10. [Google Scholar]
- Hoppe, H.-G. Significance of exoenzymatic activities in the ecology of brackish water: Measurements by means of methylumbelliferyl-substrates. Mar. Ecol. Prog. Ser. 1983, 11, 299–308. [Google Scholar] [CrossRef]
- Smith, D.C.; Simon, M.; Alldredge, A.L.; Azam, F. Intense hydrolytic enzyme activity on marine aggregates and implications for rapid particle dissolution. Nature 1992, 359, 139–142. [Google Scholar] [CrossRef]
- Fu, Y.; Yin, Z.; Wu, L.; Yin, C. Diversity of cultivable β-glycosidase-producing micro-organisms isolated from the soil of a ginseng field and their ginsenosides-hydrolysing activity. Lett. Appl. Microbiol. 2014, 58, 138–144. [Google Scholar] [CrossRef]
- Tiwari, R.; Kumar, K.; Singh, S.; Nain, L.; Shukla, P. Molecular detection and environment-specific diversity of glycosyl hydrolase family 1 β-glucosidase in different habitats. Front. Microbiol. 2016, 7, 1597. [Google Scholar] [CrossRef]
- Rath, J.; Herndl, G.J. Characteristics and diversity of beta-d-glucosidase (ec 3.2.1.21) activity in marine snow. Appl. Environ. Microbiol. 1994, 60, 807–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrieta, J.M.; Herndl, G.J. Assessing the diversity of marine bacterial beta-glucosidases by capillary electrophoresis zymography. Appl. Environ. Microbiol. 2001, 67, 4896–4900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebauer, D.S. Litter degradation rate and β-glucosidase activity increase with fungal diversity. Can. J. For. Res. 2010, 40, 1076–1085. [Google Scholar] [CrossRef]
- De Gannes, V.; Bekele, I.; Dipchansingh, D.; Wuddivira, M.N.; De Cairies, S.; Boman, M.; Hickey, W.J. Microbial community structure and function of soil following ecosystem conversion from native forests to teak plantation forests. Front. Microbiol. 2016, 7, 1976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrieta, J.M.; Herndl, G.J. Changes in bacterial β-glucosidase diversity during a coastal phytoplankton bloom. Limnol. Oceanogr. 2002, 47, 594–599. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira, C.B.; Cavalcanti, G.S.; Walter, J.M.; Silva-Lima, A.W.; Dinsdale, E.A.; Bourne, D.G.; Thompson, C.C.; Thompson, F.L. Microbial processes driving coral reef organic carbon flow. FEMS Microbiol. Rev. 2017, 41, 575–595. [Google Scholar] [CrossRef] [Green Version]
- Smillie, C.S.; Smith, M.B.; Friedman, J.; Cordero, O.X.; David, L.A.; Alm, E.J. Ecology drives a global network of gene exchange connecting the human microbiome. Nature 2011, 480, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Bailey, V.L.; Fansler, S.J.; Stegen, J.C.; McCue, L.A. Linking microbial community structure to β-glucosidic function in soil aggregates. ISME J. 2013, 7, 2044–2053. [Google Scholar] [CrossRef]
- Prada, F.; Caroselli, E.; Mengoli, S.; Brizi, L.; Fantazzini, P.; Capaccioni, B.; Pasquini, L.; Fabricius, K.E.; Dubinsky, Z.; Falini, G.; et al. Ocean warming and acidification synergistically increase coral mortality. Sci. Rep. 2017, 7, 40842. [Google Scholar] [CrossRef] [Green Version]
- Motone, K.; Takagi, T.; Aburaya, S.; Miura, N.; Aoki, W.; Ueda, M. A zeaxanthin-producing bacterium isolated from the algal phycosphere protects coral endosymbionts from environmental stress. mBio 2020, 11, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, P.; Li, Y.; Marshall, S.; Zhang, H. High genetic diversity of microbial cellulase and hemicellulase genes in the hindgut of Holotrichia parallela larvae. Int. J. Mol. Sci. 2015, 16, 16545–16559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Pozo, M.V.; Fernández-Arrojo, L.; Gil-Martínez, J.; Montesinos, A.; Chernikova, T.N.; Nechitaylo, T.Y.; Waliszek, A.; Tortajada, M.; Rojas, A.; Huws, S.A.; et al. Microbial β-glucosidases from cow rumen metagenome enhance the saccharification of lignocellulose in combination with commercial cellulase cocktail. Biotechnol. Biofuels 2012, 5, 73. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Duan, C.-J.; Liu, L.; Tang, J.-L.; Feng, J.-X. Properties of a metagenome-derived beta-glucosidase from the contents of rabbit cecum. Biosci. Biotechnol. Biochem. 2009, 73, 1470–1473. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Feng, Y.; Mo, X.; Duan, C.; Tang, J.; Feng, J. Cloning and expression of a beta-glucosidase gene umcel3G from metagenome of buffalo rumen and characterization of the translated product. Sheng Wu Gong Cheng Xue Bao 2008, 24, 232–238. [Google Scholar] [PubMed]
- Zhang, J.; Zhao, N.; Xu, J.; Qi, Y.; Wei, X.; Fan, M. Homology analysis of 35 beta-glucosidases in Oenococcus oeni and biochemical characterization of a novel beta-glucosidase BGL0224. Food Chem. 2021, 334, 127593. [Google Scholar] [CrossRef]
- Mattéotti, C.; Haubruge, E.; Thonart, P.; Francis, F.; De Pauw, E.; Portetelle, D.; Vandenbol, M. Characterization of a new beta-glucosidase/beta-xylosidase from the gut microbiota of the termite (Reticulitermes Santonensis). FEMS Microbiol. Lett. 2011, 314, 147–157. [Google Scholar] [CrossRef]
- Mai, Z.; Yang, J.; Tian, X.; Li, J.; Zhang, S. Gene cloning and characterization of a novel salt-tolerant and glucose-enhanced β-glucosidase from a marine Streptomycete. Appl. Biochem. Biotechnol. 2013, 169, 1512–1522. [Google Scholar] [CrossRef]
- Liew, K.J.; Lim, L.; Woo, H.Y.; Chan, K.G.; Shamsir, M.S.; Goh, K.M. Purification and characterization of a novel GH1 beta-glucosidase from Jeotgalibacillus malaysiensis. Int. J. Biol. Macromol. 2018, 115, 1094–1102. [Google Scholar] [CrossRef] [PubMed]
- Mohamad Sobri, M.F.; Abd-Aziz, S.; Abu Bakar, F.D.; Ramli, N. In-silico characterization of glycosyl hydrolase family 1 beta-glucosidase from Trichoderma asperellum UPM1. Int. J. Mol. Sci. 2020, 21, 4035. [Google Scholar] [CrossRef] [PubMed]
- Shipkowski SBrenchley, J.E. Characterization of an unusual cold-active beta-glucosidase belonging to family 3 of the glycoside hydrolases from the psychrophilic isolate paenibacillus sp. Strain c7. Appl. Environ. Microbiol. 2005, 71, 4225–4232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karnaouri, A.; Topakas, E.; Paschos, T.; Taouki, I.; Christakopoulos, P. Cloning, expression and characterization of an ethanol tolerant GH3 β-glucosidase from Myceliophthora thermophila. PeerJ 2013, 1, e46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuankhayan, P.; Rimlumduan, T.; Svasti, J.; Cairns, J.R.K. Hydrolysis of soybean isoflavonoid glycosides by dalbergia beta-glucosidases. J. Agric. Food Chem. 2007, 55, 2407. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, R.; Shi, P.; Huang, H.; Meng, K.; Yuan, T.; Yang, P.; Yao, B. A novel low-temperature-active β-glucosidase from Symbiotic serratia sp. Tn49 reveals four essential positions for substrate accommodation. Appl. Microbiol. Biotechnol. 2011, 92, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Kristen, D.; Neal, E.; Scott, F.; Chiara, P.; Morgan, S. Variation in growth rates of branching corals along Australia’s Great Barrier Reef. Sci. Rep. 2017, 7, 2920. [Google Scholar]
- Yamamura, H.; Hayakawa, M.; Iimura, Y. Application of sucrose-gradient centrifugation for selective isolation of Nocardia spp. From soil. J. Appl. Microbiol. 2003, 95, 677–685. [Google Scholar] [CrossRef]
- Ritchie, K.B. Regulation of microbial populations by coral surface mucus and mucus-associated bacteria. Mar. Ecol. Prog. 2006, 322, 1–14. [Google Scholar] [CrossRef]
- Li, Y.; Wu, C.; Zhou, M.; Wang, E.T.; Zhang, Z.; Liu, W.; Ning, J.; Xie, Z. Diversity of cultivable protease-producing bacteria in Laizhou bay sediments, bohai sea, china. Front. Microbiol. 2017, 8, 405. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Odoux, E.; Escoute, J.; Verdeil, J.L.; Brillouet, J.M. Localization of beta-D-glucosidase activity and glucovanillin in Vanilla Bean (Vanilla planifolia Andrews). Ann. Bot. 2003, 92, 437–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CORAL GENERA | Total and Rate (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Pocillopora | Acropora | Porites | Favia | Turbinaria | Montipora | |||
| GENERA DISTRIBUTION | ||||||||
| Fimicutes (6) | Bacillus | 1 | 1 | 3 | 5 (5.75%) | |||
| Lysinibacillus | 1 | 1 (1.15%) | ||||||
| Brevibacterium | 1 | 1 (1.15%) | ||||||
| Cytobacillus | 1 | 2 | 3 (3.45%) | |||||
| Fictibacillus | 3 | 3 (3.45%) | ||||||
| Exiguobacterium | 2 | 2 | 2 | 3 | 9 (10.35%) | |||
| Proteobacteria (11) | ||||||||
| Photobacterium | 14 | 14 (16.09%) | ||||||
| Sphingobium | 3 | 1 | 5 | 9 (10.35%) | ||||
| Brevundimonas | 1 | 1 | 2 (2.30%) | |||||
| Paracoccus | 9 | 3 | 12 (13.79%) | |||||
| Celeribacter | 1 | 1 (1.15%) | ||||||
| Pseudovibrio | 1 | 1 | 2 (2.30%) | |||||
| Stenotrophomonas | 1 | 1 (1.15%) | ||||||
| Pseudomonas | 3 | 3 (3.45%) | ||||||
| Pseudoalteromonas | 1 | 1 (1.15%) | ||||||
| Alteromonas | 2 | 1 | 3 (3.45%) | |||||
| Vibrio | 1 | 1 | 6 | 1 | 2 | 11 (12.64%) | ||
| Actinobacteria (4) | ||||||||
| Microbacterium | 2 | 1 | 3 (3.45%) | |||||
| Rothia | 1 | 1 (1.15%) | ||||||
| Nocardia | 1 | 1 (1.15%) | ||||||
| Brachybacterium | 1 | 1 (1.15%) | ||||||
| Total strain number (87) | 15 | 9 | 16 | 26 | 10 | 10 | 87 | |
| Family | Cazy-Activities | Coral Gene | Holotrichia parallela Gene |
|---|---|---|---|
| GH1 | Glucosidase | 68 | ND |
| GH2 | Galactosidase | 27 | 24 |
| GH3 | Glucosidase | 84 | 0 |
| GH5 | Endoglucanase | 14 | 0 |
| GH8 | Endoglucanase | 11 | 27 |
| GH11 | Xylanase | 1 | 14 |
| GH16 | Glucanase | 28 | ND |
| GH39 | Xylosidase | 5 | 0 |
| GH52 | Xylosidase | 1 | 0 |
| Substance | Relative Activity (%) a | |
|---|---|---|
| 10 mM | 20 mM | |
| Control | 100 b | 100 |
| Ni+ | 65.4 ± 7.2 | 115 ± 26.9 |
| Zn2+ | 140 ± 23.2 | 76 ± 1.2 |
| Cu2+ | 43 ± 0.4 | 34 ± 4.6 |
| Mn2+ | 351 ± 19.4 | 272 ± 39.5 |
| Mg2+ | 108 ± 15.1 | 115 ± 14.8 |
| Co2+ | 222 ± 18.8 | ND |
| Hg2+ | 13 ± 0.8 | 13 ± 1.4 |
| Ca2+ | 246 ± 9.3 | 261 ± 13.9 |
| K+ | 74 ± 5 | 126 ± 10.0 |
| EDTA | 154 ± 9.8 | 218 ± 32.3 |
| Solvent | Log POW | Relative Activity (%) a | ||
|---|---|---|---|---|
| 10% (v/v) | 20% (v/v) | |||
| Control | -- | 100 b | 100 | |
| DMSO | −1.35 | 47 ± 2.2 | 19 ± 0.9 | |
| Methanol | −0.76 | 9 ± 1.6 | 10 ± 0.5 | |
| Acetone | −0.24 | 30 ± 2.2 | 15 ± 2.4 | |
| Ethanol | −0.24 | 57 ± 8.8 | 29 ± 3.4 | |
| Acetonitrile | −0.34 | 15 ± 3.3 | 19 ± 3.5 | |
| Isopropyl | alcohol | 0.16 | 44 ± 6.3 | 24 ± 4.9 |
| Butanol | 0.8 | 12 ± 1.4 | 15 ± 0.3 | |
| Isoamyl | alcohol | 1.28 | 31 ± 4.6 | 54 ± 16.9 |
| Benzene | 2.13 | 79 ± 3.1 | 148 ± 30.2 | |
| Toluene | 2.4 | 88 ± 12 | 153 ± 69.0 | |
| Hexane | 3.5 | 115 ± 14.1 | 216 ± 20.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, H.; Xiao, Z.; Yu, K.; Zhang, Q.; Lu, C.; Wang, G.; Wang, Y.; Liang, J.; Huang, W.; Huang, X.; et al. High Diversity of β-Glucosidase-Producing Bacteria and Their Genes Associated with Scleractinian Corals. Int. J. Mol. Sci. 2021, 22, 3523. https://doi.org/10.3390/ijms22073523
Su H, Xiao Z, Yu K, Zhang Q, Lu C, Wang G, Wang Y, Liang J, Huang W, Huang X, et al. High Diversity of β-Glucosidase-Producing Bacteria and Their Genes Associated with Scleractinian Corals. International Journal of Molecular Sciences. 2021; 22(7):3523. https://doi.org/10.3390/ijms22073523
Chicago/Turabian StyleSu, Hongfei, Zhenlun Xiao, Kefu Yu, Qi Zhang, Chunrong Lu, Guanghua Wang, Yinghui Wang, Jiayuan Liang, Wen Huang, Xueyong Huang, and et al. 2021. "High Diversity of β-Glucosidase-Producing Bacteria and Their Genes Associated with Scleractinian Corals" International Journal of Molecular Sciences 22, no. 7: 3523. https://doi.org/10.3390/ijms22073523