
Environmental Enrichment Enhances Cav 2.1 Channel-Mediated Presynaptic Plasticity in Hypoxic–Ischemic Encephalopathy

 , ,
, ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
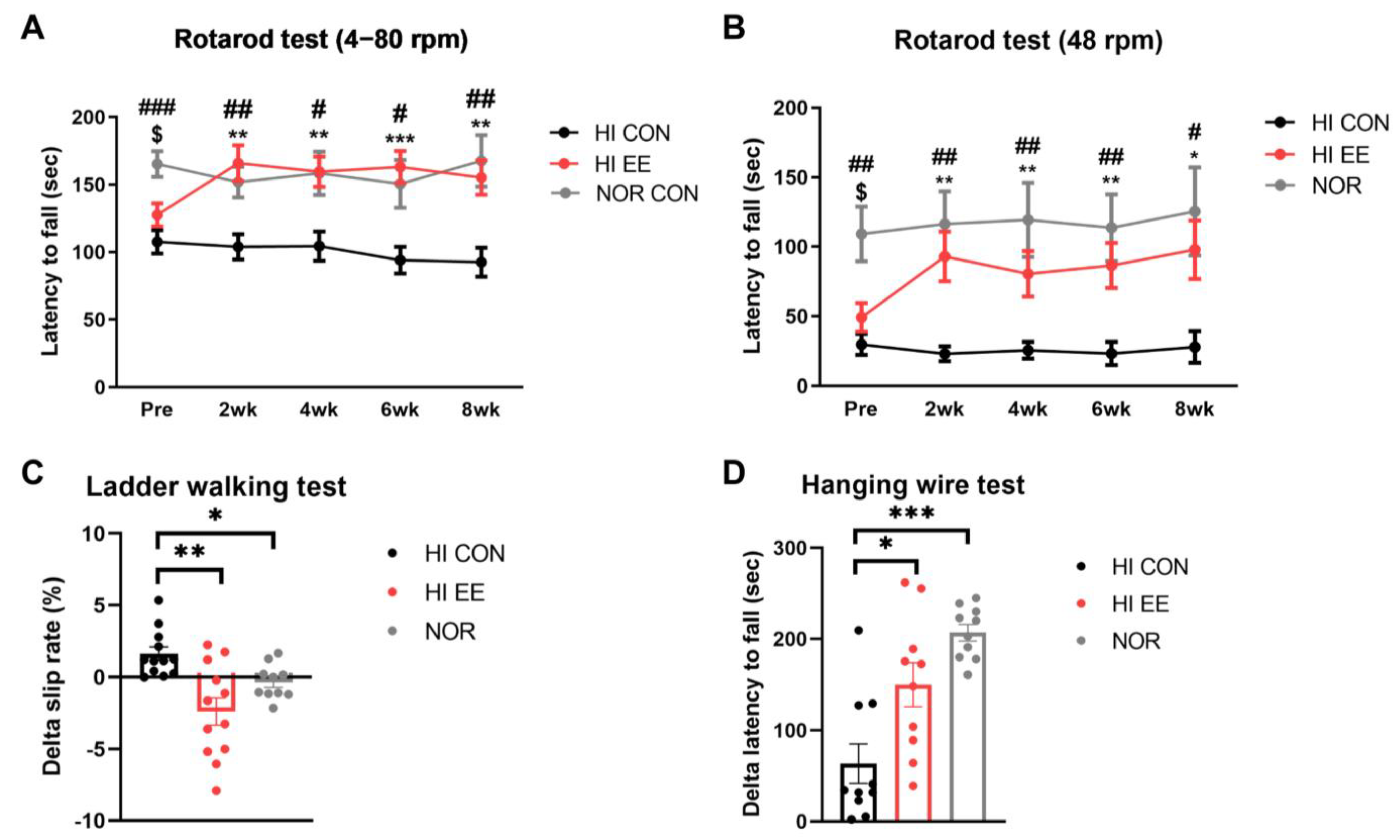
2.1. EE Improves Motor Coordination and Strength in HIE Mice
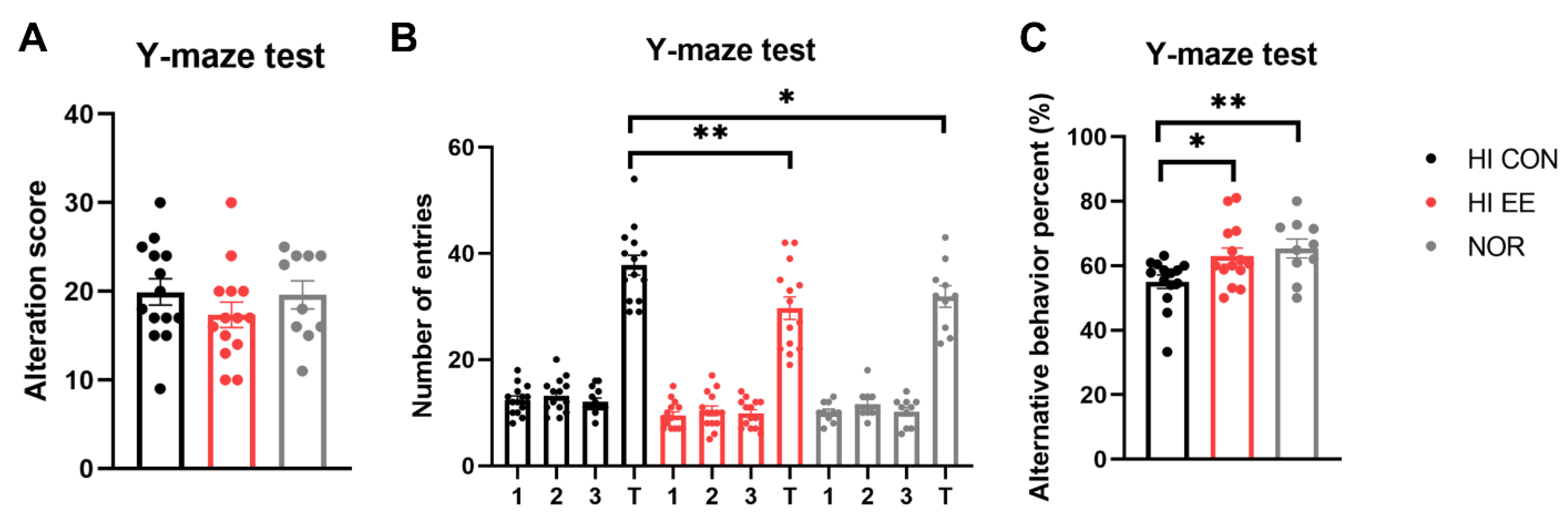
2.2. EE Improves Short-Term Spatial Memory in HIE Mice
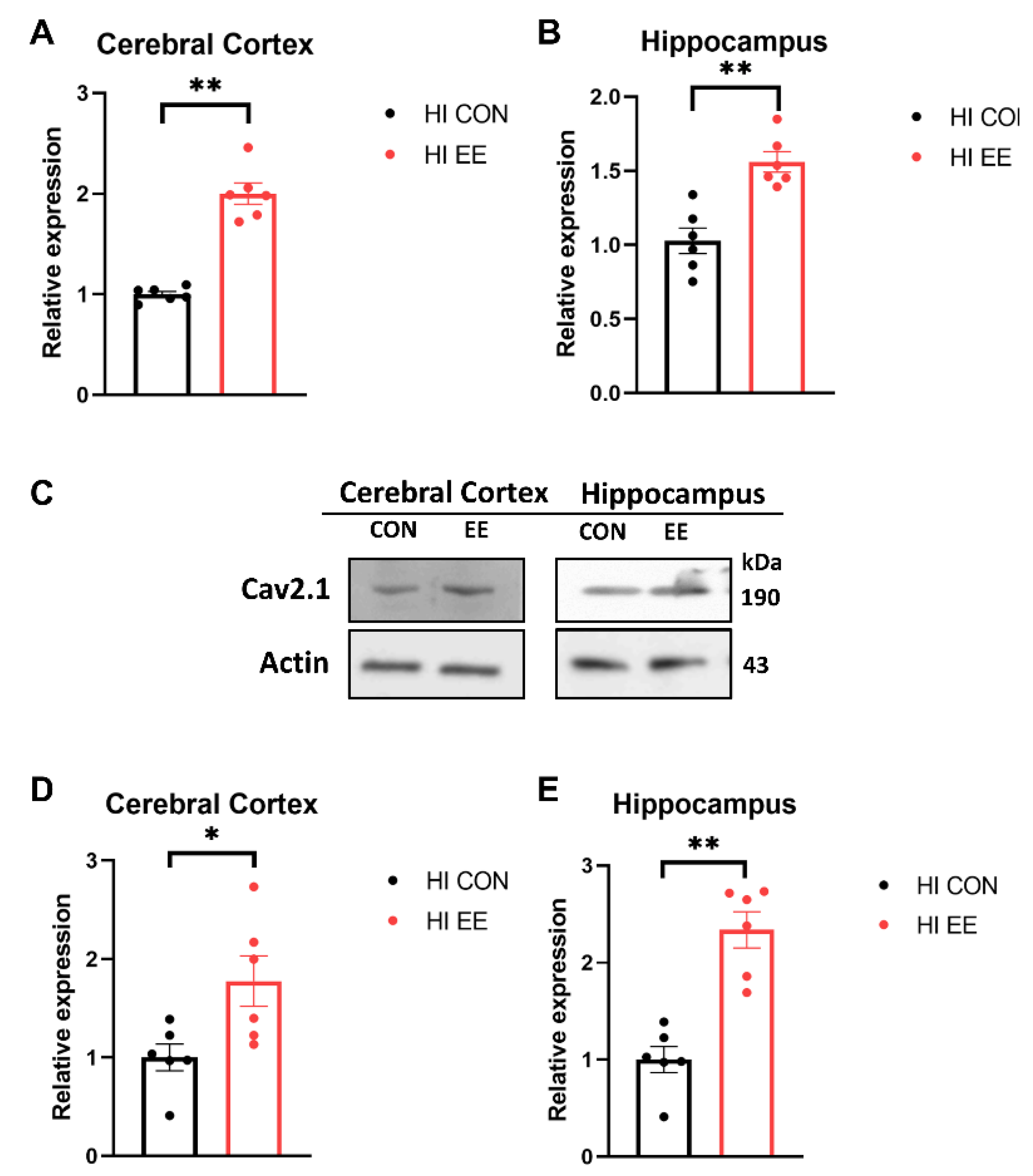
2.3. EE Upregulates the Expression of Cav 2.1 in the Cerebral Cortex and Hippocampus in HIE Mice
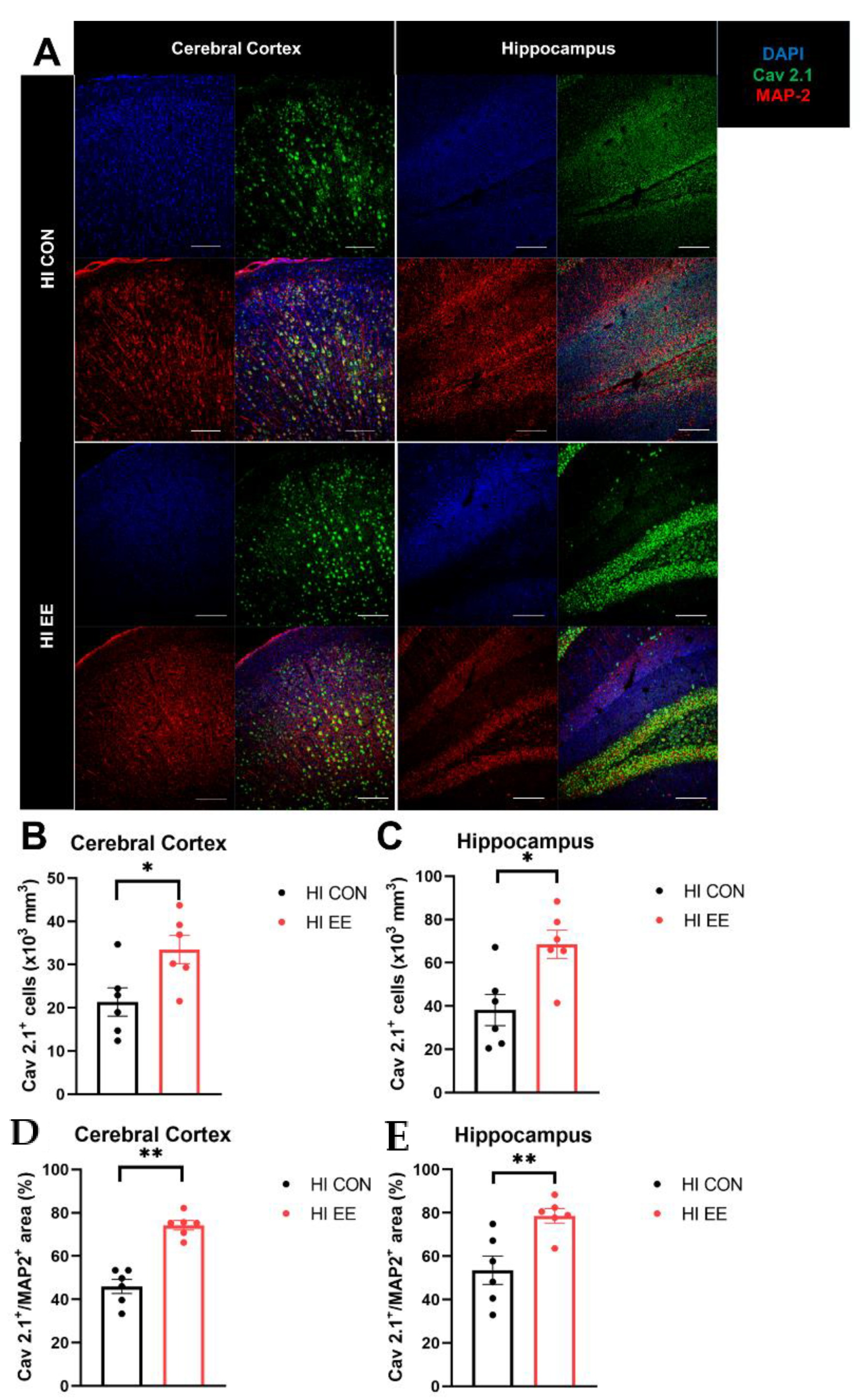
2.4. EE Induces Presynaptic Plasticity through the Higher Colocalization of Cav2.1 with MAP2 in the Cerebral Cortex and Hippocampus in HIE Mice
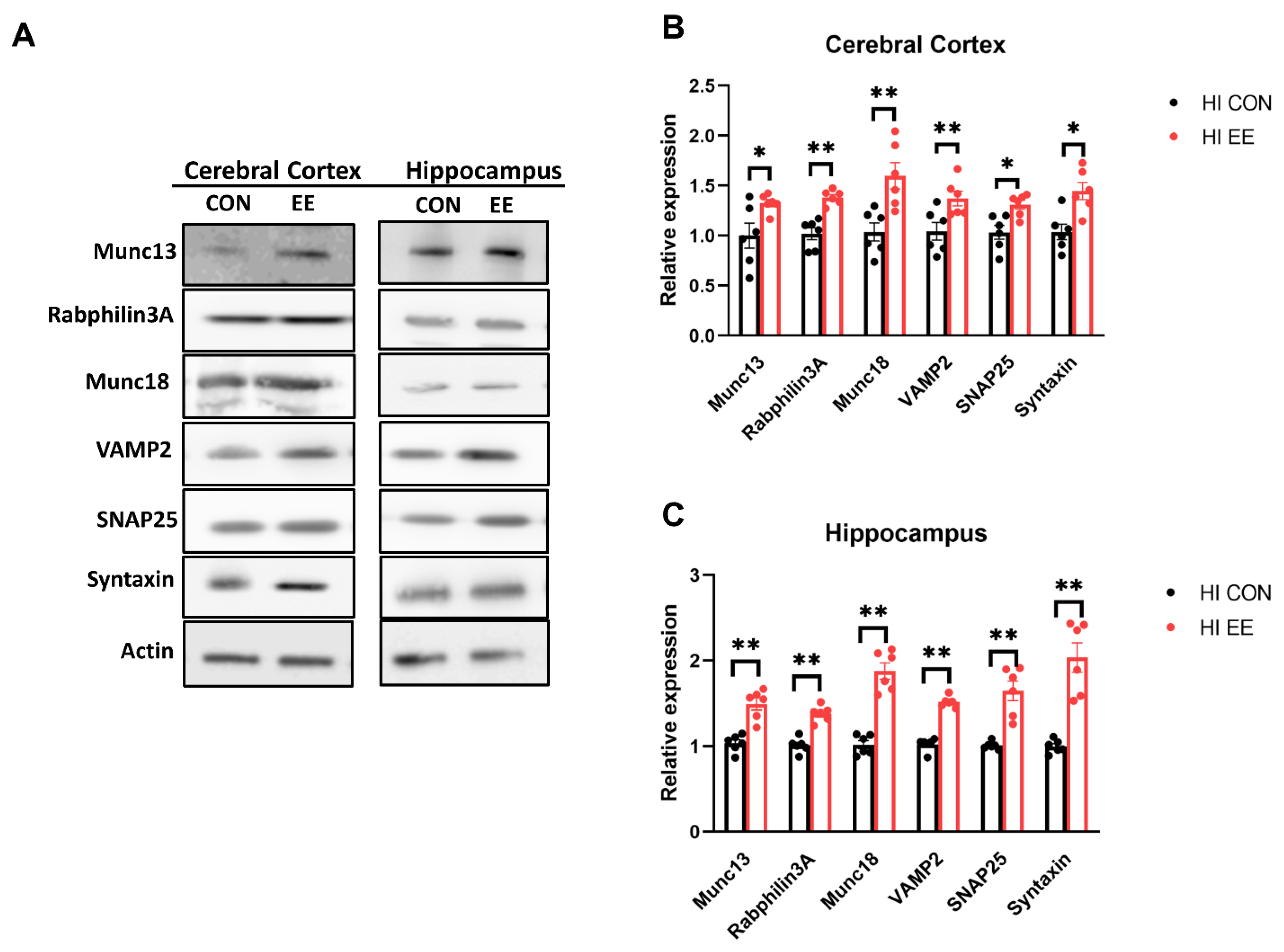
2.5. EE Upregulates the Expression of Presynaptic Proteins in the Cerebral Cortex and Hippocampus in HIE Mice
3. Discussion
4. Materials and Methods
4.1. Ethics Statement and Experimental Animals
4.2. Construction of Hypoxic–Ischemic Encephalopathy (HIE) Model
4.3. Experimental Procedures and Cage Condition
4.4. Behavioral Assessments
4.4.1. Rotarod Test
4.4.2. Ladder Walking Test
4.4.3. Hanging Wire Test
4.4.4. Y-Maze Test
4.5. Molecular Assessments
4.5.1. Quantitative Real-Time PCR (qRT-PCR)
4.5.2. Western Blot
4.6. Immunohistochemistry
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Allen, K.A.; Brandon, D.H. Hypoxic Ischemic Encephalopathy: Pathophysiology and Experimental Treatments. Newborn Infant Nurs. Rev. 2011, 11, 125–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, C.M.; Perlman, M. Follow-Up of the Term Infant after Hypoxic-Ischemic Encephalopathy. Paediatr. Child Health 2006, 11, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Thornton, C.; Leaw, B.; Mallard, C.; Nair, S.; Jinnai, M.; Hagberg, H. Cell Death in the Developing Brain after Hypoxia-Ischemia. Front. Cell. Neurosci. 2017, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Northington, F.J.; Chavez-Valdez, R.; Martin, L.J. Neuronal Cell Death in Neonatal Hypoxia-Ischemia. Ann. Neurol. 2011, 69, 743–758. [Google Scholar] [CrossRef] [PubMed]
- Reiss, J.; Sinha, M.; Gold, J.; Bykowski, J.; Lawrence, S.M. Outcomes of Infants with Mild Hypoxic Ischemic Encephalopathy Who Did Not Receive Therapeutic Hypothermia. Biomed. Hub 2019, 4, 1–9. [Google Scholar] [CrossRef]
- Kim, M.; Yu, J.H.; Seo, J.H.; Shin, Y.-K.; Wi, S.; Baek, A.; Song, S.-Y.; Cho, S.-R. Neurobehavioral Assessments in a Mouse Model of Neonatal Hypoxic-ischemic Brain Injury. J. Vis. Exp. 2017, e55838. [Google Scholar] [CrossRef]
- Hofmeijer, J.; Van Putten, M.J. Ischemic Cerebral Damage: An Appraisal of Synaptic Failure. Stroke 2012, 43, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Tong, X.; Huang, W.; Fu, J.; Xue, X. Synaptic Injury in the Thalamus Accompanies White Matter Injury in Hypoxia/Ischemia-Mediated Brain Injury in Neonatal Rats. BioMed Res. Int. 2019, 2019, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ten, V.S.; Bradley-Moore, M.; Gingrich, J.A.; Stark, R.I.; Pinsky, D.J. Brain Injury and Neurofunctional Deficit in Neonatal Mice with Hypoxic-Ischemic Encephalopathy. Behav. Brain Res. 2003, 145, 209–219. [Google Scholar] [CrossRef]
- Muntsant, A.; Shrivastava, K.; Recasens, M.; Giménez-Llort, L. Severe Perinatal Hypoxic-Ischemic Brain Injury Induces Long-Term Sensorimotor Deficits, Anxiety-Like Behaviors and Cognitive Impairment in a Sex-, Age- and Task-Selective Manner in C57BL/6 Mice but Can Be Modulated by Neonatal Handling. Front. Behav. Neurosci. 2019, 13, 7. [Google Scholar] [CrossRef]
- Berger, D.; Varriale, E.; Van Kessenich, L.M.; Herrmann, H.J.; De Arcangelis, L. Three Cooperative Mechanisms Required for Recovery after Brain Damage. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Calabresi, P.; Centonze, D.; Pisani, A.; Cupini, L.M.; Bernardi, G. Synaptic Plasticity in the Ischaemic Brain. Lancet Neurol. 2003, 2, 622–629. [Google Scholar] [CrossRef]
- Catterall, W.A.; Few, A.P. Calcium Channel Regulation and Presynaptic Plasticity. Neuron 2008, 59, 882–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezprozvanny, I.; Hiesinger, P.R. The Synaptic Maintenance Problem: Membrane Recycling, Ca2+ Homeostasis and Late Onset Degeneration. Mol. Neurodegener. 2013, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Yuan, H.; Yuan, F.; Yang, S.-H. Neuroprotection Targeting Ischemic Penumbra and Beyond for the Treatment of Ischemic Stroke. Neurol. Res. 2012, 34, 331–337. [Google Scholar] [CrossRef]
- Nie, J.; Yang, X. Modulation of Synaptic Plasticity by Exercise Training as a Basis for Ischemic Stroke Rehabilitation. Cell. Mol. Neurobiol. 2017, 37, 5–16. [Google Scholar] [CrossRef]
- Van Praag, H.; Kempermann, G.; Gage, F.H. Neural Consequences of Enviromental Enrichment. Nat. Rev. Neurosci. 2000, 1, 191–198. [Google Scholar] [CrossRef]
- Song, S.-Y.; Chae, M.; Yu, J.H.; Lee, M.Y.; Pyo, S.; Shin, Y.-K.; Baek, A.; Park, J.-W.; Park, E.S.; Choi, J.Y.; et al. Environmental Enrichment Upregulates Striatal Synaptic Vesicle-Associated Proteins and Improves Motor Function. Front. Neurol. 2018, 9, 465. [Google Scholar] [CrossRef] [Green Version]
- Rosbergen, I.C.; Grimley, R.S.; Hayward, K.S.; Brauer, S.G. The Impact of Environmental Enrichment in an Acute Stroke Unit on How and When Patients Undertake Activities. Clin. Rehabil. 2019, 33, 784–795. [Google Scholar] [CrossRef]
- McDonald, M.W.; Hayward, K.S.; Rosbergen, I.C.M.; Jeffers, M.S.; Corbett, D. Is Environmental Enrichment Ready for Clinical Application in Human Post-stroke Rehabilitation? Front. Behav. Neurosci. 2018, 12, 135. [Google Scholar] [CrossRef]
- Diaz, R.; Miguel, P.M.; Deniz, B.F.; Confortim, H.D.; Barbosa, S.; Mendonça, M.C.P.; Da Cruz-Höfling, M.A.; Pereira, L.O. Environmental Enrichment Attenuates the Blood Brain Barrier Dysfunction Induced by the Neonatal Hypoxia-Ischemia. Int. J. Dev. Neurosci. 2016, 53, 35–45. [Google Scholar] [CrossRef]
- Cechetti, F.; Worm, P.V.; Lovatel, G.; Moysés, F.; Siqueira, I.R.; Netto, C.A. Environmental Enrichment Prevents Behavioral Deficits and Oxidative Stress Caused by Chronic Cerebral Hypoperfusion in the Rat. Life Sci. 2012, 91, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Rojas, J.; Deniz, B.; Schuch, C.; Carletti, J.; Deckmann, I.; Diaz, R.; Matte, C.; Dos Santos, T.; Wyse, A.; Netto, C.; et al. Environmental Stimulation Improves Performance in the Ox-Maze Task and Recovers Na+,K+-ATPase Activity in the Hippocampus of Hypoxic–Ischemic rats. Neuroscience 2015, 291, 118–127. [Google Scholar] [CrossRef]
- Chae, S.Y.; Jang, J.H.; Im, G.H.; Jeong, J.-H.; Jung, W.-B.; Ko, S.; Jie, H.; Kim, J.H.; Chang, Y.S.; Chung, S.; et al. Physical Exercise Enhances Adult Cortical Plasticity in a Neonatal Rat Model of Hypoxic-Ischemic Injury: Evidence from BOLD-fMRI and Electrophysiological Recordings. NeuroImage 2019, 188, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Nudo, R.J. Recovery after Brain Injury: Mechanisms and Principles. Front. Hum. Neurosci. 2013, 7, 887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, W.-B.; Im, G.H.; Chung, J.J.; Ahn, S.-Y.; Jeon, T.Y.; Chang, Y.S.; Park, W.S.; Kim, J.H.; Kim, K.-S.; Lee, J.H. Neuroplasticity for Spontaneous Functional Recovery after Neonatal Hypoxic Ischemic Brain Injury in Rats Observed by Functional MRI and Diffusion Tensor Imaging. NeuroImage 2016, 126, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Ferreira, E.; Hristova, M. Plasticity in the Neonatal Brain following Hypoxic-Ischaemic Injury. Neural Plast. 2016, 2016, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Vive, S.; Bunketorp-Käll, L.; Carlsson, G. Experience of Enriched Rehabilitation in the Chronic Phase of Stroke. Disabil. Rehabil. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- De Boer, A.; Storm, A.; Soler, M.G.; Smolders, S.; Rué, L.; Poppe, L.; Pasquale, E.B.; Robberecht, W.; Lemmens, R. Environmental Enrichment during the Chronic Phase after Experimental Stroke Promotes Functional Recovery without Synergistic Effects of EphA4 Targeted Therapy. Hum. Mol. Genet. 2019, 29, 605–617. [Google Scholar] [CrossRef] [Green Version]
- Vive, S.; Geijerstam, J.-L.A.; Kuhn, H.G.; Bunketorp-Käll, L. Enriched, Task-Specific Therapy in the Chronic Phase After Stroke: An Exploratory Study. J. Neurol. Phys. Ther. 2020, 44, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Südhof, T.C. The Synaptic Vesicle Cycle. Annu. Rev. Neurosci. 2004, 27, 509–547. [Google Scholar] [CrossRef] [Green Version]
- Sakane, A.; Manabe, S.; Ishizaki, H.; Tanaka-Okamoto, M.; Kiyokage, E.; Toida, K.; Yoshida, T.; Miyoshi, J.; Kamiya, H.; Takai, Y.; et al. Rab3 GTPase-Activating Protein Regulates Synaptic Transmission and Plasticity through the Inactivation of Rab3. Proc. Natl. Acad. Sci. USA 2006, 103, 10029–10034. [Google Scholar] [CrossRef] [Green Version]
- Kaeser, P.S.; Kwon, H.-B.; Blundell, J.; Chevaleyre, V.; Morishita, W.; Malenka, R.C.; Powell, C.M.; Castillo, P.E.; Südhof, T.C. RIM1 Phosphorylation at Serine-413 by Protein Kinase A is not Required for Presynaptic Long-Term Plasticity or Learning. Proc. Natl. Acad. Sci. USA 2008, 105, 14680–14685. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Gong, J.; Zhu, L.; Wang, S.; Yang, X.; Xu, Y.; Yang, X.; Ma, C. Munc13 Activates the Munc18-1/Syntaxin-1 Complex and Enables Munc18-1 to Prime SNARE Assembly. EMBO J. 2020, 39, e103631. [Google Scholar] [CrossRef]
- Yang, X.; Wang, S.; Sheng, Y.; Zhang, M.; Zou, W.; Wu, L.; Kang, L.; Rizo, J.; Zhang, R.; Xu, T.; et al. Syntaxin Opening by the MUN Domain Underlies the Function of Munc13 in Synaptic-Vesicle Priming. Nat. Struct. Mol. Biol. 2015, 22, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Shu, T.; Jin, H.; Rothman, J.E.; Zhang, Y. Munc13-1 MUN Domain and Munc18-1 Cooperatively Chaperone SNARE Assembly through a Tetrameric Complex. Proc. Natl. Acad. Sci. USA 2020, 117, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Lipstein, N.; Sakaba, T.; Cooper, B.H.; Lin, K.-H.; Strenzke, N.; Ashery, U.; Rhee, J.-S.; Taschenberger, H.; Neher, E.; Brose, N. Dynamic Control of Synaptic Vesicle Replenishment and Short-Term Plasticity by Ca2+-Calmodulin-Munc13-1 Signaling. Neuron 2013, 79, 82–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genc, O.; Kochubey, O.; Toonen, R.F.; Verhage, M.; Schneggenburger, R. Munc18-1 is a Dynamically Regulated PKC Target during Short-Term Enhancement of Transmitter Release. Elife 2014, 3, e01715. [Google Scholar] [CrossRef] [Green Version]
- Toonen, R.F.G.; Wierda, K.; Sons, M.S.; De Wit, H.; Cornelisse, L.N.; Brussaard, A.; Plomp, J.J.; Verhage, M. Munc18-1 Expression Levels Control Synapse Recovery by Regulating Readily Releasable Pool Size. Proc. Natl. Acad. Sci. USA 2006, 103, 18332–18337. [Google Scholar] [CrossRef] [Green Version]
- Jahn, R.; Fasshauer, D. Molecular Machines Governing Exocytosis of Synaptic Vesicles. Nature 2012, 490, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Südhof, T.C.; Rothman, J.E. Membrane Fusion: Grappling with SNARE and SM Proteins. Science 2009, 323, 474–477. [Google Scholar] [CrossRef] [Green Version]
- Bonifacino, J.S.; Glick, B.S. The Mechanisms of Vesicle Budding and Fusion. Cell 2004, 116, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Araújo, S.J.; Ulloa, F.; Cotrufo, T.; Ricolo, D.; Soriano, E. Snare Complex in Axonal Guidance and Neuroregeneration. Neural Regen. Res. 2018, 13, 386–392. [Google Scholar] [CrossRef]
- Bloom, O.E.; Morgan, J.R. Membrane Trafficking Events Underlying Axon Repair, Growth, and Regeneration. Mol. Cell. Neurosci. 2011, 48, 339–348. [Google Scholar] [CrossRef]
- Lübbert, M.; Goral, R.O.; Keine, C.; Thomas, C.; Guerrero-Given, D.; Putzke, T.; Satterfield, R.; Kamasawa, N.; Young, S.M., Jr. CaV2.1 α1 Subunit Expression Regulates Presynaptic CaV2.1 Abundance and Synaptic Strength at a Central Synapse. Neuron 2019, 101, 260–273. [Google Scholar] [CrossRef]
- Nanou, E.; Sullivan, J.M.; Scheuer, T.; Catterall, W.A. Calcium Sensor Regulation of the CaV2.1 Ca2+ Channel Contributes to Short-Term Synaptic Plasticity in Hippocampal Neurons. Proc. Natl. Acad. Sci. USA 2016, 113, 1062–1067. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Leal, K.; Magupalli, V.G.; Nanou, E.; Martinez, G.Q.; Scheuer, T.; Catterall, W.A. Modulation of CaV2.1 Channels by Neuronal Calcium Sensor-1 Induces Short-Term Synaptic Facilitation. Mol. Cell. Neurosci. 2014, 63, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Nanou, E.; Scheuer, T.; Catterall, W.A. Calcium Sensor Regulation of the CaV2.1 Ca2+ Channel Contributes to Long-Term Potentiation and Spatial Learning. Proc. Natl. Acad. Sci. USA 2016, 113, 13209–13214. [Google Scholar] [CrossRef] [Green Version]
- Anderson, B.J.; Rapp, D.N.; Baek, D.H.; McCloskey, D.P.; Coburn-Litvak, P.S.; Robinson, J.K. Exercise Influences Spatial Learning in the Radial Arm Maze. Physiol. Behav. 2000, 70, 425–429. [Google Scholar] [CrossRef]
- McQuaid, R.J.; Dunn, R.; Jacobson-Pick, S.; Anisman, H.; Audet, M.-C. Post-Weaning Environmental Enrichment in Male CD-1 Mice: Impact on Social Behaviors, Corticosterone Levels and Prefrontal Cytokine Expression in Adulthood. Front. Behav. Neurosci. 2018, 12, 145. [Google Scholar] [CrossRef]
- Leuner, B.; Gould, E. Structural Plasticity and Hippocampal Function. Annu. Rev. Psychol. 2010, 61, 111–140. [Google Scholar] [CrossRef] [Green Version]
- Kitabatake, Y.; Sailor, K.A.; Ming, G.-L.; Song, H. Adult Neurogenesis and Hippocampal Memory Function: New Cells, More Plasticity, New Memories? Neurosurg. Clin. North Am. 2007, 18, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Wang, X. Selective Neuronal Vulnerability to Oxidative Stress in the Brain. Front. Aging Neurosci. 2010, 2, 12. [Google Scholar] [CrossRef]
- Nikonenko, A.G.; Radenovic, L.; Andjus, P.R.; Skibo, G.G. Structural Features of Ischemic Damage in the Hippocampus. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2009, 292, 1914–1921. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, C.-J.; Zhang, Q.; Yu, K.-W.; Wang, Y.-Y. An Enriched Environment Promotes Synaptic Plasticity and Cognitive Recovery after Permanent Middle Cerebral Artery Occlusion in Mice. Neural Regen. Res. 2019, 14, 462–469. [Google Scholar] [CrossRef]
- Lee, M.-Y.; Yu, J.H.; Kim, J.Y.; Seo, J.H.; Park, E.S.; Kim, C.H.; Kim, H.; Cho, S.-R. Alteration of Synaptic Activity–Regulating Genes Underlying Functional Improvement by Long-term Exposure to an Enriched Environment in the Adult Brain. Neurorehabilit Neural Repair 2013, 27, 561–574. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Li, M.-Y.; Zhang, X.; Jin, X.; Liu, J.; Wei, P.-H. Delayed Exposure to Environmental Enrichment Improves Functional Outcome after Stroke. J. Pharmacol. Sci. 2019, 140, 137–143. [Google Scholar] [CrossRef]
- Hong, M.; Kim, M.; Kim, T.-W.; Park, S.-S.; Kim, M.-K.; Park, Y.H.; Sung, Y.-H.; Shin, M.-S. Treadmill Exercise Improves Motor Function and Short-term Memory by Enhancing Synaptic Plasticity and Neurogenesis in Photothrombotic Stroke Mice. Int. Neurourol. J. 2020, 24, S28–S38. [Google Scholar] [CrossRef]
- Catterall, W.A. Voltage-Gated Calcium Channels. Cold Spring Harb. Perspect. Biol. 2011, 3, a003947. [Google Scholar] [CrossRef]
- Simms, B.A.; Zamponi, G.W. Neuronal Voltage-Gated Calcium Channels: Structure, Function, and Dysfunction. Neuron 2014, 82, 24–45. [Google Scholar] [CrossRef] [Green Version]
- Lübbert, M.; Goral, R.O.; Satterfield, R.; Putzke, T.; van den Maagdenberg, A.M.; Kamasawa, N.; Young, S.M., Jr. A Novel Region in the CaV2.1 α1 Subunit C-Terminus Regulates Fast Synaptic Vesicle Fusion and Vesicle Docking at the Mammalian Presynaptic Active Zone. Elife 2017, 6, e28412. [Google Scholar] [CrossRef] [Green Version]
- Heck, J.; Parutto, P.; Ciuraszkiewicz, A.; Bikbaev, A.; Freund, R.; Mitlöhner, J.; Andres-Alonso, M.; Fejtova, A.; Holcman, D.; Heine, M. Transient Confinement of CaV2.1 Ca2+-Channel Splice Variants Shapes Synaptic Short-Term Plasticity. Neuron 2019, 103, 66–79.e12. [Google Scholar] [CrossRef]
- Leal, K.; Mochida, S.; Scheuer, T.; Catterall, W.A. Fine-Tuning Synaptic Plasticity by Modulation of CaV2.1 Channels with Ca2+ Sensor Proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 17069–17074. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Kutner, M.; Nachmanni, D.; Atlas, D. CaV2.1 (P/Q channel) Interaction with Synaptic Proteins is Essential for Depolarization-Evoked Release. Channels 2010, 4, 266–277. [Google Scholar] [CrossRef] [Green Version]
- Gandini, M.A.; Souza, I.A.; Fan, J.; Li, K.; Wang, D.; Zamponi, G.W. Interactions of Rabconnectin-3 with Cav2 Calcium Channels. Mol. Brain 2019, 12, 1–7. [Google Scholar] [CrossRef]
- Dolphin, A.C. Functions of Presynaptic Voltage-gated Calcium Channels. Function 2020, 2, zqaa027. [Google Scholar] [CrossRef]
- Chung, J.-W.; Ryu, W.-S.; Kim, B.J.; Yoon, B.-W. Elevated Calcium after Acute Ischemic Stroke: Association with a Poor Short-Term Outcome and Long-Term Mortality. J. Stroke 2015, 17, 54–59. [Google Scholar] [CrossRef]
- Kim, S.Y.; Hsu, J.E.; Husbands, L.C.; Kleim, J.A.; Jones, T.A. Coordinated Plasticity of Synapses and Astrocytes Underlies Practice-Driven Functional Vicariation in Peri-Infarct Motor Cortex. J. Neurosci. 2018, 38, 93–107. [Google Scholar] [CrossRef]
- Jones, T.A.; Adkins, D.L. Motor System Reorganization After Stroke: Stimulating and Training Toward Perfection. Physiology 2015, 30, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Hatem, S.M.; Saussez, G.; Della Faille, M.; Prist, V.; Zhang, X.; Dispa, D.; Bleyenheuft, Y. Rehabilitation of Motor Function after Stroke: A Multiple Systematic Review Focused on Techniques to Stimulate Upper Extremity Recovery. Front. Hum. Neurosci. 2016, 10, 442. [Google Scholar] [CrossRef] [Green Version]
- Ballester, B.R.; Maier, M.; Duff, A.; Cameirão, M.; Bermúdez, S.; Duarte, E.; Cuxart, A.; Rodríguez, S.; Mozo, R.M.S.S.; Verschure, P.F.M.J. A Critical Time Window for Recovery Extends beyond One-Year Post-Stroke. J. Neurophysiol. 2019, 122, 350–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.-Y.L.; Kim, S.W.; Kim, J.H.; Park, C.K. Increased Levels of Synaptic Proteins Involved in Synaptic Plasticity after Chronic Intraocular Pressure Elevation and Modulation by Brain-Derived Neurotrophic Factor in a Glaucoma Animal Model. Dis. Model. Mech. 2019, 12, dmm037184. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, T.; Gal-Ben-Ari, S.; Dieterich, D.C.; Kreutz, M.R.; Ziv, N.E.; Gundelfinger, E.D.; Rosenblum, K. The Roles of Protein Expression in Synaptic Plasticity and Memory Consolidation. Front. Mol. Neurosci. 2014, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Torres, D.M.C.; Cardenas, F.P. Synaptic Plasticity in Alzheimer’s Disease and Healthy Aging. Rev. Neurosci. 2020, 31, 245–268. [Google Scholar] [CrossRef]
- Gutierrez, R.M.S.; Real, C.C.; Scaranzi, C.R.; Garcia, P.C.; Oliveira, D.L.; Britto, L.R.; Pires, R.S. Motor Improvement Requires an Increase in Presynaptic Protein Expression and Depends on Exercise Type and Age. Exp. Gerontol. 2018, 113, 18–28. [Google Scholar] [CrossRef]
- Grant, S.G.N. Synapse Molecular Complexity and the Plasticity Behaviour Problem. Brain Neurosci. Adv. 2018, 2, 1–7. [Google Scholar] [CrossRef]
- Metz, G.A.; Whishaw, I.Q. The Ladder Rung Walking Task: A Scoring System and its Practical Application. J. Vis. Exp. 2009, e1204. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, E.; Winder, S.J. A Modified Wire Hanging Apparatus for Small Animal Muscle Function Testing. PLoS Curr. 2016, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Sarnyai, Z.; Sibille, E.L.; Pavlides, C.; Fenster, R.J.; McEwen, B.S.; Tóth, M. Impaired Hippocampal-Dependent Learning and Functional Abnormalities in the Hippocampus in Mice Lacking Serotonin1a Receptors. Proc. Natl. Acad. Sci. USA 2000, 97, 14731–14736. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, S.-Y.; Pyo, S.; Choi, S.; Oh, H.S.; Seo, J.H.; Yu, J.H.; Baek, A.; Shin, Y.-K.; Lee, H.Y.; Choi, J.Y.; et al. Environmental Enrichment Enhances Cav 2.1 Channel-Mediated Presynaptic Plasticity in Hypoxic–Ischemic Encephalopathy. Int. J. Mol. Sci. 2021, 22, 3414. https://doi.org/10.3390/ijms22073414
Song S-Y, Pyo S, Choi S, Oh HS, Seo JH, Yu JH, Baek A, Shin Y-K, Lee HY, Choi JY, et al. Environmental Enrichment Enhances Cav 2.1 Channel-Mediated Presynaptic Plasticity in Hypoxic–Ischemic Encephalopathy. International Journal of Molecular Sciences. 2021; 22(7):3414. https://doi.org/10.3390/ijms22073414
Chicago/Turabian StyleSong, Suk-Young, Soonil Pyo, Sungchul Choi, Hee Sang Oh, Jung Hwa Seo, Ji Hea Yu, Ahreum Baek, Yoon-Kyum Shin, Hoo Young Lee, Ja Young Choi, and et al. 2021. "Environmental Enrichment Enhances Cav 2.1 Channel-Mediated Presynaptic Plasticity in Hypoxic–Ischemic Encephalopathy" International Journal of Molecular Sciences 22, no. 7: 3414. https://doi.org/10.3390/ijms22073414