
Granulibacter bethesdensis, a Pathogen from Patients with Chronic Granulomatous Disease, Produces a Penta-Acylated Hypostimulatory Glycero-D-talo-oct-2-ulosonic Acid–Lipid A Glycolipid (Ko-Lipid A)

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
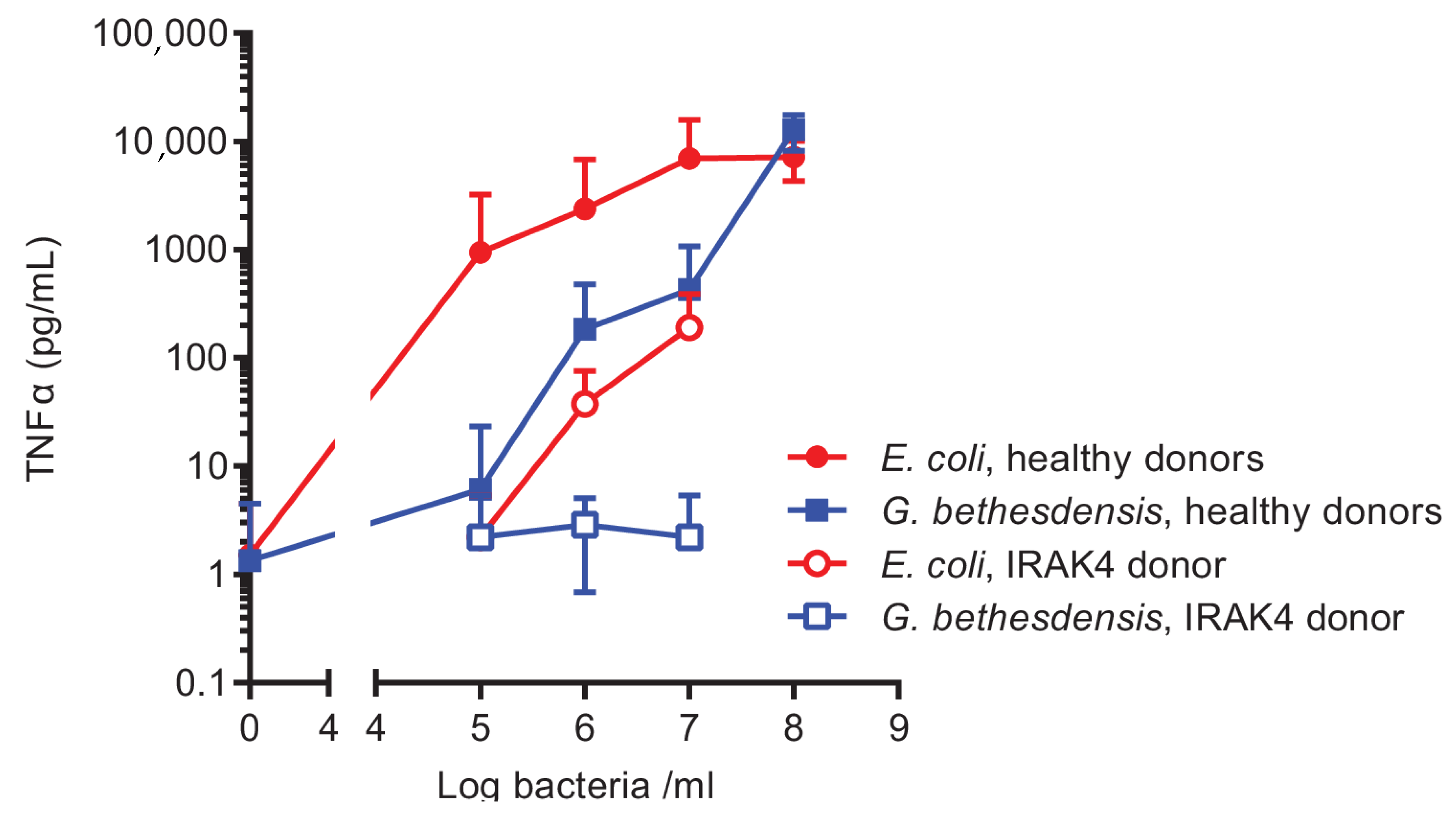
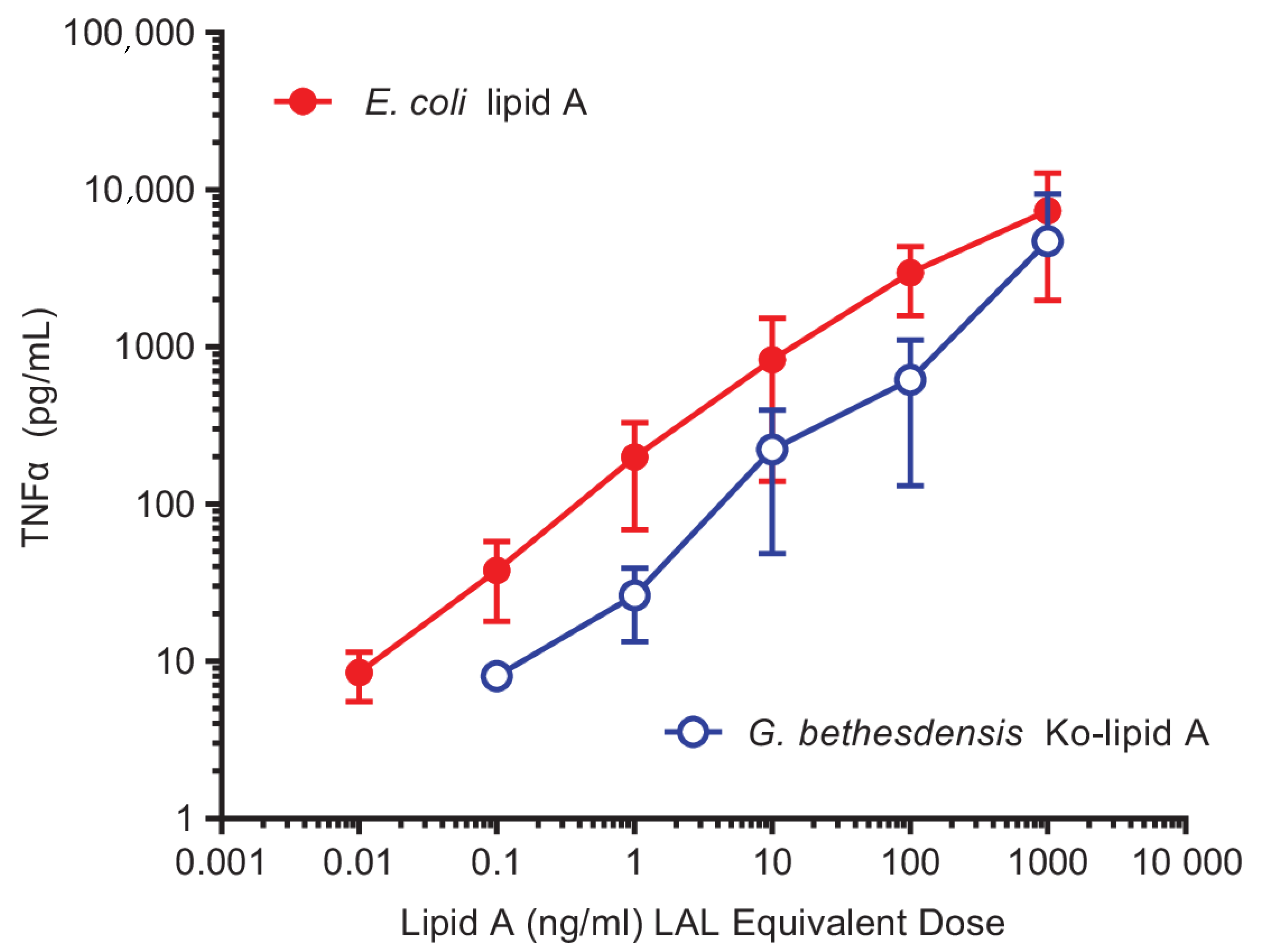
2.1. Hypostimulatory Effect of G. bethesdensis on Human Blood Cells
2.2. Characterization of the G. bethesdensis LPS
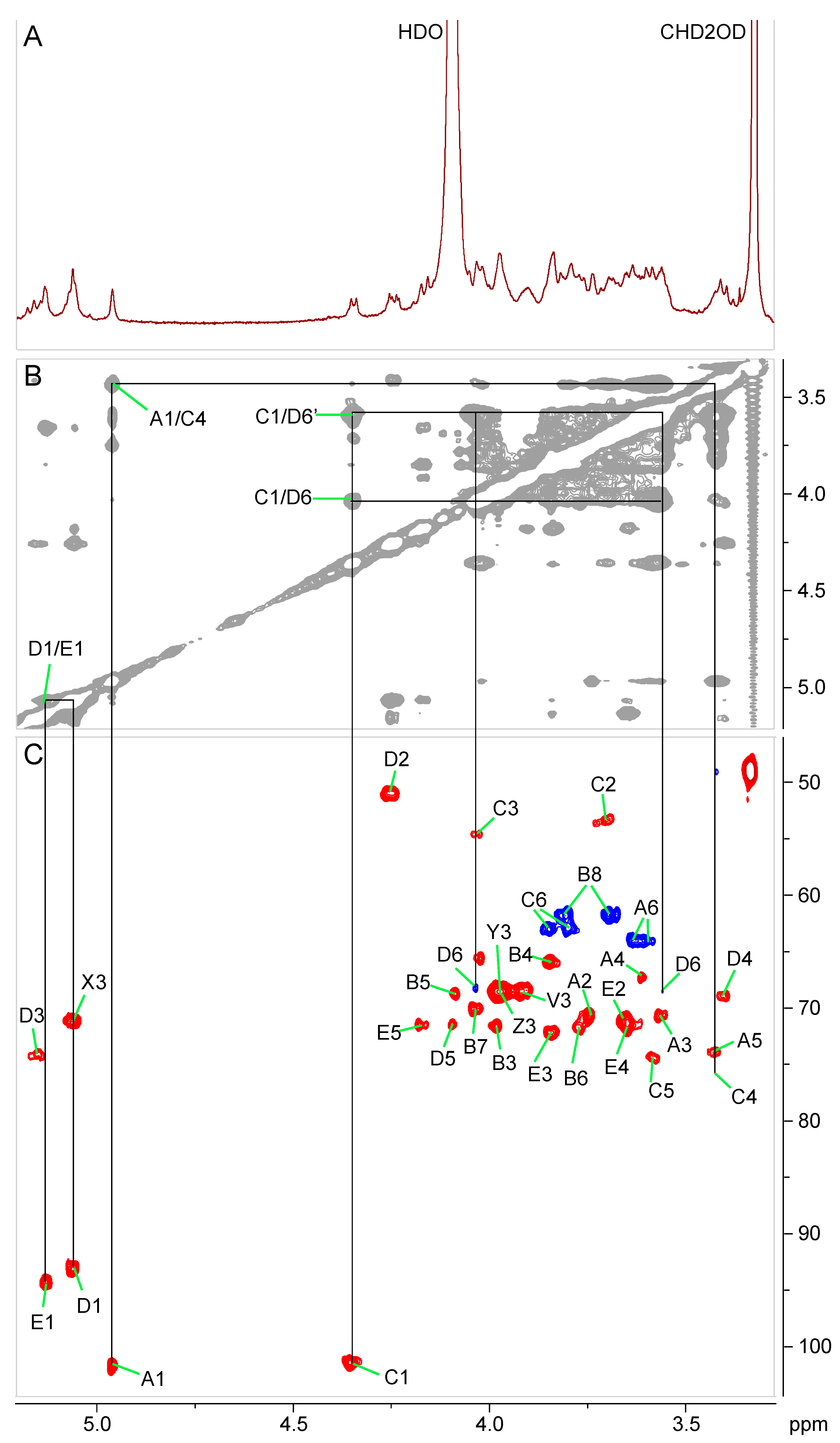
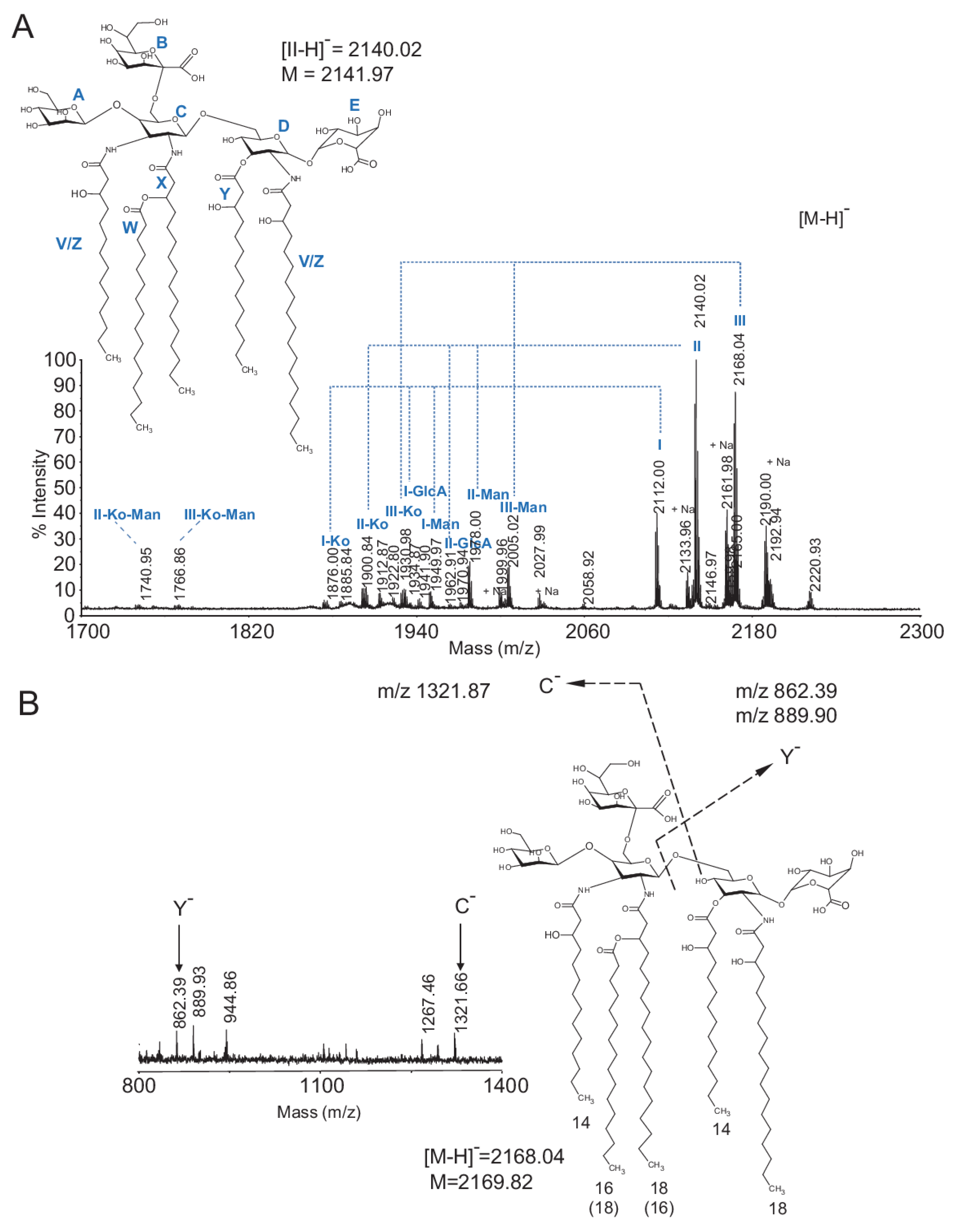
2.3. Chemical and Structural Characterization of Ko-Containing Lipid A Glycosyl Substituents
| α-Manp-(1→4)(α-Kop-(2→6))-β-GlcpN3N-(1→6)-α-GlcpN-(1↔1)-α-GlcpA |
| A B C D E |
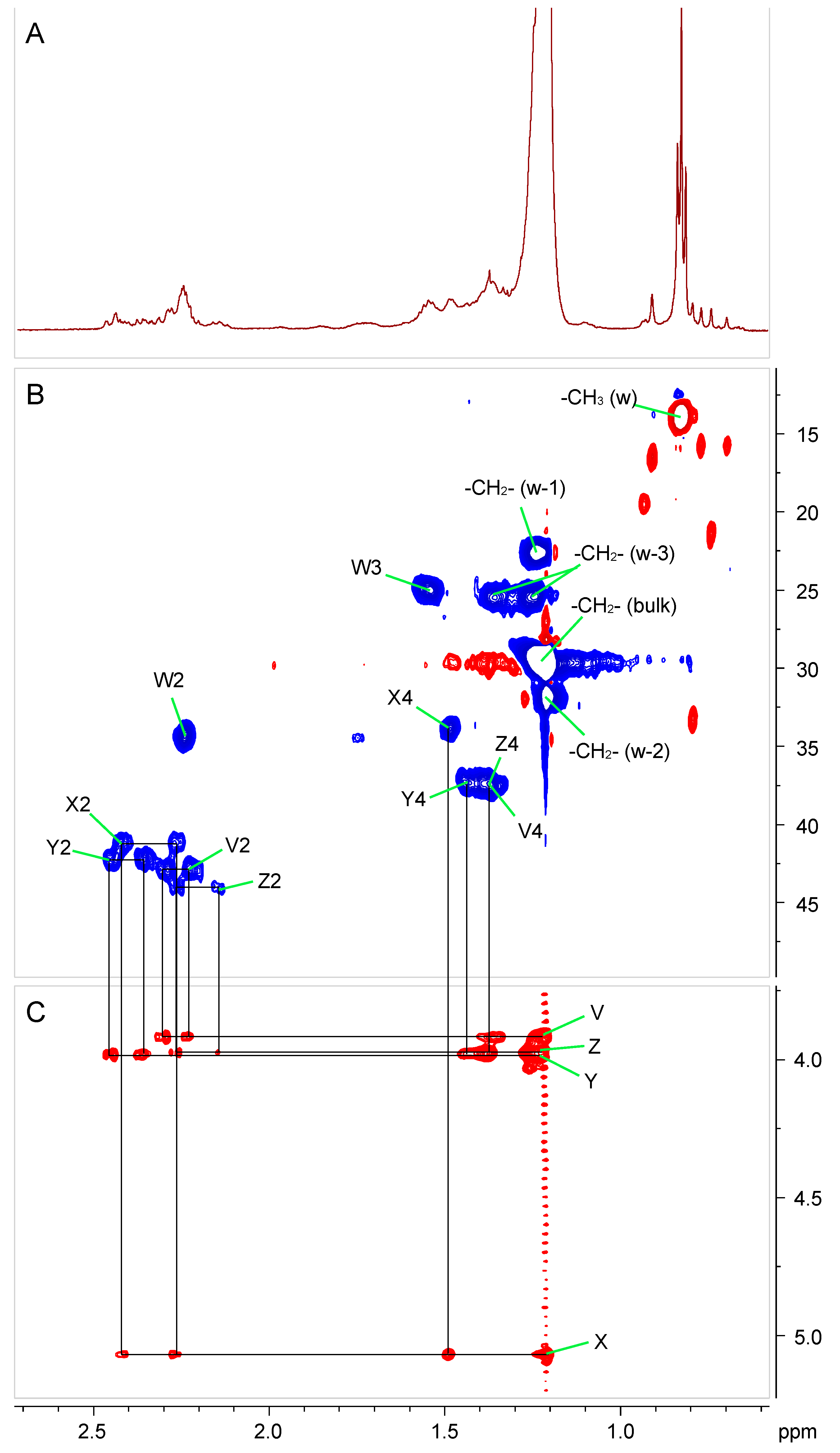
2.4. Fatty Acid Substituents of Ko–Lipid A
2.5. Induction of Cytokine Production by Purified Granulibacter Ko–Lipid A in Human Whole Blood
3. Discussion
4. Materials and Methods
4.1. Bacteria
4.2. Extraction and Purification of LPS
4.3. Isolation of Gb Ko–Lipid A Glycolipid
4.4. Compositional Analysis of LPS
4.5. Analysis of O-deacylated Lipid A
4.6. MS Analysis of Lipid A
4.7. Nuclear Magnetic Resonance (NMR) Spectroscopy
4.8. Gel Electrophoresis and Staining
4.9. Endotoxin Measurement
4.10. Whole-Blood Cytokine Assay
4.11. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Alauzet, C.; Teyssier, C.; Jumas-Bilak, E.; Gouby, A.; Chiron, R.; Rabaud, C.; Counil, F.; Lozniewski, A.; Marchandin, H. Gluconobacter as Well as Asaia species, newly emerging opportunistic human pathogens among acetic acid bacteria. J. Clin. Microbiol. 2010, 48, 3935–3942. [Google Scholar] [CrossRef] [Green Version]
- Falcone, E.L.; Petts, J.R.; Fasano, M.B.; Ford, B.; Nauseef, W.M.; Neves, J.F.; Simões, M.J.; Tierce, M.L.; De La Morena, M.T.; Greenberg, D.E.; et al. Methylotroph infections and chronic granulomatous disease. Emerg. Infect. Dis. 2016, 22, 404–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouby, A.; Teyssier, C.; Vecina, F.; Marchandin, H.; Granolleras, C.; Zorgniotti, I.; Jumas-Bilak, E. Acetobacter cibinongensis Bacteremia in human. Emerg. Infect. Dis. 2007, 13, 784–785. [Google Scholar] [CrossRef]
- Greenberg, D.E.; Ding, L.; Zelazny, A.M.; Stock, F.; Wong, A.; Anderson, V.L.; Miller, G.; Kleiner, D.E.; Tenorio, A.R.; Brinster, L.; et al. A novel bacterium associated with lymphadenitis in a patient with chronic granulomatous disease. PLoS Pathog. 2006, 2, e28. [Google Scholar] [CrossRef]
- Rihs, J.D.; Brenner, D.J.; Weaver, R.E.; Steigerwalt, A.G.; Hollis, D.G.; Yu, V.L. Roseomonas, a new genus associated with bacteremia and other human infections. J. Clin. Microbiol. 1993, 31, 3275–3283. [Google Scholar] [CrossRef] [Green Version]
- Tuuminen, T.; Heinäsmäki, T.; Kerttula, T. First Report of bacteremia by Asaia bogorensis, in a patient with a history of intravenous-drug abuse. J. Clin. Microbiol. 2006, 44, 3048–3050. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, D.E.; Porcella, S.F.; Stock, F.; Wong, A.; Conville, P.S.; Murray, P.R.; Holland, S.M.; Zelazny, A.M. Granulibacter bethesdensis gen. nov., sp. nov., a distinctive pathogenic acetic acid bacterium in the family Acetobacteraceae. Int. J. Syst. Evol. Microbiol. 2006, 56, 2609–2616. [Google Scholar] [CrossRef] [PubMed]
- Zarember, K.A.; Soule, B.P.; Gallin, J.I. Chronic granulomatous disease: From lethal pediatric mystery to complex chronic disease. In National Institute of Allergy and Infectious Diseases, NIH; Georgiev, V.S., Ed.; Humana Press: Totowa, NJ, USA, 2010; Volume 3, pp. 319–352. [Google Scholar]
- Mayer, E.F.; Gialanella, P.; Munjal, I.; Cunningham-Rundles, C.; Dara, J. Fulminant Sepsis Due to Granulibacter bethesdensis in a 4-year-old boy with X-linked chronic granulomatous disease. Pediatr. Infect. Dis. J. 2017, 36, 1165–1166. [Google Scholar] [CrossRef]
- Greenberg, D.E.; Shoffner, A.R.; Marshall-Batty, K.R.; Arora, K.; Zhao, M.; Martin, R.; Ding, L.; Hammer, C.H.; Shaw, P.A.; Kuhns, U.B.; et al. Serologic reactivity to the emerging pathogen Granulibacter bethesdensis. J. Infect. Dis. 2012, 206, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.; Song, H.H.; Zarember, K.A.; Mills, T.A.; Gallin, J.I. Persistence of the bacterial pathogen Granulibacter bethesdensisin chronic granulomatous disease monocytes and macrophages lacking a functional NADPH oxidase. J. Immunol. 2013, 191, 3297–3307. [Google Scholar] [CrossRef] [Green Version]
- Zarember, K.A.; Marshall-Batty, K.R.; Cruz, A.R.; Chu, J.; Fenster, M.E.; Shoffner, A.R.; Rogge, L.S.; Whitney, A.R.; Czapiga, M.; Song, H.H.; et al. Innate immunity against Granulibacter bethesdensis, an emerging gram-negative bacterial pathogen. Infect. Immun. 2011, 80, 975–981. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.; Smelkinson, M.G.; Dorward, D.W.; Zarember, K.A.; Gallin, J.I. Early intracellular trafficking of Granulibacter bethesdensis in human macrophages. Infect. Immun. 2017, 85, e00847-16. [Google Scholar] [CrossRef] [Green Version]
- Bera, R.; Nayak, A.; Sen, A.K.; Chowdhury, B.P.; Bhadra, R. Isolation and characterisation of the lipopolysaccharide from Acidiphiliumstrain GS18h/ATCC55963, a soil isolate of Indian copper mine. FEMS Microbiol. Lett. 2005, 246, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Bera, R.; Nayak, A.; Sarkar, C.; Singh, S.K.; Ratha, J.; Bhadra, R. Immunobiological activities of a new nontoxic lipopolysaccharide fromAcidiphiliumGS18h/ATCC55963, a soil isolate from an Indian copper mine. FEMS Immunol. Med. Microbiol. 2006, 48, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, Y.; Nishizawa, T.; Kouhchi, C.; Inagawa, H.; Yamaguchi, T.; Nagai, S.; Tamura, A.; Soma, G.-I. Identification and characterization of lipopolysaccharide in acetic acid bacteria. Anticancer. Res. 2007, 26, 3997–4002. [Google Scholar]
- Dekker, R.F.H.; Rietschel, E.T.; Sandermann, H. Isolation of α-glucan and lipopolysaccharide fractions from Acetobacter xylinum. Arch. Microbiol. 1977, 115, 353–357. [Google Scholar] [CrossRef]
- Hashimoto, M.; Ozono, M.; Furuyashiki, M.; Baba, R.; Hashiguchi, S.; Suda, Y.; Fukase, K.; Fujimoto, Y. Characterization of a Novel d-Glycero-d-talo-oct-2-ulosonic acid-substituted lipid a moiety in the lipopolysaccharide produced by the acetic acid bacterium Acetobacter pasteurianus NBRC 3283. J. Biol. Chem. 2016, 291, 21184–21194. [Google Scholar] [CrossRef] [Green Version]
- Pallach, M.; Di Lorenzo, F.; Facchini, F.A.; Gully, D.; Giraud, E.; Peri, F.; Duda, K.A.; Molinaro, A.; Silipo, A. Structure and inflammatory activity of the LPS isolated from Acetobacter pasteurianus CIP103108. Int. J. Biol. Macromol. 2018, 119, 1027–1035. [Google Scholar] [CrossRef]
- Medvedev, A.E.; Lentschat, A.; Kuhns, D.B.; Blanco, J.C.; Salkowski, C.; Zhang, S.; Arditi, M.; Gallin, J.I.; Vogel, S.N. Distinct Mutations in IRAK-4 Confer hyporesponsiveness to lipopolysaccharide and Interleukin-1 in a Patient with recurrent bacterial infections. J. Exp. Med. 2003, 198, 521–531. [Google Scholar] [CrossRef]
- Westphal, O.; Jann, K. Extraction with phenol-water and further applications of the procedure. Methods Carbohydr. Chem. 1965, 5, 83–91. [Google Scholar]
- Vinogradov, E.V.; Muller-Loennies, S.; Petersen, B.O.; Meshkov, S.; Thomas-Oates, J.E.; Holst, O.; Brade, H. Structural investigation of the lipopolysaccharide from Acinetobacter haemolyticus strain NCTC 10305 (ATCC 17906, DNA group 4). JBIC J. Biol. Inorg. Chem. 1997, 247, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Zähringer, U.; Kawahara, K.; Kosma, P. Isolation and structural characterization of a (Kdo-isosteric) d-glycero-α-d-talo-oct-2-ulopyranosidonic acid (Ko) interlinking lipid A and core oligosaccharide in the lipopolysaccharide of Acinetobacter calcoaceticus NCTC 10305. Carbohydr. Res. 2013, 378, 63–70. [Google Scholar] [CrossRef]
- Brown, D.B.; Muszyński, A.; Carlson, R.W. Elucidation of a novel lipid A α-(1,1)-GalA transferase gene (rgtF) from Mesorhizobium loti: Heterologous expression of RGTF causes Rhizobium ETLI to synthesize lipid A with α-(1,1)-GalA. Glycobiology 2013, 23, 546–558. [Google Scholar] [CrossRef] [Green Version]
- Moran, A.P.; Zahringer, U.; Seydel, U.; Scholz, D.; Stutz, P.; Rietschel, E.T. Structural analysis of the lipid A component of Campylobacter jejuni CCUG 10936 (serotype O:2) lipopolysaccharide. Description of a lipid A containing a hybrid backbone of 2-amino-2-deoxy-D-glucose and 2,3-diamino-2,3-dideoxy-D-glucose. JBIC J. Biol. Inorg. Chem. 1991, 198, 459–469. [Google Scholar] [CrossRef]
- Chung, H.S.; Yang, E.G.; Hwang, D.; Lee, J.E.; Guan, Z.; Raetz, C.R. Kdo hydroxylase is an inner core assembly enzyme in the Ko-containing lipopolysaccharide biosynthesis. Biochem. Biophys. Res. Commun. 2014, 452, 789–794. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, S.G.; Taylor, D.P. Occurrence of 2,3-Diamino-2,3-dideoxy-D-glucose in Lipid A from lipopolysaccharide of Pseudomonas diminuta. J. Gen. Microbiol. 1978, 109, 367–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zähringer, U.; Lindner, B.; Knirel, Y.A.; Akker, W.M.R.V.D.; Hiestand, R.; Heine, H.; Dehio, C. Structure and biological activity of the short-chain lipopolysaccharide from Bartonella henselae ATCC 49882T. J. Biol. Chem. 2004, 279, 21046–21054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malgorzata-Miller, G.; Heinbockel, L.; Brandenburg, K.; Van Der Meer, J.W.M.; Netea, M.G.; Joosten, L.A.B. Bartonella quintana lipopolysaccharide (LPS): Structure and characteristics of a potent TLR4 antagonist for in-vitro and in-vivo applications. Sci. Rep. 2016, 6, 34221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenberg, D.E.; Porcella, S.F.; Zelazny, A.M.; Virtaneva, K.; Sturdevant, D.E.; Kupko, J.J.; Barbian, K.D.; Babar, A.; Dorward, D.W.; Holland, S.M. Genome sequence analysis of the emerging human pathogenic acetic acid bacterium Granulibacter bethesdensis. J. Bacteriol. 2007, 189, 8727–8736. [Google Scholar] [CrossRef] [Green Version]
- Molinaro, A.; Holst, O.; Di Lorenzo, F.; Callaghan, M.; Nurisso, A.; D’Errico, G.; Zamyatina, A.; Peri, F.; Berisio, R.; Jerala, R.; et al. chemistry of lipid a: At the heart of innate immunity. Chem. A Eur. J. 2015, 21, 500–519. [Google Scholar] [CrossRef]
- Sweet, C.R.; Ribeiro, A.A.; Raetz, C.R.H. Oxidation and transamination of the 3″-Position of UDP-N-acetylglucosamine by Enzymes from Acidithiobacillus ferrooxidans. J. Biol. Chem. 2004, 279, 25400–25410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Mourik, A.; Steeghs, L.; van Laar, J.; Meiring, H.D.; Hamstra, H.J.; van Putten, J.P.; Wösten, M.M. Altered linkage of hydroxyacyl chains in lipid A of Campylobacter jejuni reduces TLR4 activation and antimicrobial resistance. J. Biol. Chem. 2010, 285, 15828–15836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunn, J.S. The Salmonella PmrAB regulon: Lipopolysaccharide modifications, antimicrobial peptide resistance and more. Trends Microbiol. 2008, 16, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Ematsuura, M. Structural modifications of bacterial lipopolysaccharide that facilitate gram-negative bacteria evasion of host innate immunity. Front. Immunol. 2013, 4, 109. [Google Scholar] [CrossRef] [Green Version]
- Raetz, C.R.; Reynolds, C.M.; Trent, M.S.; Bishop, R.E. Lipid A modification systems in gram-negative bacteria. Annu. Rev. Biochem. 2007, 76, 295–329. [Google Scholar] [CrossRef] [Green Version]
- Rolin, O.; Muse, S.J.; Safi, C.; Elahi, S.; Gerdts, V.; Hittle, L.E.; Ernst, R.K.; Harvill, E.T.; Preston, A. Enzymatic modification of lipid A by ArnT Protects Bordetella bronchiseptica against cationic peptides and is required for transmission. Infect. Immun. 2013, 82, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, D.E.; Sturdevant, D.E.; Marshall-Batty, K.R.; Chu, J.; Pettinato, A.M.; Virtaneva, K.; Lane, J.; Geller, B.L.; Porcella, S.F.; Gallin, J.I.; et al. Simultaneous host-pathogen transcriptome analysis during Granulibacter bethesdensis infection of neutrophils from healthy subjects and patients with chronic granulomatous disease. Infect. Immun. 2015, 83, 4277–4292. [Google Scholar] [CrossRef] [Green Version]
- Karbarz, M.J.; Six, D.A.; Raetz, C.R.H. Purification and characterization of the Lipid A 1-Phosphatase LpxE of Rhizobium leguminosarum. J. Biol. Chem. 2009, 284, 414–425. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ribeiro, A.A.; Guan, Z.; Abraham, S.N.; Raetz, C.R.H. Attenuated virulence of a Francisella mutant lacking the lipid A 4′-phosphatase. Proc. Natl. Acad. Sci. USA 2007, 104, 4136–4141. [Google Scholar] [CrossRef] [Green Version]
- Carlson, R.W.; Forsberg, L.S.; Kannenberg, E.L. Lipopolysaccharides in rhizobium-legume symbioses. In Endotoxins: Structure, Function and Recognition; Wang, X., Quinn, P.J., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 339–386. [Google Scholar]
- Grimmecke, H.D.; Mamat, U.; Lauk, W.; Shashkov, A.S.; Knirel, Y.A.; Vinogradov, E.V.; Kochetkov, N.K. Structure of the capsular polysaccharide and the O-side- chain of the lipopolysaccharide from Acetabacter methanolicus MB 584 (IMET (10945), and of oligosaccharides resulting from their degradation by the bacteriophage Acml. Carbohydr. Res. 1991, 220, 165–172. [Google Scholar] [CrossRef]
- Previato, J.; Jones, C.; Stephan, M.P.; Almeida, L.P.; Mendonça-Previato, L.; Stephen, M.P. Structure of the repeating oligosaccharide from the lipopolysaccharide of the nitrogen-fixing bacterium Acetobacter diazotrophicus strain PAL 5. Carbohydr. Res. 1997, 298, 311–318. [Google Scholar] [CrossRef]
- Greenberg, D.E.; Shoffner, A.R.; Zelazny, A.M.; Fenster, M.E.; Zarember, K.A.; Stock, F.; Ding, L.; Marshall-Batty, K.R.; Wasserman, R.L.; Welch, D.F.; et al. Recurrent Granulibacter bethesdensis infections and chronic granulomatous disease. Emerg. Infect. Dis. 2010, 16, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Muszyński, A.; Laus, M.; Kijne, J.W.; Carlson, R.W. Structures of the lipopolysaccharides from Rhizobium leguminosarum RBL5523 and its UDP-glucose dehydrogenase mutant (exo5). Glycobiology 2010, 21, 55–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, U.; Forsberg, L.; Carlson, R. Structure of lipid A component of Rhizobium leguminosarum bv. phaseoli lipopolysaccharide. Unique nonphosphorylated lipid A containing 2-amino-2-deoxygluconate, galacturonate, and glucosamine. J. Biol. Chem. 1994, 269, 14402–14410. [Google Scholar] [CrossRef]
- York, W.S.; Darvill, A.G.; McNeil, M.; Stevenson, T.T.; Albersheim, P. Isolation and characterization of plant cell walls and cell wall components. In Methods Enzymol; Jackman, J., Ed.; Academic Press: Cambridge, MA, USA, 1986; Volume 118, pp. 3–40. [Google Scholar]
- Roppel, J.; Mayer, H.; Weckesser, J. Identification of a 2,3-diamino-2,3-dideoxyhexose in the lipid a component of lipopolysaccharides of Rhodopseudomonas viridis and Rhodopseudomonas palustris. Carbohydr. Res. 1975, 40, 31–40. [Google Scholar] [CrossRef]
- Kelly, S.J.; Muszyński, A.; Kawaharada, Y.; Hubber, A.M.; Sullivan, J.T.; Sandal, N.; Carlson, R.W.; Stougaard, J.; Ronson, C.W. Conditional requirement for exopolysaccharide in the mesorhizobium–lotus symbiosis. Mol. Plant Microbe Interact. 2013, 26, 319–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krauss, J.H.; Weckesser, J.; Mayer, H. Electrophoretic analysis of lipopolysaccharides of purple nonsulfur bacteria. Int. J. Syst. Bacteriol. 1988, 38, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Corzo, J.; Pérez-Galdona, R.; León-Barrios, M.; Gutiérrez-Navarro, A.M. Alcian blue fixation allows silver staining of the isolated polysaccharide component of bacterial lipopolysaccharides in polyacrylamide gels. Electrophoresis 1991, 12, 439–441. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Residue | Chemical Shift (ppm) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| D | 6-α-GlcN | 5.06 | 4.26 | 5.16 | 3.41 | 4.10 | 4.05/3.58 | ||
| 93.1 | 51.1 | 74.3 | 69.0 | 71.4 | 68.3 | ||||
| E | α-GlcA | 5.13 | 3.65 | 3.85 | 3.65 | 4.17 | |||
| 94.3 | 71.1 | 72.3 | 72.0 | 71.5 | ND | ||||
| A | α-Man | 4.96 | 3.74 | 3.56 | 3.61 | 3.43 | 3.63/3.60 | ||
| 101.8 | 70.7 | 70.7 | 67.4 | 73.9 | 64.1 | ||||
| C | 4,6-β-GlcN3N | 4.36 | 3.70 | 4.03 | 3.42 | 3.60 | 3.85/3.80 | ||
| 101.4 | 53.4 | 54.7 | 75.8 | 74.3 | 63.1 | ||||
| B | α-Ko | 3.98 | 3.85 | 4.09 | 3.77 | 4.03 | 3.81/3.69 | ||
| ND | ND | 71.6 | 65.9 | 68.8 | 71.8 | 70.1 | 61.8 | ||
| Acyl | Residue | Chemical Shift (ppm) | ||||
|---|---|---|---|---|---|---|
| Chain No. | 2 | 3 | 4 | |||
| V | (3-OH)-FA | 2.33/2.26 | 3.92 | 1.44/1.39 | ||
| amide linked | 42.9 | 68.5 | 37.4 | |||
| W | FA | 2.27/2.27 | 1.58/1.58 | 1.27/1.27 | ||
| ester linked | 34.5 | 25.1 | 29.6 | |||
| X | (3-OR)-FA | 2.45/2.31 | 5.07 | 1.52/1.52 | ||
| amide linked | 41.2 | 71.0 | 34.0 | |||
| Y | (3-OH)-FA | 2.49/2.39 | 3.98 | 1.48/1.41 | ||
| ester linked | 42.2 | 68.5 | 37.2 | |||
| Z | (3-OH)-FA | 2.31/2.18 | 3.97 | 1.41/1.41 | ||
| amide linked | 44.2 | 68.5 | 37.5 | |||
| bulk -CH2- | ω-3 | ω-2 | ω-1 | ω | ||
| All fatty acids | 1.27/1.27 | 1.38/1.29 | 1.24/1.24 | 1.27/1.27 | 0.87 | |
| 29.6 | 25.5 | 31.9 | 22.6 | 14.0 | ||
| Ion, Peak Label | Obs. [M-H]− | Predicted [M-H]− | MW [M] | Glycosyl Composition | Proposed FA Composition (penta-acyl Lipid A) |
|---|---|---|---|---|---|
| I | 2112.00 | 2111.36 | 2113.72 | Man, GlcN3N(Ko), GlcN, GlcA | (14:0(3-OH))3; 16:0; 18:0(3-OH) |
| II | 2140.02 | 2139.39 | 2141.79 | Man, GlcN3N(Ko), GlcN, GlcA | (14:0(3-OH))2; 16:0(3-OH); 16:0; 18:0(3-OH) |
| III | 2168.04 | 2167.42 | 2169.82 | Man, GlcN3N(Ko), GlcN, GlcA | (14:0(3-OH))2; 16:0; 18:0(3-OH)2 |
| I-GlcA | 1934.87 | 1935.33 | 1937.59 | Man, GlcN3N(Ko), GlcN | (14:0(3-OH))3; 16:0; 18:0(3-OH) |
| II-GlcA | 1963.91 | 1963.36 | 1965.65 | Man, GlcN3N(Ko), GlcN | (14:0(3-OH))2; 16:0(3-OH); 16:0; 18:0(3-OH) |
| I-Man | 1949.97 | 1949.30 | 1951.58 | GlcN3N(Ko), GlcN, GlcA | (14:0(3-OH))3; 16:0; 18:0(3-OH) |
| II-Man | 1978.00 | 1977.34 | 1979.63 | GlcN3N(Ko), GlcN, GlcA | (14:0(3-OH))2; 16:0(3-OH); 16:0; 18:0(3-OH) |
| III-Man | 2005.02 | 2005.37 | 2007.68 | GlcN3N(Ko), GlcN, GlcA | (14:0(3-OH))2; 16:0; 18:0(3-OH)2 |
| I-Ko | 1876.00 | 1875.31 | 1877.54 | Man, GlcN3N(Ko), GlcN, GlcA | (14:0(3-OH))3; 16:0; 18:0(3-OH) |
| II-Ko | 1902.95 | 1903.34 | 1905.59 | Man, GlcN3N(Ko), GlcN, GlcA | (14:0(3-OH))2; 16:0(3-OH); 16:0; 18:0(3-OH) |
| III-Ko | 1930.98 | 1931.37 | 1933.65 | Man, GlcN3N(Ko), GlcN, GlcA | (14:0(3-OH))2; 16:0; 18:0(3-OH)2 |
| II-Ko-Man | 1740.95 | 1741.28 | 1743.45 | GlcN3N(Ko), GlcN, GlcA | (14:0(3-OH))2; 16:0(3-OH); 16:0; 18:0(3-OH) |
| II-Ko-Man | 1769.85 | 1769.31 | 1771.50 | GlcN3N(Ko), GlcN, GlcA | (14:0(3-OH))2; 16:0; 18:0(3-OH)2 |
| Ion | Obs. [M-H]− | Predicted [M-H]− | MW [M] | Glycosyl Composition | Proposed FA Composition de-O-acylated (Triacyl-Lipid A) |
|---|---|---|---|---|---|
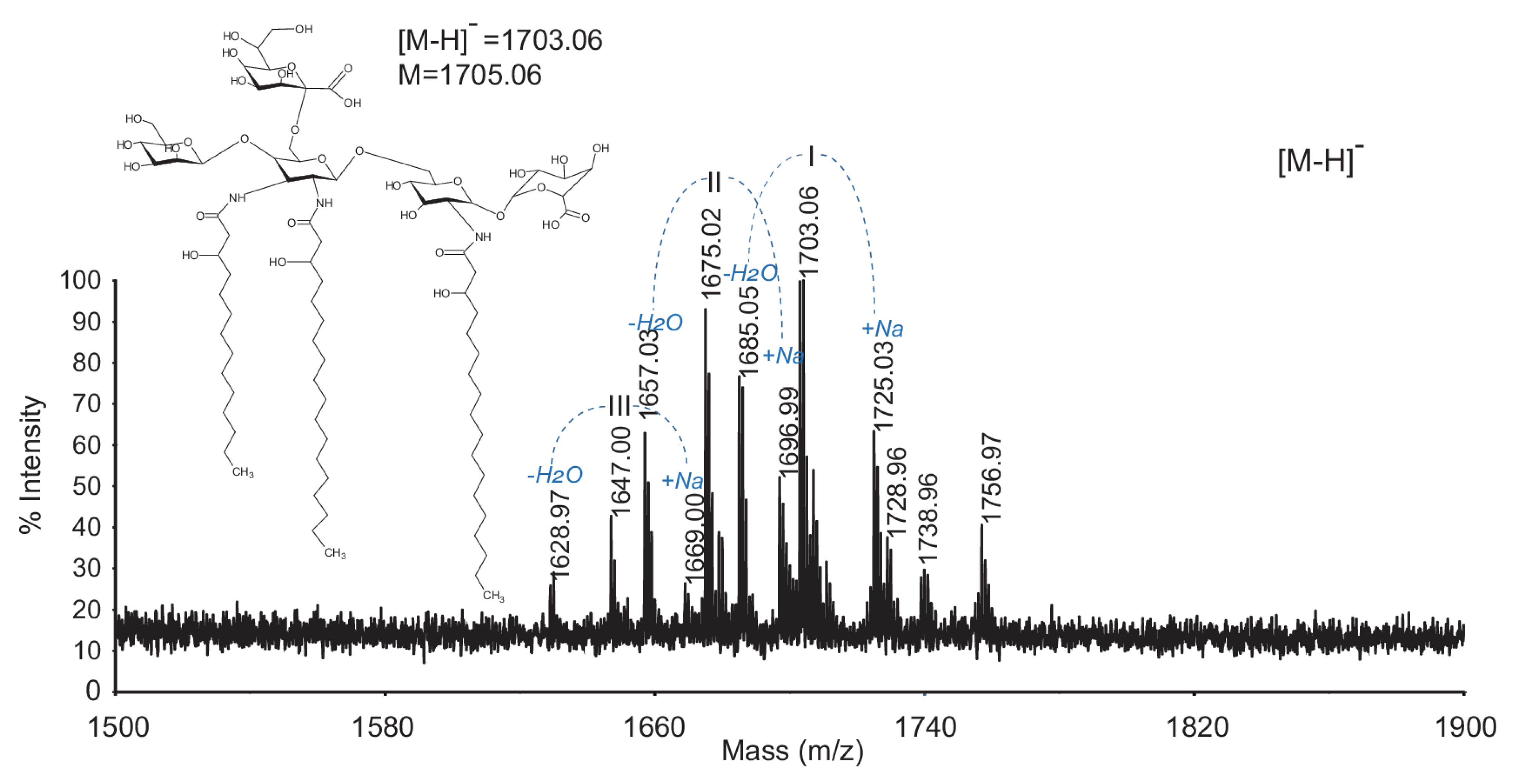
| I | 1703.06 | 1703.00 | 1705.06 | Man, GlcN3N(Ko), GlcN, GlcA | 18:0(3-OH); 16:0(3-OH); 14:0(3-OH) |
| 1725.02 | 1725.99 | 1728.05 | (Man, GlcN3N(Ko), GlcN, GlcA)Na | 18:0(3-OH); 16:0(3-OH); 14:0(3-OH) | |
| 1685.05 | 1684.99 | 1687.04 | (Man, GlcN3N(Ko), GlcN, GlcA)anhydro | 18:0(3-OH); 16:0(3-OH); 14:0(3-OH) | |
| II | 1675.03 | 1674.97 | 1677.01 | Man, GlcN3N(Ko), GlcN, GlcA | 18:0(3-OH); 16:0(3-OH); 14:0(3-OH) |
| 1696.99 | 1697.96 | 1699.99 | (Man, GlcN3N(Ko), GlcN, GlcA)Na | 18:0(3-OH); 16:0(3-OH); 14:0(3-OH) | |
| 1657.02 | 1656.96 | 1658.99 | (Man, GlcN3N(Ko), GlcN, GlcA)anhydro | 18:0(3-OH); 16:0(3-OH); 14:0(3-OH) | |
| III | 1647.00 | 1646.94 | 1648.95 | Man, GlcN3N(Ko), GlcN, GlcA | 18:0(3-OH); 14:0(3-OH) |
| 1669.99 | 1699.93 | 1671.94 | (Man, GlcN3N(Ko), GlcN, GlcA)Na | 18:0(3-OH); 14:0(3-OH) | |
| 1628.97 | 1628.92 | 1630.93 | (Man, GlcN3N(Ko), GlcN, GlcA)anhydro | 18:0(3-OH); 14:0(3-OH) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muszyński, A.; Zarember, K.A.; Heiss, C.; Shiloach, J.; Berg, L.J.; Audley, J.; Kozyr, A.; Greenberg, D.E.; Holland, S.M.; Malech, H.L.; et al. Granulibacter bethesdensis, a Pathogen from Patients with Chronic Granulomatous Disease, Produces a Penta-Acylated Hypostimulatory Glycero-D-talo-oct-2-ulosonic Acid–Lipid A Glycolipid (Ko-Lipid A). Int. J. Mol. Sci. 2021, 22, 3303. https://doi.org/10.3390/ijms22073303
Muszyński A, Zarember KA, Heiss C, Shiloach J, Berg LJ, Audley J, Kozyr A, Greenberg DE, Holland SM, Malech HL, et al. Granulibacter bethesdensis, a Pathogen from Patients with Chronic Granulomatous Disease, Produces a Penta-Acylated Hypostimulatory Glycero-D-talo-oct-2-ulosonic Acid–Lipid A Glycolipid (Ko-Lipid A). International Journal of Molecular Sciences. 2021; 22(7):3303. https://doi.org/10.3390/ijms22073303
Chicago/Turabian StyleMuszyński, Artur, Kol A. Zarember, Christian Heiss, Joseph Shiloach, Lars J. Berg, John Audley, Arina Kozyr, David E. Greenberg, Steven M. Holland, Harry L. Malech, and et al. 2021. "Granulibacter bethesdensis, a Pathogen from Patients with Chronic Granulomatous Disease, Produces a Penta-Acylated Hypostimulatory Glycero-D-talo-oct-2-ulosonic Acid–Lipid A Glycolipid (Ko-Lipid A)" International Journal of Molecular Sciences 22, no. 7: 3303. https://doi.org/10.3390/ijms22073303