
Identification of Novel FNIN2 and FNIN3 Fibronectin-Derived Peptides That Promote Cell Adhesion, Proliferation and Differentiation in Primary Cells and Stem Cells

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Interaction of Fibronectin with Integrin Receptors
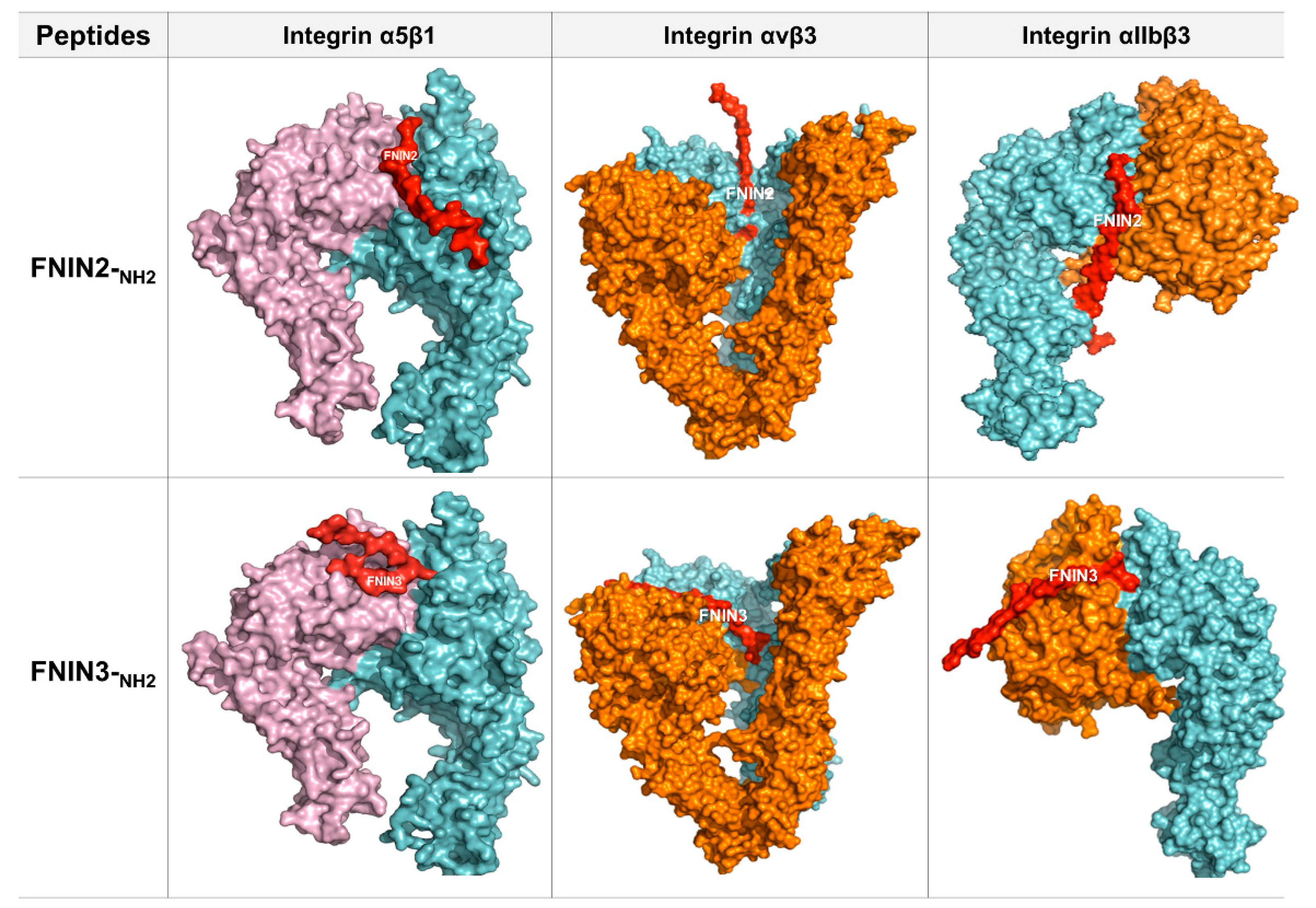
2.2. Interactions between FNIN2 and FNIN3 and Integrin Receptors
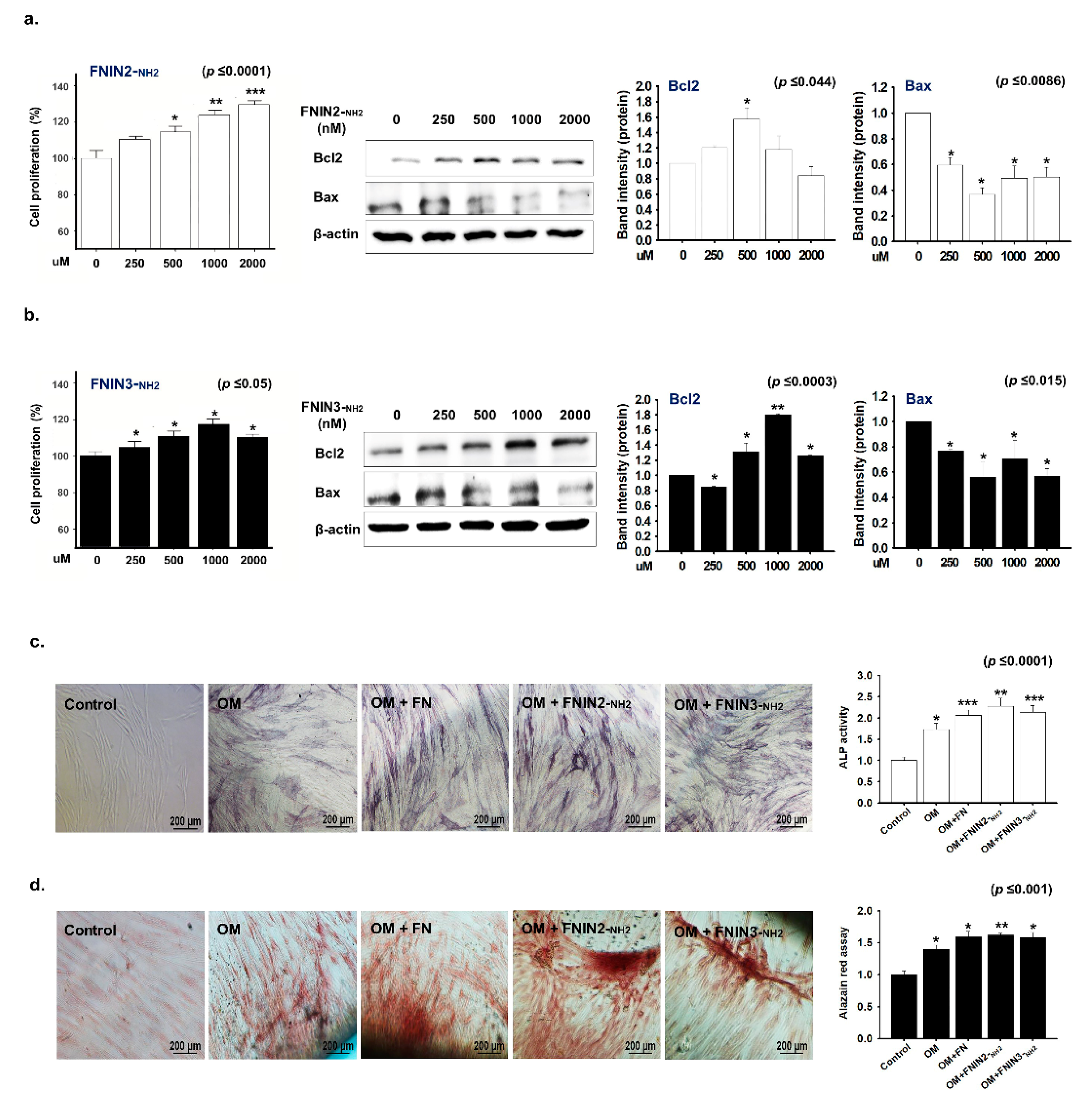
2.3. Cells Proliferation Induction by FNINs
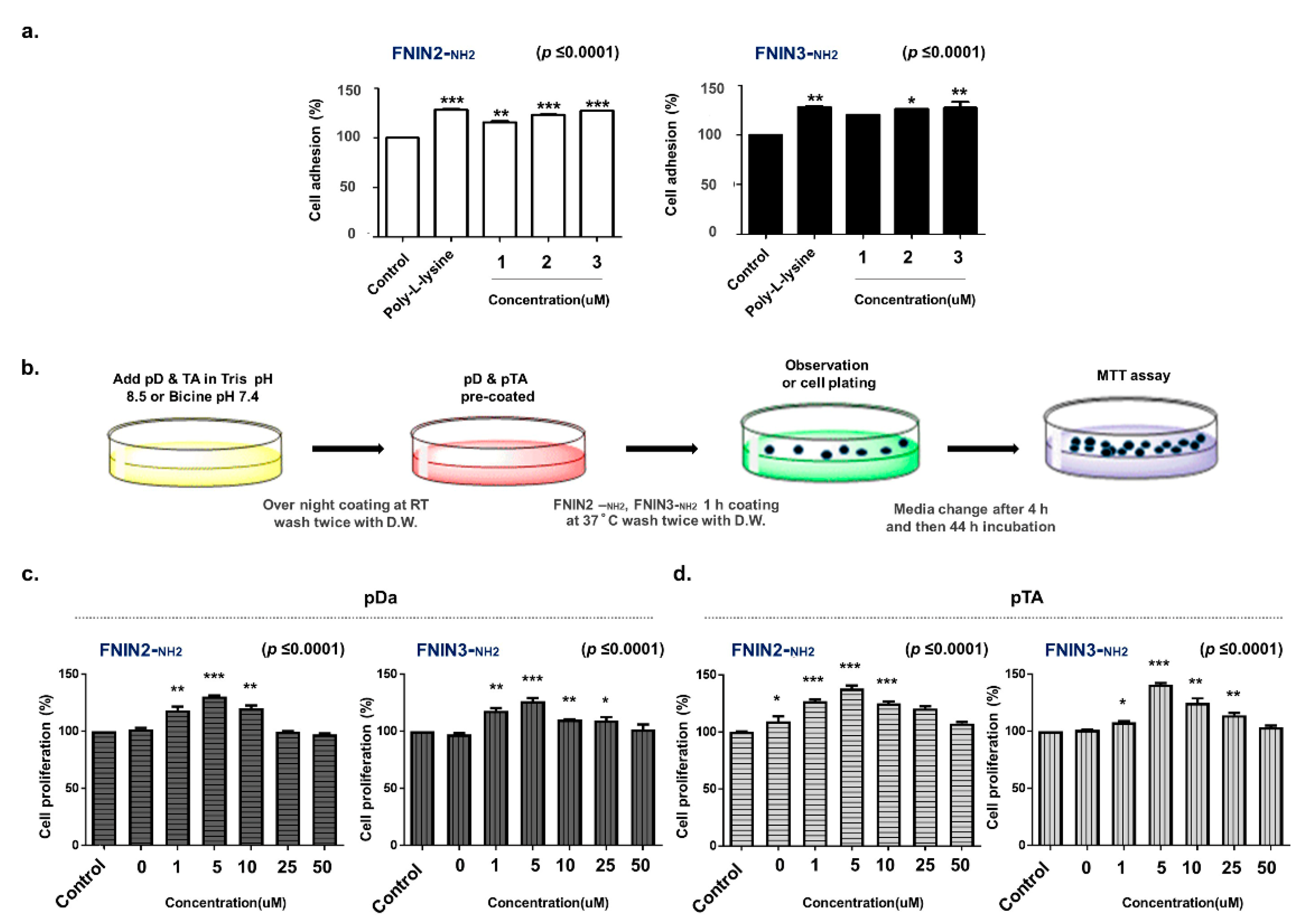
2.4. Assessment of Cell Attachment and Proliferation in FNINs Coated Plate
2.5. Effects of FNIN2 or FNIN3 on the Proliferation and Osteogenic Differentiation of Human Mesenchymal Stem Cells (MSCs)
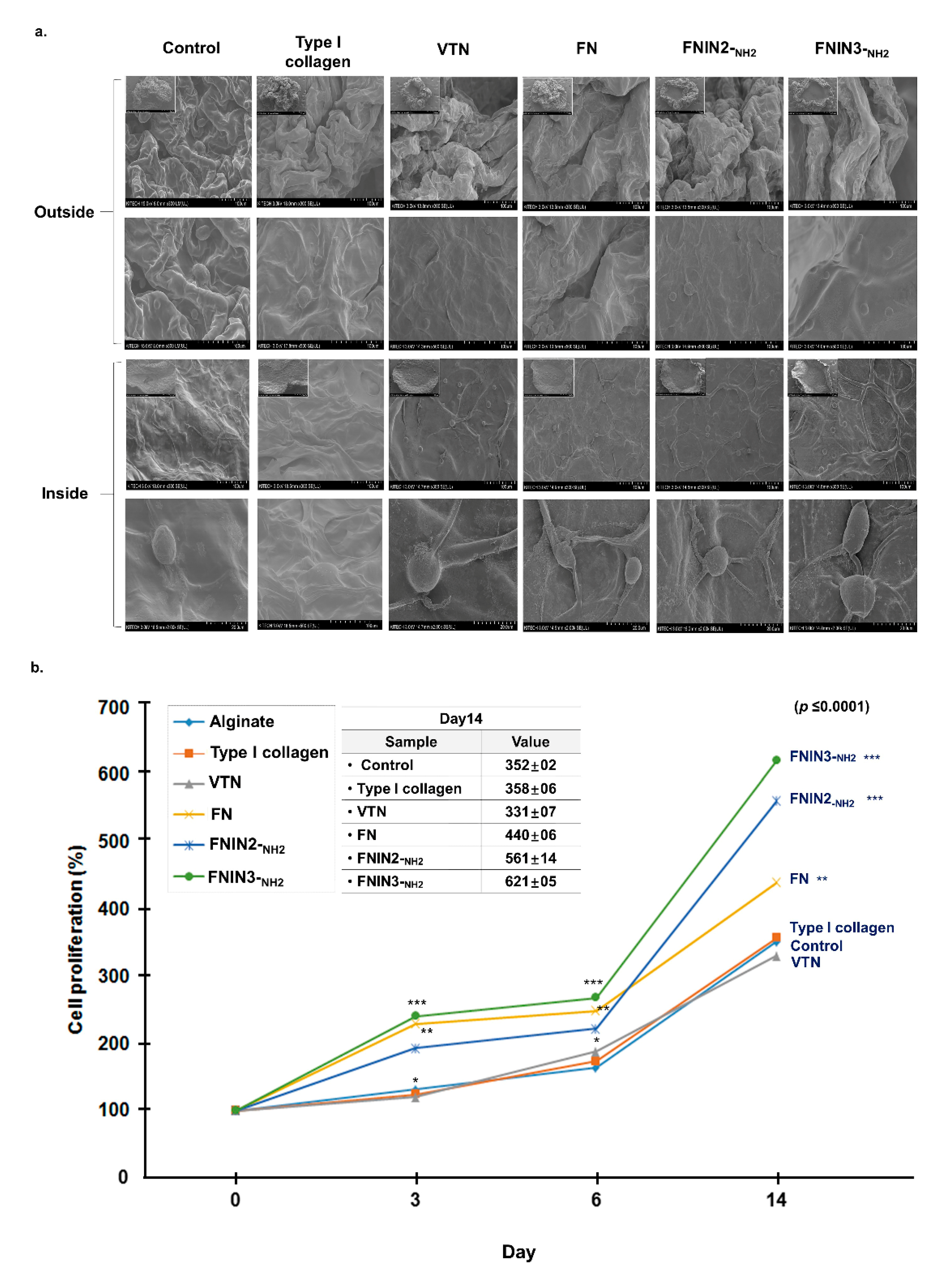
2.6. C2C12 Cell Proliferation Induced by FNIN2-NH2 or FNIN3-NH2 in Alginate Beads
3. Discussion
4. Materials and Methods
4.1. In Silico Experiments
4.1.1. D Structures of Fibronectin and Integrin
4.1.2. Interactions between Fibronectin and Integrins (α5β1, αvβ3, and αIIbβ3)
4.1.3. Prediction of Fibronectin Short Peptides
4.1.4. Interactions between the Designed Peptides and Integrins
4.2. FNIN Peptide Preparation
4.3. Cell Proliferation
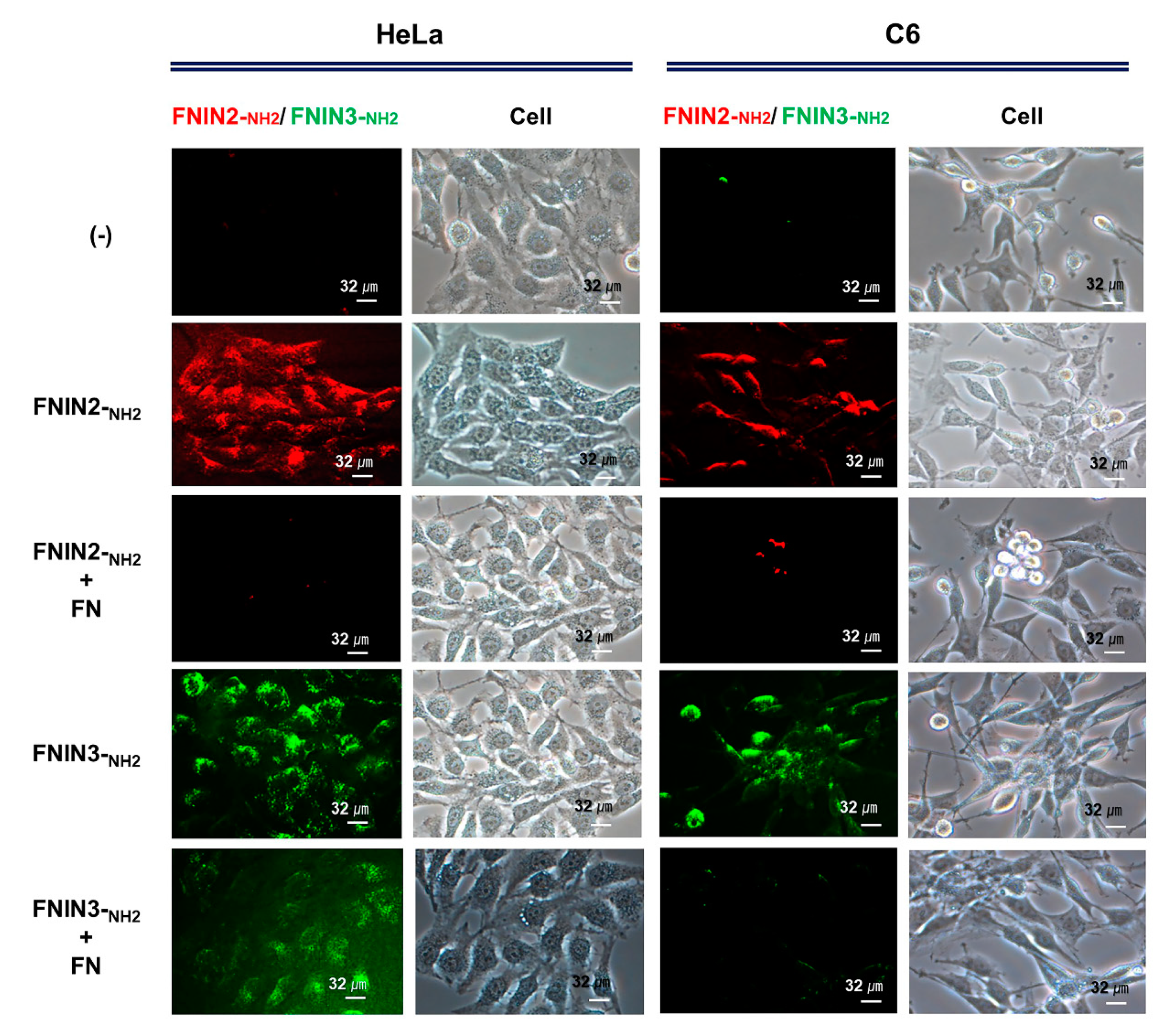
4.4. Binding of FNIN2-NH2 Peptide to Cells
4.5. Binding of FNIN2-NH2 and FNIN3-NH2 Peptides with FN Protein to Cells
4.6. Polymerized Dopamine (pD) or Polymerized TA (pTA) Pre-Coating Prior to FNIN2-NH2 or FNIN3-NH2 Coating on Plates
4.7. Cell Adhesion and Proliferation Analysis in HBEpiC Cells
4.8. Proliferation of MSCs by FNIN2# and FNIN3#
4.9. Osteogenic Differentiation of MSC by FNIN2-NH2 and FNIN3-NH2
4.10. Alginate Bead Preparation, 3D Culture, and Cell Proliferation
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys. Acta 2014, 1840, 2506–2519. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, K.; Lee, E.J.; Moon, J.S.; Park, S.Y.; Choi, I. Multifaceted Interweaving between Extracellular Matrix, Insulin Resistance, and Skeletal Muscle. Cells 2018, 7, 148. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Qiu, Y.; Zeng, X.; Ding, Y.; Zeng, J.; Lu, K.; Li, D. Effect of a feeder layer composed of mouse embryonic and human foreskin fibroblasts on the proliferation of human embryonic stem cells. Exp. Ther. Med. 2016, 11, 2321–2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimanskaya, I.; Chung, Y.; Meisner, L.; Johnson, J.; West, M.D.; Lanza, R. Human embryonic stem cells derived without feeder cells. Lancet 2005, 365, 1636–1641. [Google Scholar] [CrossRef]
- Antoni, D.; Burckel, H.; Josset, E.; Noel, G. Three-dimensional cell culture: A breakthrough in vivo. Int. J. Mol. Sci. 2015, 16, 5517–5527. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning from 2D to 3D Cell Culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Karsdal, M.A.; Nielsen, M.J.; Sand, J.M.; Henriksen, K.; Genovese, F.; Bay-Jensen, A.C.; Smith, V.; Adamkewicz, J.I.; Christiansen, C.; Leeming, D.J. Extracellular matrix remodeling: The common denominator in connective tissue diseases. Possibilities for evaluation and current understanding of the matrix as more than a passive architecture, but a key player in tissue failure. Assay Drug Dev. Technol. 2013, 11, 70–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular matrix assembly: A multiscale deconstruction. Nat. Rev. Mol. Cell Biol. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Vanhee, P.; van der Sloot, A.M.; Verschueren, E.; Serrano, L.; Rousseau, F.; Schymkowitz, J. Computational design of peptide ligands. Trends Biotechnol. 2011, 29, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Benson, N.; Cucurull-Sanchez, L.; Demin, O.; Smirnov, S.; van der Graaf, P. Reducing systems biology to practice in pharmaceutical company research; selected case studies. Adv. Exp. Med. Biol. 2012, 736, 607–615. [Google Scholar] [CrossRef]
- Baig, M.H.; Ahmad, K.; Saeed, M.; Alharbi, A.M.; Barreto, G.E.; Ashraf, G.M.; Choi, I. Peptide based therapeutics and their use for the treatment of neurodegenerative and other diseases. Biomed. Pharmacother. 2018, 103, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Patel, L.N.; Zaro, J.L.; Shen, W.C. Cell penetrating peptides: Intracellular pathways and pharmaceutical perspectives. Pharm. Res. 2007, 24, 1977–1992. [Google Scholar] [CrossRef] [PubMed]
- Baig, M.H.; Ahmad, K.; Rabbani, G.; Choi, I. Use of peptides for the management of Alzheimer’s disease: Diagnosis and inhibition. Front. Aging Neurosci. 2018, 10, 21. [Google Scholar] [CrossRef]
- Baig, M.H.; Ahmad, K.; Roy, S.; Ashraf, J.M.; Adil, M.; Siddiqui, M.H.; Khan, S.; Kamal, M.A.; Provaznik, I.; Choi, I. Computer Aided Drug Design: Success and Limitations. Curr. Pharm. Des. 2016, 22, 572–581. [Google Scholar] [CrossRef]
- Mabonga, L.; Kappo, A.P. Protein-protein interaction modulators: Advances, successes and remaining challenges. Biophys. Rev. 2019, 11, 559–581. [Google Scholar] [CrossRef] [PubMed]
- Humphries, J.D.; Byron, A.; Humphries, M.J. Integrin ligands at a glance. J. Cell Sci. 2006, 119, 3901–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiss, M.; Beckmann, K.; Giros, A.; Costell, M.; Fassler, R. The role of integrin binding sites in fibronectin matrix assembly in vivo. Curr. Opin. Cell Biol. 2008, 20, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Aota, S.; Yamada, K.M. Fibronectin and cell adhesion: Specificity of integrin-ligand interaction. Adv. Enzymol. Relat. Areas Mol. Biol. 1995, 70, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, K.; Lee, E.J.; Shaikh, S.; Kumar, A.; Rao, K.M.; Park, S.Y.; Jin, J.O.; Han, S.S.; Choi, I. Targeting integrins for cancer management using nanotherapeutic approaches: Recent advances and challenges. Semin Cancer Biol. 2019. [Google Scholar] [CrossRef]
- Nagae, M.; Re, S.; Mihara, E.; Nogi, T.; Sugita, Y.; Takagi, J. Crystal structure of alpha5beta1 integrin ectodomain: Atomic details of the fibronectin receptor. J. Cell Biol. 2012, 197, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Pimton, P.; Sarkar, S.; Sheth, N.; Perets, A.; Marcinkiewicz, C.; Lazarovici, P.; Lelkes, P.I. Fibronectin-mediated upregulation of alpha5beta1 integrin and cell adhesion during differentiation of mouse embryonic stem cells. Cell Adhes. Migr. 2011, 5, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaffner, F.; Ray, A.M.; Dontenwill, M. Integrin alpha5beta1, the Fibronectin Receptor, as a Pertinent Therapeutic Target in Solid Tumors. Cancers 2013, 5, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, S.K.; Olden, K.; Yamada, K.M. Fibronectin and integrins in invasion and metastasis. Cancer Metastasis Rev. 1995, 14, 173–189. [Google Scholar] [CrossRef]
- Johansson, S.; Svineng, G.; Wennerberg, K.; Armulik, A.; Lohikangas, L. Fibronectin-integrin interactions. Front. Biosci. 1997, 2, d126–d146. [Google Scholar] [CrossRef] [Green Version]
- Missirlis, D.; Haraszti, T.; Kessler, H.; Spatz, J.P. Fibronectin promotes directional persistence in fibroblast migration through interactions with both its cell-binding and heparin-binding domains. Sci. Rep. 2017, 7, 3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashino, K.; Uemori, Y.; Kimizuka, F.; Kato, I.; Titani, K. A 31-kDa recombinant fibronectin cell-binding domain fragment: Its binding to receptor, cell adhesive activity, and fusion proteins. J. Biochem. 1996, 119, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, F.; Gu, W.; Yang, H.; Meng, Q.; Zhang, Y.; Yang, H.; Duan, Q. The roles of platelet GPIIb/IIIa and αvβ3 integrins during HeLa cells adhesion, migration, and invasion to monolayer endothelium under static and dynamic shear flow. J. Biomed. Biotechnol. 2009, 2009, 829243. [Google Scholar] [CrossRef]
- Taherian, A.; Li, X.; Liu, Y.; Haas, T.A. Differences in integrin expression and signaling within human breast cancer cells. BMC Cancer 2011, 11, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermann, M.-R.; Jakobson, M.; Colo, G.P.; Rognoni, E.; Jakobson, M.; Kupatt, C.; Posern, G.; Fässler, R. Integrins synergise to induce expression of the MRTF-A–SRF target gene ISG15 for promoting cancer cell invasion. J. Cell Sci. 2016, 129, 1391–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Dellatore, S.M.; Miller, W.M.; Messersmith, P.B. Mussel-inspired surface chemistry for multifunctional coatings. Science 2007, 318, 426–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abouelmagd, S.A.; Meng, F.F.; Kim, B.K.; Hyun, H.; Yeo, Y. Tannic Acid-Mediated Surface Functionalization of Polymeric Nanoparticles. ACS Biomater. Sci. Eng. 2016, 2, 2294–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, K.; Choi, I.; Lee, Y.H. Implications of Skeletal Muscle Extracellular Matrix Remodeling in Metabolic Disorders: Diabetes Perspective. Int. J. Mol. Sci. 2020, 21, 3845. [Google Scholar] [CrossRef]
- Reilly, G.C.; Engler, A.J. Intrinsic extracellular matrix properties regulate stem cell differentiation. J. Biomech. 2010, 43, 55–62. [Google Scholar] [CrossRef]
- Guilak, F.; Cohen, D.M.; Estes, B.T.; Gimble, J.M.; Liedtke, W.; Chen, C.S. Control of stem cell fate by physical interactions with the extracellular matrix. Cell Stem Cell 2009, 5, 17–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.J.; Jan, A.T.; Baig, M.H.; Ahmad, K.; Malik, A.; Rabbani, G.; Kim, T.; Lee, I.-K.; Lee, Y.H.; Park, S.-Y. Fibromodulin and regulation of the intricate balance between myoblast differentiation to myocytes or adipocyte-like cells. FASEB J. 2017, 32, 768–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampe, K.J.; Heilshorn, S.C. Building stem cell niches from the molecule up through engineered peptide materials. Neurosci. Lett. 2012, 519, 138–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachman, H.; Nicosia, J.; Dysart, M.; Barker, T.H. Utilizing Fibronectin Integrin-Binding Specificity to Control Cellular Responses. Adv. Wound Care 2015, 4, 501–511. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, C.T.; Cheng, H.W.; Huang, C.M.; Li, H.R.; Ou, M.H.; Huang, J.R.; Khoo, K.H.; Yu, H.W.; Chen, Y.Q.; Wang, Y.K.; et al. Fibronectin in cell adhesion and migration via N-glycosylation. Oncotarget 2017, 8, 70653–70668. [Google Scholar] [CrossRef] [Green Version]
- Plow, E.F.; Haas, T.A.; Zhang, L.; Loftus, J.; Smith, J.W. Ligand binding to integrins. J. Biol. Chem. 2000, 275, 21785–21788. [Google Scholar] [CrossRef] [Green Version]
- Ruoslahti, E. RGD and other recognition sequences for integrins. Annu. Rev. Cell Dev. Biol. 1996, 12, 697–715. [Google Scholar] [CrossRef] [PubMed]
- Pierschbacher, M.D.; Ruoslahti, E. Cell attachment activity of fibronectin can be duplicated by small synthetic fragments of the molecule. Nature 1984, 309, 30–33. [Google Scholar] [CrossRef]
- Hersel, U.; Dahmen, C.; Kessler, H. RGD modified polymers: Biomaterials for stimulated cell adhesion and beyond. Biomaterials 2003, 24, 4385–4415. [Google Scholar] [CrossRef]
- Zhou, P.; Tian, F.; Shang, Z. 2D depiction of nonbonding interactions for protein complexes. J. Comput. Chem. 2009, 30, 940–951. [Google Scholar] [CrossRef] [PubMed]
- Raab-Westphal, S.; Marshall, J.F.; Goodman, S.L. Integrins as Therapeutic Targets: Successes and Cancers. Cancers 2017, 9, 110. [Google Scholar] [CrossRef] [PubMed]
- Aoshiba, K.; Rennard, S.I.; Spurzem, J.R. Fibronectin supports bronchial epithelial cell adhesion and survival in the absence of growth factors. Am. J. Physiol. 1997, 273, 684–693. [Google Scholar] [CrossRef]
- Zhang, Z.-Y.; Teoh, S.-H.; Chong, M.S.K.; Lee, E.S.M.; Tan, L.-G.; Mattar, C.N.; Fisk, N.M.; Choolani, M.; Chan, J. Neo-vascularization and bone formation mediated by fetal mesenchymal stem cell tissue-engineered bone grafts in critical-size femoral defects. Biomaterials 2010, 31, 608–620. [Google Scholar] [CrossRef]
- Mann, B.K.; Tsai, A.T.; Scott-Burden, T.; West, J.L. Modification of surfaces with cell adhesion peptides alters extracellular matrix deposition. Biomaterials 1999, 20, 2281–2286. [Google Scholar] [CrossRef]
- Meinel, L.; Hofmann, S.; Betz, O.; Fajardo, R.; Merkle, H.P.; Langer, R.; Evans, C.H.; Vunjak-Novakovic, G.; Kaplan, D.L. Osteogenesis by human mesenchymal stem cells cultured on silk biomaterials: Comparison of adenovirus mediated gene transfer and protein delivery of BMP-2. Biomaterials 2006, 27, 4993–5002. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.M.; Han, M.; Park, I.S.; Jung, Y.; Kim, S.H.; Kim, S.-H. Adhesion and differentiation of adipose-derived stem cells on a substrate with immobilized fibroblast growth factor. Acta Biomater. 2012, 8, 1759–1767. [Google Scholar] [CrossRef] [PubMed]
- Sogo, Y.; Ito, A.; Matsuno, T.; Oyane, A.; Tamazawa, G.; Satoh, T.; Yamazaki, A.; Uchimura, E.; Ohno, T. Fibronectin-calcium phosphate composite layer on hydroxyapatite to enhance adhesion, cell spread and osteogenic differentiation of human mesenchymal stem cells in vitro. Biomed. Mater. 2007, 2, 116–123. [Google Scholar] [CrossRef]
- Moursi, A.M.; Globus, R.K.; Damsky, C.H. Interactions between integrin receptors and fibronectin are required for calvarial osteoblast differentiation in vitro. J. Cell Sci. 1997, 110, 2187–2196. [Google Scholar]
- Langenbach, F.; Handschel, J. Effects of dexamethasone, ascorbic acid and beta-glycerophosphate on the osteogenic differentiation of stem cells in vitro. Stem Cell Res. Ther. 2013, 4, 117. [Google Scholar] [CrossRef] [Green Version]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.-Y.; Tsai, W.-B.; Voelcker, N.H. Screening of rat mesenchymal stem cell behaviour on polydimethylsiloxane stiffness gradients. Acta Biomater. 2012, 8, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, Z.; Huang, H.; Suzek, B.E.; Wu, C.H.; UniProt, C. A fast Peptide Match service for UniProt Knowledgebase. Bioinformatics 2013, 29, 2808–2809. [Google Scholar] [CrossRef] [Green Version]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. LIGPLOT: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. 1995, 8, 127–134. [Google Scholar] [CrossRef]
- Pathak, S.; Regmi, S.; Nguyen, T.T.; Gupta, B.; Gautam, M.; Yong, C.S.; Kim, J.O.; Son, Y.; Kim, J.R.; Park, M.H.; et al. Polymeric microsphere-facilitated site-specific delivery of quercetin prevents senescence of pancreatic islets in vivo and improves transplantation outcomes in mouse model of diabetes. Acta Biomater. 2018, 75, 287–299. [Google Scholar] [CrossRef]
- Regmi, S.; Cao, J.; Pathak, S.; Gupta, B.; Kumar Poudel, B.; Tung, P.T.; Yook, S.; Park, J.-B.; Yong, C.S.; Kim, J.O.; et al. A three-dimensional assemblage of gingiva-derived mesenchymal stem cells and NO-releasing microspheres for improved differentiation. Int. J. Pharm. 2017, 520, 163–172. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | ||||||||||
| No. | Name | Sequence | Length | Gravy | Molecular Weight | M.W Monoisotopic | Isoelectric Point | Net Charge at pH7.0 | Average Hydrophilicity | Ratio of Hydrophilic Residues/Total Number of Residues (%) |
| 1 | FNIN2-NH2 | LSISPSDNAVVLTNLLPTGE | 20 | 0.425 | 2040.3 | 2039.0787 | 4.1 | −1 | −0.3 | 35 |
| 3 | FNIN3-NH2 | TVYAVTGRGDSPASSKPC | 18 | −0.36 | 1795.01 | 1794.8571 | 9.8 | 2 | 0.1 | 33 |
| (b) | ||||||||||
| Integrin Type | FN | FNIN2-NH2 | FNIN3-NH2 | |||||||
| α5β1 | −58.51 | −77.86 | −67.08 | |||||||
| αIIbβ3 | −40.19 | −62.57 | −56.51 | |||||||
| αvβ3 | −25.27 | −65.57 | −59.18 | |||||||
| (a) | ||||||
| No. | Cells | FNIN2-NH2 | ||||
| 0 nM | 250 nM | 500 nM | 1000 nM | p Value | ||
| 1 | C2C12 | 100 ± 0 | 97 ± 3 | 95 ± 2 | 104 ± 2 | 0.0476 |
| 2 | HeLa | 100 ± 0 | 103 ± 4 | 90 ± 5 | 115 ± 7 | 0.0500 |
| 3 | HepG2 | 100 ± 0 | 99 ± 1 | 97 ± 3 | 108 ± 1 | 0.0213 |
| 4 | A498 | 100 ± 0 | 111 ± 5 | 102 ± 1 | 114 ± 4 | 0.0203 |
| 5 | Du145 | 100 ± 0 | 100 ± 1 | 94 ± 1 | 117 ± 5 | 0.0003 |
| 6 | MDA-MB-231 | 100 ± 0 | 106 ± 2 | 96 ± 2 | 91 ± 1 | 0.0002 |
| 7 | MRC-5 | 100 ± 0 | 112 ± 5 | 101 ± 3 | 107 ± 1 | 0.0261 |
| 8 | HT29 | 100 ± 0 | 98 ± 1 | 102 ± 0 | 111 ± 3 | 0.0002 |
| 9 | A431 | 100 ± 0 | 103 ± 1 | 107 ± 0 | 102 ± 0 | 0.022 |
| 10 | Fibroblast | 100 ± 0 | 91 ± 0 | 88 ± 2 | 97 ± 11 | 0.5261 |
| 11 | Cos7 | 100 ± 0 | 97 ± 4 | 94 ± 1 | 102 ± 2 | 0.1778 |
| 12 | Raw246.7 | 100 ± 0 | 110 ± 2 | 108 ± 4 | 109 ± 2 | 0.0988 |
| 13 | 3T3L1 | 100 ± 0 | 99 ± 2 | 99 ± 1 | 102 ± 3 | 0.5346 |
| 14 | Vero | 100 ± 0 | 81 ± 2 | 90 ± 3 | 93 ± 3 | 0.0027 |
| 15 | Hek293 | 100 ± 0 | 100 ± 4 | 81 ± 4 | 95 ± 6 | 0.0424 |
| 16 | C6 | 100 ± 0 | 90 ± 4 | 94 ± 1 | 113 ± 2 | 0.0114 |
| 17 | MKN28 | 100 ± 0 | 95 ± 1 | 93 ± 1 | 97 ± 1 | 0.0114 |
| (b) | ||||||
| No. | Cells | FNIN3-NH2 | ||||
| 0 nM | 250 nM | 500 nM | 1000 nM | p Value | ||
| 1 | C2C12 | 100 ± 0 | 99 ± 1 | 95 ± 1 | 99 ± 1 | 0.0069 |
| 2 | HeLa | 100 ± 0 | 97 ± 1.4 | 111 ± 0.2 | 115 ± 3 | 0.0034 |
| 3 | HepG2 | 100 ± 0 | 99 ± 6 | 117 ± 3 | 116 ± 3 | 0.0001 |
| 4 | A498 | 100 ± 0 | 103 ± 0 | 122 ± 3 | 111 ± 3 | 0.0002 |
| 5 | Du145 | 100 ± 0 | 95 ± 1 | 106 ± 1 | 99 ± 1 | 0.0005 |
| 6 | MDA-MB-231 | 100 ± 0 | 96 ± 1 | 91 ± 2 | 96 ± 1 | 0.003 |
| 7 | MRC-5 | 100 ± 0 | 90 ± 1 | 93 ± 0 | 95 ± 1 | 0.0001 |
| 8 | HT29 | 100 ± 0 | 92 ± 0 | 95 ± 1 | 93 ± 1 | 0.0001 |
| 9 | A431 | 100 ± 0 | 98 ± 1 | 104 ± 1 | 93 ± 1 | 0.0001 |
| 10 | Fibroblast | 100 ± 0 | 112 ± 4 | 92 ± 1 | 112 ± 2 | 0.0076 |
| 11 | Cos7 | 100 ± 0 | 100 ± 0 | 96 ± 1 | 102 ± 1 | 0.0165 |
| 12 | Raw246.7 | 100 ± 0 | 93 ± 3 | 82 ± 3 | 90 ± 2 | 0.0339 |
| 13 | 3T3L1 | 100 ± 0 | 98 ± 2 | 96 ± 2 | 98 ± 1 | 0.4426 |
| 14 | Vero | 100 ± 0 | 92 ± 2 | 88 ± 0 | 88 ± 3 | 0.0068 |
| 15 | Hek293 | 100 ± 0 | 71 ± 1 | 94 ± 1 | 82 ± 1 | 0.0001 |
| 16 | C6 | 100 ± 0 | 106 ± 4 | 109 ± 0 | 112 ± 7 | 0.329 |
| 17 | MKN28 | 100 ± 0 | 93 ± 0 | 96 ± 1 | 89 ± 1 | 0.0006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, E.-J.; Ahmad, K.; Pathak, S.; Lee, S.; Baig, M.H.; Jeong, J.-H.; Doh, K.-O.; Lee, D.-M.; Choi, I. Identification of Novel FNIN2 and FNIN3 Fibronectin-Derived Peptides That Promote Cell Adhesion, Proliferation and Differentiation in Primary Cells and Stem Cells. Int. J. Mol. Sci. 2021, 22, 3042. https://doi.org/10.3390/ijms22063042
Lee E-J, Ahmad K, Pathak S, Lee S, Baig MH, Jeong J-H, Doh K-O, Lee D-M, Choi I. Identification of Novel FNIN2 and FNIN3 Fibronectin-Derived Peptides That Promote Cell Adhesion, Proliferation and Differentiation in Primary Cells and Stem Cells. International Journal of Molecular Sciences. 2021; 22(6):3042. https://doi.org/10.3390/ijms22063042
Chicago/Turabian StyleLee, Eun-Ju, Khurshid Ahmad, Shiva Pathak, SunJu Lee, Mohammad Hassan Baig, Jee-Heon Jeong, Kyung-Oh Doh, Dong-Mok Lee, and Inho Choi. 2021. "Identification of Novel FNIN2 and FNIN3 Fibronectin-Derived Peptides That Promote Cell Adhesion, Proliferation and Differentiation in Primary Cells and Stem Cells" International Journal of Molecular Sciences 22, no. 6: 3042. https://doi.org/10.3390/ijms22063042