
NFAT5 Deficiency Alleviates Formalin-Induced Inflammatory Pain Through mTOR

 , , and
, , and 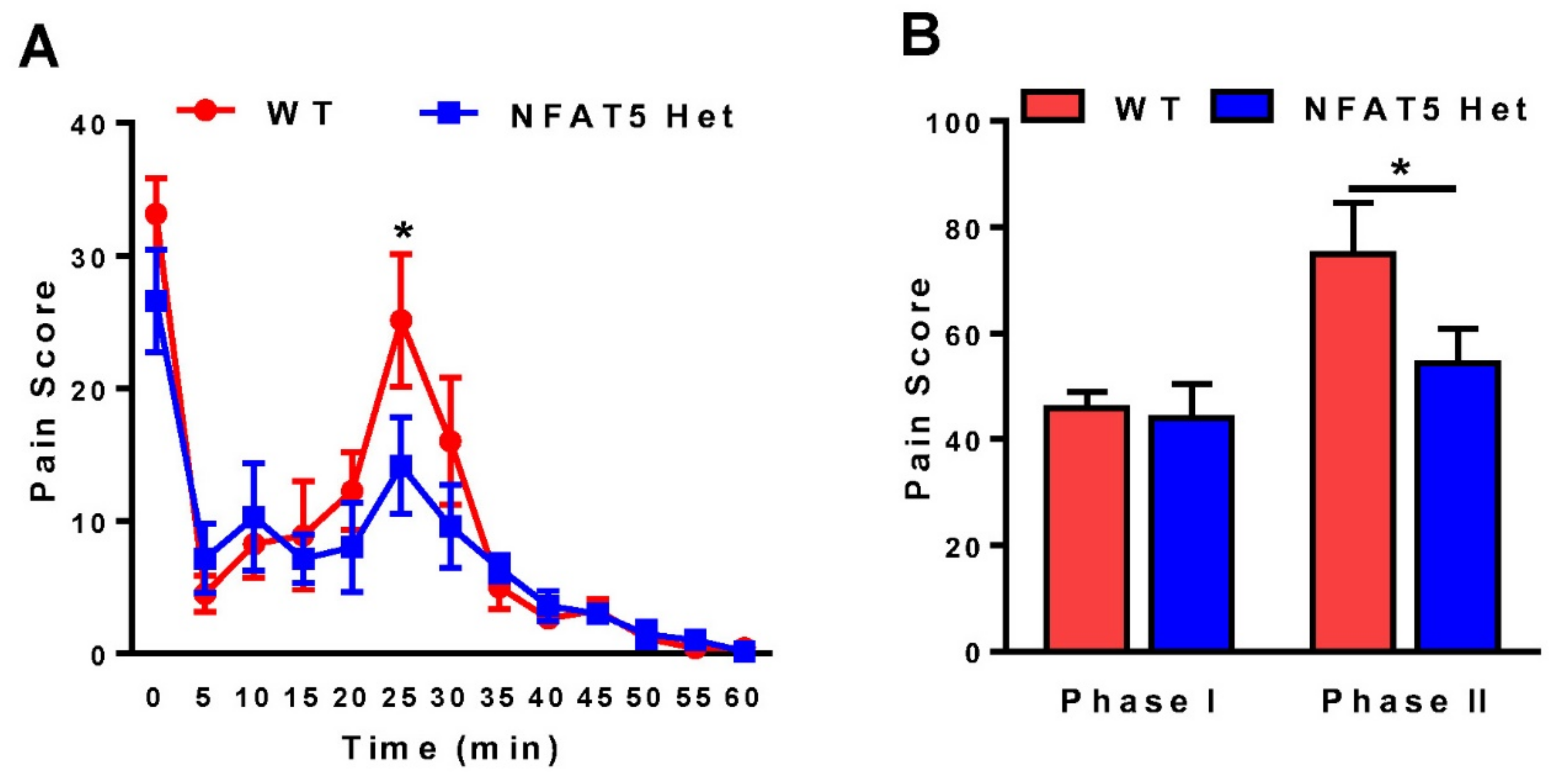{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
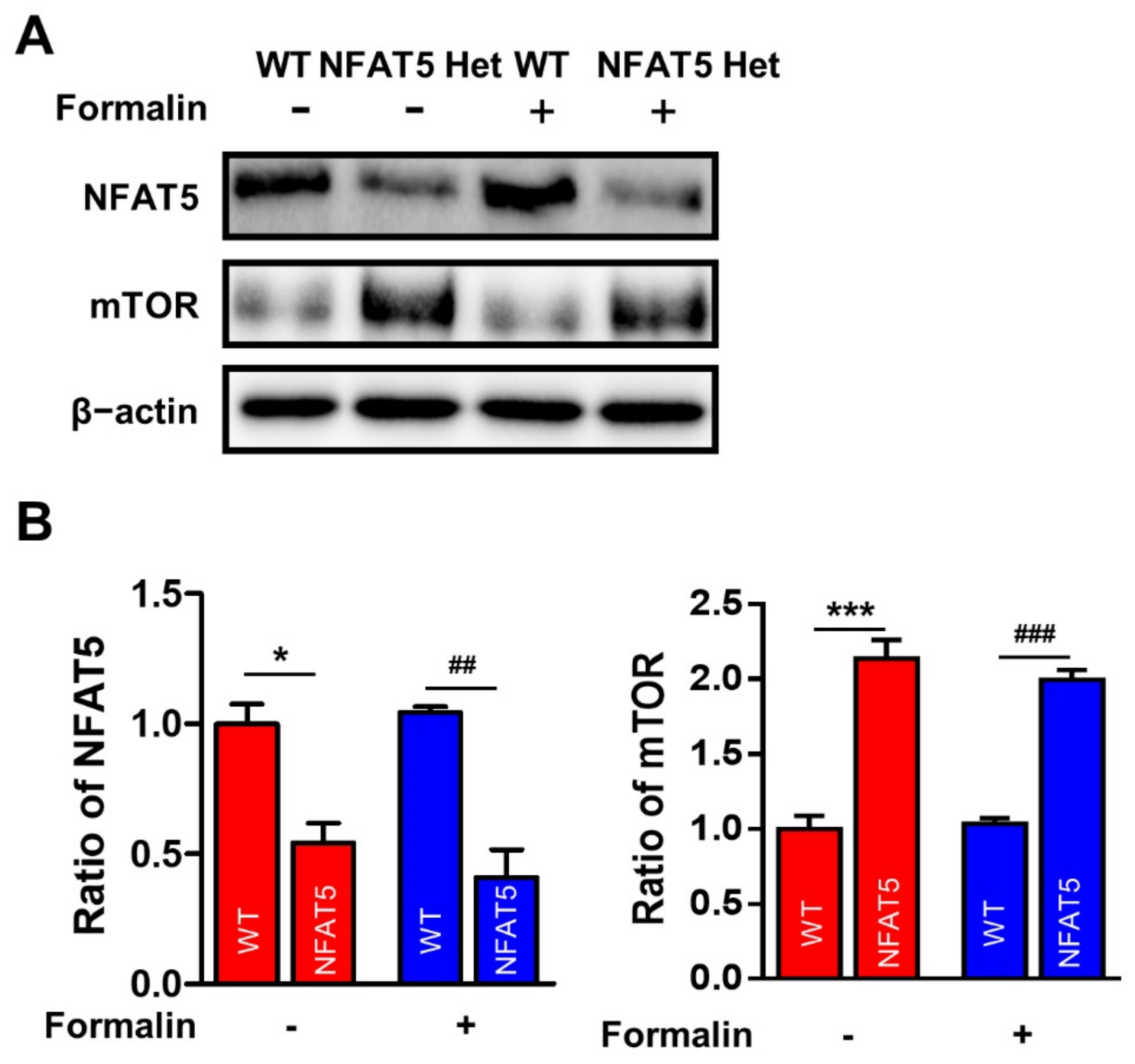
2.1. Formalin-Induced Inflammatory Pain Is Decreased in NFAT5-Het Mice
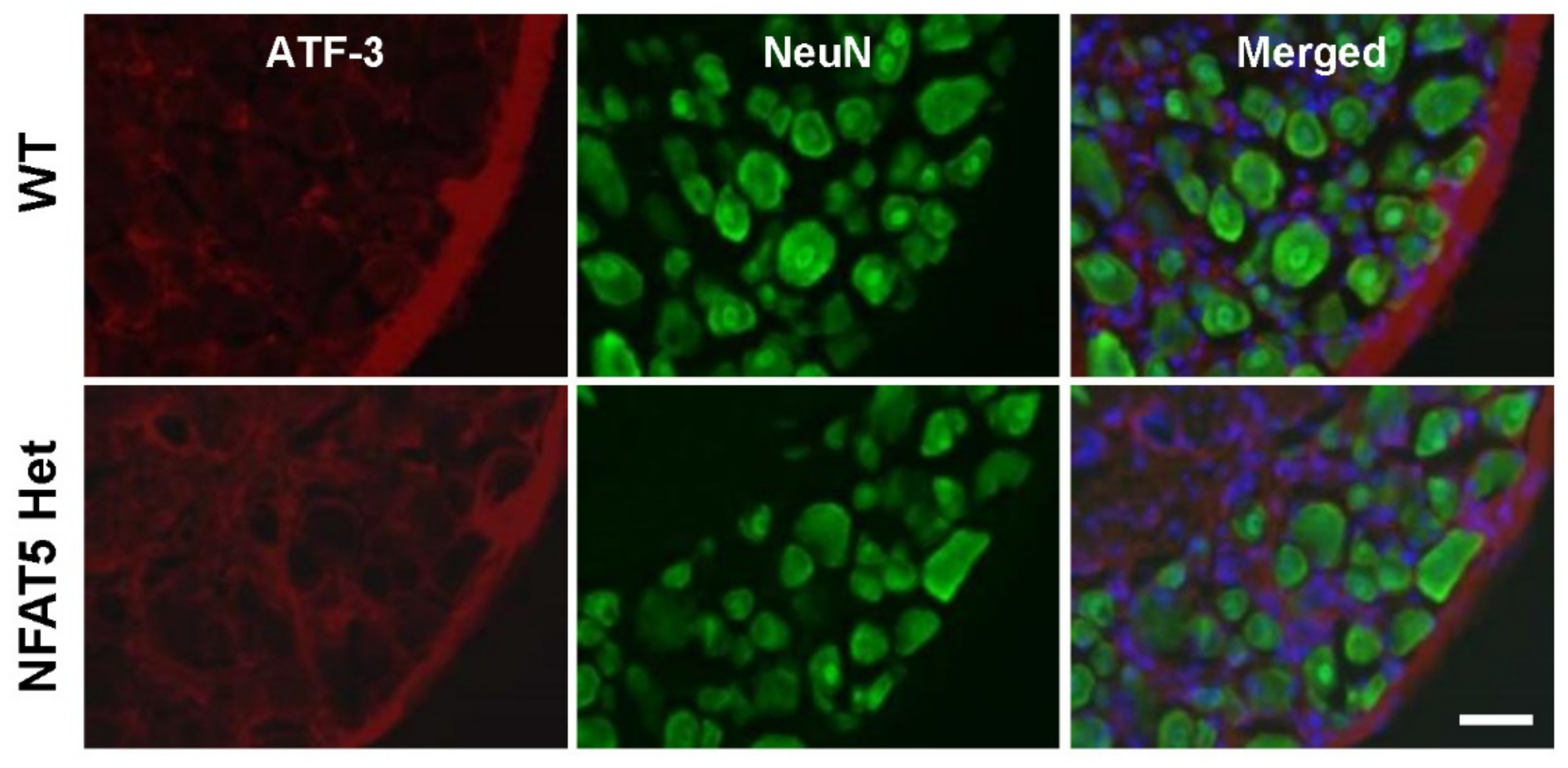
2.2. Activating Transcription Factor 3 Expression in DRG Neurons Is not Altered by NFAT5 Depletion in Formalin-Induced Inflammatory Pain
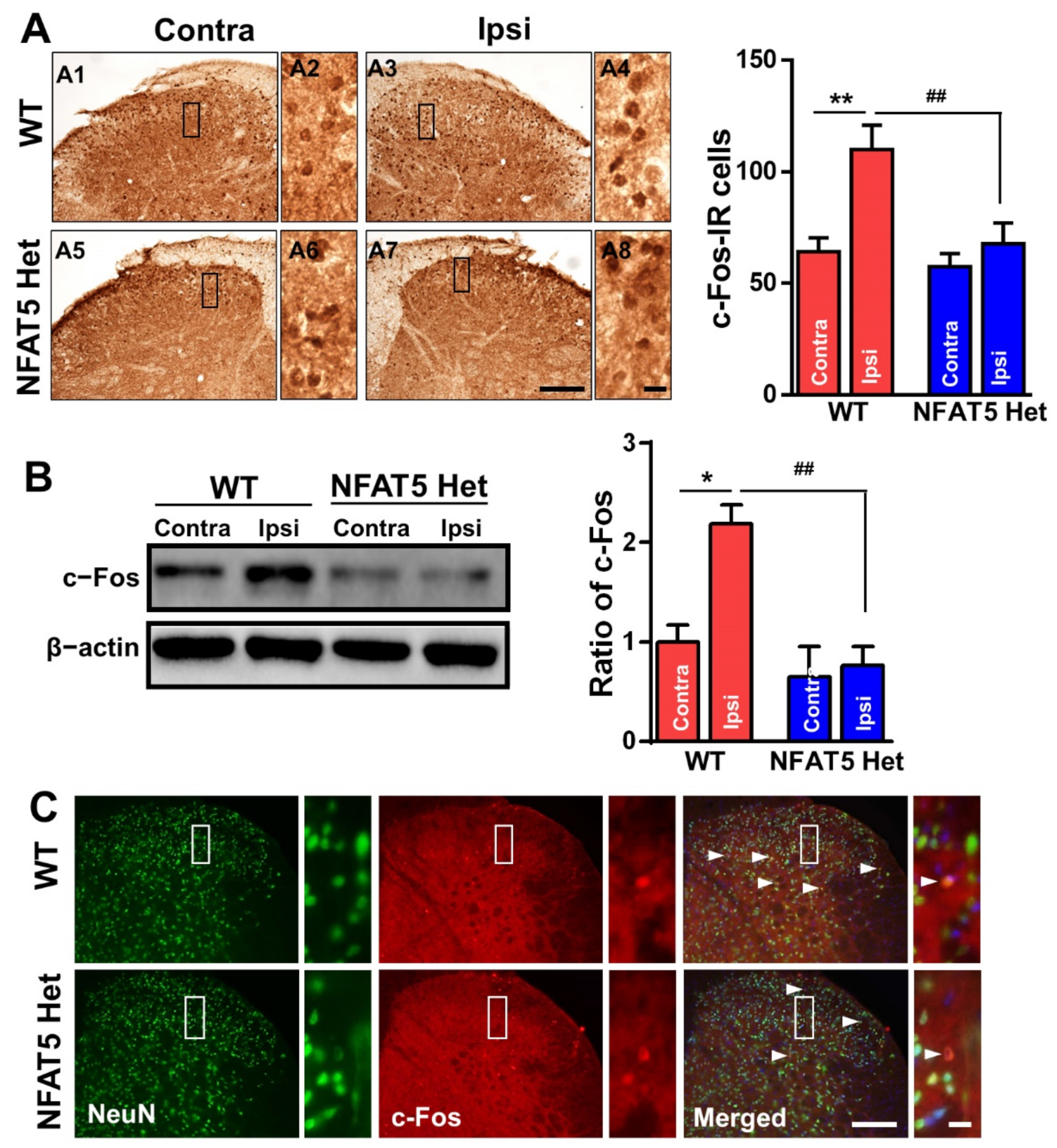
2.3. c-Fos Expression Is not Induced by Formalin Administration in NFAT5-Het Mice
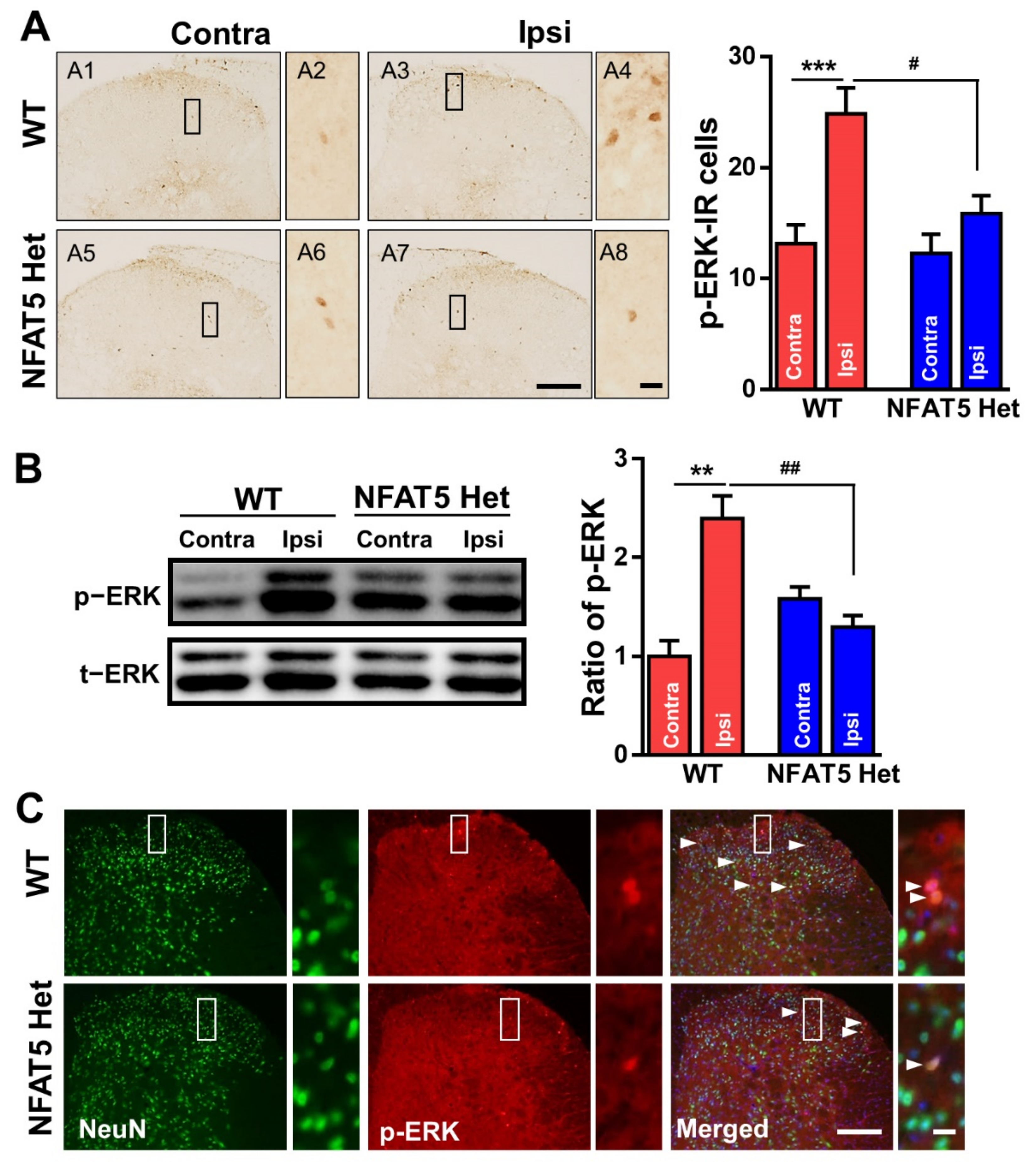
2.4. ERK Is not Activated by Formalin Injection in NFAT5-Het Mice
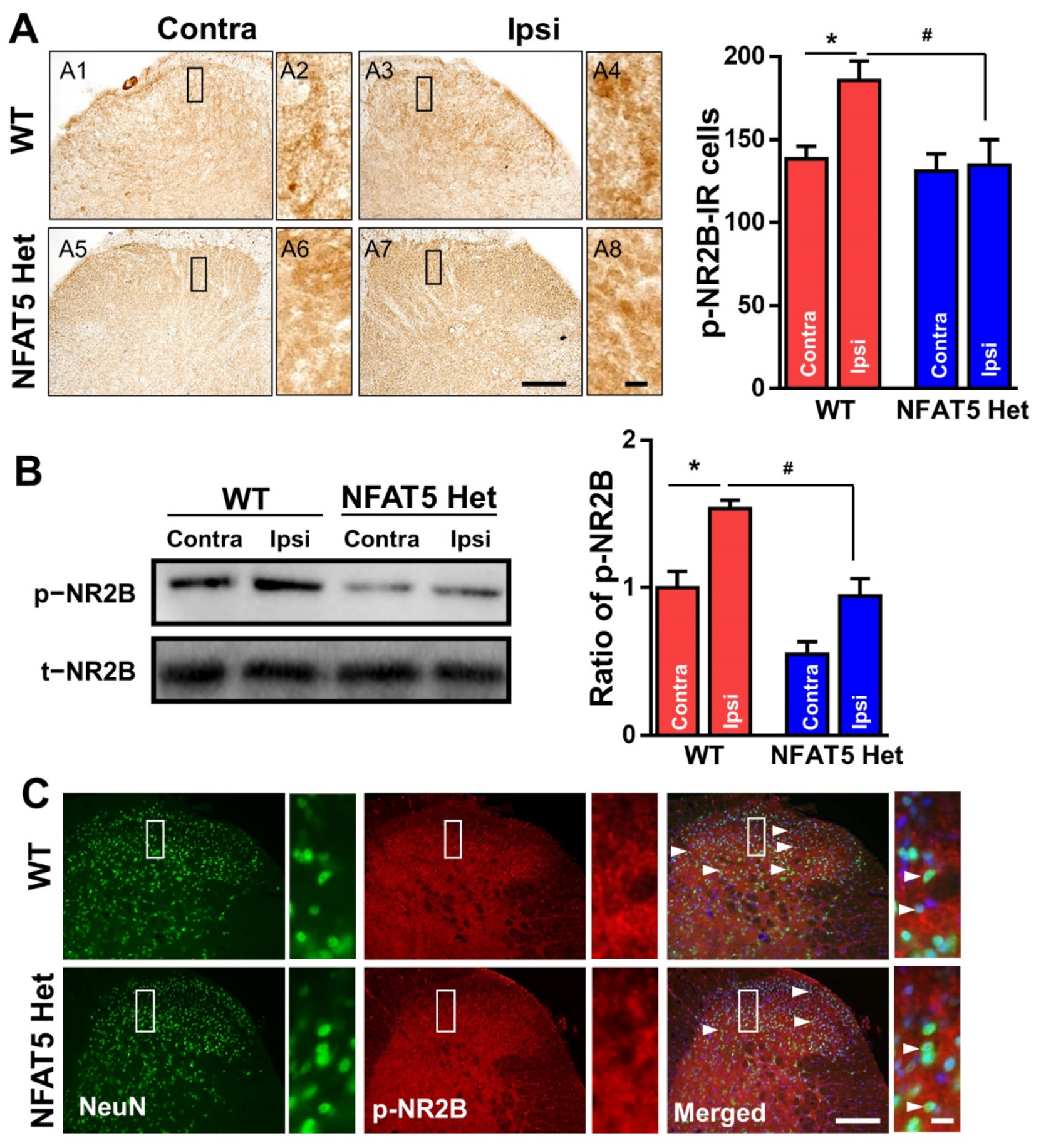
2.5. NR2B Is Phosphorylated in Spinal Neurons in Response to Formalin-Induced Inflammatory Pain
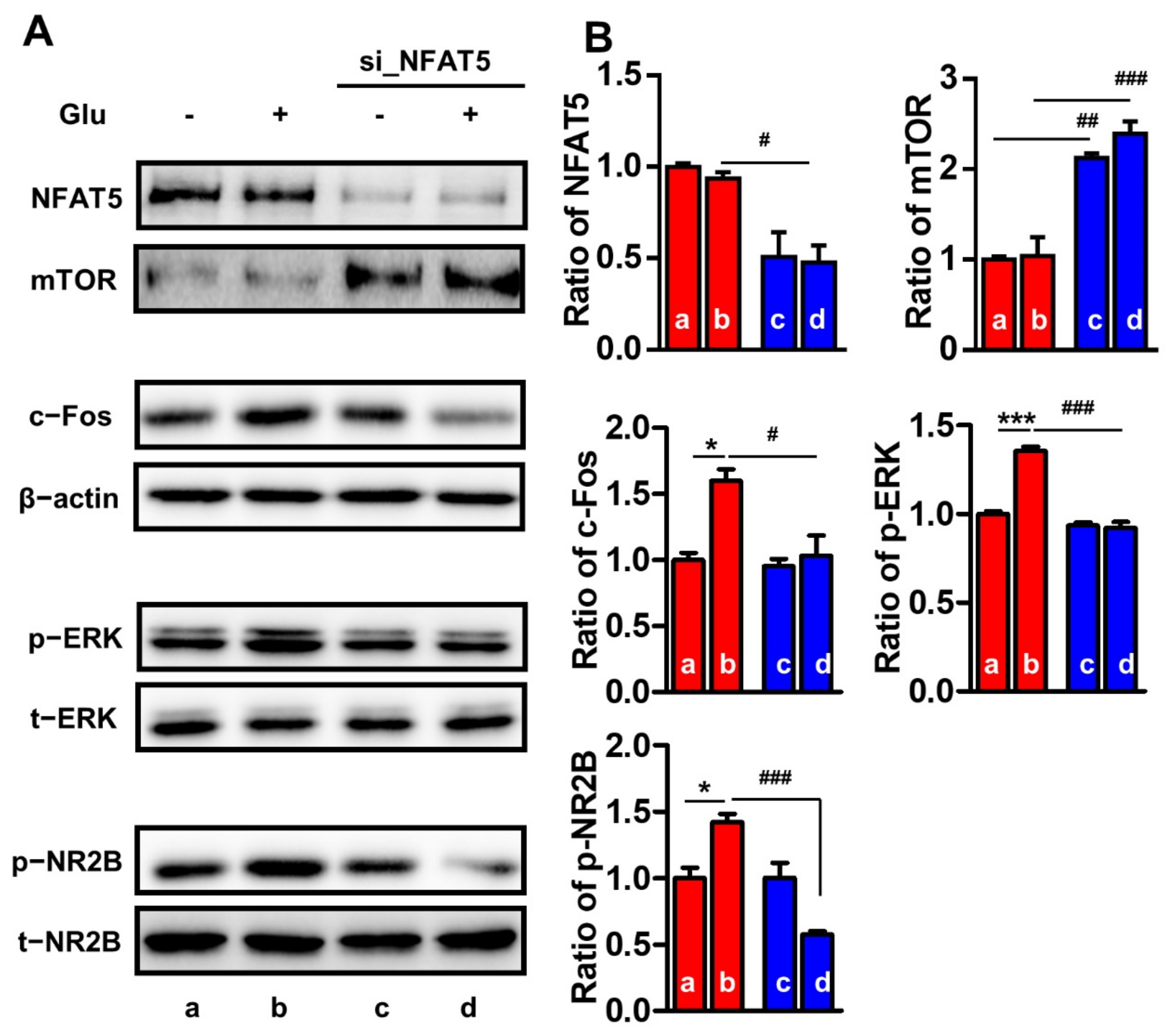
2.6. NFAT5 Upregulates c-Fos, p-ERK, and p-NR2B via mTOR in Glutamate-Stimulated PC12 Cells
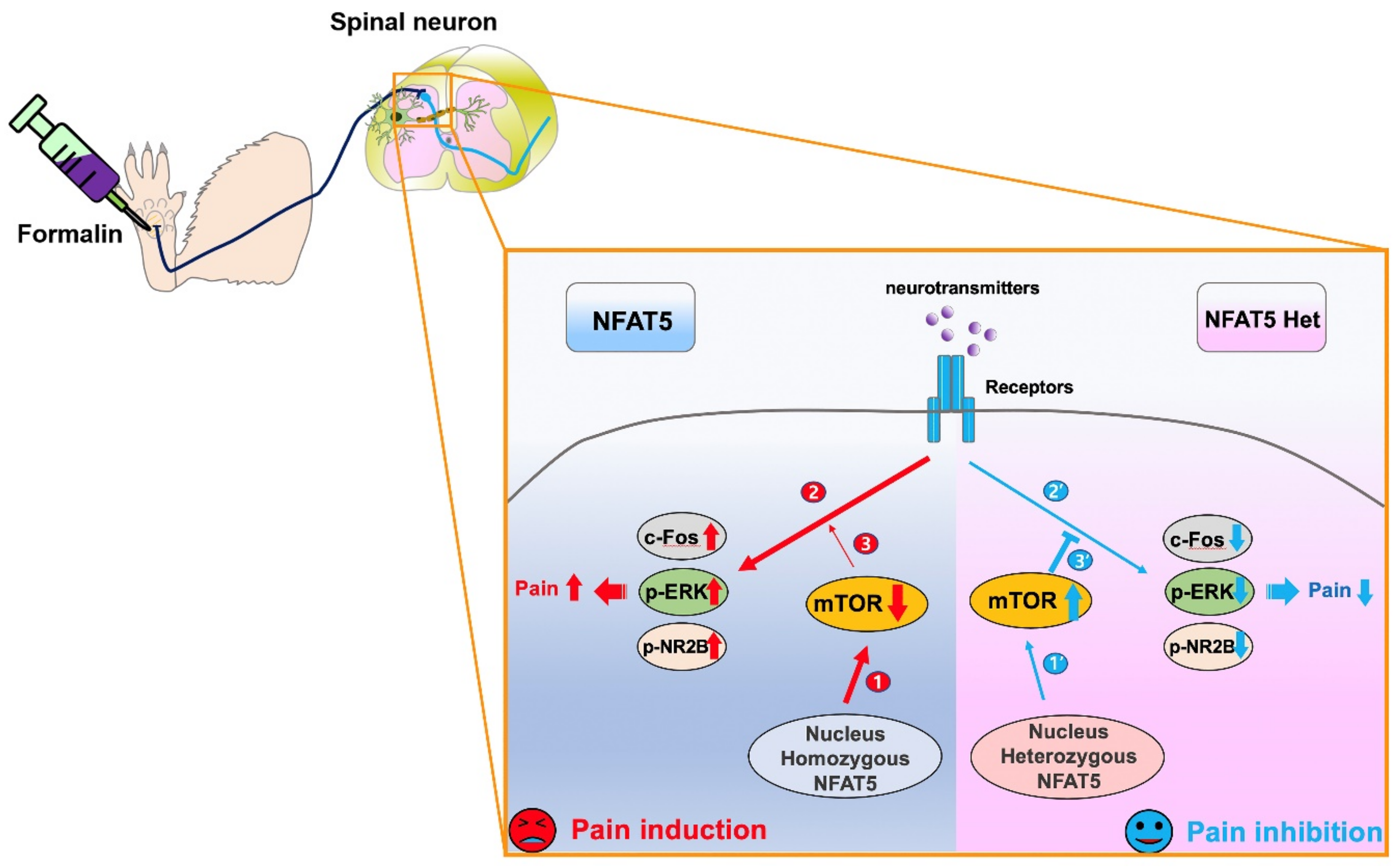
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Formalin Test
4.3. Tissue Processing and Immunostaining
4.4. Western Blot Analysis
4.5. Cell Culture and Nfat5 siRNA Transfection
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Akasha, M.; Anderson, R.R. Thyroxine and Triiodothyronine in Milk of Cows, Goats, Sheep, and Guinea Pigs. Exp. Biol. Med. 1984, 177, 360–371. [Google Scholar] [CrossRef]
- Brugman, C.; Hooff, H.V.D.; Blangé, T. Some aspects of the role of quantum mechanics in the theory of muscle contraction. J. Theor. Biol. 1984, 107, 173–177. [Google Scholar] [CrossRef]
- Lu, P.; Graham, L.; Wang, Y.; Wu, D.; Tuszynski, M. Promotion of Survival and Differentiation of Neural Stem Cells with Fibrin and Growth Factor Cocktails after Severe Spinal Cord Injury. J. Vis. Exp. 2014, 89, e50641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNamara, C.R.; Mandel-Brehm, J.; Bautista, D.M.; Siemens, J.; Deranian, K.L.; Zhao, M.; Hayward, N.J.; Chong, J.A.; Julius, D.; Moran, M.M.; et al. TRPA1 mediates formalin-induced pain. Proc. Natl. Acad. Sci. USA 2007, 104, 13525–13530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, G.-N.; Liu, Y.-L.; Lin, H.-M.; Yang, S.-L.; Feng, Y.-L.; Reid, P.F.; Qin, Z.-H. Involvement of cholinergic system in suppression of formalin-induced inflammatory pain by cobratoxin. Acta Pharmacol. Sin. 2011, 32, 1233–1238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.; Kim, J.-J.; Shin, N.; Yin, Y.; Nan, Y.; Xu, Y.; Hong, J.; Hsu, T.M.; Chung, W.; Ko, Y.; et al. High Omega-3 Polyunsaturated Fatty Acids in fat-1 Mice Reduce Inflammatory Pain. J. Med. Food 2017, 20, 535–541. [Google Scholar] [CrossRef]
- Choi, J.-G.; Kim, J.-M.; Kang, D.-W.; Choi, J.-W.; Park, J.B.; Ahn, S.-H.; Ryu, Y.; Kim, H.-W. Inoculation of Lewis lung carcinoma cells enhances formalin-induced pain behavior and spinal Fos expression in mice. J. Veter. Sci. 2017, 18, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Tjølsen, A.; Berge, O.-G.; Hunskaar, S.; Rosland, J.H.; Hole, K. The formalin test: An evaluation of the method. Pain 1992, 51, 5–17. [Google Scholar] [CrossRef]
- Dubuisson, D.; Dennis, S.G. The formalin test: A quantitative study of the analgesic effects of morphine, meperidine, and brain stem stimulation in rats and cats. Pain 1977, 4, 161–174. [Google Scholar] [CrossRef]
- Wheeler-Aceto, H.; Cowan, A. Standardization of the rat paw formalin test for the evaluation of analgesics. Psychopharmacology 1991, 104, 35–44. [Google Scholar] [CrossRef]
- Mai, L.; Zhu, X.; Huang, F.; He, H.; Fan, W. p38 mitogen-activated protein kinase and pain. Life Sci. 2020, 256, 117885. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, Y.T.J.A. Erythronium japonicum Alleviates Inflammatory Pain by Inhibiting MAPK Activation and by Suppressing NF-κB Activation via ERK/Nrf2/HO-1 Signaling Pathway. Antioxidants 2020, 9, 626. [Google Scholar] [CrossRef] [PubMed]
- Quintero, G.C.; Herrera, J.; Bethancourt, J. Cortical NR2B NMDA subunit antagonism reduces inflammatory pain in male and female rats. J. Pain Res. 2011, 4, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhang, N.; Zhang, R.; Zhao, W.; Chen, Y.; Wang, Z.; Xu, B.; Zhang, M.; Shi, X.; Zhang, Q.; et al. Preemptive intrathecal administration of endomorphins relieves inflammatory pain in male mice via inhibition of p38 MAPK signaling and regulation of inflammatory cytokines. J. Neuroinflammation 2018, 15, 320. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Ma, J.; Yang, X.; He, H.; Wu, H.; Ma, T.; Lü, J. Water-Soluble Polymer Assists N-Methyl-D-Aspartic Acid Receptor 2B siRNA Delivery to Relieve Chronic Inflammatory Pain In Vitro and In Vivo. Pain Res. Manag. 2018, 2018, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhao, W.; Zhang, M.; Xu, B.; Shi, X.; Zhang, Q.; Guo, Y.; Xiao, J.; Chen, D.; Fang, Q.; et al. Analgesic activities of the mixed opioid and NPFF receptors agonist DN-9 in a mouse model of formalin-induced orofacial inflammatory pain. Peptides 2018, 110, 30–39. [Google Scholar] [CrossRef]
- Choi, S.; You, S.; Kim, D.; Choi, S.Y.; Kwon, H.M.; Kim, H.-S.; Hwang, D.; Park, Y.-J.; Cho, C.-S.; Kim, W.-U. Transcription factor NFAT5 promotes macrophage survival in rheumatoid arthritis. J. Clin. Investig. 2017, 127, 954–969. [Google Scholar] [CrossRef] [PubMed]
- Han, E.-J.; Kim, H.Y.; Lee, N.; Kim, N.-H.; Yoo, S.-A.; Kwon, H.M.; Jue, D.-M.; Park, Y.-J.; Cho, C.-S.; De, T.Q. Suppression of NFAT5-mediated inflammation and chronic arthritis by novel κB-binding inhibitors. E. Bio. Medicine. 2017, 18, 261–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, N.-H.; Choi, S.; Han, E.-J.; Hong, B.-K.; Choi, S.Y.; Kwon, H.M.; Hwang, S.-Y.; Cho, C.-S.; Kim, W.-U. The xanthine oxidase-NFAT5 pathway regulates macrophage activation and TLR-induced inflammatory arthritis. Eur. J. Immunol. 2014, 44, 2721–2736. [Google Scholar] [CrossRef] [PubMed]
- Buxadé, M.; Lunazzi, G.; Minguillón, J.; Iborra, S.; Berga-Bolaños, R.; Del Val, M.; Aramburu, J.; López-Rodríguez, C. Gene expression induced by Toll-like receptors in macrophages requires the transcription factor NFAT5. J. Exp. Med. 2012, 209, 379–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.Y.; Li, F.; Wen, Y.B.; Cui, H.X.; Guo, M.L.; Zhang, L.; Zhang, Y.F.; Guo, Y.J.; Guo, Y.X. Melatonin inhibits Sirt1-dependent NAMPT and NFAT5 signaling in chondrocytes to attenuate osteoarthritis. Oncotarget 2017, 8, 55967–55983. [Google Scholar] [CrossRef] [Green Version]
- Aramburu, J.; López-Rodríguez, C. Regulation of inflammatory functions of macrophages and T lymphocytes by NFAT5. Frontiers in immunology 2019, 10, 535. [Google Scholar] [CrossRef]
- Go, W.Y.; Liu, X.; Roti, M.A.; Liu, F.; Ho, S.N. NFAT5/TonEBP mutant mice define osmotic stress as a critical feature of the lymphoid microenvironment. Proc. Natl. Acad. Sci. USA 2004, 101 , 10673–10678. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Jeong, E.A.; Kim, K.E.; Yi, C.-O.; Jin, Z.; Lee, J.E.; Lee, D.H.; Kim, H.J.; Kang, S.S.; Cho, G.J.; et al. TonEBP/NFAT5 haploinsufficiency attenuates hippocampal inflammation in high-fat diet/streptozotocin-induced diabetic mice. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.R.; Kim, D.H.; Kim, K.K.; Jeong, B.; Kang, D.; Lee, T.H.; Park, J.W.; Kwon, H.M.; Lee, B.J. Tonicity-responsive enhancer binding protein (TonEBP) regulates TNF-α-induced hypothalamic inflammation. FEBS Lett. 2019, 593, 2762–2770. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Han, J.; Kim, U.J.; Cha, M.; Um, S.W.; Bai, S.J.; Hong, S.-K.; Lee, B.H. Inhibition of Mammalian Target of Rapamycin (mTOR) Signaling in the Insular Cortex Alleviates Neuropathic Pain after Peripheral Nerve Injury. Front. Mol. Neurosci. 2017, 10, 79. [Google Scholar] [CrossRef]
- Um, S.W.; Kim, M.J.; Leem, J.W.; Bai, S.J.; Lee, B.H. Pain-Relieving Effects of mTOR Inhibitor in the Anterior Cingulate Cortex of Neuropathic Rats. Mol. Neurobiol. 2019, 56, 2482–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asante, O.C.; Wallace, V.C.; Dickenson, A.H. Formalin-Induced Behavioural Hypersensitivity and Neuronal Hyperexcitability are Mediated by Rapid Protein Synthesis at the Spinal Level. Mol. Pain 2009, 5, 1744–8069. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, L.; Wang, H.; Su, Z.; Pang, X. Neuroinflammation and central PI3K/Akt/mTOR signal pathway contribute to bone cancer pain. Mol. Pain 2019, 15, 1744806919830240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shortland, P.J.; Baytug, B.; Krzyzanowska, A.; McMahon, S.B.; Priestley, J.V.; Averill, S. ATF3 expression in L4 dorsal root ganglion neurons after L5 spinal nerve transection. Eur. J. Neurosci. 2006, 23, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Barr, G.A. Formalin-Induced c-fos Expression in the Brain of Infant Rats. J. Pain 2011, 12, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Kültz, D.; Burg, M. Evolution of osmotic stress signaling via MAP kinase cascades. J. Exp. Biol. 1998, 201, 3015–3021. [Google Scholar]
- Pearson, G.; Robinson, F.; Gibson, T.B.; Xu, B.-E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-Activated Protein (MAP) Kinase Pathways: Regulation and Physiological Functions*. Endocr. Rev. 2001, 22, 153–183. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Song, N.; Liu, F.; Lin, J.; Liu, M.; Huang, C.; Liao, D.; Zhou, C.; Wang, H.; Shen, J. Activation of mitogen-activated protein kinases in satellite glial cells of the trigeminal ganglion contributes to substance P-mediated inflammatory pain. Int. J. Oral Sci. 2019, 11, 24. [Google Scholar] [CrossRef]
- Khan, S.; Shehzad, O.; Chun, J.; Kim, Y.S. Mechanism underlying anti-hyperalgesic and anti-allodynic properties of anomalin in both acute and chronic inflammatory pain models in mice through inhibition of NF-κB, MAPKs and CREB signaling cascades. Eur. J. Pharmacol. 2013, 718, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Petrenko, A.B.; Yamakura, T.; Baba, H.; Shimoji, K. The Role of N-Methyl-d-Aspartate (NMDA) Receptors in Pain: A Review. Anesth. Analg. 2003, 97, 1108–1116. [Google Scholar] [CrossRef] [PubMed]
- Lipton, J.O.; Sahin, M. The Neurology of mTOR. Neuron 2014, 84, 275–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hay, N.; Sonenberg, N. Upstream and downstream of mTOR. Genes Dev. 2004, 18, 1926–1945. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Zhou, H.; Huang, S.; Liu, J. MiR-1906 attenuates neuropathic pain in rats by regulating the TLR4/mTOR/ Akt signaling pathway. Transl. Neurosci. 2019, 10, 175–179. [Google Scholar] [CrossRef] [Green Version]
- Duan, Z.; Li, J.; Pang, X.; Wang, H.; Su, Z. Blocking Mammalian Target of Rapamycin (mTOR) Alleviates Neuropathic Pain Induced by Chemotherapeutic Bortezomib. Cell. Physiol. Biochem. 2018, 48, 54–62. [Google Scholar] [CrossRef]
- Lutz, B.M.; Nia, S.; Xiong, M.; Tao, Y.-X.; Bekker, A. mTOR, a New Potential Target for Chronic Pain and Opioid-Induced Tolerance and Hyperalgesia. Mol. Pain 2015, 11, 32. [Google Scholar] [CrossRef] [Green Version]
- Sluka, K.A.; Jordan, H.H.; Westlund, K.N. Reduction in joint swelling and hyperalgesia following post-treatment with a non-NMDA glutamate receptor antagonist. Pain 1994, 59, 95–100. [Google Scholar] [CrossRef]
- Walker, K.; Reeve, A.; Bowes, M.; Winter, J.; Wotherspoon, G.; Davis, A.; Schmid, P.; Gasparini, F.; Kuhn, R.; Urban, L. mGlu5 receptors and nociceptive function II. mGlu5 receptors functionally expressed on peripheral sensory neurones mediate inflammatory hyperalgesia. Neuropharmacol. 2001, 40, 10–19. [Google Scholar] [CrossRef]
- Hunt, S.P.; Pini, A.; Evan, G. Induction of c-fos-like protein in spinal cord neurons following sensory stimulation. Nat. Cell Biol. 1987, 328, 632–634. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, M. c-Fos and the changing face of pain. Trends Neurosci. 1990, 13, 439–440. [Google Scholar] [CrossRef]
- Munglani, R.; Fleming, B.G.; Hunt, S.P. Remembrance of times past: The significance of c-fos in pain. Br. J. Anaesth. 1996, 76, 1–4. [Google Scholar] [CrossRef]
- Lee, J.-W.; Nam, H.; Kim, L.E.; Jeon, Y.; Min, H.; Ha, S.; Lee, Y.; Kim, S.-Y.; Lee, S.J.; Kim, E.-K.; et al. TLR4 (toll-like receptor 4) activation suppresses autophagy through inhibition of FOXO3 and impairs phagocytic capacity of microglia. Autophagy 2019, 15, 753–770. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.; Yi, M.-H.; Ko, Y.; Kim, H.-W.; Seo, J.H.; Lee, Y.H.; Lee, W.; Kim, D.W. Expression of LC3 and Beclin 1 in the spinal dorsal horn following spinal nerve ligation-induced neuropathic pain. Brain Res. 2013, 1519, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.-J.; Zhang, L.; Ji, R.-R. Spinal injection of TNF-α-activated astrocytes produces persistent pain symptom mechanical allodynia by releasing monocyte chemoattractant protein-1. Glia 2010, 58, 1871–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Wu, X.; Fossett, N. Upregulation of the Drosophila Friend of GATA Gene u-shaped by JAK/STAT Signaling Maintains Lymph Gland Prohemocyte Potency. Mol. Cell. Biol. 2009, 29, 6086–6096. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gwon, D.H.; Kim, S.I.; Lee, S.H.; Noh, C.; Kim, Y.; Yun, S.; Lee, W.H.; Oh, J.Y.; Kim, D.W.; Hong, J.; et al. NFAT5 Deficiency Alleviates Formalin-Induced Inflammatory Pain Through mTOR. Int. J. Mol. Sci. 2021, 22, 2587. https://doi.org/10.3390/ijms22052587
Gwon DH, Kim SI, Lee SH, Noh C, Kim Y, Yun S, Lee WH, Oh JY, Kim DW, Hong J, et al. NFAT5 Deficiency Alleviates Formalin-Induced Inflammatory Pain Through mTOR. International Journal of Molecular Sciences. 2021; 22(5):2587. https://doi.org/10.3390/ijms22052587
Chicago/Turabian StyleGwon, Do Hyeong, Song I. Kim, Seoung Hun Lee, Chan Noh, Yeojung Kim, Sangwon Yun, Won Hyung Lee, Jun Young Oh, Dong Woon Kim, Jinpyo Hong, and et al. 2021. "NFAT5 Deficiency Alleviates Formalin-Induced Inflammatory Pain Through mTOR" International Journal of Molecular Sciences 22, no. 5: 2587. https://doi.org/10.3390/ijms22052587