
Senescence Marker Protein 30 (SMP30): A Novel Pan-Species Diagnostic Marker for the Histopathological Diagnosis of Breast Cancer in Humans and Animals

 , and
, and
Abstract
:1. Introduction
2. Results
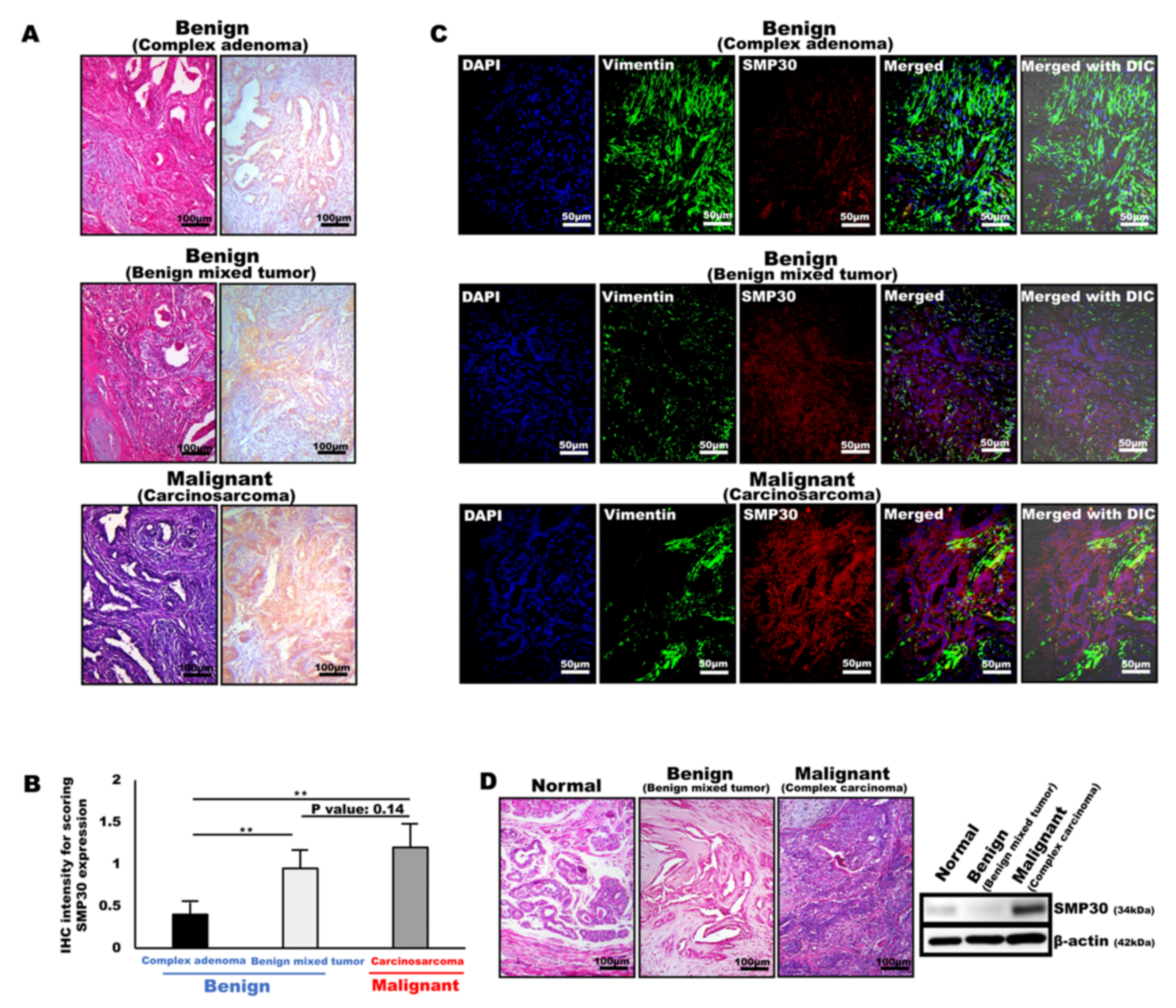
2.1. Elevated SMP30 Expression Levels in Mammary Tumors Correspond to Their Malignancy
2.2. SMP30 Expression Is Highly Associated with the Differentiation of the Tumor Cells in Various Types of Mammary Gland Tumors
2.3. SMP30 Expressions of Canine Primary Carcinoma Cells Are Colocalized with Pan-Cytokeratin, a Marker of Epithelial Origin in Mammary Glands
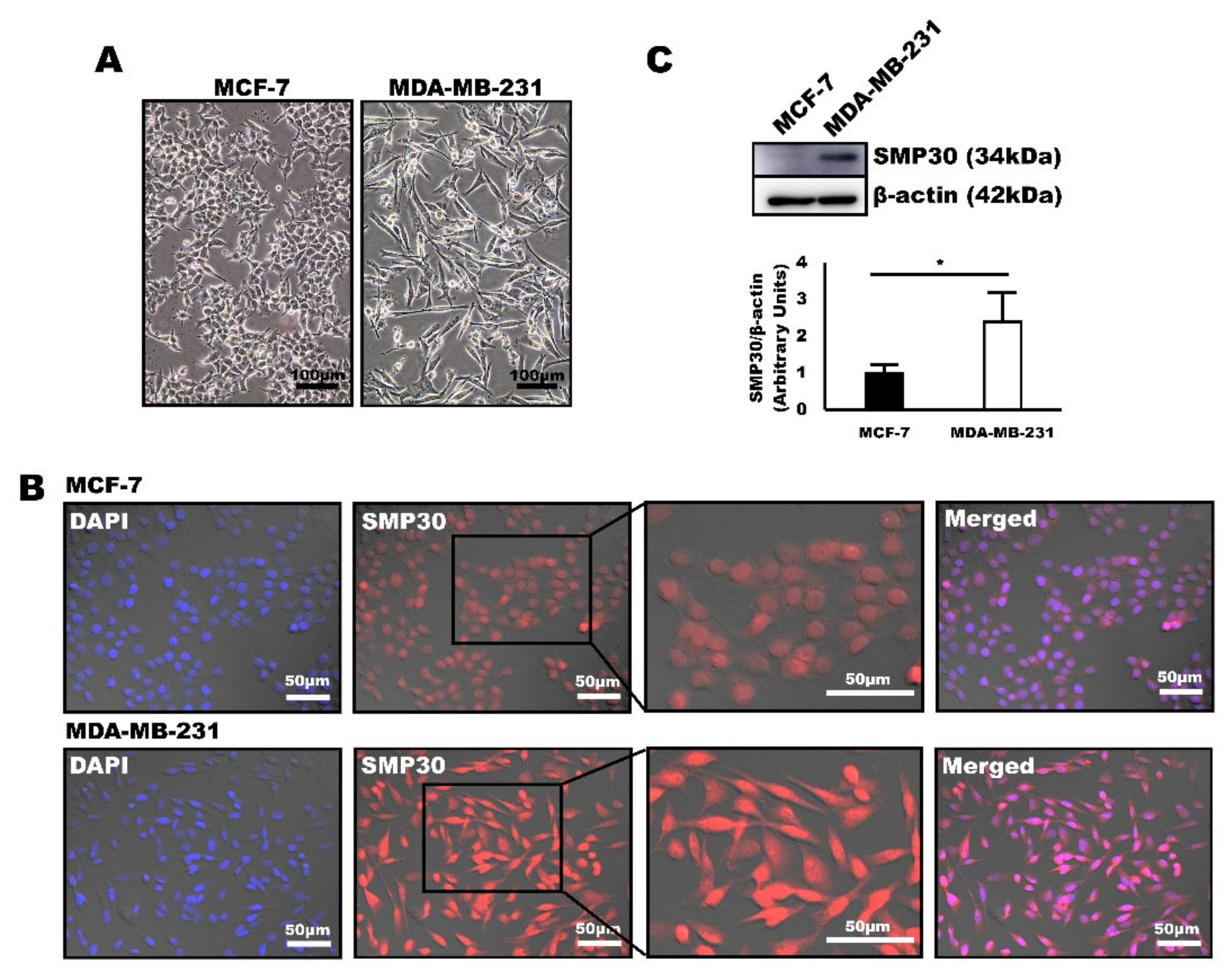
2.4. SMP30 Levels Increased with Malignancy in Human Breast Cancer Cells
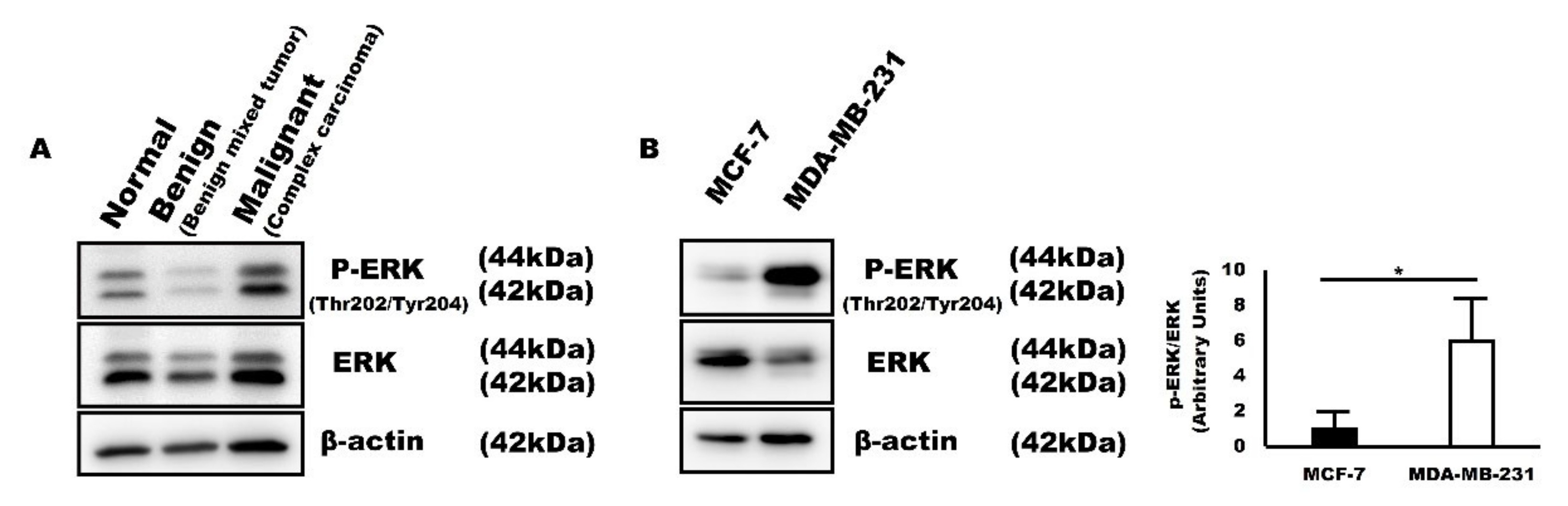
2.5. P-ERK Expression Corresponds to SMP30 Expression in Canine Mammary Gland Tumors and Human Breast Cancer Cells
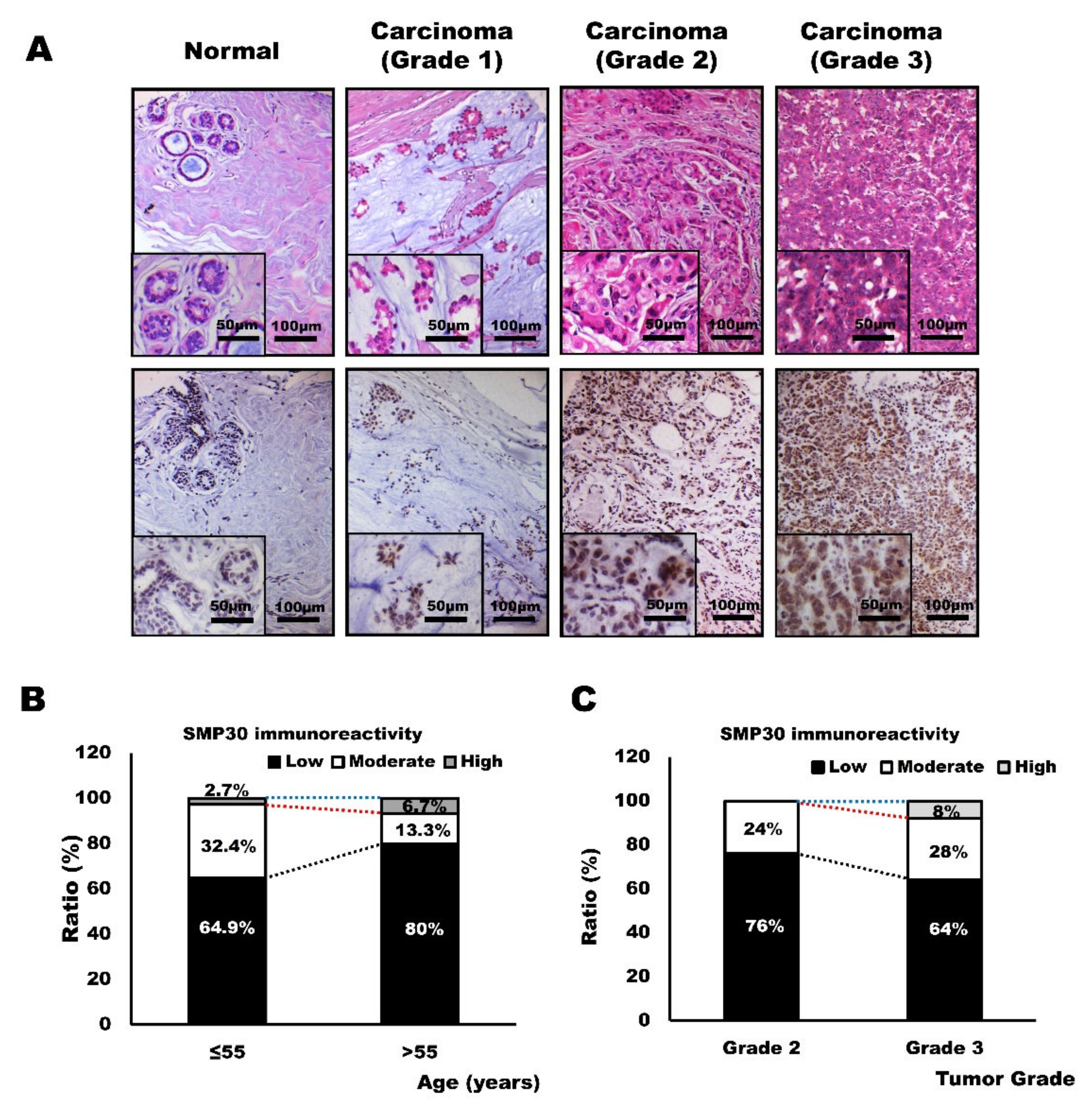
2.6. SMP30 Expression Levels in Human Breast Cancer Tissues and Its Correlation with Age and Histopathological Grades of Carcinomas
3. Discussion
4. Materials and Methods
4.1. Tissue Samples and Histopathologic Examination
4.2. Immunohistochemistry and Immunoreactivity Evaluation
4.3. Primary Cell Isolation of Canine Mammary Adenocarcinoma
4.4. Cell Culture
4.5. Western Blot Analysis
4.6. Immunofluorescence
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BID | Bis in die |
| DAB | 3,3-diaminobenzidine |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DPBS | Dulbecco’s phosphate-buffered saline |
| ER | Estrogen receptor |
| FBS | Fetal bovine serum |
| FFPE | Formalin fixed, paraffin embedded |
| H&E | Hematoxylin and eosin |
| HER2 | Human epidermal growth factor 2 |
| HRP | Horseradish peroxidase |
| LPS | Lipopolysaccharide |
| NO | Nitric oxide |
| PBS | Phosphate-buffered saline |
| PFA | Paraformaldehyde |
| PR | Progesterone receptor |
| PVDF | Polyvinylidene difluoride |
| ROS | Reactive oxygen species |
| SDS-PAGE | Sodium dodecyl sulfate–polyacrylamide gel electrophoresis |
| SMP30 | Senescence marker protein 30 |
| TBS | Tris-buffered saline |
| TMA | Tissue microarray |
| TNF-α | Tumor necrosis factor alpha |
References
- Rahman, S.A.; Al-Marzouki, A.; Otim, M.; Khalil Khayat, N.E.H.; Yousuf, R.; Rahman, P. Awareness about breast cancer and breast self-examination among female students at the university of Sharjah: A cross-sectional study. Asian Pac. J. Cancer Prev. 2019, 20, 1901–1908. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.M.; Montazeri, A.; Mohagheghi, M.A.; Jarrahi, A.M.; Harirchi, I.; Najafi, M.; Ebrahimi, M. Breast cancer in Iran: An epidemiological review. Breast J. 2007, 13, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Kubo, K.; Matsuyama, S.; Katayama, K.; Tsutsumi, C.; Yonezawa, K.; Shimada, T.; Kotani, T.; Sakuma, S.; Ohashi, F.; Takamori, Y. Frequent Expression of the c-kit Proto-Oncogene in Canine Malignant Mammary Tumor. J. Vet. Med. Sci. 1998, 60, 1335–1340. [Google Scholar] [CrossRef] [Green Version]
- Philibert, J.C.; Snyder, P.W.; Glickman, N.; Glickman, L.T.; Knapp, D.W.; Waters, D.J. Influence of host factors on survival in dogs with malignant mammary gland tumors. J. Vet. Intern. Med. 2003, 17, 102–106. [Google Scholar] [CrossRef]
- Moe, L. Population-based incidence of mammary tumours in some dog breeds. J. Reprod. Fertil. Suppl. 2001, 57, 439–443. [Google Scholar] [PubMed]
- Goldschmidt, M.H.; Peña, L.; Zappulli, V. Tumors of the Mammary gland. In Tumors in Domestic Animal, 5th ed.; Meuten, D.J., Ed.; Wiley-Blackwell: Ames, IA, USA, 2018; pp. 723–765. [Google Scholar]
- Wang, L. Early Diagnosis of Breast Cancer. Sensors 2017, 17, 1572. [Google Scholar] [CrossRef] [PubMed]
- Novosad, C.A. Principles of treatment for mammary gland tumors. Clin. Tech. Small Anim. Pract. 2003, 18, 107–109. [Google Scholar] [CrossRef]
- Spitale, A.; Mazzola, P.; Soldini, D.; Mazzucchelli, L.; Bordoni, A. Breast cancer classification according to immunohistochemical markers: Clinicopathologic features and short-term survival analysis in a population-based study from the south of Switzerland. Ann. Oncol. 2009, 20, 628–635. [Google Scholar] [CrossRef]
- Van de Ven, S.; Smit, V.T.; Dekker, T.J.; Nortier, J.W.; Kroep, J.R. Discordances in ER, PR and HER2 receptors after neoadjuvant chemotherapy in breast cancer. Cancer Treat. Rev. 2011, 37, 422–430. [Google Scholar] [CrossRef]
- Abdelmegeed, S.M.; Mohammed, S. Canine mammary tumors as a model for human disease. Oncol. Lett. 2018, 15, 8195–8205. [Google Scholar] [CrossRef] [Green Version]
- Dowsett, M.; Dunbier, A.K. Emerging Biomarkers and New Understanding of Traditional Markers in Personalized Therapy for Breast Cancer. Clin. Cancer. Res. 2008, 14, 8019–8026. [Google Scholar] [CrossRef] [Green Version]
- Hammond, M.E.; Hayes, D.F.; Dowsett, M.; Allred, D.C.; Hagerty, K.L.; Badve, S.; Fitzgibbons, P.L.; Francis, G.; Goldstein, N.S.; Hayes, M.; et al. American Society of Clinical Oncology/College of American Pathologists guideline recommendations for immunohistochemical testing of estrogen and progesterone receptors in breast cancer. Arch. Pathol. Lab. Med. 2010, 134, 907–922. [Google Scholar] [CrossRef]
- Toniti, W.; Buranasinsup, S.; Kongcharoen, A.; Charoonrut, P.; Puchadapirom, P.; Kasorndorkbua, C. Immunohistochemical determination of estrogen and progesterone receptors in canine mammary tumors. Asian Pac. J. Cancer Prev. 2009, 10, 907–911. [Google Scholar]
- Peña, L.; Gama, A.; Goldschmidt, M.H.; Abadie, J.; Benazzi, C.; Castagnaro, M.; Díez, L.; Gärtner, F.; Hellmén, E.; Kiupel, M.; et al. Canine Mammary Tumors: A Review and Consensus of Standard Guidelines on Epithelial and Myoepithelial Phenotype Markers, HER2, and Hormone Receptor Assessment Using Immunohistochemistry. Vet. Pathol. 2014, 51, 127–145. [Google Scholar] [CrossRef]
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nature 2001, 411, 342–348. [Google Scholar] [CrossRef]
- Gerl, R.; Vaux, D.L. Apoptosis in the development and treatment of cancer. Carcinogenesis 2005, 26, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Kasibhatla, S.; Tseng, B. Why Target Apoptosis in Cancer Treatment? Mol. Cancer Ther. 2003, 2, 573–580. [Google Scholar]
- Scott, S.H.; Bahnson, B.J. Senescence Marker Protein 30: Functional and Structural Insights to its Unknown Physiological Function. Biomol. Concepts 2011, 2, 469–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, M. The anti-apoptotic effect of regucalcin is mediated through multisignaling pathways. Apoptosis 2013, 18, 1145–1153. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, N.; Ishigami, A.; Kondo, Y. Pathophysiological significance of senescence marker protein-30. Geriatr. Gerontol. Int. 2010, 10 (Suppl. 1), S88–S98. [Google Scholar] [CrossRef]
- Park, J.K.; Hong, I.H.; Ki, M.R.; Chung, H.Y.; Ishigami, A.; Ji, A.R.; Goo, M.J.; Kim, D.H.; Kwak, J.H.; Min, C.W.; et al. Vitamin C deficiency increases the binucleation of hepatocytes in SMP30 knock-out mice. J. Gastroenterol. Hepatol. 2010, 25, 1769–1776. [Google Scholar] [CrossRef]
- Son, T.G.; Kim, S.J.; Kim, K.; Kim, M.S.; Chung, H.Y.; Lee, J. Cytoprotective roles of senescence marker protein 30 against intracellular calcium elevation and oxidative stress. Arch. Pharm. Res. 2008, 31, 872–877. [Google Scholar] [CrossRef]
- Choo, J.; Heo, G.; Kim, S.J.; Lee, Y.; Ishigami, A.; Maruyama, N.; Chung, H.Y.; Im, E. Senescence marker protein 30 protects intestinal epithelial cells against inflammation-induced cell death by enhancing Nrf2 activity. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3668–3678. [Google Scholar] [CrossRef]
- Kadowaki, S.; Shishido, T.; Sasaki, T.; Sugai, T.; Narumi, T.; Honda, Y.; Otaki, Y.; Kinoshita, D.; Takahashi, T.; Nishiyama, S.; et al. Deficiency of Senescence Marker Protein 30 Exacerbates Cardiac Injury after Ischemia/Reperfusion. Int. J. Mol. Sci. 2016, 17, 542. [Google Scholar] [CrossRef]
- Izumi, T.; Yamaguchi, M. Overexpression of regucalcin suppresses cell death and apoptosis in cloned rat hepatoma H4-II-E cells induced by lipopolysaccharide, PD 98059, dibucaine, or Bay K 8644. J. Cell. Biochem. 2004, 93, 598–608. [Google Scholar] [CrossRef]
- Kim, H.S.; Son, T.G.; Park, H.R.; Lee, Y.; Jung, Y.; Ishigami, A.; Lee, J. Senescence marker protein 30 deficiency increases Parkinson’s pathology by impairing astrocyte activation. Neurobiol. Aging 2013, 34, 1177–1183. [Google Scholar] [CrossRef]
- Son, T.G.; Park, H.R.; Kim, S.J.; Kim, K.; Kim, M.S.; Ishigami, A.; Handa, S.; Maruyama, N.; Chung, H.Y.; Lee, J. Senescence marker protein 30 is up-regulated in kainate-induced hippocampal damage through ERK-mediated astrocytosis. J. Neurosci. Res. 2009, 87, 2890–2897. [Google Scholar] [CrossRef]
- Salaroglio, I.C.; Mungo, E.; Gazzano, E.; Kopecka, J.; Riganti, C. ERK is a Pivotal Player of Chemo-Immune-Resistance in Cancer. Int. J. Mol. Sci. 2019, 20, 2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sar, P.; Peter, R.; Rath, B.; Das Mohapatra, A.; Mishra, S.K. 3,3′5 Triiodo L Thyronine Induces Apoptosis in Human Breast Cancer MCF-7 cells, Repressing SMP30 Expression through Negative Thyroid Response Elements. PLoS ONE 2011, 6, e20861. [Google Scholar] [CrossRef] [Green Version]
- Maia, C.; Santos, C.; Schmitt, F.; Socorro, S. Regucalcin is under-expressed in human breast and prostate cancers: Effect of sex steroid hormones. J. Cell. Biochem. 2009, 107, 667–676. [Google Scholar] [CrossRef]
- Sommers, C.L.; Byers, S.W.; Thompson, E.W.; Torri, J.A.; Gelmann, E.P. Differentiation state and invasiveness of human breast cancer cell lines. Breast Cancer Res. Treat. 1994, 31, 325–335. [Google Scholar] [CrossRef]
- Benz, C.C. Impact of aging on the biology of breast cancer. Crit. Rev. Oncol. Hematol. 2008, 66, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Yancik, R.; Wesley, M.N.; Ries, L.A.; Havlik, R.J.; Edwards, B.K.; Yates, J.W. Effect of age and comorbidity in postmenopausal breast cancer patients aged 55 years and older. JAMA 2001, 285, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Kaszak, I.; Ruszczak, A.; Kanafa, S.; Kacprzak, K.; Król, M.; Jurka, P. Current biomarkers of canine mammary tumors. Acta Vet. Scand. 2018, 60, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatloukal, J.; Lorenzová, J.; Tichý, F.; Nečas, A.; Kecová, H.; Kohout, P. Breed and Age as Risk Factors for Canine Mammary Tumours. Acta Vet. Brno 2005, 74, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Marques, C.; Correia, J.; Ferreira, F. HER2-positive feline mammary carcinoma. Aging 2016, 8, 1574–1575. [Google Scholar] [CrossRef] [Green Version]
- Sorenmo, K. Canine mammary gland tumors. Vet. Clin. N. Am. Small Anim. Pract. 2003, 33, 573–596. [Google Scholar] [CrossRef]
- Sorenmo, K.U.; Rasotto, R.; Zappulli, V.; Goldschmidt, M.H. Development, Anatomy, Histology, Lymphatic Drainage, Clinical Features, and Cell Differentiation Markers of Canine Mammary Gland Neoplasms. Vet. Pathol. 2011, 48, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dooijeweert, C.; Deckers, I.A.G.; Baas, I.O.; van der Wall, E.; van Diest, P.J. Hormone- and HER2-receptor assessment in 33,046 breast cancer patients: A nationwide comparison of positivity rates between pathology laboratories in the Netherlands. Breast Cancer Res. Treat. 2019, 175, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Al-Mansour, M.A.; Kubba, M.A.G.; Al-Azreg, S.A.; Dribika, S.A. Comparative histopathology and immunohistochemistry of human and canine mammary tumors. Open Vet. J. 2018, 8, 243–249. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB J. 2003, 17, 1195–1214. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Masutomi, H.; Noda, Y.; Ozawa, Y.; Takahashi, K.; Handa, S.; Maruyama, N.; Shimizu, T.; Ishigami, A. Senescence marker protein-30/superoxide dismutase 1 double knockout mice exhibit increased oxidative stress and hepatic steatosis. FEBS Open Bio 2014, 4, 522–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, K.; Kondo, Y.; Sekiya, M.; Sato, Y.; Tobino, K.; Iwakami, S.I.; Goto, S.; Takahashi, K.; Maruyama, N.; Seyama, K.; et al. Complete lack of vitamin C intake generates pulmonary emphysema in senescence marker protein-30 knockout mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 298, L784–L792. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yi, J. Cancer cell killing via ROS: To increase or decrease, that is the question. Cancer Biol. Ther. 2008, 7, 1875–1884. [Google Scholar] [CrossRef] [PubMed]
- Pelicano, H.; Carney, D.; Huang, P. ROS stress in cancer cells and therapeutic implications. Drug Resist. Update 2004, 7, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [Green Version]
- Benhar, M.; Engelberg, D.; Levitzki, A. ROS, stress-activated kinases and stress signaling in cancer. EMBO Rep. 2002, 3, 420–425. [Google Scholar] [CrossRef] [Green Version]
- Lau, A.T.; Wang, Y.; Chiu, J.F. Reactive oxygen species: Current knowledge and applications in cancer research and therapeutic. J. Cell. Biochem. 2008, 104, 657–667. [Google Scholar] [CrossRef]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Mo, Z.; Zheng, S.; Lv, Z.; Zhuang, Y.; Lan, X.; Wang, F.; Lu, X.; Zhao, Y.; Zhou, S. Senescence marker protein 30 (SMP30) serves as a potential prognostic indicator in hepatocellular carcinoma. Sci. Rep. 2016, 6, 39376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sariego, J.; Zrada, S.; Byrd, M.; Matsumoto, T. Breast cancer in young patients. Am. J. Surg. 1995, 170, 243–245. [Google Scholar] [CrossRef]
- Maggard, M.A.; O’Connell, J.B.; Lane, K.E.; Liu, J.H.; Etzioni, D.A.; Ko, C.Y. Do young breast cancer patients have worse outcomes? J. Surg. Res. 2003, 113, 109–113. [Google Scholar] [CrossRef]
- Fujita, T.; Uchida, K.; Maruyama, N. Purification of senescence marker protein-30 (SMP30) and its androgen-independent decrease with age in the rat liver. Biochim. Biophys. Acta 1992, 1116, 122–128. [Google Scholar] [CrossRef]
- Sidoni, A.; Cavaliere, A.; Bellezza, G.; Scheibel, M.; Bucciarelli, E. Breast cancer in young women: Clinicopathological features and biological specificity. Breast 2003, 12, 247–250. [Google Scholar] [CrossRef]
- Bakkach, J.; Mansouri, M.; Derkaoui, T.; Loudiyi, A.; Fihri, M.; Hassani, S.; Barakat, A.; Ghailani Nourouti, N.; Bennani Mechita, M. Clinicopathologic and prognostic features of breast cancer in young women: A series from North of Morocco. BMC Womens Health 2017, 17, 106. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immunohistochemical Grading Standard for SMP30 Expression | |||
|---|---|---|---|
| SMP30 Intensity (per 200× Field) | Score | SMP30 Positive Cell Ratio (per 200× Field) | Score |
| Negative | 0 | <5% | 0 |
| Weak | 1 | 5–15% | 0.5 |
| Moderate | 2 | 16–25% | 1 |
| Strong | 3 | 26–33% | 1.5 |
| Intense | 4 | 34–50% | 2 |
| 51–66% | 2.5 | ||
| 67–75% | 3 | ||
| 76–100% | 4 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baek, S.-M.; Lee, S.-W.; Kim, T.-U.; Choi, S.-K.; Yun, S.; Lee, W.-J.; Han, S.-H.; Hong, I.-H.; Park, S.-J.; Kim, T.-H.; et al. Senescence Marker Protein 30 (SMP30): A Novel Pan-Species Diagnostic Marker for the Histopathological Diagnosis of Breast Cancer in Humans and Animals. Int. J. Mol. Sci. 2021, 22, 2340. https://doi.org/10.3390/ijms22052340
Baek S-M, Lee S-W, Kim T-U, Choi S-K, Yun S, Lee W-J, Han S-H, Hong I-H, Park S-J, Kim T-H, et al. Senescence Marker Protein 30 (SMP30): A Novel Pan-Species Diagnostic Marker for the Histopathological Diagnosis of Breast Cancer in Humans and Animals. International Journal of Molecular Sciences. 2021; 22(5):2340. https://doi.org/10.3390/ijms22052340
Chicago/Turabian StyleBaek, Su-Min, Seoung-Woo Lee, Tae-Un Kim, Seong-Kyoon Choi, Sungho Yun, Won-Jae Lee, Se-Hyeon Han, Il-Hwa Hong, Sang-Joon Park, Tae-Hwan Kim, and et al. 2021. "Senescence Marker Protein 30 (SMP30): A Novel Pan-Species Diagnostic Marker for the Histopathological Diagnosis of Breast Cancer in Humans and Animals" International Journal of Molecular Sciences 22, no. 5: 2340. https://doi.org/10.3390/ijms22052340