
Yes-Associated Protein 1 Is a Novel Calcium Sensing Receptor Target in Human Parathyroid Tumors

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
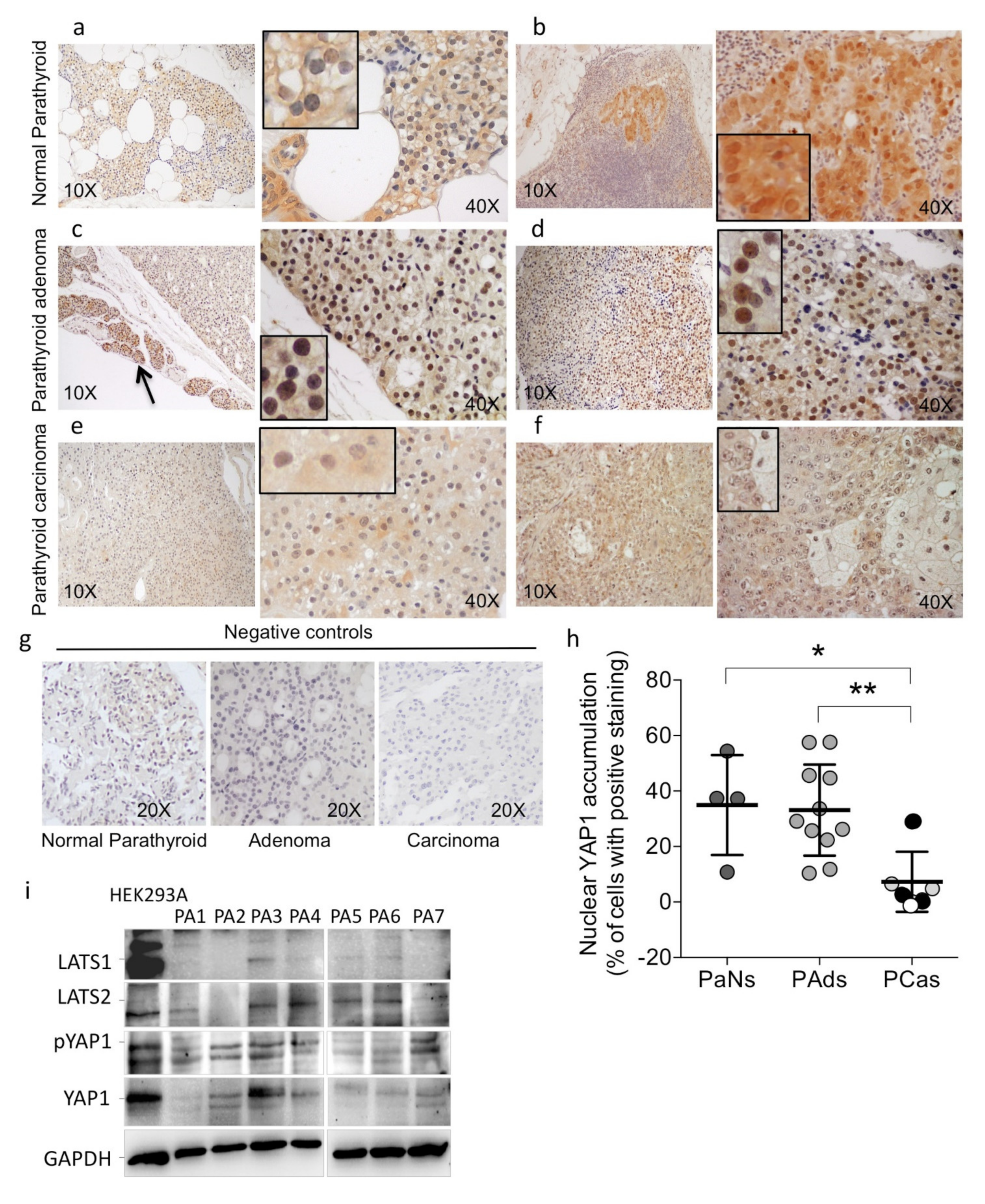
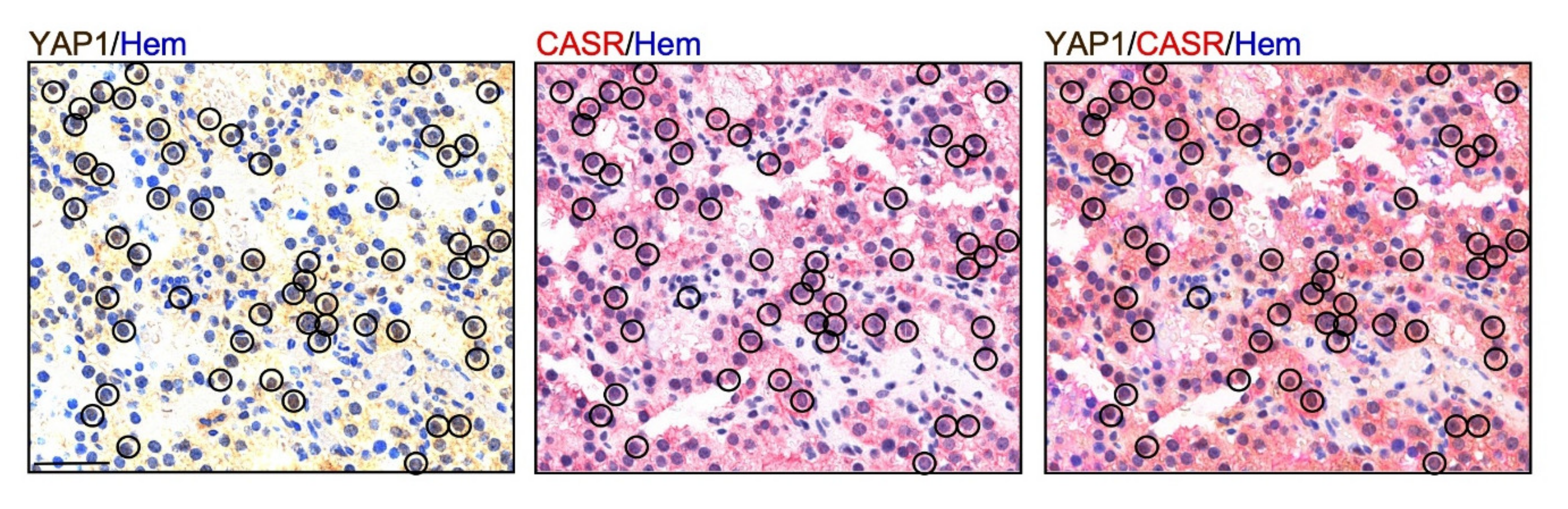
2.1. YA1P Expression in Human Parathyroid Tumors
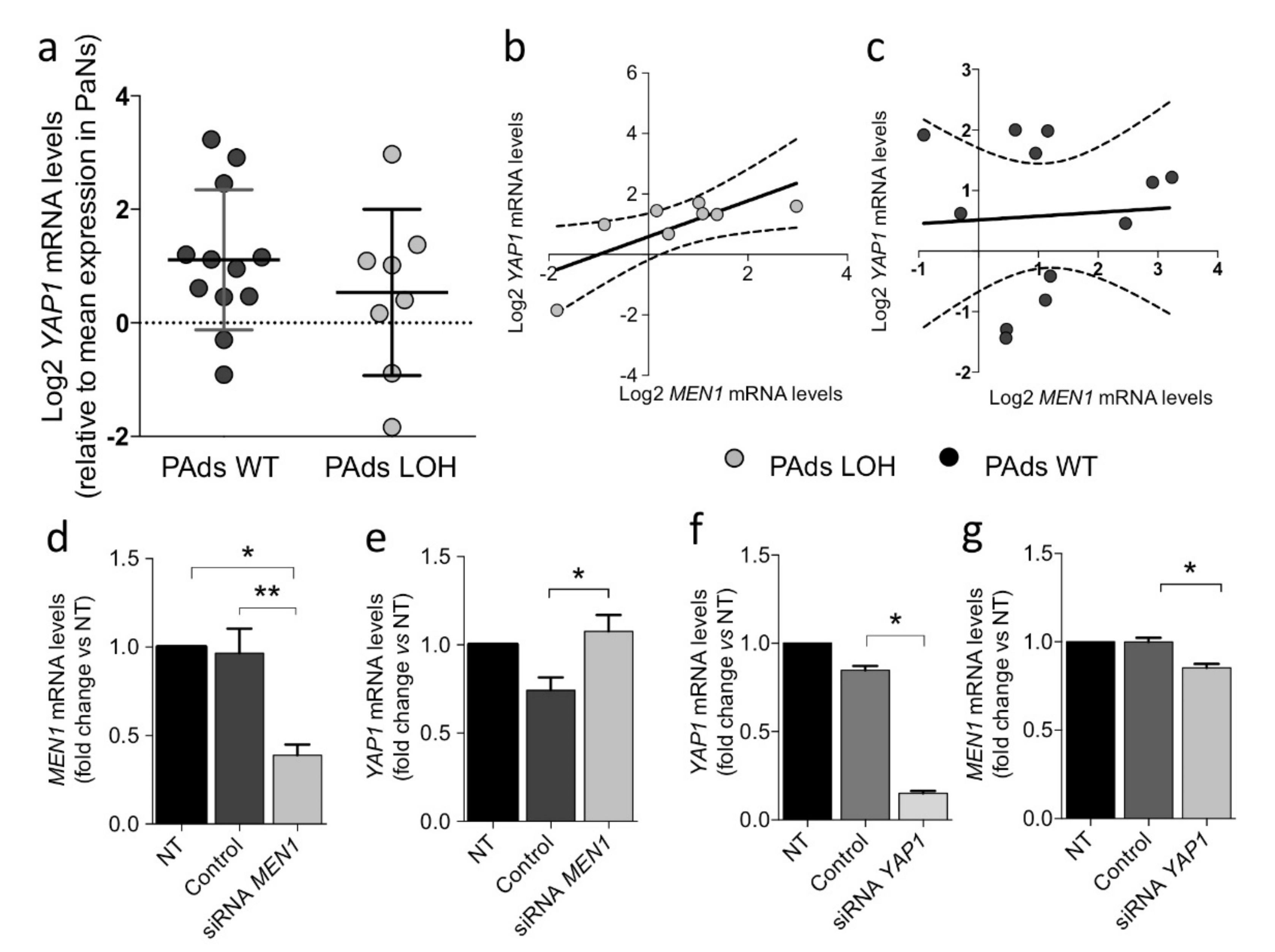
2.2. MEN1 Aberrations Do Not Directly Modulate YAP1 Expression in Parathyroid Tumors
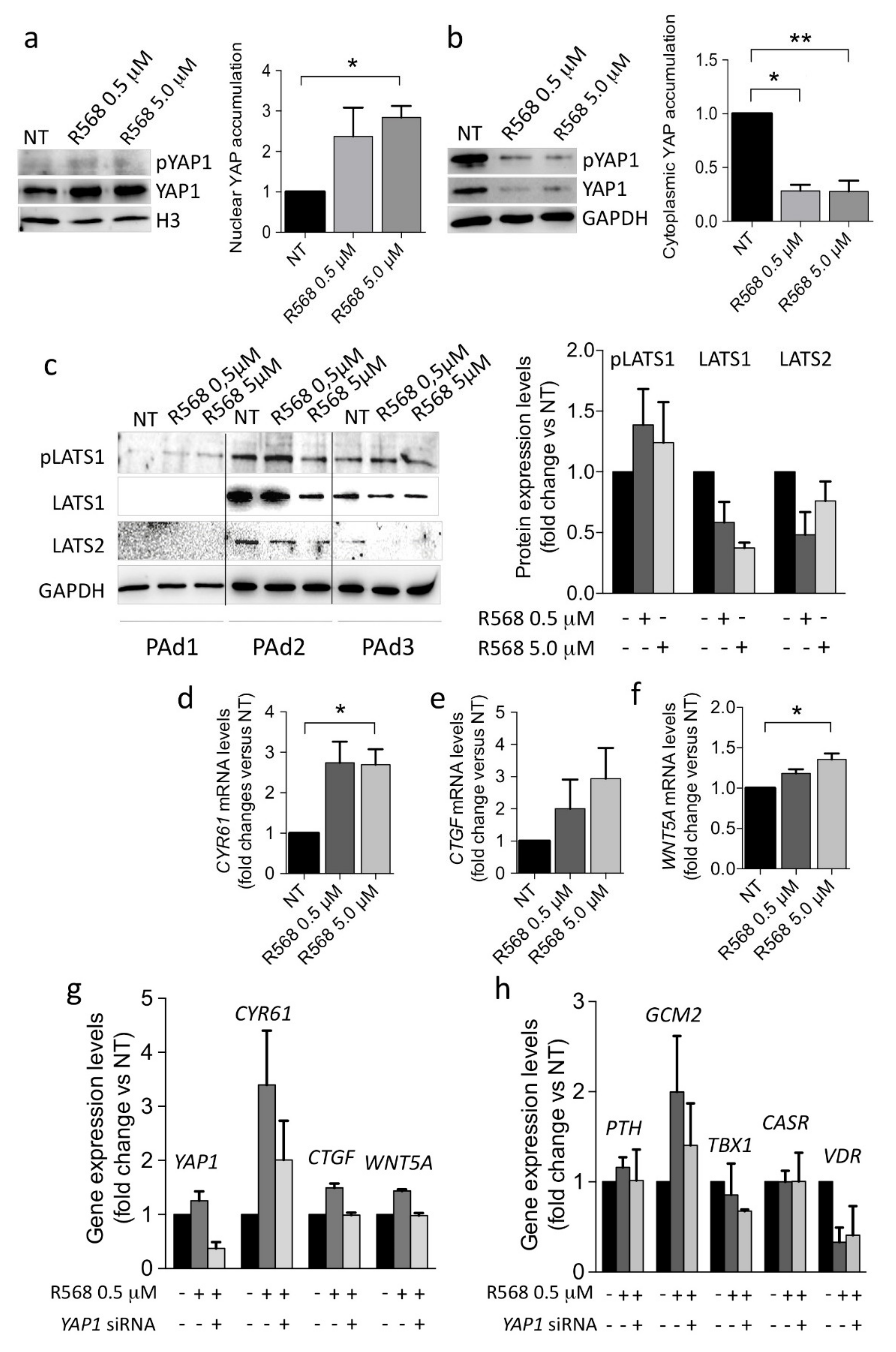
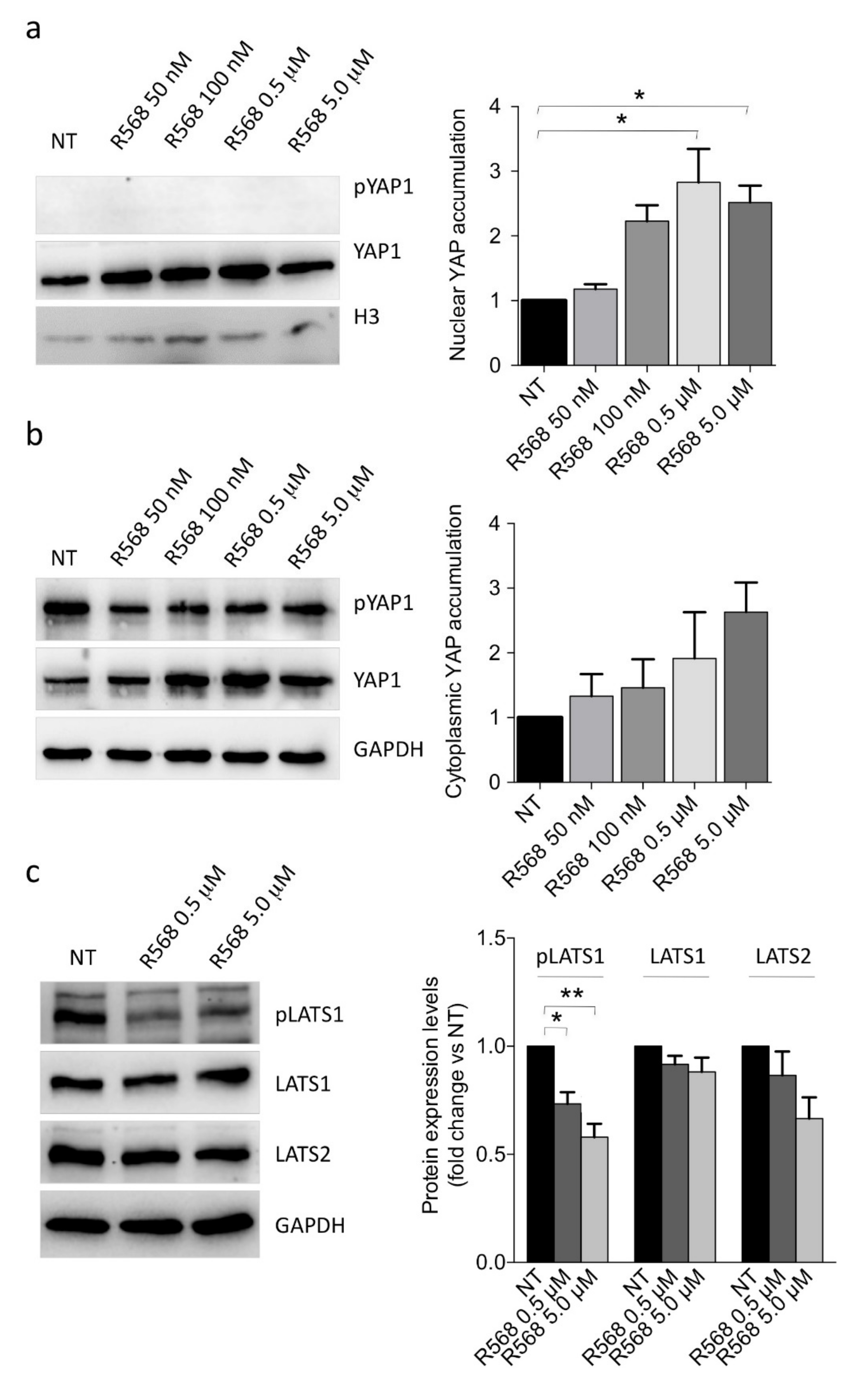
2.3. CASR Activation Induces Nuclear YAP1 Accumulation and Inhibits LATS2 Expression
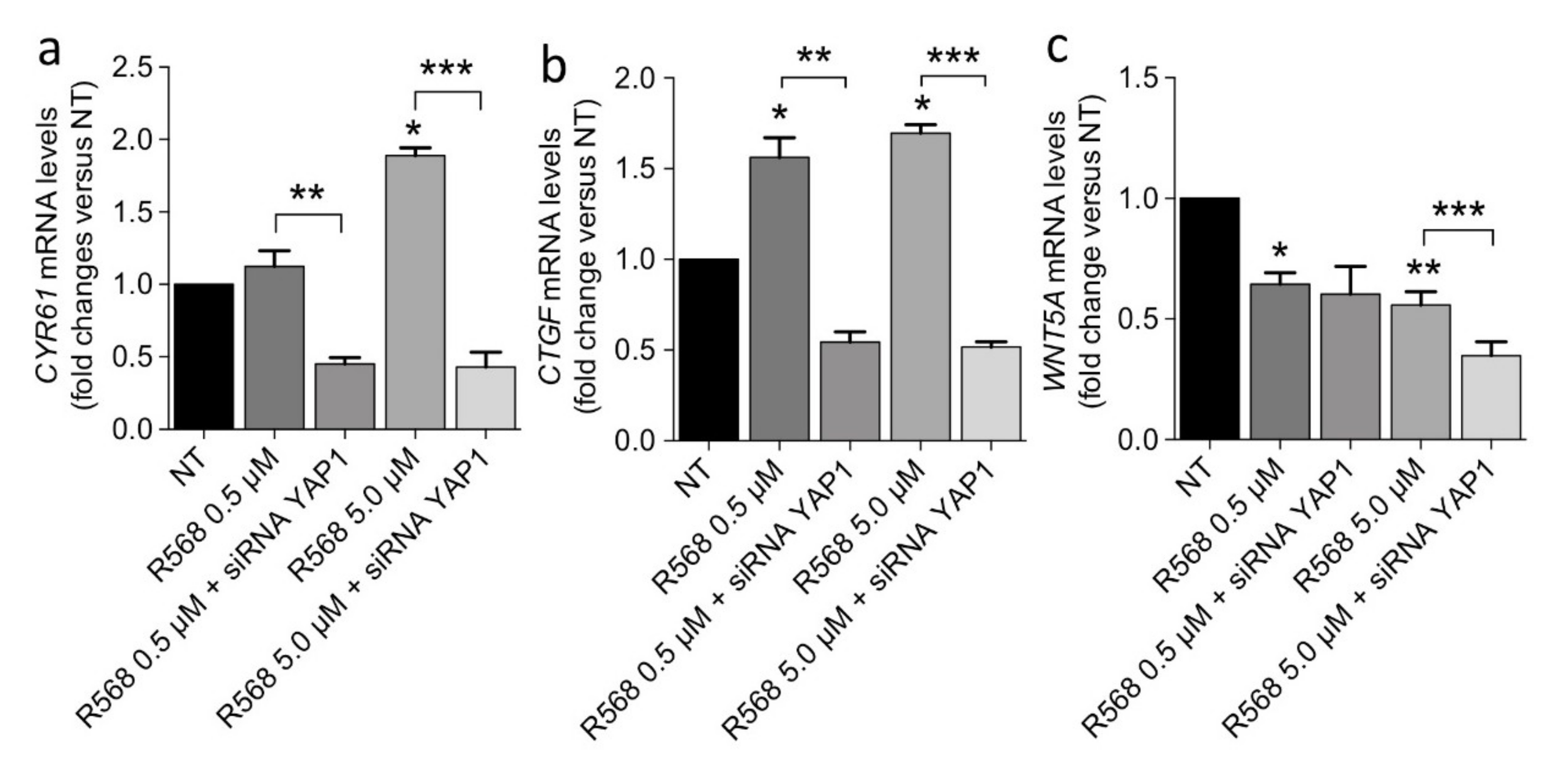
2.4. CASR Activation Modulates the Expression of YAP1 Target Genes in PAds-Derived Cells
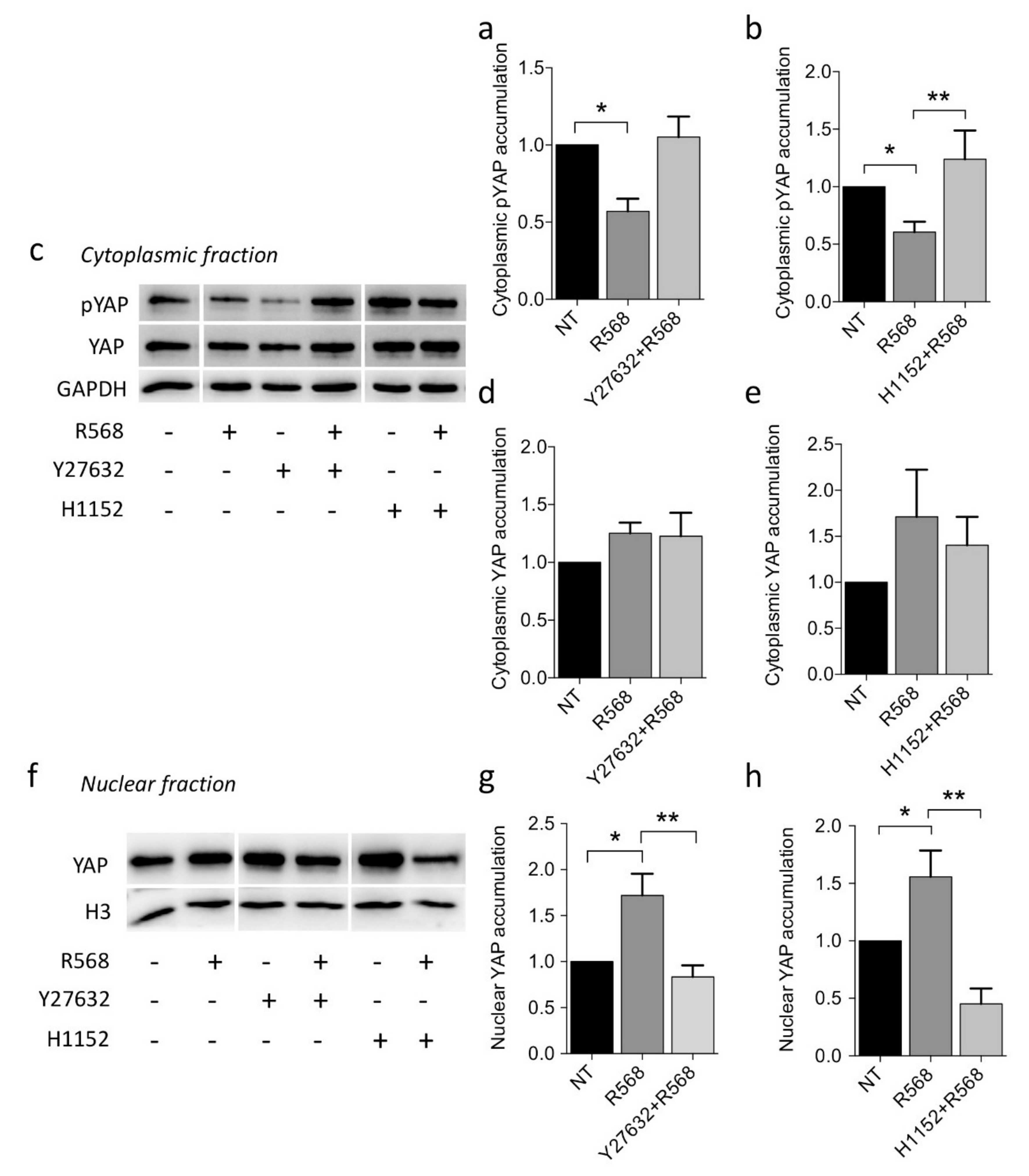
2.5. Dissecting CASR-YAP1 Signaling in CASR-HEK293A Cells
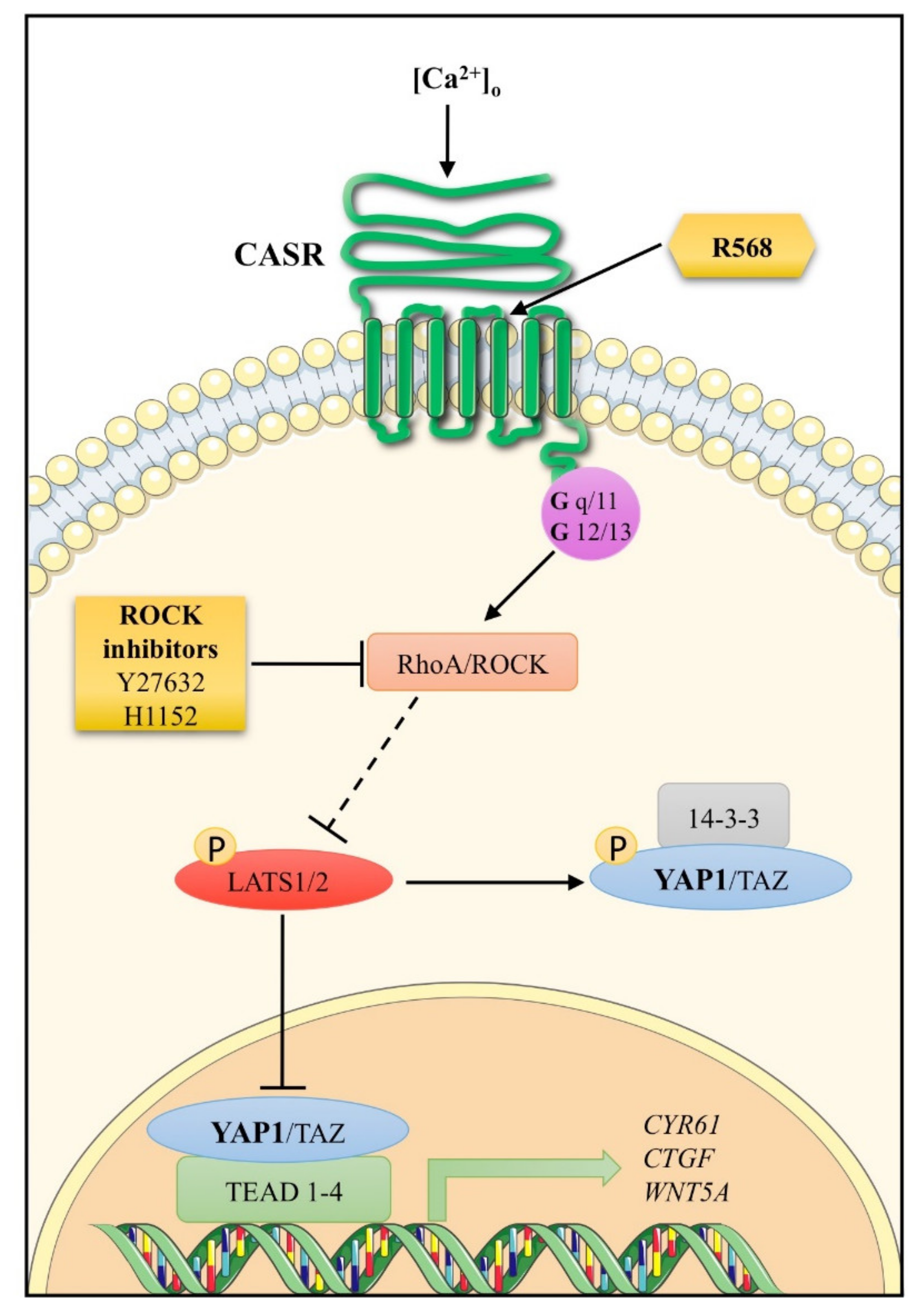
3. Discussion
4. Materials and Methods
4.1. Parathyroid Tissue Samples
4.2. Cell Cultures
4.3. DNA Extraction and Array Comparative Genomic Hybridization (aCGH) Analysis
4.4. Peripheral Blood DNA Extraction and Direct Sequencing of CDC73 and MEN1 Genes
4.5. RNA Isolation and Real-Time Quantitative Reverse Transcription (qRT-PCR)
4.6. Protein Extraction and Western Blot Analysis
4.7. Immunohistochemistry
4.8. YAP1 and MEN1 Gene Silencing
4.9. CASR Transfection
4.10. Treatments of CASR-HEK293A and PAds-Derived Cells with [Ca2+]o or R568
4.11. Treatment of CASR-HEK293A Cells with Rho-Kinase Inhibitors
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CASR | Calcium sensing receptor |
| ROCK | Rho-associated coiled-coil-containing protein kinase |
| LATS1/2 | Large tumor suppressor 1/2 |
| YAP1 | Yes-associated protein 1 |
| TAZ | Tafazzin, coactivator with PDZ-binding motif |
| HEK293A | Human embryonic kidney 293A cells |
| CDC73 | Cell division cycle 73 |
| MEN1 | Multiple endocrine neoplasia type 1 |
References
- Pan, D. The hippo signaling pathway in development and cancer. Dev. Cell 2010, 19, 491–505. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.X.; Zhao, B.; Guan, K.L. Hippo pathway in organ size control, tissue homeostasis, and cancer. Cell 2015, 163, 811–828. [Google Scholar] [CrossRef] [Green Version]
- Zanconato, F.; Cordenonsi, M.; Piccolo, S. YAP/TAZ at the roots of cancer. Cancer Cell 2016, 29, 783–803. [Google Scholar] [CrossRef] [Green Version]
- Park, H.W.; Kim, Y.C.; Yu, B.; Moroishi, T.; Mo, J.S.; Plouffe, S.W.; Meng, Z.; Lin, K.C.; Yu, F.X.; Alexander, C.M.; et al. Alternative Wnt signaling activates YAP/TAZ. Cell 2015, 162, 780–794. [Google Scholar] [CrossRef] [Green Version]
- Sebio, A.; Lenz, H.-J. Molecular pathways: Hippo signaling, a critical tumor suppressor. Clin. Cancer Res. 2015, 21, 5002–5007. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic signaling pathways in The Cancer Genome Atlas. Cell 2018, 173, 321–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, J.S.; Park, H.W.; Guan, K.L. The Hippo signaling pathway in stem cell biology and cancer. EMBO Rep. 2014, 15, 642–656. [Google Scholar] [CrossRef] [Green Version]
- Conigrave, A.D.; Ward, D.T. Calcium-sensing receptor (CaSR): Pharmacological properties and signaling pathways. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 27, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Mingione, A.; Verdelli, C.; Ferrero, S.; Vaira, V.; Guarnieri, V.; Scillitani, A.; Vicentini, L.; Balza, G.; Beretta, E.; Terranegra, A.; et al. Filamin A is reduced and contributes to the CASR sensitivity in human parathyroid tumors. J. Mol. Endocrinol. 2017, 58, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Corbetta, S.; Mantovani, G.; Lania, A.; Borgato, S.; Vicentini, L.; Beretta, E.; Faglia, G.; Di Blasio, A.M.; Spada, A. Calcium-sensing receptor expression and signalling in human parathyroid adenomas and primary hyperplasia. Clin. Endocrinol. (Oxf.) 2000, 52, 339–348. [Google Scholar] [CrossRef]
- Corbetta, S.; Lania, A.; Filopanti, M.; Vicentini, L.; Ballaré, E.; Spada, A. Mitogen-activated protein kinase cascade in human normal and tumoral parathyroid cells. J. Clin. Endocrinol. Metab. 2002, 87, 2201–2205. [Google Scholar] [CrossRef]
- Davies, S.L.; Gibbons, C.E.; Vizard, T.; Ward, D.T. Ca2+-sensing receptor induces Rho kinase-mediated actin stress fiber assembly and altered cell morphology, but not in response to aromatic amino acids. Am. J. Physiol. Cell Physiol. 2006, 290, C1543–C1551. [Google Scholar] [CrossRef] [Green Version]
- Pi, M.; Spurney, R.F.; Tu, Q.; Hinson, T.; Quarles, L.D. Calcium-sensing receptor activation of rho involves filamin and rho-guanine nucleotide exchange factor. Endocrinology 2002, 143, 3830–3838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Hujer, K.M.; Wu, Z.; Miller, R.T. The Ca2+-sensing receptor couples to Galpha12/13 to activate phospholipase D in Madin-Darby canine kidney cells. Am. J. Physiol. Cell Physiol. 2004, 286, C22–C30. [Google Scholar] [CrossRef] [PubMed]
- Cetani, F.; Pardi, E.; Vignali, E.; Borsari, S.; Picone, A.; Cianferotti, L.; Ambrogini, E.; Miccoli, P.; Pinchera, A.; Marcocci, C. MEN1 gene alterations do not correlate with the phenotype of sporadic primary hyperparathyroidism. J. Endocrinol. Investig. 2002, 25, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Dwight, T.; Twigg, S.; Delbridge, L.; Wong, F.K.; Farnebo, F.; Richardson, A.L.; Nelson, A.; Zedenius, J.; Philips, J.; Larsson, C.; et al. Loss of heterozygosity in sporadic parathyroid tumours: Involvement of chromosome 1 and the MEN1 gene locus in 11q13. Clin. Endocrinol. (Oxf.) 2000, 53, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Verdelli, C.; Forno, I.; Morotti, A.; Creo, P.; Guarnieri, V.; Scillitani, A.; Cetani, F.; Vicentini, L.; Balza, G.; Beretta, E.; et al. The aberrantly expressed miR-372 partly impairs sensitivity to apoptosis in parathyroid tumor cells. Endocr. Relat. Cancer 2018, 25, 761–771. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.; Yang, J.; DeRan, M.; Wu, C.; Su, A.I.; Bonamy, G.M.; Liu, J.; Peters, E.C.; Wu, X. Identification of serum-derived sphingosine-1-phosphate as a small molecule regulator of YAP. Chem. Biol. 2012, 19, 955–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, J.S.; Yu, F.X.; Gong, R.; Brown, J.H.; Guan, K.L. Regulation of the Hippo-YAP pathway by protease-activated receptors (PARs). Genes Dev. 2012, 26, 2138–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.-X.; Zhao, B.; Panupinyhu, N.; Jewell, J.L.; Lian, I.; Wang, L.H.; Zhao, J.; Yuan, H.; Tumareneng, K.; Li, H.; et al. Regulation of the Hippo-YAP pathway by G-protein-coupled receptor signaling. Cell 2012, 150, 780–791. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.X.; Luo, J.; Mo, J.S.; Liu, G.; Kim, Y.C.; Meng, Z.; Zhao, L.; Peyman, G.; Ouyang, H.; Jiang, W.; et al. Mutant Gq/11 promote uveal melanoma tumorigenesis by activating YAP. Cancer Cell 2014, 25, 822–830. [Google Scholar] [CrossRef] [Green Version]
- Shahbazi, R.; Baradaran, B.; Khordadmehr, M.; Safaei, S.; Baghbanzadeh, A.; Jigari, F.; Ezzati, H. Targeting ROCK signaling in health, malignant and non-malignant diseases. Immunol. Lett. 2020, 219, 15–26. [Google Scholar] [CrossRef]
- Mingione, A.; Verdelli, C.; Terranegra, A.; Soldati, L.; Corbetta, S. Molecular and clinical aspects of the target therapy with the calcimimetic cinacalcet in the treatment of parathyroid tumors. Curr. Cancer Drug Targets 2015, 15, 563–574. [Google Scholar] [CrossRef]
- Verdelli, C.; Tavanti, G.S.; Corbetta, S. Intratumor heterogeneity in human parathyroid tumors. Histol. Histopathol. 2020, 18230. [Google Scholar] [CrossRef]
- Guarnieri, V.; Battista, C.; Muscarella, L.A.; Bisceglia, M.; de Martino, D.; Baorda, F.; Maiello, E.; D’Agruma, L.; Chiodini, I.; Clemente, C.; et al. CDC73 mutations and parafibromin immunohistochemistry in parathyroid tumors: Clinical correlations in a single-centre patient cohort. Cell Oncol. (Dordr.) 2012, 6, 411–422. [Google Scholar] [CrossRef]
- Muscarella, L.A.; Turchetti, D.; Fontana, A.; Baorda, F.; Palumbo, O.; la Torre, A.; de Martino, D.; Franco, R.; Losito, N.S.; Repaci, A.; et al. Large deletion at the CDC73 gene locus and search for predictive markers of the presence of a CDC73 genetic lesion. Oncotarget 2018, 9, 20721–20733. [Google Scholar] [CrossRef] [Green Version]
- Juhlin, C.C.; Nilsson, I.L.; Lagerstedt-Robinson, K.; Stenman, A.; Bränström, R.; Tham, E.; Höög, A. Parafibromin immunostainings of parathyroid tumors in clinical routine: A near-decade experience from a tertiary center. Mod. Pathol. 2019, 32, 1082–1094. [Google Scholar] [CrossRef]
- Jho, E. Dual role of YAP: Oncoprotein and tumor suppressor. J. Thorac. Dis. 2018, 10, S3895–S3898. [Google Scholar] [CrossRef]
- Wang, H.; Du, Y.C.; Zhou, X.J.; Liu, H.; Tang, S.C. The dual functions of YAP-1 to promote and inhibit cell growth in human malignancy. Cancer Metastasis. Rev. 2014, 33, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Totty, N.F.; Irwin, M.S.; Sudol, M.; Downward, J. Akt phosphorylates the Yes-associated protein, YAP, to induce interaction with 14-3-3 and attenuation of p73-mediated apoptosis. Mol. Cell 2003, 11, 11–23. [Google Scholar] [CrossRef]
- Strano, S.; Monti, O.; Pediconi, N.; Baccarini, A.; Fontemaggi, G.; Lapi, E.; Mantovani, F.; Damalas, A.; Citro, G.; Sacchi, A.; et al. The transcriptional coactivator Yes-associated protein drives p73 gene-target specificity in response to DNA Damage. Mol. Cell 2005, 18, 447–459. [Google Scholar] [CrossRef]
- Cottini, F.; Hideshima, T.; Xu, C.; Sattler, M.; Dori, M.; Agnelli, L.; ten Hacken, E.; Bertilacciom, M.T.; Antonini, E.; Neri, A.; et al. Rescue of Hippo coactivator YAP1 triggers DNA damage-induced apoptosis in hematological cancers. Nat. Med. 2014, 20, 599–606. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Takahashi-Kanemitsu, A.; Kikuchi, I.; Ben, C.; Hatakeyama, M. Transcriptional co-activator functions of YAP and TAZ are inversely regulated by tyrosine phosphorylation status of parafibromin. iScience 2018, 2, 103. [Google Scholar] [CrossRef] [Green Version]
- Falchetti, A. Genetics of parathyroids disorders: Overview. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 781–790. [Google Scholar] [CrossRef]
- Aycicek, G.S.; Aydogan, B.I.; Sahin, M.; Ersoz, C.C.; Sak, S.D.; Baskal, N. Clinical impact of p27kip1 and CaSR expression on primary hyperparathyroidism. Endocr. Pathol. 2018, 29, 250–258. [Google Scholar] [CrossRef]
- Singh, P.; Bhadada, S.K.; Dahiya, D.; Arya, A.K.; Saikia, U.N.; Sachdeva, N.; Kaur, J.; Brandi, M.L.; Rao, S.D. Reduced calcium sensing receptor (CaSR) expression is epigenetically deregulated in parathyroid adenomas. J. Clin. Endocrinol. Metab. 2020, 105, 3015–3024. [Google Scholar] [CrossRef]
- Xu, B.; Li, S.H.; Zheng, R.; Gao, S.B.; Ding, L.H.; Yin, Z.Y.; Lin, X.; Feng, Z.J.; Zhang, S.; Wang, X.M.; et al. Menin promotes hepatocellular carcinogenesis and epigenetically up-regulates Yap1 transcription. Proc. Natl. Acad. Sci. USA 2013, 110, 17480–17485. [Google Scholar] [CrossRef] [Green Version]
- Verdelli, C.; Forno, I.; Vaira, V.; Corbetta, S. Epigenetic alterations in human parathyroid tumors. Endocrine 2015, 49, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Lu, J.; Goyal, A.; Wong, T.; Lian, G.; Zhang, J.; Hecht, J.L.; Feng, Y.; Sheen, V.L. Opposing FlnA and FlnB interactions regulate RhoA activation in guiding dynamic actin stress fiber formation and cell spreading. Hum. Mol. Genet. 2017, 26, 1294–1304. [Google Scholar] [CrossRef]
- Park, H.W.; Guan, K.L. Regulation of the Hippo pathway and implications for anticancer drug development. Trends Pharmacol. Sci. 2013, 34, 581–589. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Son, S.; Shin, I. Role of the CCN protein family in cancer. BMB Rep. 2018, 51, 486–492. [Google Scholar] [CrossRef] [Green Version]
- Totaro, A.; Panciera, T.; Piccolo, S. YAP/TAZ upstream signals and downstream responses. Nat. Cell Biol. 2018, 20, 888–899. [Google Scholar] [CrossRef]
- Verdelli, C.; Avagliano, L.; Guarnieri, V.; Cetani, F.; Ferrero, S.; Vicentini, L.; Beretta, E.; Scillitani, A.; Creo, P.; Bulfamante, G.P.; et al. Expression, function, and regulation of the embryonic transcription factor TBX1 in parathyroid tumors. Lab. Investig. 2017, 97, 1488–1499. [Google Scholar] [CrossRef]
- Mizobuchi, M.; Ritter, C.S.; Krits, I.; Slatopolsky, E.; Sicard, G.; Brown, A.J. Calcium-sensing receptor expression is regulated by glial cells missing-2 in human parathyroid cells. J. Bone Miner. Res. 2009, 24, 1173–1179. [Google Scholar] [CrossRef] [Green Version]
- Correa, P.; Akerström, G.; Westin, G. Underexpression of Gcm2, a master regulatory gene of parathyroid gland development, in adenomas of primary hyperparathyroidism. Clin. Endocrinol. (Oxf.) 2002, 57, 501–505. [Google Scholar] [CrossRef]
- Morotti, A.; Forno, I.; Verdelli, C.; Guarnieri, V.; Cetani, F.; Terrasi, A.; Silipigni, R.; Guerneri, S.; Andrè, V.; Scillitani, A.; et al. The oncosuppressors MEN1 and CDC73 are involved in lncRNA deregulation in human parathyroid tumors. J. Bone Miner. Res. 2020. [Google Scholar] [CrossRef]
- Cinque, L.; Sparaneo, A.; Salcuni, A.S.; de Martino, D.; Battista, C.; Logoluso, F.; Palumbo, O.; Cocchi, R.; Maiello, E.; Graziano, P.; et al. MEN1 gene mutation with parathyroid carcinoma: First report of a familial case. Endocr. Connect. 2017, 8, 886–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarnieri, V.; Scillitani, A.; Muscarella, L.A.; Battista, C.; Bonfitto, N.; Bisceglia, M.; Minisola, S.; Mascia, M.L.; D’Agruma, L.; Cole, D.E.C. Diagnosis of parathyroid tumors in familial isolated hyperparathyroidism with HRPT2 mutation: Implications for cancer surveillance. J. Clin. Endocrinol. Metab. 2006, 8, 2827–2832. [Google Scholar] [CrossRef]
- Cinque, L.; Sparaneo, A.; Cetani, F.; Coco, M.; Clemente, C.; Chetta, M.; Balsamo, T.; Battista, C.; Sanpaolo, E.; Pardi, E.; et al. Novel association of MEN1 gene mutations with parathyroid carcinoma. Oncol. Lett. 2017, 1, 23–30. [Google Scholar] [CrossRef]
- DeLellis, R.; Larsson, C.; Arnold, A.; Lloy, R.; Bilezikian, J.; Mete, O.; Eng, C. Tumors of the parathyroid glands. In WHO Classification of Tumors of Endocrine Organs, 4th ed.; Lloyd, R., Osamura, R., Kloppel, G., Rosai, J., Eds.; IARC Press: Lyon, France, 2017; pp. 145–159. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCa Sample | Germline/ Somatic | CDC73 Mutation | MEN1 Mutation | SCa mg/dL | PTH pg/mL | Recur. | Metas. |
|---|---|---|---|---|---|---|---|
| PCa1 | Germline | c.518_521del4 | WT | 13.0 | 660 | yes | no |
| PCa2 | Germline | p.R415X | WT | 12.7 | 367 | no | yes |
| PCa3 | Germline/Somatic | WT | WT | 20.2 | 940 | no | no |
| PCa4 | Germline | WT | p.D418N | Na | na | no | no |
| PCa5 | Germline | c.685_688 delAGAG | WT | 10.0 | 101 | yes | no |
| PCa6 | Germline | WT | p.Q166L/A167S | 11.8 | 160 | yes | no |
| Gene Name | Probe Number |
|---|---|
| YAP1 | Hs00371735_m1 |
| GCM2 | Hs00899403_m1 |
| PTH | Hs00757710_g1 |
| TBX1 | Hs00271949_m1 |
| YAP1 | Hs00371735_m1 |
| CYR61 | Hs00155479_m1 |
| CTGF | Hs00170014_m1 |
| CASR | Hs01047795_m1 |
| WNT5A | Hs00998537_m1 |
| VDR | Hs01045843_m1 |
| MEN1 | Hs00365720_m1 |
| HMBS | Hs00609297_m1 |
| B2M | Hs99999907_m1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tavanti, G.S.; Verdelli, C.; Morotti, A.; Maroni, P.; Guarnieri, V.; Scillitani, A.; Silipigni, R.; Guerneri, S.; Maggiore, R.; Mari, G.; et al. Yes-Associated Protein 1 Is a Novel Calcium Sensing Receptor Target in Human Parathyroid Tumors. Int. J. Mol. Sci. 2021, 22, 2016. https://doi.org/10.3390/ijms22042016
Tavanti GS, Verdelli C, Morotti A, Maroni P, Guarnieri V, Scillitani A, Silipigni R, Guerneri S, Maggiore R, Mari G, et al. Yes-Associated Protein 1 Is a Novel Calcium Sensing Receptor Target in Human Parathyroid Tumors. International Journal of Molecular Sciences. 2021; 22(4):2016. https://doi.org/10.3390/ijms22042016
Chicago/Turabian StyleTavanti, Giulia Stefania, Chiara Verdelli, Annamaria Morotti, Paola Maroni, Vito Guarnieri, Alfredo Scillitani, Rosamaria Silipigni, Silvana Guerneri, Riccardo Maggiore, Gilberto Mari, and et al. 2021. "Yes-Associated Protein 1 Is a Novel Calcium Sensing Receptor Target in Human Parathyroid Tumors" International Journal of Molecular Sciences 22, no. 4: 2016. https://doi.org/10.3390/ijms22042016