
Feasibility of Human Platelet Lysate as an Alternative to Foetal Bovine Serum for In Vitro Expansion of Chondrocytes



Abstract
:1. Introduction
2. Cartilage Anatomy and Physiology
3. Osteoarthritis
4. Autologous Chondrocyte Implantation
5. Conventional Expansion of Chondrocytes Using Foetal Bovine Serum
6. Human Platelet Lysate as Replacement for Foetal Bovine Serum
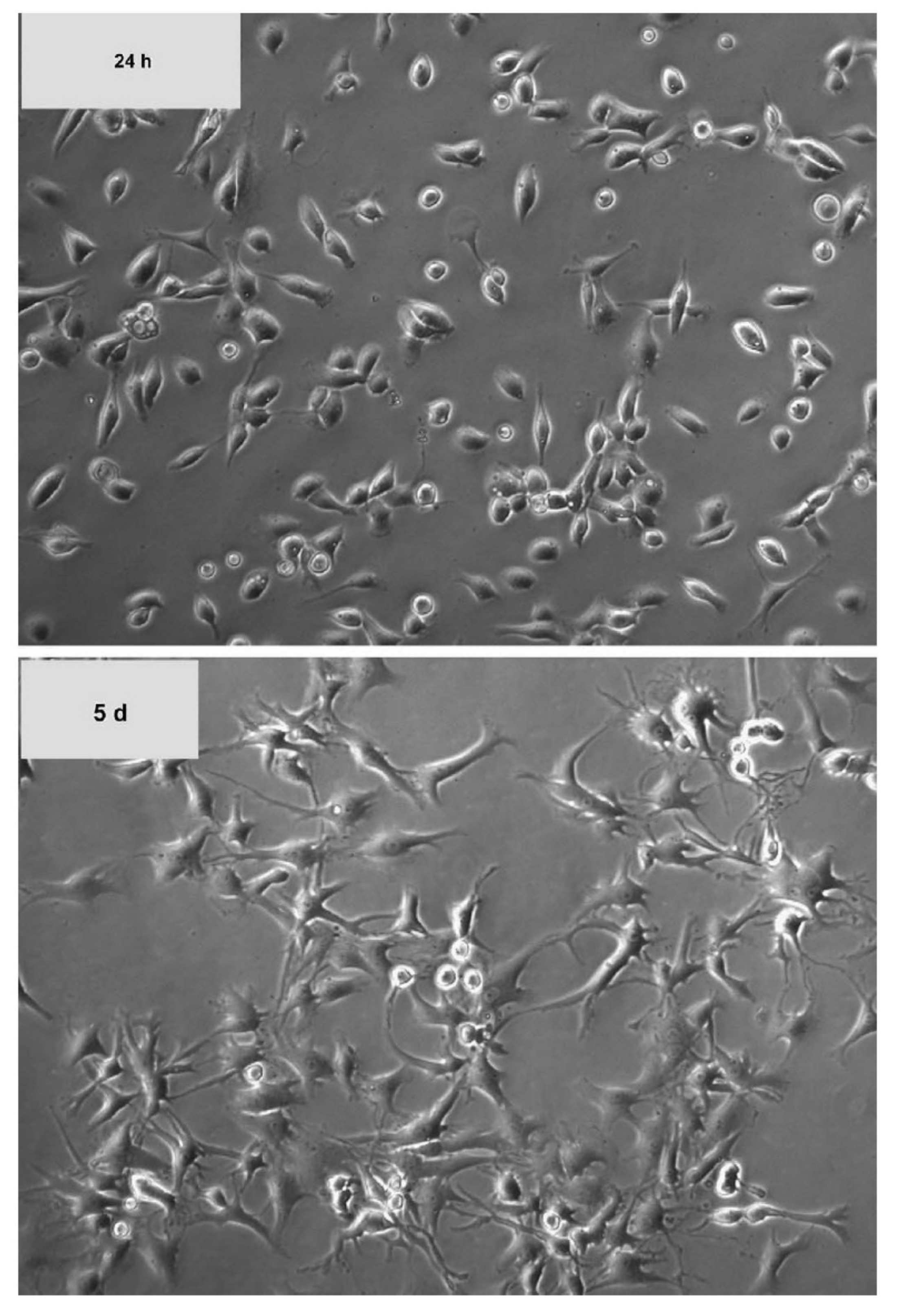
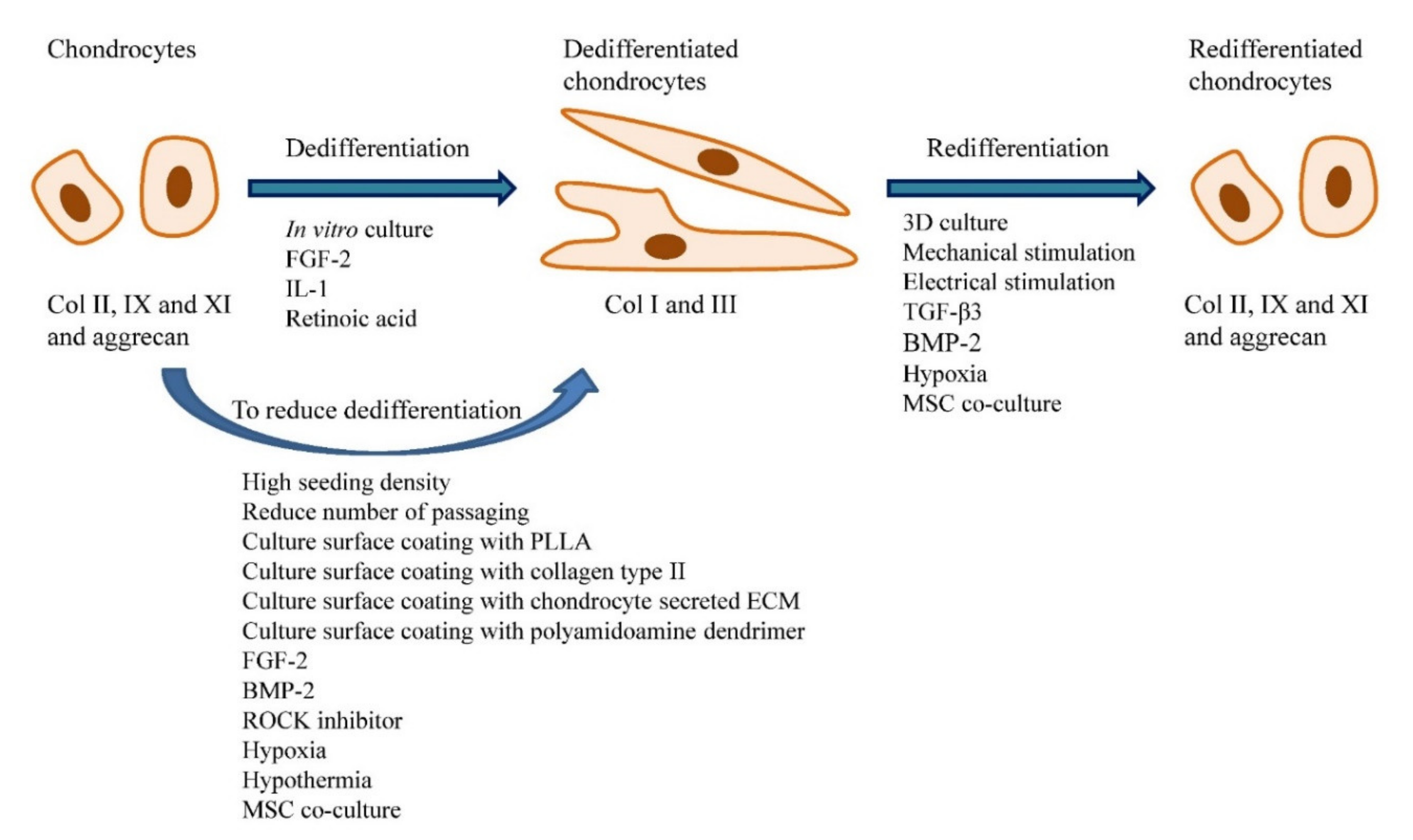
7. Chondrocyte Dedifferentiation and Redifferentiation of Dedifferentiated Cells
8. Other Alternatives for Foetal Bovine Serum
8.1. Human Serum
8.2. Serum-Free Medium
8.3. Others
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Jordan, J.M. Epidemiology of Osteoarthritis. Clin. Geriatr. Med. 2010, 26, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, E. Pharmacologic and Non-Pharmacologic Treatment of Osteoarthritis. Curr. Treat. Opt. Rheumatol. 2016, 2, 111–125. [Google Scholar] [CrossRef] [Green Version]
- Ogura, T.; Merkely, G.; Bryant, T.; Winalski, C.S.; Minas, T. Autologous Chondrocyte Implantation “Segmental-Sandwich” Technique for Deep Osteochondral Defects in the Knee: Clinical Outcomes and Correlation with Magnetic Resonance Imaging Findings. Orthop. J. Sports Med. 2019, 7, 2325967119847173. [Google Scholar] [CrossRef]
- Haleem, A.M.; Chu, C.R. Advances in Tissue Engineering Techniques for Articular Cartilage Repair. Oper. Tech. Orthop. 2010, 20, 76–89. [Google Scholar] [CrossRef] [Green Version]
- Batty, L.; Dance, S.; Bajaj, S.; Cole, B.J. Autologous Chondrocyte Implantation: An Overview of Technique and Outcomes. ANZ J. Surg. 2011, 81, 18–25. [Google Scholar] [CrossRef]
- Davies, R.L.; Kuiper, N.J. Regenerative Medicine: A Review of the Evolution of Autologous Chondrocyte Implantation (ACI) Therapy. Bioengineering 2019, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Bieback, K. Platelet Lysate as Replacement for Fetal Bovine Serum in Mesenchymal Stromal Cell Cultures. Transfus. Med. Hemother. 2013, 40, 326–335. [Google Scholar] [CrossRef] [Green Version]
- Sykes, J.G.; Kuiper, J.; Richardson, J.B.; Wright, K.T.; Kuiper, N.J. Impact of Human Platelet Lysate on the Expansion and Chondrogenic Capacity of Cultured Human Chondrocytes for Cartilage Cell Therapy. Osteoarthr. Cartil. 2018, 26, S103. [Google Scholar] [CrossRef] [Green Version]
- Sophia Fox, A.J.; Bedi, A.; Rodeo, S.A. The Basic Science of Articular Cartilage: Structure, Composition, and Function. Sports Health 2009, 1, 461–468. [Google Scholar] [CrossRef]
- Karuppal, R. Current Concepts in the Articular Cartilage Repair and Regeneration. J. Orthop. 2017, 14, A1–A3. [Google Scholar] [CrossRef]
- Luo, Y.; Sinkeviciute, D.; He, Y.; Karsdal, M.; Henrotin, Y.; Mobasheri, A.; Önnerfjord, P.; Bay-Jensen, A. The Minor Collagens in Articular Cartilage. Protein Cell 2017, 8, 560–572. [Google Scholar] [CrossRef] [Green Version]
- Camarero-Espinosa, S.; Rothen-Rutishauser, B.; Foster, E.J.; Weder, C. Articular Cartilage: From Formation to Tissue Engineering. Biomater. Sci. 2016, 4, 734–767. [Google Scholar] [CrossRef]
- Shen, S.; Chen, M.; Guo, W.; Li, H.; Li, X.; Huang, S.; Luo, X.; Wang, Z.; Wen, Y.; Yuan, Z.; et al. Three Dimensional Printing-Based Strategies for Functional Cartilage Regeneration. Tissue Eng. Part B Rev. 2019, 25, 187–201. [Google Scholar] [CrossRef] [Green Version]
- Van Tunen, J.A.C.; Peat, G.; Bricca, A.; Larsen, L.B.; Søndergaard, J.; Thilsing, T.; Roos, E.M.; Thorlund, J.B. Association of Osteoarthritis Risk Factors with Knee and Hip Pain in a Population-Based Sample of 29–59 Year Olds in Denmark: A Cross-Sectional Analysis. BMC Musculoskelet. Disord. 2018, 19, 300. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, E.; Goncalves, G.H.; Bricca, A.; Roos, E.M.; Thorlund, J.B.; Juhl, C.B. Knee Osteoarthritis Risk Is Increased 4-6 Fold after Knee Injury—A Systematic Review and Meta-Analysis. Br. J. Sports Med. 2019, 53, 1454–1463. [Google Scholar] [CrossRef]
- Yucesoy, B.; Charles, L.E.; Baker, B.; Burchfiel, C.M. Occupational and Genetic Risk Factors for Osteoarthritis: A Review. Work 2015, 50, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Buckwalter, J.A.; Mankin, H.J. Articular Cartilage: Degeneration and Osteoarthritis, Repair, Regeneration, and Transplantation. Instr. Course Lect. 1998, 47, 487–504. [Google Scholar]
- Lespasio, M.J.; Piuzzi, N.S.; Husni, M.E.; Muschler, G.F.; Guarino, A.; Mont, M.A. Knee Osteoarthritis: A Primer. Perm. J. 2017, 21, 16–183. [Google Scholar] [CrossRef] [Green Version]
- Sandell, L.J.; Aigner, T. Articular Cartilage and Changes in Arthritis: Cell Biology of Osteoarthritis. Arthritis Res. Ther. 2001, 3, 107. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Tian, B.O.; Qu, X.; Liu, F.; Tang, T.; Qin, A.N.; Zhu, Z.; Dai, K. MicroRNAs Play a Role in Chondrogenesis and Osteoarthritis. Int. J. Mol. Med. 2014, 34, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Hu, X.; Cheng, J.; Zhang, X.; Zhao, F.; Shi, W.; Ren, B.; Yu, H.; Yang, P.; Li, Z.; et al. A Small Molecule Promotes Cartilage Extracellular Matrix Generation and Inhibits Osteoarthritis Development. Nat. Commun. 2019, 10, 1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemeyer, P.; Albrecht, D.; Andereya, S.; Angele, P.; Ateschrang, A.; Aurich, M.; Baumann, M.; Bosch, U.; Erggelet, C.; Fickert, S.; et al. Autologous Chondrocyte Implantation (ACI) for Cartilage Defects of the Knee: A Guideline by the Working Group “Clinical Tissue Regeneration” of the German Society of Orthopaedics and Trauma (DGOU). Knee 2016, 23, 426–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoburg, A.; Löer, I.; Körsmeier, K.; Siebold, R.; Niemeyer, P.; Fickert, S.; Ruhnau, K. Matrix-Associated Autologous Chondrocyte Implantation Is an Effective Treatment at Midterm Follow-up in Adolescents and Young Adults. Orthop. J. Sports Med. 2019, 7, 2325967119841077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jochems, C.E.A.; van der Valk, J.B.F.; Stafleu, F.R.; Baumans, V. The Use of Fetal Bovine Serum: Ethical or Scientific Problem? Altern. Lab. Anim. 2002, 30, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Gstraunthaler, G. Alternatives to the Use of Fetal Bovine Serum: Serum-Free Cell Culture. ALTEX Altern. Anim. Exp. 2003, 20, 275–281. [Google Scholar]
- Otero, M.; Favero, M.; Dragomir, C.; Hachem, K.E.; Hashimoto, K.; Plumb, D.A.; Goldring, M.B. Human chondrocyte cultures as models of cartilage-specific gene regulation. In Human Cell Culture Protocols; Mitry, R.R., Hughes, R.D., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 301–336. ISBN 978-1-61779-367-7. [Google Scholar]
- Lee, J.; Lee, J.-Y.; Chae, B.-C.; Jang, J.; Lee, E.; Son, Y. Fully Dedifferentiated Chondrocytes Expanded in Specific Mesenchymal Stem Cell Growth Medium with FGF2 Obtains Mesenchymal Stem Cell Phenotype in Vitro but Retains Chondrocyte Phenotype in Vivo. Cell Transplant. 2017, 26, 1673–1687. [Google Scholar] [CrossRef] [Green Version]
- Duan, L.; Ma, B.; Liang, Y.; Chen, J.; Zhu, W.; Li, M.; Wang, D. Cytokine Networking of Chondrocyte Dedifferentiation in Vitro and Its Implications for Cell-Based Cartilage Therapy. Am. J. Transl. Res. 2015, 7, 194–208. [Google Scholar]
- Ma, B.; Leijten, J.C.H.; Wu, L.; Kip, M.; van Blitterswijk, C.A.; Post, J.N.; Karperien, M. Gene Expression Profiling of Dedifferentiated Human Articular Chondrocytes in Monolayer Culture. Osteoarthr. Cartil. 2013, 21, 599–603. [Google Scholar] [CrossRef] [Green Version]
- Angelozzi, M.; Penolazzi, L.; Mazzitelli, S.; Lambertini, E.; Lolli, A.; Piva, R.; Nastruzzi, C. Dedifferentiated Chondrocytes in Composite Microfibers as Tool for Cartilage Repair. Front. Bioeng. Biotechnol. 2017, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Schulze-Tanzil, G. Activation and dedifferentiation of chondrocytes: Implications in cartilage injury and repair. Ann. Anat. Anat. Anz. 2009, 191, 325–338. [Google Scholar] [CrossRef]
- Henschler, R.; Gabriel, C.; Schallmoser, K.; Burnouf, T.; Koh, M.B.C. Human Platelet Lysate Current Standards and Future Developments. Transfusion 2019, 59, 1407–1413. [Google Scholar] [CrossRef] [PubMed]
- Schallmoser, K.; Strunk, D. Preparation of Pooled Human Platelet Lysate (PHPL) as an Efficient Supplement for Animal Serum-Free Human Stem Cell Cultures. J. Vis. Exp. 2009, e1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astori, G.; Amati, E.; Bambi, F.; Bernardi, M.; Chieregato, K.; Schäfer, R.; Sella, S.; Rodeghiero, F. Platelet Lysate as a Substitute for Animal Serum for the Ex-Vivo Expansion of Mesenchymal Stem/Stromal Cells: Present and Future. Stem Cell Res. Ther. 2016, 7, 93. [Google Scholar] [CrossRef] [Green Version]
- Prana, H.B.B.H.; Hanif, B.M.M.; Hanim, B.O.A.; Hanani, B.N.N.; Saihah, B.M.A.N.; Fathi, B.H.M.N.; Hamdan, M.Y.N.; Rizal, A.R.; Leong, C.F.; Ng, M.H.; et al. Human Platelet Lysate Promotes Proliferation but Fails to Maintain Chondrogenic Markers of Chondrocytes. Sains Malays. 2019, 48, 2169–2176. [Google Scholar]
- Dessels, C.; Durandt, C.; Pepper, M.S. Comparison of Human Platelet Lysate Alternatives Using Expired and Freshly Isolated Platelet Concentrates for Adipose-Derived Stromal Cell Expansion. Platelets 2019, 30, 356–367. [Google Scholar] [CrossRef] [Green Version]
- Antoninus, A.A.; Widowati, W.; Wijaya, L.; Agustina, D.; Puradisastra, S.; Sumitro, S.B.; Widodo, M.A.; Bachtiar, I. Human Platelet Lysate Enhances the Proliferation of Wharton’s Jelly-Derived Mesenchymal Stem Cells. Biomark. Genom. Med. 2015, 7, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Klatte-Schulz, F.; Schmidt, T.; Uckert, M.; Scheffler, S.; Kalus, U.; Rojewski, M.; Schrezenmeier, H.; Pruss, A.; Wildemann, B. Comparative Analysis of Different Platelet Lysates and Platelet Rich Preparations to Stimulate Tendon Cell Biology: An in Vitro Study. Int. J. Mol. Sci. 2018, 19, 212. [Google Scholar] [CrossRef] [Green Version]
- Pereira, R.C.; Scaranari, M.; Benelli, R.; Strada, P.; Reis, R.L.; Cancedda, R.; Gentili, C. Dual Effect of Platelet Lysate on Human Articular Cartilage: A Maintenance of Chondrogenic Potential and a Transient Proinflammatory Activity Followed by an Inflammation Resolution. Tissue Eng. Part A 2013, 19, 1476–1488. [Google Scholar] [CrossRef]
- Hildner, F.; Eder, M.J.; Hofer, K.; Aberl, J.; Redl, H.; van Griensven, M.; Gabriel, C.; Peterbaue-Scherb, A. Human Platelet Lysate Successfully Promotes Proliferation and Subsequent Chondrogenic Differentiation of Adipose-Derived Stem Cells: A Comparison with Articular Chondrocytes. J. Tissue Eng. Regen. Med. 2015, 9, 808–818. [Google Scholar] [CrossRef]
- Kaps, C.; Loch, A.; Haisch, A.; Smolian, H.; Burmester, G.R.; Häupl, T.; Sittinger, M. Human Platelet Supernatant Promotes Proliferation but Not Differentiation of Articular Chondrocytes. Med. Biol. Eng. Comput. 2002, 40, 485–490. [Google Scholar] [CrossRef]
- Muraglia, A.; Nguyen, V.T.; Nardini, M.; Mogni, M.; Coviello, D.; Dozin, B.; Strada, P.; Baldelli, I.; Formica, M.; Cancedda, R.; et al. Culture Medium Supplements Derived from Human Platelet and Plasma: Cell Commitment and Proliferation Support. Front. Bioeng. Biotechnol. 2017, 5, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kachroo, U.; Zachariah, S.M.; Thambaiah, A.; Tabasum, A.; Livingston, A.; Rebekah, G.; Srivastava, A.; Vinod, E. Comparison of Human Platelet Lysate versus Fetal Bovine Serum for Expansion of Human Articular Cartilage-Derived Chondroprogenitors. Cartilage 2020. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.; González-García, C.; Gómez Ribelles, J.L.; Salmerón-Sánchez, M. Maintenance of Chondrocyte Phenotype during Expansion on PLLA Microtopographies. J. Tissue Eng. 2018, 9, 2041731418789829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, K.H.; Aminuddin, B.S.; Fuzina, N.H.; Ruszymah, B.H. Insulin-Transferrin-Selenium Prevent Human Chondrocyte Dedifferentiation and Promote the Formation of High Quality Tissue Engineered Human Hyaline Cartilage. Eur. Cell Mater. 2005, 9, 58–67. [Google Scholar] [CrossRef]
- Wongin, S.; Waikakul, S.; Chotiyarnwong, P.; Siriwatwechakul, W.; Kino-oka, M.; Kim, M.-H.; Viravaidya-Pasuwat, K. Maintenance of Human Chondrogenic Phenotype on a Dendrimer-Immobilized Surface for an Application of Cell Sheet Engineering. BMC Biotechnol. 2018, 18, 14. [Google Scholar] [CrossRef]
- Mandl, E.W.; Jahr, H.; Koevoet, J.L.M.; van Leeuwen, J.P.T.M.; Weinans, H.; Verhaar, J.A.N.; van Osch, G.J.V.M. Fibroblast Growth Factor-2 in Serum-Free Medium Is a Potent Mitogen and Reduces Dedifferentiation of Human Ear Chondrocytes in Monolayer Culture. Matrix Biol. 2004, 23, 231–241. [Google Scholar] [CrossRef]
- Matsumoto, E.; Furumatsu, T.; Kanazawa, T.; Tamura, M.; Ozaki, T. ROCK Inhibitor Prevents the Dedifferentiation of Human Articular Chondrocytes. Biochem. Biophys. Res. Commun. 2012, 420, 124–129. [Google Scholar] [CrossRef]
- Mennan, C.; Garcia, J.; McCarthy, H.; Owen, S.; Perry, J.; Wright, K.; Banerjee, R.; Richardson, J.B.; Roberts, S. Human Articular Chondrocytes Retain Their Phenotype in Sustained Hypoxia While Normoxia Promotes Their Immunomodulatory Potential. Cartilage 2018, 10, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, D.H.; Matmati, M.; Khayat, G.; Chaudhry, S.; Hinz, B.; Quinn, T.M. Culture of Primary Bovine Chondrocytes on a Continuously Expanding Surface Inhibits Dedifferentiation. Tissue Eng. Part A 2012, 18, 2466–2476. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Q.; Cui, W.; Liu, F.; Wang, Q.; Chen, Z.; Fan, W. Co-Cultivated Mesenchymal Stem Cells Support Chondrocytic Differentiation of Articular Chondrocytes. Int. Orthop. 2013, 37, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Von Bomhard, A.; Faust, J.; Elsaesser, A.F.; Schwarz, S.; Pippich, K.; Rotter, N. Impact of Expansion and Redifferentiation under Hypothermia on Chondrogenic Capacity of Cultured Human Septal Chondrocytes. J. Tissue Eng. 2017, 8, 2041731417732655. [Google Scholar] [CrossRef] [PubMed]
- Gunja, N.J.; Athanasiou, K.A. Passage and Reversal Effects on Gene Expression of Bovine Meniscal Fibrochondrocytes. Arthritis Res. Ther. 2007, 9, R93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watt, F.M. Effect of Seeding Density on Stability of the Differentiated Phenotype of Pig Articular Chondrocytes in Culture. J. Cell Sci. 1988, 89, 373–378. [Google Scholar] [PubMed]
- Mao, Y.; Block, T.; Singh-Varma, A.; Sheldrake, A.; Leeth, R.; Griffey, S.; Kohn, J. Extracellular Matrix Derived from Chondrocytes Promotes Rapid Expansion of Human Primary Chondrocytes in Vitro with Reduced Dedifferentiation. Acta Biomater. 2019, 85, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; De Souza, P.; Merker, H.-J. Integrin Expression and Collagen Type II Implicated in Maintenance of Chondrocyte Shape in Monolayer Culture: An Immunomorphological Study. Cell Biol. Int. 1997, 21, 115–125. [Google Scholar] [CrossRef]
- Claus, S.; Aubert-Foucher, E.; Demoor, M.; Camuzeaux, B.; Paumier, A.; Piperno, M.; Damour, O.; Duterque-Coquillaud, M.; Galéra, P.; Mallein-Gerin, F. Chronic Exposure of Bone Morphogenetic Protein-2 Favors Chondrogenic Expression in Human Articular Chondrocytes Amplified in Monolayer Cultures. J. Cell. Biochem. 2010, 111, 1642–1651. [Google Scholar] [CrossRef] [PubMed]
- Egli, R.J.; Bastian, J.D.; Ganz, R.; Hofstetter, W.; Leunig, M. Hypoxic Expansion Promotes the Chondrogenic Potential of Articular Chondrocytes. J. Orthop. Res. 2008, 26, 977–985. [Google Scholar] [CrossRef]
- Li, X.; Liang, Y.; Xu, X.; Xiong, J.; Ouyang, K.; Duan, L.; Wang, D. Cell-to-Cell Culture Inhibits Dedifferentiation of Chondrocytes and Induces Differentiation of Human Umbilical Cord-Derived Mesenchymal Stem Cells. Biomed Res. Int. 2019, 2019, 5871698. [Google Scholar] [CrossRef] [Green Version]
- Aurich, M.; Hofmann, G.O.; Best, N.; Rolauffs, B. Induced Redifferentiation of Human Chondrocytes from Articular Cartilage Lesion in Alginate Bead Culture after Monolayer Dedifferentiation: An Alternative Cell Source for Cell-Based Therapies? Tissue Eng. Part A 2017, 24, 275–286. [Google Scholar] [CrossRef]
- Caron, M.M.J.; Emans, P.J.; Coolsen, M.M.E.; Voss, L.; Surtel, D.A.M.; Cremers, A.; van Rhijn, L.W.; Welting, T.J.M. Redifferentiation of Dedifferentiated Human Articular Chondrocytes: Comparison of 2D and 3D Cultures. Osteoarthr. Cartil. 2012, 20, 1170–1178. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Kawaguchi, H.; Ogasawara, T.; Asawa, Y.; Kishimoto, J.; Takahashi, T.; Chung, U.; Yamaoka, H.; Asato, H.; Nakamura, K. Optimal Combination of Soluble Factors for Tissue Engineering of Permanent Cartilage from Cultured Human Chondrocytes. J. Biol. Chem. 2007, 282, 20407–20415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, C.L.; Polak, J.M. Control of Human Articular Chondrocyte Differentiation by Reduced Oxygen Tension. J. Cell. Physiol. 2004, 199, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Markway, B.D.; Cho, H.; Johnstone, B. Hypoxia Promotes Redifferentiation and Suppresses Markers of Hypertrophy and Degeneration in both Healthy and Osteoarthritic Chondrocytes. Arthritis Res. Ther. 2013, 15, R92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, X.; Xie, Y.; Wang, G. Mechanical Stimulation Promotes the Proliferation and the Cartilage Phenotype of Mesenchymal Stem Cells and Chondrocytes Co-Cultured in Vitro. Biomed. Pharmacother. 2019, 117, 109146. [Google Scholar] [CrossRef]
- Krueger, S.; Achilles, S.; Zimmermann, J.; Tischer, T.; Bader, R.; Jonitz-Heincke, A. Re-Differentiation Capacity of Human Chondrocytes in Vitro Following Electrical Stimulation with Capacitively Coupled Fields. J. Clin. Med. 2019, 8, 1771. [Google Scholar] [CrossRef] [Green Version]
- Meretoja, V.V.; Dahlin, R.L.; Wright, S.; Kasper, F.K.; Mikos, A.G. Articular Chondrocyte Redifferentiation in 3D Co-Cultures with Mesenchymal Stem Cells. Tissue Eng. Part C. Methods 2014, 20, 514–523. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.W.; Lee, M.C.; Bae, H.C.; Han, H.-S. Direct Coculture of Human Chondrocytes and Synovium-Derived Stem Cells Enhances In Vitro Chondrogenesis. Cell J. 2018, 20, 53–60. [Google Scholar]
- Hendriks, J.A.A.; Miclea, R.L.; Schotel, R.; de Bruijn, E.; Moroni, L.; Karperien, M.; Riesle, J.; van Blitterswijk, C.A. Primary Chondrocytes Enhance Cartilage Tissue Formation upon Co-Culture with a Range of Cell Types. Soft Matter 2010, 6, 5080–5088. [Google Scholar] [CrossRef]
- Kudva, A.K.; Luyten, F.P.; Patterson, J. Initiating Human Articular Chondrocyte Re-Differentiation in a 3D System after 2D Expansion. J. Mater. Sci. Mater. Med. 2017, 28, 156. [Google Scholar] [CrossRef] [Green Version]
- Benya, P.D.; Shaffer, J.D. Dedifferentiated Chondrocytes Reexpress the Differentiated Collagen Phenotype When Cultured in Agarose Gels. Cell 1982, 30, 215–224. [Google Scholar] [CrossRef]
- Grigolo, B.; Lisignoli, G.; Piacentini, A.; Fiorini, M.; Gobbi, P.; Mazzotti, G.; Duca, M.; Pavesio, A.; Facchini, A. Evidence for Redifferentiation of Human Chondrocytes Grown on a Hyaluronan-Based Biomaterial (HYAFF®11): Molecular, Immunohistochemical and Ultrastructural Analysis. Biomaterials 2002, 23, 1187–1195. [Google Scholar] [CrossRef]
- Levett, P.A.; Melchels, F.P.W.; Schrobback, K.; Hutmacher, D.W.; Malda, J.; Klein, T.J. Chondrocyte Redifferentiation and Construct Mechanical Property Development in Single-Component Photocrosslinkable Hydrogels. J. Biomed. Mater. Res. Part A 2014, 102, 2544–2553. [Google Scholar] [CrossRef] [PubMed]
- Meinert, C.; Schrobback, K.; Hutmacher, D.W.; Klein, T.J. A Novel Bioreactor System for Biaxial Mechanical Loading Enhances the Properties of Tissue-Engineered Human Cartilage. Sci. Rep. 2017, 7, 16997. [Google Scholar] [CrossRef] [Green Version]
- Heyland, J.; Wiegandt, K.; Goepfert, C.; Nagel-Heyer, S.; Ilinich, E.; Schumacher, U.; Pörtner, R. Redifferentiation of Chondrocytes and Cartilage Formation under Intermittent Hydrostatic Pressure. Biotechnol. Lett. 2006, 28, 1641–1648. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-J.; Bhang, S.H.; La, W.-G.; Yang, H.S.; Seong, J.Y.; Lee, H.; Im, G.-I.; Lee, S.-H.; Kim, B.-S. Spinner-Flask Culture Induces Redifferentiation of de-Differentiated Chondrocytes. Biotechnol. Lett. 2011, 33, 829–836. [Google Scholar] [CrossRef]
- Hiemer, B.; Krogull, M.; Bender, T.; Ziebart, J.; Krueger, S.; Bader, R.; Jonitz-Heincke, A. Effect of Electric Stimulation on Human Chondrocytes and Mesenchymal Stem Cells under Normoxia and Hypoxia. Mol. Med. Rep. 2018, 18, 2133–2141. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, V.J.; Lee, A.; Anderson, J.; Parreno, J.; Theodoropoulos, J.; Backstein, D.; Kandel, R. Redifferentiated Chondrocytes in Fibrin Gel for the Repair of Articular Cartilage Lesions. Am. J. Sports Med. 2019, 47, 2348–2359. [Google Scholar] [CrossRef]
- Payr, S.; Tichy, B.; Atteneder, C.; Michel, M.; Tiefenboeck, T.; Lang, N.; Nuernberger, S.; Hajdu, S.; Rosado-Balmayor, E.; Marlovits, S.; et al. Redifferentiation of Aged Human Articular Chondrocytes by Combining Bone Morphogenetic Protein-2 and Melanoma Inhibitory Activity Protein in 3D-Culture. PLoS ONE 2017, 12, e0179729. [Google Scholar] [CrossRef] [Green Version]
- Jakob, M.; Démarteau, O.; Schäfer, D.; Hintermann, B.; Dick, W.; Heberer, M.; Martin, I. Specific Growth Factors during the Expansion and Redifferentiation of Adult Human Articular Chondrocytes Enhance Chondrogenesis and Cartilaginous Tissue Formation in Vitro. J. Cell. Biochem. 2001, 81, 368–377. [Google Scholar] [CrossRef]
- Jeyakumar, V.; Niculescu-Morzsa, E.; Bauer, C.; Lacza, Z.; Nehrer, S. Platelet-Rich Plasma Supports Proliferation and Redifferentiation of Chondrocytes during in Vitro Expansion. Front. Bioeng. Biotechnol. 2017, 5, 75. [Google Scholar] [CrossRef]
- Domm, C.; Schünke, M.; Christesen, K.; Kurz, B. Redifferentiation of Dedifferentiated Bovine Articular Chondrocytes in Alginate Culture under Low Oxygen Tension. Osteoarthr. Cartil. 2002, 10, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foldager, C.B.; Nielsen, A.B.; Munir, S.; Ulrich-Vinther, M.; Søballe, K.; Bünger, C.; Lind, M. Combined 3D and Hypoxic Culture Improves Cartilage-Specific Gene Expression in Human Chondrocytes. Acta Orthop. 2011, 82, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Meretoja, V.V.; Dahlin, R.L.; Kasper, F.K.; Mikos, A.G. Enhanced Chondrogenesis in Co-Cultures with Articular Chondrocytes and Mesenchymal Stem Cells. Biomaterials 2012, 33, 6362–6369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlin, R.L.; Ni, M.; Meretoja, V.V.; Kasper, F.K.; Mikos, A.G. TGF-Β3-Induced Chondrogenesis in Co-Cultures of Chondrocytes and Mesenchymal Stem Cells on Biodegradable Scaffolds. Biomaterials 2014, 35, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Song, X.; Li, T.; Xiao, J.; Chen, Y.; Gong, X.; Zeng, W.; Yang, L.; Chen, C. Pellet Coculture of Osteoarthritic Chondrocytes and Infrapatellar Fat Pad-Derived Mesenchymal Stem Cells with Chitosan/Hyaluronic Acid Nanoparticles Promotes Chondrogenic Differentiation. Stem Cell Res. Ther. 2017, 8, 264. [Google Scholar] [CrossRef]
- Scalzone, A.; Ferreira, A.M.; Tonda-Turo, C.; Ciardelli, G.; Dalgarno, K.; Gentile, P. The Interplay between Chondrocyte Spheroids and Mesenchymal Stem Cells Boosts Cartilage Regeneration within a 3D Natural-Based Hydrogel. Sci. Rep. 2019, 9, 14630. [Google Scholar] [CrossRef] [Green Version]
- Gan, L.; Kandel, R.A. In Vitro Cartilage Tissue Formation by Co-Culture of Primary and Passaged Chondrocytes. Tissue Eng. 2007, 13, 831–842. [Google Scholar] [CrossRef]
- Ecke, A.; Lutter, A.-H.; Scholka, J.; Hansch, A.; Becker, R.; Anderer, U. Tissue Specific Differentiation of Human Chondrocytes Depends on Cell Microenvironment and Serum Selection. Cells 2019, 8, 934. [Google Scholar] [CrossRef] [Green Version]
- Tallheden, T.; van der Lee, J.; Brantsing, C.; Månsson, J.-E.; Sjögren-Jansson, E.; Lindahl, A. Human Serum for Culture of Articular Chondrocytes. Cell Transplant. 2005, 14, 469–479. [Google Scholar] [CrossRef] [Green Version]
- Chua, K.H.; Aminuddin, B.S.; Fuzina, N.H.; Ruszymah, B.H. Basic Fibroblast Growth Factor with Human Serum Supplementation: Enhancement of Human Chondrocyte Proliferation and Promotion of Cartilage Regeneration. Singap. Med. J. 2007, 48, 324–332. [Google Scholar]
- Anderer, U.; Libera, J. In Vitro Engineering of Human Autogenous Cartilage. J. Bone Miner. Res. 2002, 17, 1420–1429. [Google Scholar] [CrossRef] [PubMed]
- Malpeli, M.; Randazzo, N.; Cancedda, R.; Dozin, B. Serum-Free Growth Medium Sustains Commitment of Human Articular Chondrocyte through Maintenance of Sox9 Expression. Tissue Eng. 2004, 10, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P.; Cheng, K.H.; Yan, C.H.; Chan, B.P. Collagen Microsphere Based 3D Culture System for Human Osteoarthritis Chondrocytes (HOACs). Sci. Rep. 2019, 9, 12453. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Neil, D.A.; Lin, L.; Fu, X.; Zhang, J.; Yu, C. Serum-Free Media for Articular Chondrocytes in Vitro Expansion. Chin. Med. J. (Engl.) 2013, 126, 2523–2529. [Google Scholar] [PubMed]
- Ho, S.T.B.; Yang, Z.; Ho, S.T.B.; Yang, Z.; Hui, H.P.J.; Ho, S.T.B.; Yang, Z.; Hui, H.P.J.; Oh, K.W.S.; Ho, S.T.B.; et al. A Serum Free Approach towards the Conservation of Chondrogenic Phenotype during in Vitro Cell Expansion. Growth Factors 2009, 27, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Martinez, I.; Elvenes, J.; Olsen, R.; Bertheussen, K.; Johansen, O. Redifferentiation of in Vitro Expanded Adult Articular Chondrocytes by Combining the Hanging-Drop Cultivation Method with Hypoxic Environment. Cell Transplant. 2008, 17, 987–996. [Google Scholar] [CrossRef]
- Gigout, A.; Jolicoeur, M.; Buschmann, M.D. Low Calcium Levels in Serum-Free Media Maintain Chondrocyte Phenotype in Monolayer Culture and Reduce Chondrocyte Aggregation in Suspension Culture. Osteoarthr. Cartil. 2005, 13, 1012–1024. [Google Scholar] [CrossRef] [Green Version]
- Stewart, M.C.; Saunders, K.M.; Burton-Wurster, N.; Macleod, J.N. Phenotypic Stability of Articular Chondrocytes in Vitro: The Effects of Culture Models, Bone Morphogenetic Protein 2, and Serum Supplementation. J. Bone Miner. Res. 2000, 15, 166–174. [Google Scholar] [CrossRef]
- Law, J.X.; Chowdhury, S.R.; Saim, A.B.; Idrus, R.B.H. Platelet-Rich Plasma with Keratinocytes and Fibroblasts Enhance Healing of Full-Thickness Wounds. J. Tissue Viability 2017, 26, 208–215. [Google Scholar] [CrossRef]
- Xian, L.J.; Chowdhury, S.R.; Saim, A.B.; Idrus, R.B.H. Concentration-Dependent Effect of Platelet-Rich Plasma on Keratinocyte and Fibroblast Wound Healing. Cytotherapy 2015, 17, 293–300. [Google Scholar] [CrossRef]
- Kardos, D.; Simon, M.; Vácz, G.; Hinsenkamp, A.; Holczer, T.; Cseh, D.; Sárközi, A.; Szenthe, K.; Bánáti, F.; Szathmary, S.; et al. The Composition of Hyperacute Serum and Platelet-Rich Plasma Is Markedly Different despite the Similar Production Method. Int. J. Mol. Sci. 2019, 20, 721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akeda, K.; An, H.S.; Okuma, M.; Attawia, M.; Miyamoto, K.; Thonar, E.-M.A.; Lenz, M.E.; Sah, R.L.; Masuda, K. Platelet-Rich Plasma Stimulates Porcine Articular Chondrocyte Proliferation and Matrix Biosynthesis. Osteoarthr. Cartil. 2006, 14, 1272–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drengk, A.; Zapf, A.; Stürmer, E.K.; Stürmer, K.M.; Frosch, K.-H. Influence of Platelet-Rich Plasma on Chondrogenic Differentiation and Proliferation of Chondrocytes and Mesenchymal Stem Cells. Cells Tissues Organs 2009, 189, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Balakumar, B.; Rajagopal, K.; Madhuri, V. Bone Marrow Extract as a Growth Supplement for Human Iliac Apophyseal Chondrocyte Culture. Indian J. Med. Res. 2016, 144, 831–837. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| FBS | HPL | Defined Medium | |
|---|---|---|---|
| Pros | Abundant and easily available | Easy to produce | Components are well-defined |
| Cheaper compared to HPL and chemically defined medium | A universal growth supplement that is suitable for most human and animal cells | No batch-to-batch variation | |
| A universal growth supplement that is suitable for most human and animal cells | Contains most of the factors required for cell survival and proliferation | No risk of disease transmission | |
| Contains most of the factors required for cell survival and proliferation | No risk of xenogeneic immune reaction | No risk of xenogeneic/allogeneic protein contamination | |
| Used by most of the studies and publications | Can be prepared using autologous blood | No unintended interaction with test substances | |
| Cons | Components are ill-defined | Components are ill-defined | Not available for certain cells |
| Batch-to-batch variation | Batch-to-batch variation (reduced by pooling) | Some defined medium required extra coating to promote cell attachment | |
| A potential source of animal microbial contaminants | Unintended interaction with test substances | More expansive compared to FBS and HPL | |
| Risk of animal protein contamination on cells prepared for clinical usage | Fewer vendors distribute the product | Time consuming and difficult to develop | |
| Unintended interaction with test substances | Potential eliciting allogeneic immune response | ||
| Ethical concerns with animal welfare | Risk of transmitting human viruses |
| References | Source of Chondrocytes | Proliferation | Dedifferentiation | Redifferentiation | Other Key Findings |
|---|---|---|---|---|---|
| [35] | Human | HPL increases chondrocyte proliferation compared to FBS. | HPL and FBS fail to prevent chondrocyte dedifferentiation. | - | Chondrocytes cultured with HPL have lower expression of col X, a chondrocyte hypertrophic marker compared to those cultured with FBS. |
| [39] | Human | HPL increases chondrocyte proliferation compared to FBS. | HPL and FBS fail to prevent chondrocyte dedifferentiation. | Chondrocytes cultured with HPL maintains its redifferentiation potential after 10 population doublings and those cultured with FBS lost its redifferentiation potential after 3 population doublings. | Chondrocytes cultured with HPL in the presence of IL-1α demonstrate a transient activation and resolution of inflammation. Chondrocytes cultured with HPL secrete chondrocyte chemoattractants. |
| [8] | Human | HPL increases chondrocyte proliferation compared to FBS. | - | Chondrocytes cultured with HPL have poorer redifferentiation potential. | - |
| [40] | Human | HPL increases chondrocyte proliferation compared to FBS. | - | At the same population doubling number, chondrocytes cultured with HPL show better redifferentiation potential compared to those cultured with FBS. | - |
| [41] | Bovine | HPL increases chondrocyte proliferation compared to FBS. | - | - | Chondrocytes cultured in 3D with HPL showed poorer formation of cartilaginous matrix compared to those cultured with FBS. |
| [42] | Human | Addition of HPL to FBS promotes chondrocyte proliferation compared to FBS alone. | - | - | Chondrocytes cultured in 3D with FBS + HPL form cartilage-like tissue. |
| [43] | Human | HPL increases chondroprogenitor cell proliferation compared to FBS. | - | - | Chondroprogenitor cells cultured HPL have lower expression of col I, II, X, and aggrecan compared to those expanded with FBS. |
| Strategy | Specific Factor | References |
|---|---|---|
| Seeding density | High seeding density | [54] |
| Cell passaging | Reduce number of passaging | [50] |
| Surface substrate | Polyamidoamine dendrimer | [46] |
| PLLA | [44] | |
| Chondrocytes secreted ECM | [55] | |
| Collagen type II | [56] | |
| Chemical compound | Insulin-transferrin-selenium | [45] |
| FGF-2 | [47] | |
| BMP-2 | [57] | |
| ROCK inhibitor | [48] | |
| Hypoxia | 2% O2 | [49] |
| 1.5% O2 | [58] | |
| Hypothermia | 32.2 °C | [52] |
| Co-culture | Ratio of chondrocytes: MSCs; 2:1 | [51] |
| Ratio of chondrocytes: MSCs; 1:1 | [59] |
| Strategy | Specific Factor | References |
|---|---|---|
| 3D culture | Barium-alginate nanofiber | [30] |
| Alginate bead | [60] | |
| Cell pellet | [61] | |
| Polyethylene glycol hydrogel | [70] | |
| Agarose hydrogel | [71] | |
| Hyaluronan hydrogel | [72] | |
| Hyaluronic acid methyacrylate hydrogel | [73] | |
| Mechanical stimulation | Cyclic sinusoidal dynamic tensile mechanical stimulation | [65] |
| Shear and/or compression stress and gelatin methacryloyl/hyaluronic acid methacrylate | [74] | |
| Intermittent hydrostatic pressure and alginate bead | [75] | |
| Spinner flask | [76] | |
| Electrical stimulation | Capacitively coupled electric field stimulation | [66] |
| Alternating electrical stimulation and hypoxia | [77] | |
| Chemical compound | TGF-β3 and high seeding density | [78] |
| BMP-2 and alginate bead | [79] | |
| BMP-2 and insulin | [62] | |
| TGF-β1 and dexamethasone | [80] | |
| Platelet-rich plasma | [81] | |
| Hypoxia and 3D culture | 5% O2 and alginate bead | [82] |
| 1% and 5% O2 and MPEG-PLGA scaffold | [83] | |
| 3D co-culture | Ratio of chondrocytes: MSCs; 3:7 | [67] |
| Ratio of chondrocytes: MSCs; 1:3 | [84] | |
| Ratio of chondrocytes: MSCs; 1:3 | [85] | |
| Ratio of chondrocytes: MSCs; 1:1 | [86] | |
| Chondrocyte pellet on MSCs-laden chitosan/β-glycerophosphate hydrogel | [87] | |
| Ratio of chondrocytes: synovium derived stem cells; 1:1 | [68] | |
| Ratio of chondrocytes: primary chondrocytes; 4:1 | [88] | |
| Ratio of chondrocytes: primary chondrocytes; 4:1 | [69] |
| References | Source of Chondrocytes | Proliferation | Dedifferentiation | Redifferentiation | Other Key Findings |
|---|---|---|---|---|---|
| [89] | Human | HS increases chondrocyte proliferation compared to FBS. | HS is inferior compared to FBS in preventing chondrocyte dedifferentiation. | HS is inferior compared to FBS in promoting chondrocyte redifferentiation. | - |
| [90] | Human | HS increases chondrocyte proliferation compared to FBS. | - | - | - |
| [91] | Human | HS increases chondrocyte proliferation compared to FBS. | HS and FBS are comparable in maintenance of chondrocyte markers. | - | Supplement of 5 ng/mL bFGF enhances chondrocyte proliferation. The HS expanded chondrocytes form good cartilage in vivo. |
| [92] | Human | HS increases chondrocyte proliferation compared to FBS. | - | - | Substitution of HS with FBS delayed cartilage-like tissue formation and a hollow central area is formed at 3 months. |
| References | Medium | Source of Chondrocytes | Proliferation | Dedifferentiation | Redifferentiation | Other Key Findings |
|---|---|---|---|---|---|---|
| [93] | Coon’s modified Ham’s F12 medium with 5 ng/mL each of FGF-2, PDGF-bb, and EGF, 5 µg/mL insulin, 10−8 M dexamethasone, 50 µg/mL ascorbic acid, 50 µg/mL human transferrin, 2% human serum albumin, 6.25 µM linoleic acid, 30 µg/mL cholesterol, 5 × 10−5 M 2-mercaptoethanol, 30 nM selenium, 33 µM biotin, and 17 µM sodium pantothenate | Human | SFM increases chondrocyte proliferation compared to FBS. | SFM preserves the SOX9 expression better than FBS. | Chondrocytes cultured with SFM can redifferentiate spontaneously in 3D culture while those expanded with FBS requires induction factors to redifferentiate. | - |
| [94] | DMEM medium with 1% ITS-A, 0.4 µM proline, 50 µg/mL ascorbic acid, 10 mM HEPES, 0.1 mM non-essential amino acids, 2 mM L-glutamine and 1% antibiotic-antimycotic | Human | - | SFM preserves the SOX9 expression of 3D cultured chondrocytes better than FBS. | - | - |
| [95] | DMEM/F12 medium with 10 ng/mL FGF-2 and 10 ng/mL PDGF. DMEM/F12 medium with 10 ng/mL FGF-2 and 10 ng/mL IGF-1. | Bovine | Chondrocytes expanded with SFM and FBS have similar pace of cell proliferation. | SFM is superior compared to FBS in preventing chondrocyte dedifferentiation. | Chondrocytes expanded with SFM and FBS have similar redifferentiation potential in 3D culture. | - |
| [96] | DMEM/F12 medium with 10% Knockout SR, 2 ng/mL FGF-2, 2 ng/mL PDGF-AB, 2 ng/mL EGF and 10−8 M dexamethasone | Human | SFM increases chondrocyte proliferation compared to FBS. | - | Chondrocytes cultured with SFM show better redifferentiation in 3D culture compared to those expanded with FBS. | Chondrocytes cultured with SFM have lower expression of col X compared to those expanded with FBS upon redifferentiation. |
| [97] | Tissue SS supplements for biotechnology (Medi-Cult A/S) | Human | - | - | Chondrocytes expanded with SFM shows poorer redifferentiation in 3D culture compared those cultured with HS in hypoxic environment. | - |
| [98] | DMEM/F12 medium with 0.4 mM proline, 1.5 mM glutamine, 22 mM sodium bicarbonate, 8.9 mg/L alanine, 15 mg/L asparagine, 13.3 mg/L aspartic acid,14.5 mg/L glutamic acid, 7.5 mg/L glycine, 11.5 mg/L proline, 10.5 mg/L serine, 12.5 mM HEPES, 1× penicillin/streptomycin, 10 µg/mL insulin, 5.5 µg/mL transferrin, 0.05% (w/v) bovine serum albumin, 1.7 mM linoleic acid, 0.5 µg/mL sodium selenite, 5 × 10−5 M 2-mercaptoethanol and 10−8 M dexamethasone | Bovine | Chondrocytes expanded with SFM and FBS have similar pace of cell proliferation. | SFM lowers the expression of col I compared to FBS. | - | Lower calcium concentration in SFM is favourable to reduce expression of col I. |
| [99] | Opti-MEM (Gibco) | Equine | - | Chondrocytes cultured with SFM have higher expression of col II and aggrecan as well as lower col I compared to those cultured with FBS. | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liau, L.L.; Hassan, M.N.F.b.; Tang, Y.L.; Ng, M.H.; Law, J.X. Feasibility of Human Platelet Lysate as an Alternative to Foetal Bovine Serum for In Vitro Expansion of Chondrocytes. Int. J. Mol. Sci. 2021, 22, 1269. https://doi.org/10.3390/ijms22031269
Liau LL, Hassan MNFb, Tang YL, Ng MH, Law JX. Feasibility of Human Platelet Lysate as an Alternative to Foetal Bovine Serum for In Vitro Expansion of Chondrocytes. International Journal of Molecular Sciences. 2021; 22(3):1269. https://doi.org/10.3390/ijms22031269
Chicago/Turabian StyleLiau, Ling Ling, Muhammad Najib Fathi bin Hassan, Yee Loong Tang, Min Hwei Ng, and Jia Xian Law. 2021. "Feasibility of Human Platelet Lysate as an Alternative to Foetal Bovine Serum for In Vitro Expansion of Chondrocytes" International Journal of Molecular Sciences 22, no. 3: 1269. https://doi.org/10.3390/ijms22031269