Optical Assessment of Nociceptive TRP Channel Function at the Peripheral Nerve Terminal

 ,
,  ,
,
Abstract
:1. Introduction
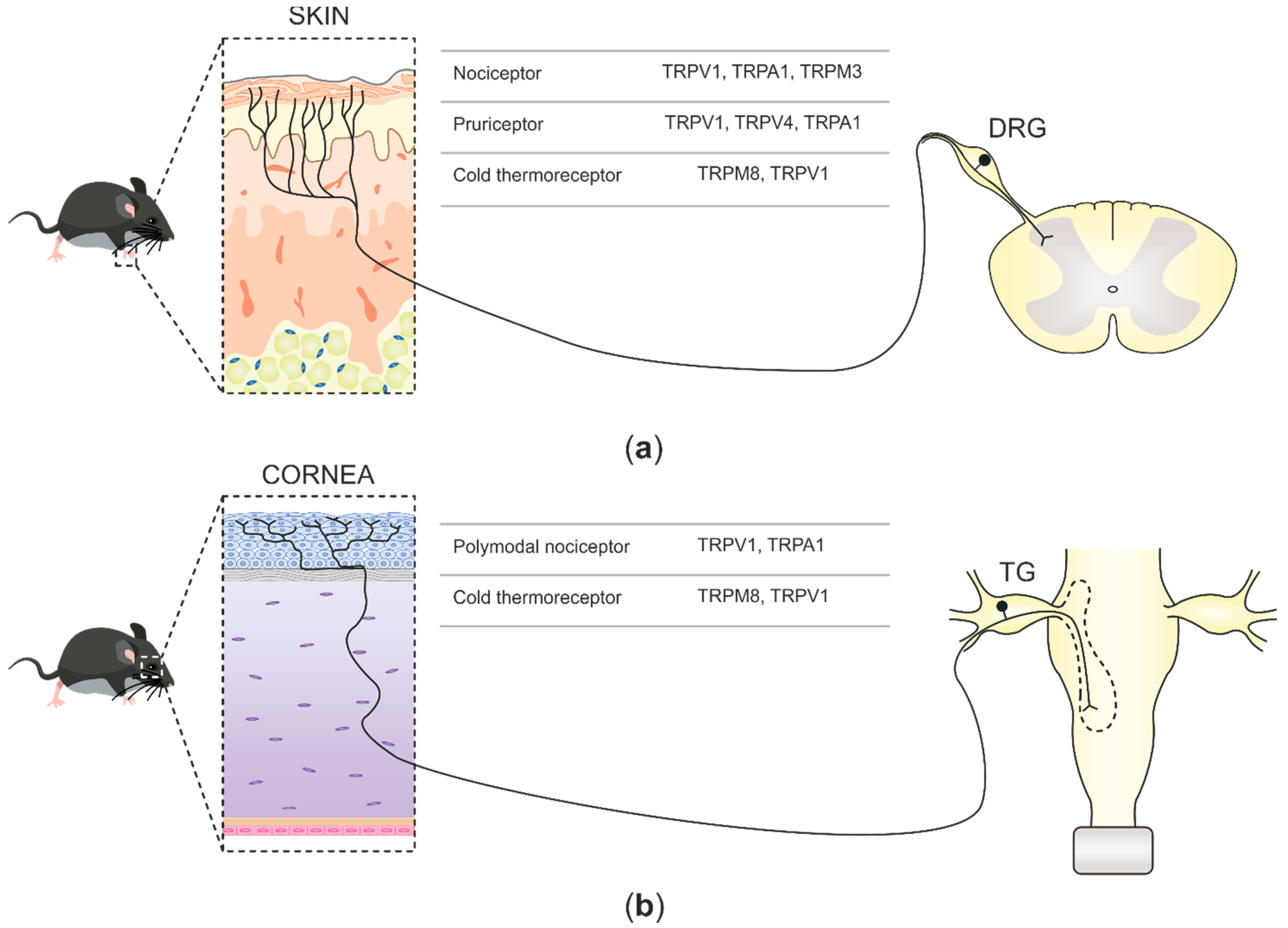
2. Nociceptive TRP Channels at Sensory Nerve Fibers
2.1. TRP Channels at Skin Nerve Endings
2.2. TRP Channels at Corneal Nerve Terminals
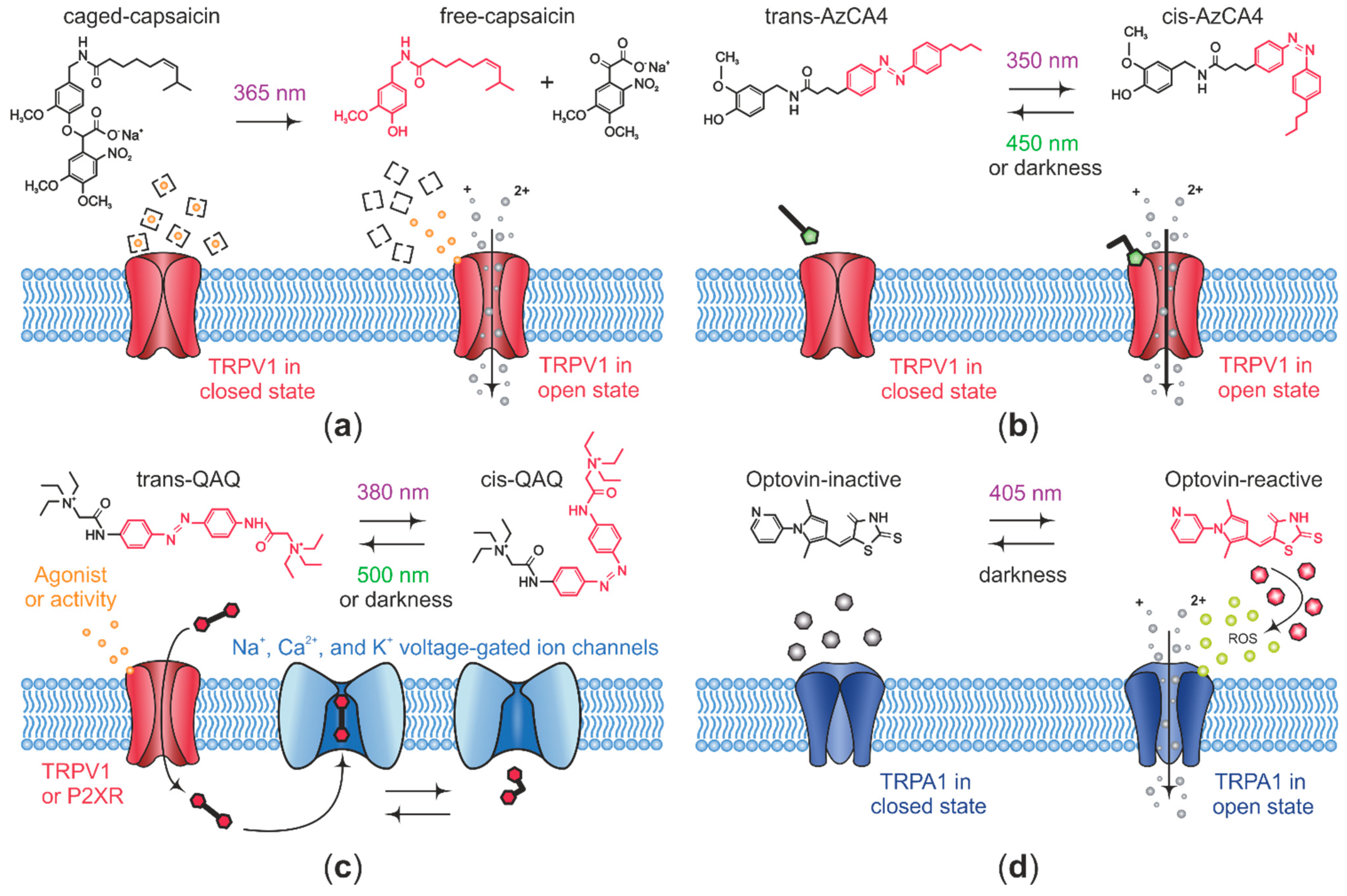
3. Opto-Pharmacological Control of Nociceptive TRP Channel Activity at the Nerve Terminal

{kind=link}
{kind=link}
{kind=link}
| Opto-Chemical Agent | Target | Activity | Opto-Chemical Strategy | Testing Method |
|---|---|---|---|---|
| Caged-capsaicin [116] | TRPV1 | agonist | Caged ligand | in vitro: rat DRG neurons. |
| AC4 1 [110] | TRPV1 | antagonist | Azobenzene-based photochromic ligand | in vitro: recombinant rat TRPV1. |
| ABCTC 2 [110] | TRPV1 | antagonist | Azobenzene-based photochromic ligand | in vitro: recombinant rat TRPV1. |
| AzCA4 3 [111] | TRPV1 | agonist | Azobenzene-based photochromic ligand | in vitro: mouse DRG neurons. ex vivo: skin nerve preparation. |
| Optovin [126] | TRPA1 | agonist | Rhodamine-containing photochromic ligand | in vitro: recombinant human TRPA1 and mouse DRG neurons. |
| TRP-switch-A [127] | zTRPA1b 4 | agonist | Azobenzene-based photochromic ligand | in vitro: recombinant zebrafish TRPA1b. |
| TRP-switch-B [127] | zTRPA1b 4 | agonist | Azopyrazole-containing photochromic ligand | in vitro: recombinant zebrafish TRPA1b. |
| QAQ 5 [118] | Navs, Kvs and VGCC | antagonist | Azobenzene-based photochromic ligand | in vitro: rat TG neurons. ex vivo: mouse DRG. in vivo: Von Frey assay. |
| QENAQ 6 [123] | Navs and Kvs | antagonist | Azobenzene-based photochromic ligand | in vitro: mouse TG neurons. ex vivo: mouse DRG. |
| PhotoS1P [121] | S1P 7 | agonist | Azobenzene-based photochromic ligand | in vitro: mouse DRG neurons. in vivo: Hargreaves test. |
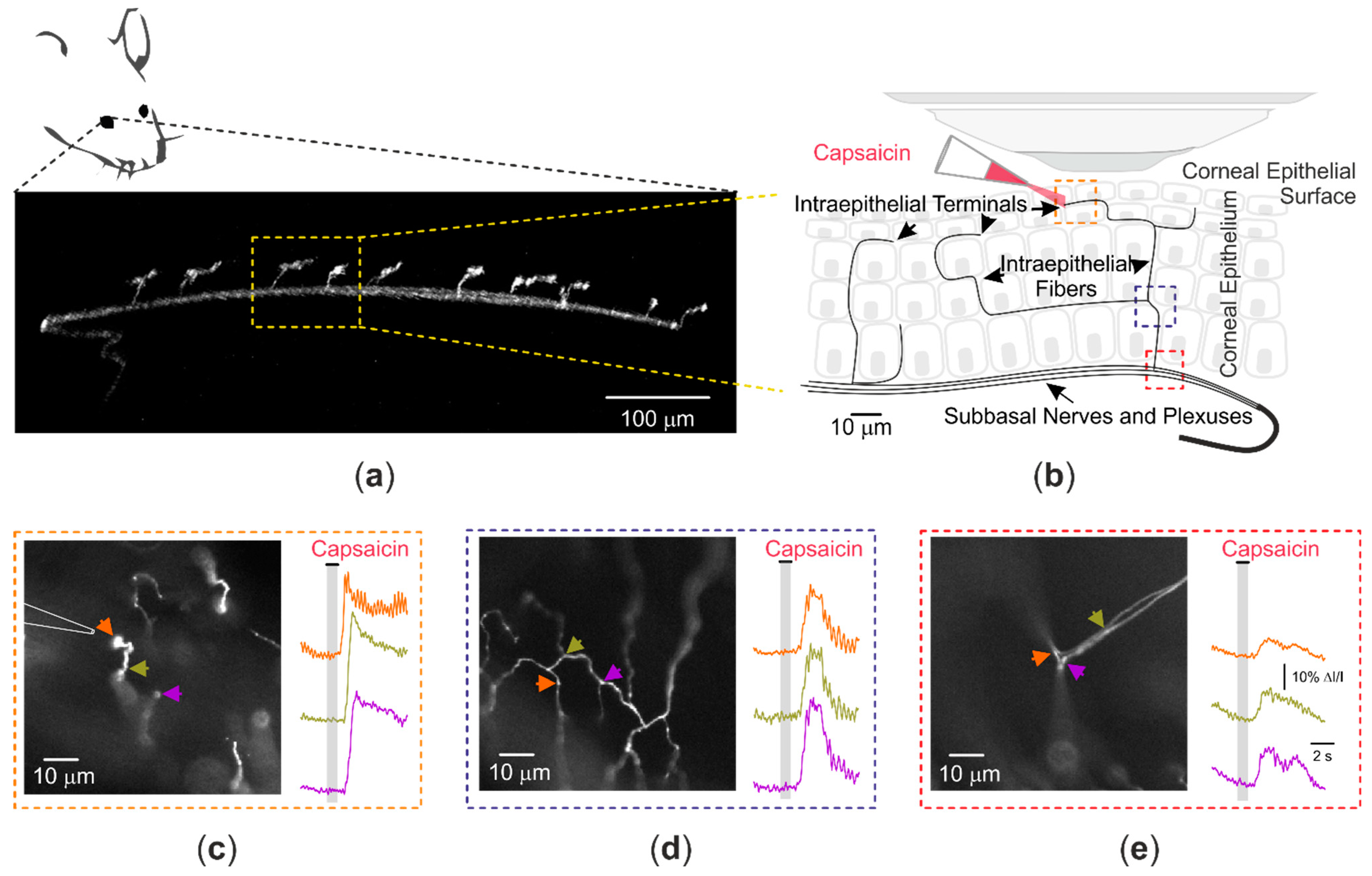
4. Optical Recording of Nociceptive TRP Channel Activity at the Nerve Terminal
5. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Amir, R.; Devor, M. Electrical excitability of the soma of sensory neurons is required for spike invasion of the soma, but not for through-conduction. Biophys. J. 2003, 84, 2181–2191. [Google Scholar] [CrossRef] [Green Version]
- Mense, S. Anatomy of Nociceptors. Senses A Compr. Ref. 2010, 5, 11–41. [Google Scholar] [CrossRef]
- Kandel, E.R.; Mack, S.; Jessell, T.M.; Schwartz, J.H.; Siegelbaum, S.A.; Hudspeth, A.J. Principles of Neural Science, 5th ed.; McGraw-Hill Education: New York, NY, USA, 2013; ISBN 9780071390118. [Google Scholar]
- Dubin, A.E.; Patapoutian, A. Nociceptors: The sensors of the pain pathway. J. Clin. Investig. 2010, 120, 3760–3772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djouhri, L.; Lawson, S.N. Aβ-fiber nociceptive primary afferent neurons: A review of incidence and properties in relation to other afferent A-fiber neurons in mammals. Brain Res. Rev. 2004, 46, 131–145. [Google Scholar] [CrossRef]
- Woolf, C.J.; Ma, Q. Nociceptors-Noxious Stimulus Detectors. Neuron 2007, 55, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Weng, X.; Smith, T.; Sathish, J.; Djouhri, L. Chronic inflammatory pain is associated with increased excitability and hyperpolarization-activated current (I h) in C- but not Aδ-nociceptors. Pain 2012, 153, 900–914. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and Molecular Mechanisms of Pain. Cell 2009, 139, 267–284. [Google Scholar] [CrossRef] [Green Version]
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef]
- Nilius, B.; Owsianik, G.; Voets, T.; Peters, J.A. Transient receptor potential cation channels in disease. Physiol. Rev. 2007, 87, 165–217. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Chen, C.C. Roles of ASICs in nociception and proprioception. Adv. Exp. Med. Biol. 2018, 1099, 37–47. [Google Scholar] [CrossRef]
- Deval, E.; Noël, J.; Lay, N.; Alloui, A.; Diochot, S.; Friend, V.; Jodar, M.; Lazdunski, M.; Lingueglia, E. ASIC3, a sensor of acidic and primary inflammatory pain. EMBO J. 2008, 27, 3047–3055. [Google Scholar] [CrossRef] [PubMed]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murthy, S.E.; Loud, M.C.; Daou, I.; Marshall, K.L.; Schwaller, F.; Kühnemund, J.; Francisco, A.G.; Keenan, W.T.; Dubin, A.E.; Lewin, G.R.; et al. The mechanosensitive ion channel Piezo2 mediates sensitivity to mechanical pain in mice. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaulieu-Laroche, L.; Christin, M.; Donoghue, A.; Agosti, F.; Yousefpour, N.; Petitjean, H.; Davidova, A.; Stanton, C.; Khan, U.; Dietz, C.; et al. TACAN Is an Ion Channel Involved in Sensing Mechanical Pain. Cell 2020, 180, 956–967.e17. [Google Scholar] [CrossRef]
- Premkumar, L.S. Methods Used for Studying TRP Channel Functions in Sensory Neurons. In TRP Channels; Zhu, M.X., Ed.; CRC Press: Boca Raton, FL, USA, 2011; ISBN 978-1-4398-1860-2. [Google Scholar]
- Malin, S.A.; Davis, B.M.; Molliver, D.C. Production of dissociated sensory neuron cultures and considerations for their use in studying neuronal function and plasticity. Nat. Protoc. 2007, 2, 152–160. [Google Scholar] [CrossRef]
- Meseguer, M.V.; L. Denlinger, B.; Talavera, K. Methodological Considerations to Understand the Sensory Function of TRP Channels. Curr. Pharm. Biotechnol. 2010, 12, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Barkai, O.; Butterman, R.; Katz, B.; Lev, S.; Binshtok, A.M. The input-output relation of primary nociceptive neurons is determined by the morphology of the peripheral nociceptive terminals. J. Neurosci. 2020, 40, 9346–9363. [Google Scholar] [CrossRef]
- Zimmermann, K.; Hein, A.; Hager, U.; Kaczmarek, J.S.; Turnquist, B.P.; Clapham, D.E.; Reeh, P.W. Phenotyping sensory nerve endings in vitro in the mouse. Nat. Protoc. 2009, 4, 174–196. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, K.; Lennerz, J.K.; Hein, A.; Link, A.S.; Stefan Kaczmarek, J.; Delling, M.; Uysal, S.; Pfeifer, J.D.; Riccio, A.; Clapham, D.E. Transient receptor potential cation channel, subfamily C, member 5 (TRPC5) is a cold-transducer in the peripheral nervous system. Proc. Natl. Acad. Sci. USA 2011, 108, 18114–18119. [Google Scholar] [CrossRef] [Green Version]
- Paricio-Montesinos, R.; Schwaller, F.; Udhayachandran, A.; Rau, F.; Walcher, J.; Evangelista, R.; Vriens, J.; Voets, T.; Poulet, J.F.A.; Lewin, G.R. The Sensory Coding of Warm Perception. Neuron 2020, 106, 830–841.e3. [Google Scholar] [CrossRef] [Green Version]
- Parra, A.; Madrid, R.; Echevarria, D.; Del Olmo, S.; Morenilla-Palao, C.; Acosta, M.C.; Gallar, J.; Dhaka, A.; Viana, F.; Belmonte, C. Ocular surface wetness is regulated by TRPM8-dependent cold thermoreceptors of the cornea. Nat. Med. 2010, 16, 1396–1399. [Google Scholar] [CrossRef] [PubMed]
- Piña, R.; Ugarte, G.; Campos, M.; Íñigo-Portugués, A.; Olivares, E.; Orio, P.; Belmonte, C.; Bacigalupo, J.; Madrid, R. Role of TRPM8 Channels in Altered Cold Sensitivity of Corneal Primary Sensory Neurons Induced by Axonal Damage. J. Neurosci. 2019, 39, 8177–8192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, R.H.; Katz, B.; Lev, S.; Binshtok, A.M. Ultrafast optical recording reveals distinct capsaicin-induced ion dynamics along single nociceptive neurite terminals in vitro. J. Biomed. Opt. 2017, 22, 076010. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, R.H.; Barkai, O.; Íñigo-Portugués, A.; Katz, B.; Lev, S.; Binshtok, A.M. Location and Plasticity of the Sodium Spike Initiation Zone in Nociceptive Terminals In Vivo. Neuron 2019, 102, 801–812.e5. [Google Scholar] [CrossRef] [PubMed]
- Mulier, M.; Van Ranst, N.; Corthout, N.; Munck, S.; Berghe, P.V.; Vriens, J.; Voets, T.; Moilanen, L. Upregulation of TRPM3 in nociceptors innervating inflamed tissue. Elife 2020, 9, e61103. [Google Scholar] [CrossRef]
- Kim, Y.S.; Chu, Y.; Han, L.; Li, M.; Li, Z.; LaVinka, P.C.; Sun, S.; Tang, Z.; Park, K.; Caterina, M.J.; et al. Central terminal sensitization of TRPV1 by descending serotonergic facilitation modulates chronic pain. Neuron 2014, 81, 873–887. [Google Scholar] [CrossRef] [Green Version]
- Baillie, L.D.; Hagen, V.; Gardner, K.M.; Mulligan, S.J. Functional imaging within individual pain fibres ex vivo with optical microscopy. J. Neurosci. Methods 2011, 198, 274–279. [Google Scholar] [CrossRef]
- Gover, T.D.; Moreira, T.H.V.; Kao, J.P.Y.; Weinreich, D. Calcium regulation in individual peripheral sensory nerve terminals of the rat. J. Physiol. 2007, 578, 481–490. [Google Scholar] [CrossRef]
- Flockerzi, V.; Nilius, B. TRPs: Truly remarkable proteins. In Handbook of Experimental Pharmacology; Flockerzi, V., Nilius, B., Eds.; Springer: Cham, Germany, 2014; Volume 222, pp. 1–12. ISBN 978-3-642-54215-2. [Google Scholar]
- Fernández-Carvajal, A.; Fernández-Ballester, G.; González-Muñiz, R.; Ferrer-Montiel, A. Pharmacology of TRP Channels. In TRP Channels in Sensory Transduction; Madrid, R., Bacigalupo, J., Eds.; Springer: Cham, Germany, 2015; pp. 41–71. ISBN 978-3-319-18705-1. [Google Scholar]
- Iseppon, F.; Arcangeletti, M. Optogenetics and photopharmacology in pain research and therapeutics. STEMedicine 2020, 1, e43. [Google Scholar] [CrossRef]
- Kramer, R.H.; Mourot, A.; Adesnik, H. Optogenetic pharmacology for control of native neuronal signaling proteins. Nat. Neurosci. 2013, 16, 816–823. [Google Scholar] [CrossRef]
- Boyden, E.S.; Zhang, F.; Bamberg, E.; Nagel, G.; Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 2005, 8, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Sun, Y.; Cai, M.; Zhao, Y.; Cao, W.; Liu, Z.; Cui, G.; Tang, B. Copper sulfide nanoparticles as a photothermal switch for TRPV1 signaling to attenuate atherosclerosis. Nat. Commun. 2018, 9, 231. [Google Scholar] [CrossRef] [PubMed]
- Munshi, R.; Qadri, S.M.; Zhang, Q.; Castellanos Rubio, I.; Del Pino, P.; Pralle, A. Magnetothermal genetic deep brain stimulation of motor behaviors in awake, freely moving mice. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Moisset, X.; Lanteri-Minet, M.; Fontaine, D. Neurostimulation methods in the treatment of chronic pain. J. Neural Transm. 2020, 127, 673–686. [Google Scholar] [CrossRef]
- Paoletti, P.; Ellis-Davies, G.C.R.; Mourot, A. Optical control of neuronal ion channels and receptors. Nat. Rev. Neurosci. 2019, 20, 514–532. [Google Scholar] [CrossRef]
- Venkatachalam, K.; Montell, C. TRP channels. Annu. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef] [Green Version]
- Mickle, A.D.; Shepherd, A.J.; Mohapatra, D.P. Sensory TRP channels: The key transducers of nociception and pain. Prog. Mol. Biol. Transl. Sci. 2015, 131, 73–118. [Google Scholar] [CrossRef] [Green Version]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Fukuoka, T.; Obata, K.; Yamanaka, H.; Dai, Y.; Tokunaga, A.; Noguchi, K. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with adelta/c-fibers and colocalization with trk receptors. J. Comp. Neurol. 2005, 493, 596–606. [Google Scholar] [CrossRef]
- Hwang, S.J.; Min Oh, J.; Valtschanoff, J.G.; Oh, J.M.; Valtschanoff, J.G. Expression of the vanilloid receptor TRPV1 in rat dorsal root ganglion neurons supports different roles of the receptor in visceral and cutaneous afferents. Brain Res. 2005, 1047, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Story, G.M.; Peier, A.M.; Reeve, A.J.; Eid, S.R.; Mosbacher, J.; Hricik, T.R.; Earley, T.J.; Hergarden, A.C.; Andersson, D.A.; Hwang, S.W.; et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Sawada, Y.; Hosokawa, H.; Hori, A.; Matsumura, K.; Kobayashi, S. Cold sensitivity of recombinant TRPA1 channels. Brain Res. 2007, 1160, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Karashima, Y.; Talavera, K.; Everaerts, W.; Janssens, A.; Kwan, K.Y.; Vennekens, R.; Nilius, B.; Voets, T. TRPA1 acts as a cold sensor in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2009, 106, 1273–1278. [Google Scholar] [CrossRef] [Green Version]
- Bautista, D.M.; Jordt, S.-E.; Nikai, T.; Tsuruda, P.R.; Read, A.J.; Poblete, J.; Yamoah, E.N.; Basbaum, A.I.; Julius, D. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell 2006, 124, 1269–1282. [Google Scholar] [CrossRef] [Green Version]
- Knowlton, W.M.; Bifolck-Fisher, A.; Bautista, D.M.; McKemy, D.D. TRPM8, but not TRPA1, is required for neural and behavioral responses to acute noxious cold temperatures and cold-mimetics in vivo. Pain 2010, 150, 340–350. [Google Scholar] [CrossRef] [Green Version]
- Vandewauw, I.; De Clercq, K.; Mulier, M.; Held, K.; Pinto, S.; Van Ranst, N.; Segal, A.; Voet, T.; Vennekens, R.; Zimmermann, K.; et al. A TRP channel trio mediates acute noxious heat sensing. Nature 2018, 555, 662–666. [Google Scholar] [CrossRef]
- Sinica, V.; Zimova, L.; Barvikova, K.; Macikova, L.; Barvik, I.; Vlachova, V. Human and Mouse TRPA1 Are Heat and Cold Sensors Differentially Tuned by Voltage. Cells 2019, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Jordt, S.-E.; Bautista, D.M.; Chuang, H.-H.; McKemy, D.D.; Zygmunt, P.M.; Högestätt, E.D.; Meng, I.D.; Julius, D. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature 2004, 427, 260–265. [Google Scholar] [CrossRef]
- Talavera, K.; Gees, M.; Karashima, Y.; Meseguer, V.M.; Vanoirbeek, J.A.J.; Damann, N.; Everaerts, W.; Benoit, M.; Janssens, A.; Vennekens, R.; et al. Nicotine activates the chemosensory cation channel TRPA1. Nat. Neurosci. 2009, 12, 1293–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meseguer, V.; Alpizar, Y.A.; Luis, E.; Tajada, S.; Denlinger, B.; Fajardo, O.; Manenschijn, J.A.; Fernández-Peña, C.; Talavera, A.; Kichko, T.; et al. TRPA1 channels mediate acute neurogenic inflammation and pain produced by bacterial endotoxins. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, D.A.; Gentry, C.; Moss, S.; Bevan, S. Transient receptor potential A1 is a sensory receptor for multiple products of oxidative stress. J. Neurosci. 2008, 28, 2485–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevisan, G.; Hoffmeister, C.; Rossato, M.F.; Oliveira, S.M.; Silva, M.A.; Silva, C.R.; Fusi, C.; Tonello, R.; Minocci, D.; Guerra, G.P.; et al. TRPA1 receptor stimulation by hydrogen peroxide is critical to trigger hyperalgesia and inflammation in a model of acute gout. Free Radic. Biol. Med. 2014, 72, 200–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenas, O.M.; Zaharieva, E.E.; Para, A.; Vásquez-Doorman, C.; Petersen, C.P.; Gallio, M. Activation of planarian TRPA1 by reactive oxygen species reveals a conserved mechanism for animal nociception. Nat. Neurosci. 2017, 20, 1686–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viana, F. TRPA1 channels: Molecular sentinels of cellular stress and tissue damage. J. Physiol. 2016, 594, 4151–4169. [Google Scholar] [CrossRef] [Green Version]
- Vriens, J.; Owsianik, G.; Hofmann, T.; Philipp, S.E.; Stab, J.; Chen, X.; Benoit, M.; Xue, F.; Janssens, A.; Kerselaers, S.; et al. TRPM3 Is a Nociceptor Channel Involved in the Detection of Noxious Heat. Neuron 2011, 70, 482–494. [Google Scholar] [CrossRef] [Green Version]
- Held, K.; Kichko, T.; De Clercq, K.; Klaassen, H.; Van Bree, R.; Vanherck, J.C.; Marchand, A.; Reeh, P.W.; Chaltin, P.; Voets, T.; et al. Activation of TRPM3 by a potent synthetic ligand reveals a role in peptide release. Proc. Natl. Acad. Sci. USA 2015, 112, E1363–E1372. [Google Scholar] [CrossRef] [Green Version]
- Straub, I.; Krügel, U.; Mohr, F.; Teichert, J.; Rizun, O.; Konrad, M.; Oberwinkler, J.; Schaefer, M. Flavanones that selectively inhibit TRPM3 attenuate thermal nociception in vivo. Mol. Pharmacol. 2013, 84, 736–750. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.-H.; McNaughton, P.A. The TRPM2 ion channel is required for sensitivity to warmth. Nature 2016, 536, 460–463. [Google Scholar] [CrossRef] [Green Version]
- Vilar, B.; Tan, C.-H.; McNaughton, P.A. Heat detection by the TRPM2 ion channel. Nature 2020, 584, E5–E12. [Google Scholar] [CrossRef] [PubMed]
- Mulier, M.; Vandewauw, I.; Vriens, J.; Voets, T. Reply to: Heat detection by the TRPM2 ion channel. Nature 2020, 584, E13–E15. [Google Scholar] [CrossRef] [PubMed]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S.; et al. A TRP channel that senses cold stimuli and menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Yang, W.; Jiang, H.; Guo, C.; Huang, A.J.W.; Hu, H.; Liu, Q. TRPV1 activity and substance P release are required for corneal cold nociception. Nat. Commun. 2019, 10, 5678. [Google Scholar] [CrossRef]
- Bautista, D.M.; Siemens, J.; Glazer, J.M.; Tsuruda, P.R.; Basbaum, A.I.; Stucky, C.L.; Jordt, S.E.; Julius, D. The menthol receptor TRPM8 is the principal detector of environmental cold. Nature 2007, 448, 204–208. [Google Scholar] [CrossRef]
- Colburn, R.W.; Lubin, M.L.; Stone, D.J.; Wang, Y.; Lawrence, D.; D’Andrea, M.R.R.; Brandt, M.R.; Liu, Y.; Flores, C.M.; Qin, N. Attenuated Cold Sensitivity in TRPM8 Null Mice. Neuron 2007, 54, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Dhaka, A.; Murray, A.N.; Mathur, J.; Earley, T.J.; Petrus, M.J.; Patapoutian, A. TRPM8 Is Required for Cold Sensation in Mice. Neuron 2007, 54, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Belmonte, C.; Nichols, J.J.; Cox, S.M.; Brock, J.A.; Begley, C.G.; Bereiter, D.A.; Dartt, D.A.; Galor, A.; Hamrah, P.; Ivanusic, J.J.; et al. TFOS DEWS II pain and sensation report. Ocul. Surf. 2017, 15, 404–437. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, A.; Bai, L.; Ginty, D.D. The gentle touch receptors of mammalian skin. Science 2014, 346, 950–954. [Google Scholar] [CrossRef] [Green Version]
- Laing, R.J.; Dhaka, A. ThermoTRPs and Pain. Neuroscientist 2016, 22, 171–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Tang, Z.; Surdenikova, L.; Kim, S.; Patel, K.N.; Kim, A.; Ru, F.; Guan, Y.; Weng, H.J.; Geng, Y.; et al. Sensory Neuron-Specific GPCR Mrgprs Are Itch Receptors Mediating Chloroquine-Induced Pruritus. Cell 2009, 139, 1353–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, W.S.; Tak, M.H.; Lee, M.H.; Kim, M.; Kim, M.; Koo, J.Y.; Lee, C.H.; Kim, M.; Oh, U. TRPV1 mediates histamine-induced itching via the activation of phospholipase A2 and 12-lipoxygenase. J. Neurosci. 2007, 27, 2331–2337. [Google Scholar] [CrossRef]
- Wilson, S.R.; Gerhold, K.A.; Bifolck-Fisher, A.; Liu, Q.; Patel, K.N.; Dong, X.; Bautista, D.M. TRPA1 is required for histamine-independent, Mas-related G protein-coupled receptor-mediated itch. Nat. Neurosci. 2011, 14, 595–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Barry, D.M.; Liu, X.-Y.; Yin, S.; Munanairi, A.; Meng, Q.-T.; Cheng, W.; Mo, P.; Wan, L.; Liu, S.-B.; et al. Facilitation of TRPV4 by TRPV1 is required for itch transmission in some sensory neuron populations. Sci. Signal. 2016, 9, ra71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pogorzala, L.A.; Mishra, S.K.; Hoon, M.A. The cellular code for mammalian thermosensation. J. Neurosci. 2013, 33, 5533–5541. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, W.M.; Palkar, R.; Lippoldt, E.K.; McCoy, D.D.; Baluch, F.; Chen, J.; McKemy, D.D. A sensory-labeled line for cold: TRPM8-expressing sensory neurons define the cellular basis for cold, cold pain, and cooling-mediated analgesia. J. Neurosci. 2013, 33, 2837–2848. [Google Scholar] [CrossRef] [Green Version]
- Peier, A.M.; Reeve, A.J.; Andersson, D.A.; Moqrich, A.; Earley, T.J.; Hergarden, A.C.; Story, G.M.; Colley, S.; Hogenesch, J.B.; McIntyre, P.; et al. A heat-sensitive TRP channel expressed in keratinocytes. Science 2002, 296, 2046–2049. [Google Scholar] [CrossRef]
- Smith, G.D.; Gunthorpe, M.J.; Kelsell, R.E.; Hayes, P.D.; Reilly, P.; Facer, P.; Wright, J.E.; Jerman, J.C.; Walhin, J.P.; Ooi, L.; et al. TRPV3 is a temperature-sensitive vanilloid receptor-like protein. Nature 2002, 418, 186–190. [Google Scholar] [CrossRef]
- Watanabe, H.; Vriens, J.; Suh, S.H.; Benham, C.D.; Droogmans, G.; Nilius, B. Heat-evoked activation of TRPV4 channels in a HEK293 cell expression system and in native mouse aorta endothelial cells. J. Biol. Chem. 2002, 277, 47044–47051. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Ramsey, I.S.; Kotecha, S.A.; Moran, M.M.; Chong, J.A.; Lawson, D.; Ge, P.; Lilly, J.; Silos-Santiago, I.; Xie, Y.; et al. TRPV3 is a calcium-permeable temperature-sensitive cation channel. Nature 2002, 418, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Abraira, V.E.; Ginty, D.D. The sensory neurons of touch. Neuron 2013, 79, 618–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belmonte, C. Pain, Dryness, and Itch Sensations in Eye Surface Disorders Are Defined by a Balance between Inflammation and Sensory Nerve Injury. Cornea 2019, 38 (Suppl. S1), S11–S24. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Trillo, J.; Florez-Paz, D.; Íñigo-Portugués, A.; González-González, O.; Del Campo, A.G.; González, A.; Viana, F.; Belmonte, C.; Gomis, A. Piezo2 Mediates Low-Threshold Mechanically Evoked Pain in the Cornea. J. Neurosci. 2020, 40, 8976–8993. [Google Scholar] [CrossRef] [PubMed]
- Parra, A.; Gonzalez-Gonzalez, O.; Gallar, J.; Belmonte, C. Tear fluid hyperosmolality increases nerve impulse activity of cold thermoreceptor endings of the cornea. Pain 2014, 155, 1481–1491. [Google Scholar] [CrossRef] [Green Version]
- Belmonte, C.; Gallar, J. Cold thermoreceptors, unexpected players in tear production and ocular dryness sensations. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3888–3892. [Google Scholar] [CrossRef] [Green Version]
- Quallo, T.; Vastani, N.; Horridge, E.; Gentry, C.; Parra, A.; Moss, S.; Viana, F.; Belmonte, C.; Andersson, D.A.; Bevan, S. TRPM8 is a neuronal osmosensor that regulates eye blinking in mice. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Gallar, J.; Pozo, M.A.; Tuckett, R.P.; Belmonte, C. Response of sensory units with unmyelinated fibres to mechanical, thermal and chemical stimulation of the cat’s cornea. J. Physiol. 1993, 468, 609–622. [Google Scholar] [CrossRef]
- González-González, O.; Bech, F.; Gallar, J.; Merayo-Lloves, J.; Belmonte, C. Functional properties of sensory nerve terminals of the mouse cornea. Investig. Ophthalmol. Vis. Sci. 2017, 58, 404–415. [Google Scholar] [CrossRef] [Green Version]
- Alcalde, I.; Íñigo-Portugués, A.; González-González, O.; Almaraz, L.; Artime, E.; Morenilla-Palao, C.; Gallar, J.; Viana, F.; Merayo-Lloves, J.; Belmonte, C. Morphological and functional changes in TRPM8-expressing corneal cold thermoreceptor neurons during aging and their impact on tearing in mice. J. Comp. Neurol. 2018, 526, 1859–1874. [Google Scholar] [CrossRef]
- Edwards, I.R.; Aronson, J.K. Adverse drug reactions: Definitions, diagnosis, and management. Lancet 2000, 356, 1255–1259. [Google Scholar] [CrossRef]
- Hüll, K.; Morstein, J.; Trauner, D. In Vivo Photopharmacology. Chem. Rev. 2018, 118, 10710–10747. [Google Scholar] [CrossRef] [PubMed]
- Velema, W.A.; Szymanski, W.; Feringa, B.L. Photopharmacology: Beyond proof of principle. J. Am. Chem. Soc. 2014, 136, 2178–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerch, M.M.; Hansen, M.J.; van Dam, G.M.; Szymanski, W.; Feringa, B.L. Emerging Targets in Photopharmacology. Angew. Chem. Int. Ed. 2016, 55, 10978–10999. [Google Scholar] [CrossRef] [Green Version]
- Fehrentz, T.; Schönberger, M.; Trauner, D. Optochemical genetics. Angew. Chem. Int. Ed. 2011, 50, 12156–12182. [Google Scholar] [CrossRef]
- Housley, G.D.; Raybould, N.P.; Thorne, P.R. Fluorescence imaging of Na+ influx via P2X receptors in cochlear hair cells. Hear. Res. 1998, 119, 1–13. [Google Scholar] [CrossRef]
- Ellis-Davies, G.C.R. Caged compounds: Photorelease technology for control of cellular chemistry and physiology. Nat. Methods 2007, 4, 619–628. [Google Scholar] [CrossRef] [Green Version]
- Kötter, R.; Schubert, D.; Dyhrfjeld-Johnsen, J.; Luhmann, H.J.; Staiger, J.F. Optical release of caged glutamate for stimulation of neurons in the in vitro slice preparation. J. Biomed. Opt. 2005, 10, 011003. [Google Scholar] [CrossRef]
- Verde, E.M.R.; Zayat, L.; Etchenique, R.; Yuste, R. Photorelease of GABA with visible light using an inorganic caging group. Front. Neural Circuits 2008, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Banghart, M.R.; Sabatini, B.L. Photoactivatable neuropeptides for spatiotemporally precise delivery of opioids in neural tissue. Neuron 2012, 73, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Mourot, A.; Kienzler, M.A.; Banghart, M.R.; Fehrentz, T.; Huber, F.M.E.; Stein, M.; Kramer, R.H.; Trauner, D. Tuning photochromic ion channel blockers. ACS Chem. Neurosci. 2011, 2, 536–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tochitsky, I.; Helft, Z.; Meseguer, V.; Fletcher, R.B.; Vessey, K.A.; Telias, M.; Denlinger, B.; Malis, J.; Fletcher, E.L.; Kramer, R.H. How Azobenzene Photoswitches Restore Visual Responses to the Blind Retina. Neuron 2016, 92, 100–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berlin, S.; Isacoff, E.Y. Synapses in the spotlight with synthetic optogenetics. EMBO Rep. 2017, 18, 677–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiner, A.; Levitz, J.; Isacoff, E.Y. Controlling ionotropic and metabotropic glutamate receptors with light: Principles and potential. Curr. Opin. Pharmacol. 2015, 20, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Broichhagen, J.; Trauner, D. The in vivo chemistry of photoswitched tethered ligands. Curr. Opin. Chem. Biol. 2014, 21, 121–127. [Google Scholar] [CrossRef]
- Stein, M.; Breit, A.; Fehrentz, T.; Gudermann, T.; Trauner, D. Optical control of TRPV1 channels. Angew. Chem. Int. Ed. 2013, 52, 9845–9848. [Google Scholar] [CrossRef]
- Frank, J.A.; Moroni, M.; Moshourab, R.; Sumser, M.; Lewin, G.R.; Trauner, D. Photoswitchable fatty acids enable optical control of TRPV1. Nat. Commun. 2015, 6, 7118. [Google Scholar] [CrossRef] [Green Version]
- Curcic, S.; Tiapko, O.; Groschner, K. Photopharmacology and opto-chemogenetics of TRPC channels-some therapeutic visions. Pharmacol. Ther. 2019, 200, 13–26. [Google Scholar] [CrossRef]
- Zemelman, B.V.; Nesnas, N.; Lee, G.A.; Miesenböck, G. Photochemical gating of heterologous ion channels: Remote control over genetically designated populations of neurons. Proc. Natl. Acad. Sci. USA 2003, 100, 1352–1357. [Google Scholar] [CrossRef] [Green Version]
- Katritzky, A.R.; Xu, Y.J.; Vakulenko, A.V.; Wilcox, A.L.; Bley, K.R. Model Compounds of Caged Capsaicin: Design, Synthesis, and Photoreactivity. J. Org. Chem. 2003, 68, 9100–9104. [Google Scholar] [CrossRef]
- Carr, J.L.; Wease, K.N.; Van Ryssen, M.P.; Paterson, S.; Agate, B.; Gallagher, K.A.; Brown, C.T.A.; Scott, R.H.; Conway, S.J. In vitro photo-release of a TRPV1 agonist. Bioorg. Med. Chem. Lett. 2006, 16, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, D.; Funk, K.; Dekowski, B.; Lechler, R.; Keller, S.; Möhrlen, F.; Frings, S.; Hagen, V. Caged capsaicins: New tools for the examination of TRPV1 channels in somatosensory neurons. ChemBioChem 2007, 8, 89–97. [Google Scholar] [CrossRef]
- Mourot, A.; Tochitsky, I.; Kramer, R.H. Light at the end of the channel: Optical manipulation of intrinsic neuronal excitability with chemical photoswitches. Front. Mol. Neurosci. 2013, 6, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourot, A.; Fehrentz, T.; Le Feuvre, Y.; Smith, C.M.; Herold, C.; Dalkara, D.; Nagy, F.; Trauner, D.; Kramer, R.H. Rapid optical control of nociception with an ion-channel photoswitch. Nat. Methods 2012, 9, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Binshtok, A.M.; Bean, B.P.; Woolf, C.J. Inhibition of nociceptors by TRPV1-mediated entry of impermeant sodium channel blockers. Nature 2007, 449, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Strichartz, G.R. The inhibition of sodium currents in myelinated nerve by quaternary derivatives of lidocaine. J. Gen. Physiol. 1973, 62, 37–57. [Google Scholar] [CrossRef] [PubMed]
- Morstein, J.; Hill, R.Z.; Novak, A.J.E.; Feng, S.; Norman, D.D.; Donthamsetti, P.C.; Frank, J.A.; Harayama, T.; Williams, B.M.; Parrill, A.L.; et al. Optical control of sphingosine-1-phosphate formation and function. Nat. Chem. Biol. 2019, 15, 623–631. [Google Scholar] [CrossRef]
- Chung, M.-K.; Güler, A.D.; Caterina, M.J. TRPV1 shows dynamic ionic selectivity during agonist stimulation. Nat. Neurosci. 2008, 11, 555–564. [Google Scholar] [CrossRef]
- Mourot, A.; Herold, C.; Kienzler, M.A.; Kramer, R.H. Understanding and improving photo-control of ion channels in nociceptors with azobenzene photo-switches. Br. J. Pharmacol. 2018, 175, 2296–2311. [Google Scholar] [CrossRef]
- Dong, M.; Babalhavaeji, A.; Samanta, S.; Beharry, A.A.; Woolley, G.A. Red-Shifting Azobenzene Photoswitches for in Vivo Use. Acc. Chem. Res. 2015, 48, 2662–2670. [Google Scholar] [CrossRef]
- Dong, M.; Babalhavaeji, A.; Hansen, M.J.; Kálmán, L.; Woolley, G.A. Red, far-red, and near infrared photoswitches based on azonium ions. Chem. Commun. 2015, 51, 12981–12984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokel, D.; Cheung, C.Y.J.; Mills, R.; Coutinho-Budd, J.; Huang, L.; Setola, V.; Sprague, J.; Jin, S.; Jin, Y.N.; Huang, X.P.; et al. Photochemical activation of TRPA1 channels in neurons and animals. Nat. Chem. Biol. 2013, 9, 257–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, P.Y.; Thawani, A.R.; Balderas, E.; White, A.J.P.; Chaudhuri, D.; Fuchter, M.J.; Peterson, R.T. TRPswitch—A Step-Function Chemo-optogenetic Ligand for the Vertebrate TRPA1 Channel. J. Am. Chem. Soc. 2020, 142, 17457–17468. [Google Scholar] [CrossRef] [PubMed]
- Julius, D. TRP channels and pain. Annu. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef] [Green Version]
- Kuffler, S.W.; Eyzaguirre, C. Synaptic inhibition in an isolated nerve cell. J. Gen. Physiol. 1955, 39, 155–184. [Google Scholar] [CrossRef] [Green Version]
- Loewenstein, W.R. Excitation and inactivation in a receptor membrane. Ann. N. Y. Acad. Sci. 1961, 94, 510–534. [Google Scholar] [CrossRef]
- Reeh, P.W. Sensory receptors in mammalian skin in an in vitro preparation. Neurosci. Lett. 1986, 66, 141–146. [Google Scholar] [CrossRef]
- Koerber, H.R.; Woodbury, C.J. Comprehensive phenotyping of sensory neurons using an ex vivo somatosensory system. Physiol. Behav. 2002, 77, 589–594. [Google Scholar] [CrossRef]
- Baumbauer, K.M.; Deberry, J.J.; Adelman, P.C.; Miller, R.H.; Hachisuka, J.; Lee, K.H.; Ross, S.E.; Koerber, H.R.; Davis, B.M.; Albers, K.M. Keratinocytes can modulate and directly initiate nociceptive responses. Elife 2015, 4, e09674. [Google Scholar] [CrossRef]
- Ritter-Jones, M.; Najjar, S.; Albers, K.M. Keratinocytes as modulators of sensory afferent firing. Pain 2016, 157, 786–787. [Google Scholar] [CrossRef]
- Israel, M.R.; Tanaka, B.S.; Castro, J.; Thongyoo, P.; Robinson, S.D.; Zhao, P.; Deuis, J.R.; Craik, D.J.; Durek, T.; Brierley, S.M.; et al. NaV1.6 regulates excitability of mechanosensitive sensory neurons. J. Physiol. 2019, 597, 3751–3768. [Google Scholar] [CrossRef] [PubMed]
- Loeza-Alcocer, E.; McPherson, T.P.; Gold, M.S. Peripheral GABA receptors regulate colonic afferent excitability and visceral nociception. J. Physiol. 2019, 597, 3425–3439. [Google Scholar] [CrossRef] [PubMed]
- Talagas, M.; Lebonvallet, N.; Berthod, F.; Misery, L. Cutaneous nociception: Role of keratinocytes. Exp. Dermatol. 2019, 28, 1466–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkai, O.; Goldstein, R.H.; Caspi, Y.; Katz, B.; Lev, S.; Binshtok, A.M. The role of Kv7/M potassium channels in controlling ectopic firing in nociceptors. Front. Mol. Neurosci. 2017, 10, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, M. Molecular Biology of Sensory Transduction. In Wall & Melzack’s Textbook of Pain, 6th ed.; McMahon, S.B., Koltzenburg, M., Tracey, I., Turk, D., Eds.; Elsevier Health Sciences: Philadelphia, PA, USA, 2013; ISBN 9780702053740. [Google Scholar]
- Brock, J.A.; McLachlan, E.M.; Belmonte, C. Tetrodotoxin-resistant impulses in single nociceptor nerve terminals in guinea-pig cornea. J. Physiol. 1998, 512, 211–217. [Google Scholar] [CrossRef]
- Brock, J.A.; Pianova, S.; Belmonte, C. Differences between nerve terminal impulses of polymodal nociceptors and cold sensory receptors of the guinea-pig cornea. J. Physiol. 2001, 533, 493–501. [Google Scholar] [CrossRef]
- Vasylyev, D.V.; Waxman, S.G. Membrane properties and electrogenesis in the distal axons of small dorsal root ganglion neurons in vitro. J. Neurophysiol. 2012, 108, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Belmonte, C.; Acosta, M.C.; Gallar, J. Neural basis of sensation in intact and injured corneas. Exp. Eye Res. 2004, 78, 513–525. [Google Scholar] [CrossRef]
- Strassman, A.M.; Levy, D. Response properties of dural nociceptors in relation to headache. J. Neurophysiol. 2006, 95, 1298–1306. [Google Scholar] [CrossRef]
- Baillie, L.D.; Ahn, A.H.; Mulligan, S.J. Sumatriptan inhibition of N-type calcium channel mediated signaling in dural CGRP terminal fibres. Neuropharmacology 2012, 63, 362–367. [Google Scholar] [CrossRef] [Green Version]
- Baillie, L.D.; Schmidhammer, H.; Mulligan, S.J. Peripheral μ-opioid receptor mediated inhibition of calcium signaling and action potential-evoked calcium fluorescent transients in primary afferent CGRP nociceptive terminals. Neuropharmacology 2015, 93, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, C.; Aracil, A.; Acosta, M.C.; Luna, C.; Gallar, J. Nerves and sensations from the eye surface. Ocul. Surf. 2004, 2, 248–253. [Google Scholar] [CrossRef]
- Ivanusic, J.J.; Wood, R.J.; Brock, J.A. Sensory and sympathetic innervation of the mouse and guinea pig corneal epithelium. J. Comp. Neurol. 2013, 521, 877–893. [Google Scholar] [CrossRef]
- Gover, T.D.; Kao, J.P.Y.; Weinreich, D. Calcium signaling in single peripheral sensory nerve terminals. J. Neurosci. 2003, 23, 4793–4797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleidervish, I.A.; Lasser-Ross, N.; Gutnick, M.J.; Ross, W.N. Na+ imaging reveals little difference in action potential-evoked Na+ influx between axon and soma. Nat. Neurosci. 2010, 13, 852–860. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleixandre-Carrera, F.; Engelmayer, N.; Ares-Suárez, D.; Acosta, M.d.C.; Belmonte, C.; Gallar, J.; Meseguer, V.; Binshtok, A.M. Optical Assessment of Nociceptive TRP Channel Function at the Peripheral Nerve Terminal. Int. J. Mol. Sci. 2021, 22, 481. https://doi.org/10.3390/ijms22020481
Aleixandre-Carrera F, Engelmayer N, Ares-Suárez D, Acosta MdC, Belmonte C, Gallar J, Meseguer V, Binshtok AM. Optical Assessment of Nociceptive TRP Channel Function at the Peripheral Nerve Terminal. International Journal of Molecular Sciences. 2021; 22(2):481. https://doi.org/10.3390/ijms22020481
Chicago/Turabian StyleAleixandre-Carrera, Fernando, Nurit Engelmayer, David Ares-Suárez, María del Carmen Acosta, Carlos Belmonte, Juana Gallar, Víctor Meseguer, and Alexander M. Binshtok. 2021. "Optical Assessment of Nociceptive TRP Channel Function at the Peripheral Nerve Terminal" International Journal of Molecular Sciences 22, no. 2: 481. https://doi.org/10.3390/ijms22020481