DAP3 Is Involved in Modulation of Cellular Radiation Response by RIG-I-Like Receptor Agonist in Human Lung Adenocarcinoma Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
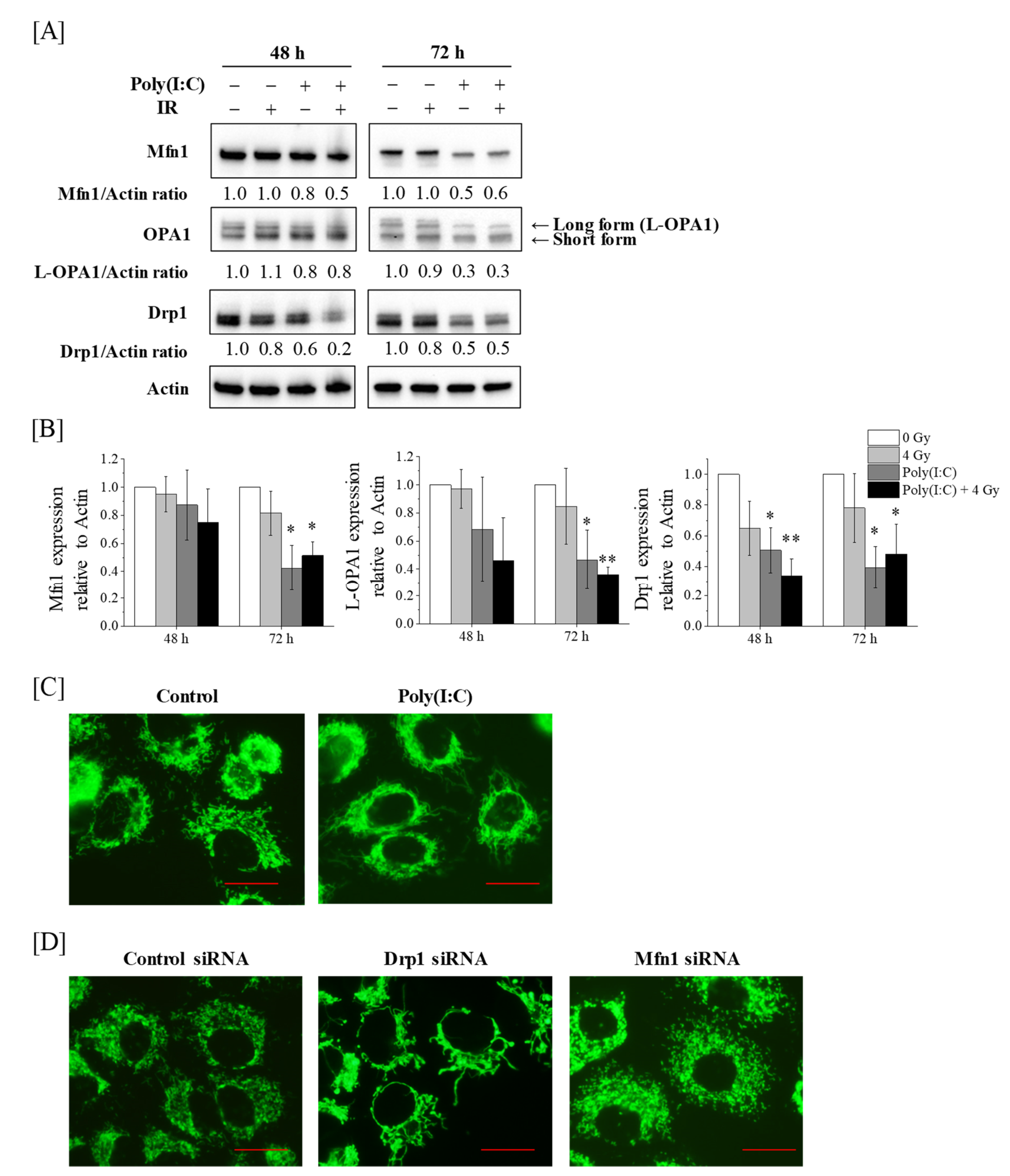
2.1. Expression of Mitochondrial Dynamics-Related Proteins and Mitochondrial Morphology in A549 Cells Treated with Poly(I:C) and/or IR
2.2. Effect of Drp1-Knockdown on IR-Induced Cell Death in A549 Cells
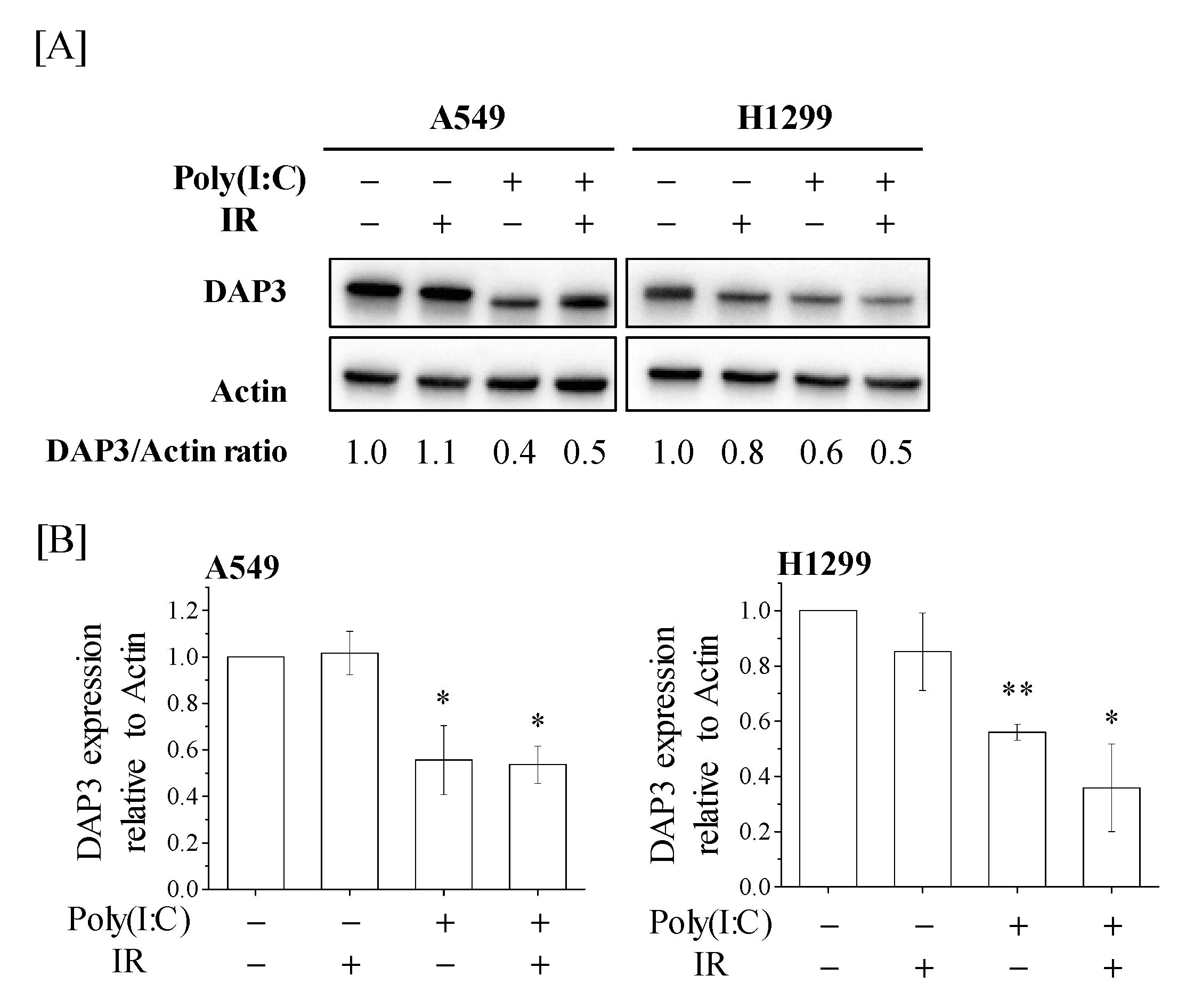
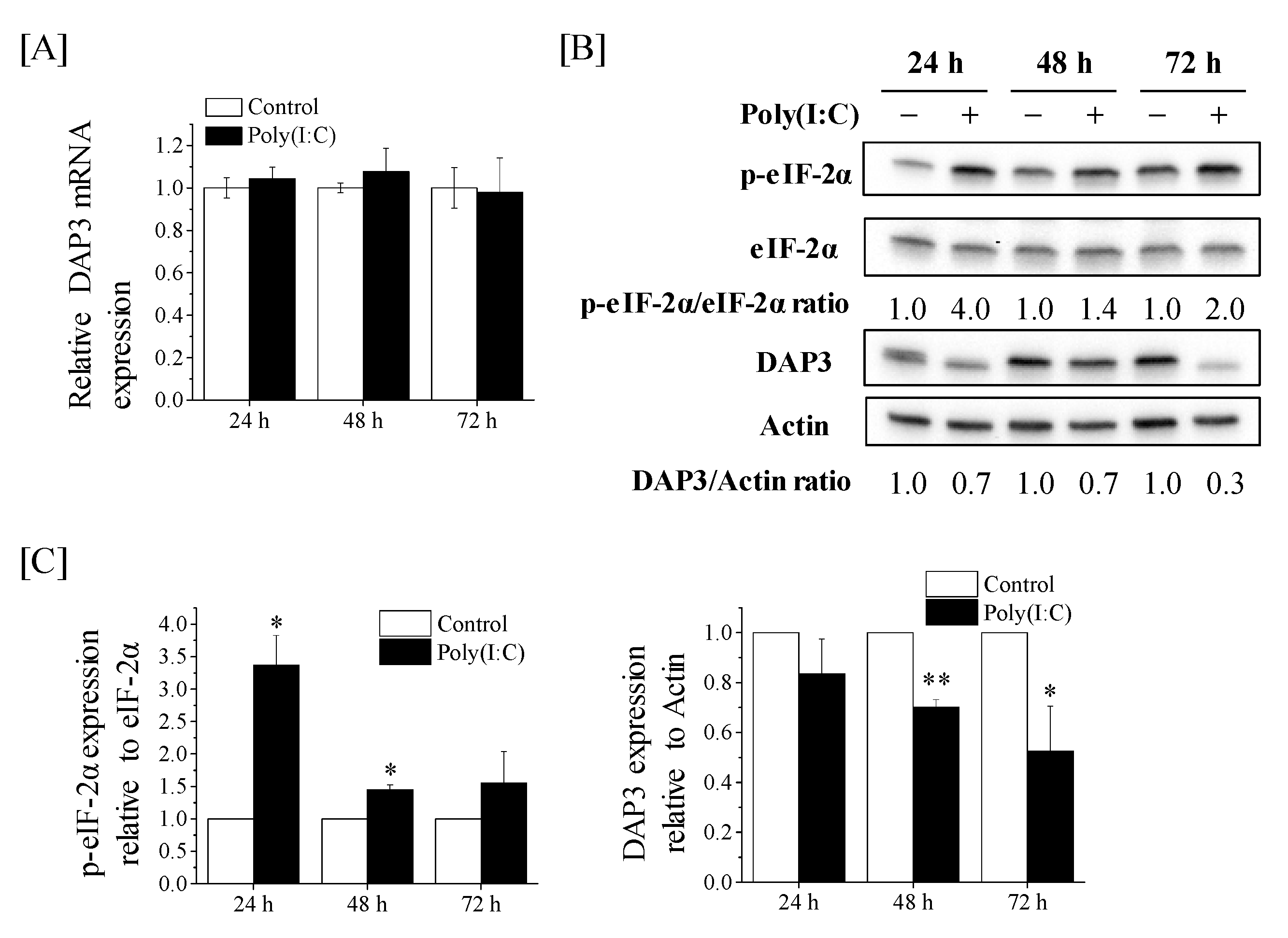
2.3. Downregulation of DAP3 Protein Expression by Poly(I:C) in Human Lung Adenocarcinoma Cells
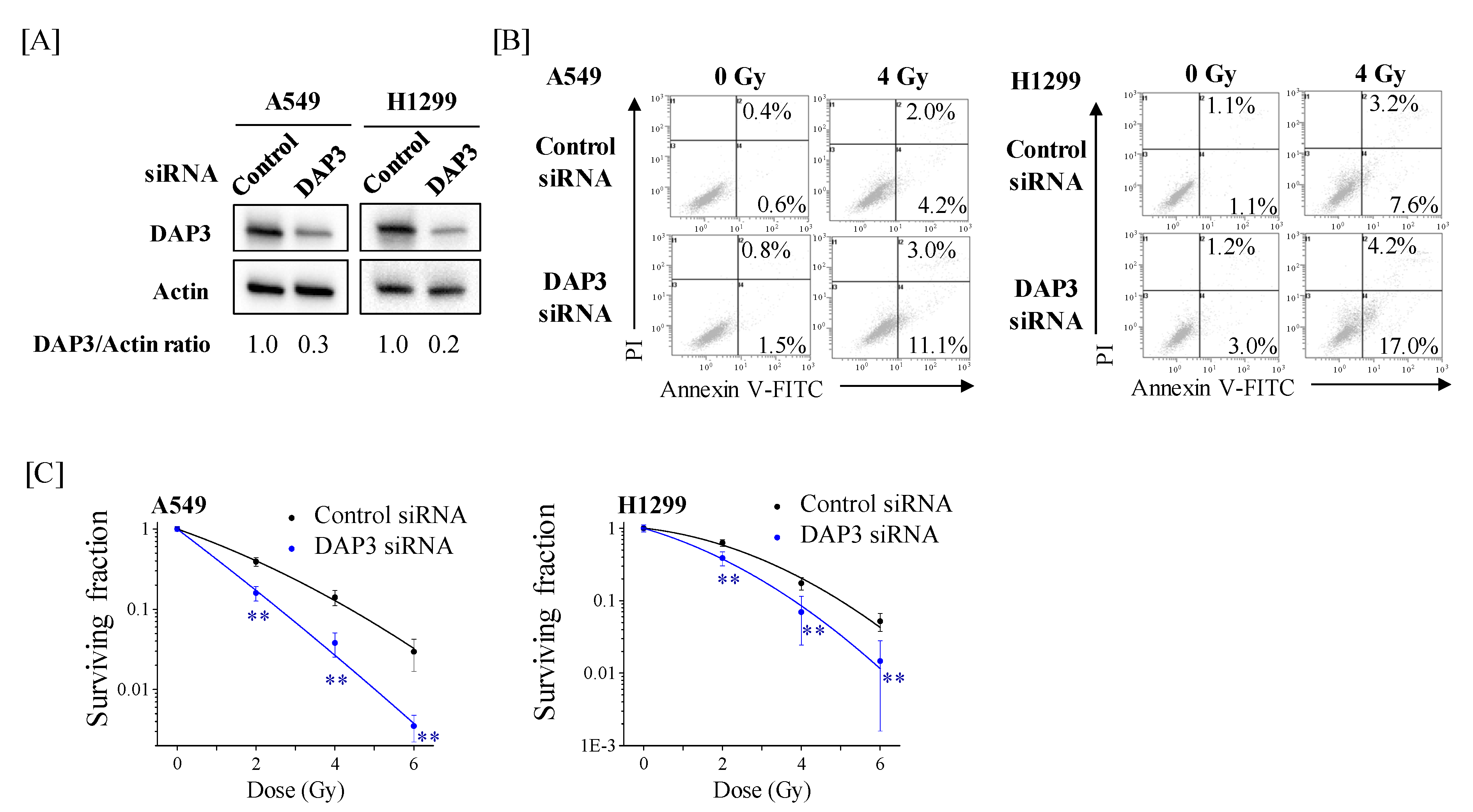
2.4. Involvement of DAP3 in Radioresistance of Human Lung Adenocarcinoma Cells
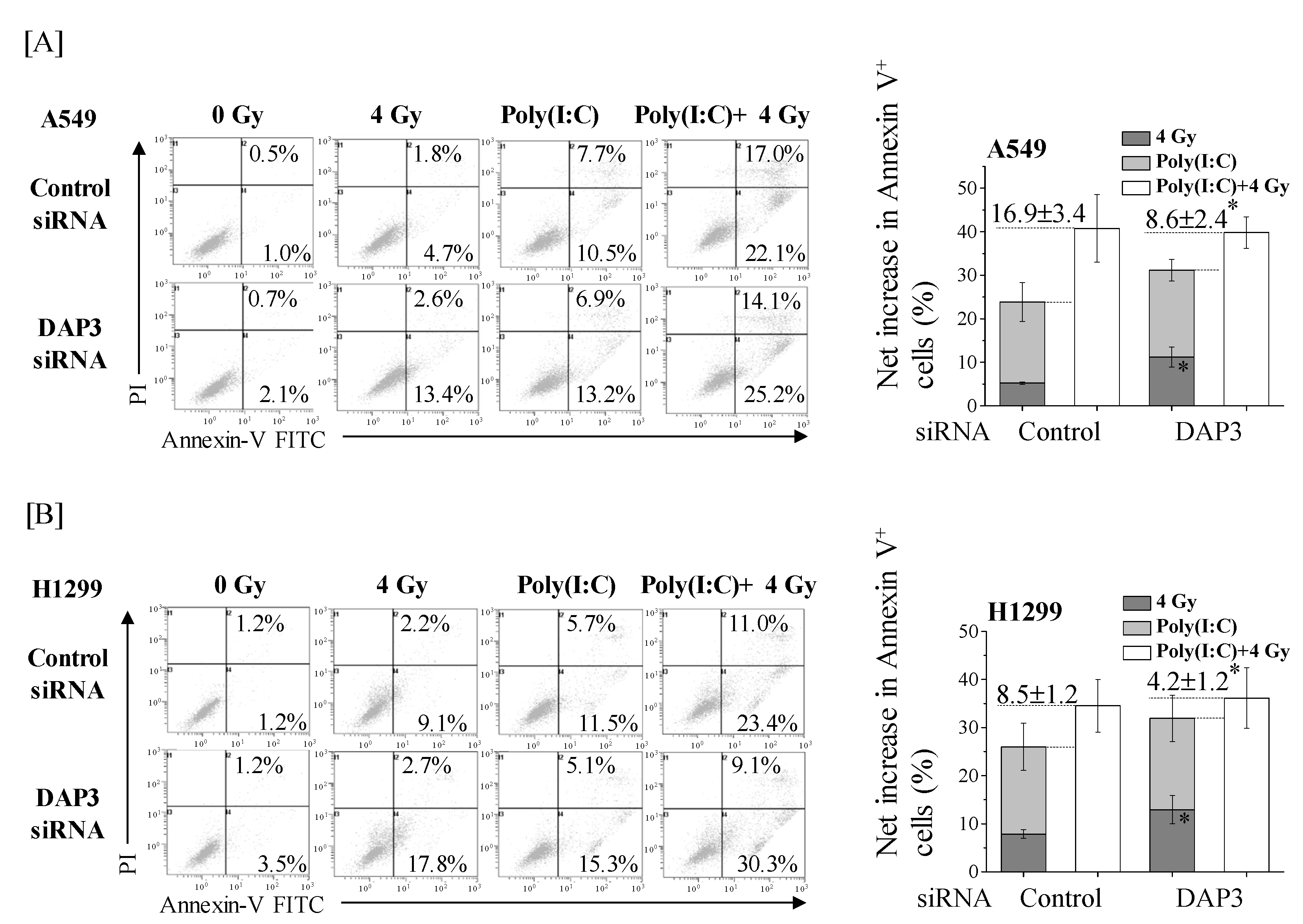
2.5. Involvement of DAP3 in the More-Than-Additive Effect of Cotreatment with Poly(I:C) and IR on Cell Death in Human Lung Adenocarcinoma Cells
2.6. Post-Transcriptional Downregulation of DAP3 Expression by Poly(I:C) in A549 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture and Treatment
4.3. In Vitro X-ray Irradiation
4.4. SDS-PAGE and Western Blotting
4.5. Mitochondrial Morphology
4.6. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.7. siRNA Transfection
4.8. Detection of Cell Death
4.9. Clonogenic Survival Assay
4.10. Statistical Analysis
4.11. URLs
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DAP3 | Death-associated protein 3 |
| Drp1 | Dynamin-related protein 1 |
| eIF-2α | eukaryotic initiation factor-2α |
| HRP | Horseradish peroxidase |
| IFNs | Interferons |
| IR | Ionizing radiation |
| L-OPA1 | long isoform OPA1 |
| mtDNA | Mitochondrial DNA |
| Mfn1/2 | Mitofusin-1/2 |
| OPA1 | Optic atrophy 1 |
| p-eIF-2α | Phosphorylated eIF-2α |
| Poly(I:C) | Poly(I:C)-HMW/LyoVec™ |
| PI | Propidium iodide |
| qRT-PCR | Quantitative reverse transcription polymerase chain reaction |
| RIG-I | Retinoic acid-inducible gene-I |
| RLRs | RIG-I-like receptors |
References
- Suen, D.F.; Norris, K.L.; Youle, R.J. Mitochondrial dynamics and apoptosis. Genes Dev. 2008, 22, 1577–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angajala, A.; Lim, S.; Phillips, J.B.; Kim, J.H.; Yates, C.; You, Z.; Tan, M. Diverse roles of mitochondria in immune responses: Novel insights into immuno-metabolism. Front. Immunol. 2018, 9, 1605. [Google Scholar] [CrossRef] [PubMed]
- Senft, D.; Ronai, Z.A. Regulators of mitochondrial dynamics in cancer. Curr. Opin. Cell Biol. 2016, 39, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, P.; Chan, D.C. Mitochondrial dynamics and inheritance during cell division, development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 634–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanier, C.; Garcin, D.; Vazquez, A.; Arnoult, D. Mitochondrial dynamics regulate the RIG-I-like receptor antiviral pathway. EMBO Rep. 2010, 11, 133–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobashigawa, S.; Suzuki, K.; Yamashita, S. Ionizing radiation accelerates Drp1-dependent mitochondrial fission, which involves delayed mitochondrial reactive oxygen species production in normal human fibroblast-like cells. Biochem. Biophys. Res. Commun. 2011, 414, 795–800. [Google Scholar] [CrossRef]
- Kwon, S.M.; Lee, Y.K.; Min, S.; Woo, H.G.; Wang, H.J.; Yoon, G. Mitoribosome defect in hepatocellular carcinoma promotes an aggressive phenotype with suppressed immune reaction. iScience 2020, 23, 101247. [Google Scholar] [CrossRef]
- Yoneyama, M.; Fujita, T. Structural mechanism of RNA recognition by the RIG-I-like receptors. Immunity 2008, 29, 178–181. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Takahashi, K.; Sato, S.; Coban, C.; Kumar, H.; Kato, H.; Ishii, K.J.; Takeuchi, O.; Akira, S. IPS-1, an adaptor triggering RIG-I- and Mda5-mediated type I interferon induction. Nat. Immunol. 2005, 6, 981–988. [Google Scholar] [CrossRef]
- Besch, R.; Poeck, H.; Hohenauer, T.; Senft, D.; Häcker, G.; Berking, C.; Hornung, V.; Endres, S.; Ruzicka, T.; Rothenfusser, S.; et al. Proapoptotic signaling induced by RIG-I and MDA-5 results in type I interferon-independent apoptosis in human melanoma cells. J. Clin. Investig. 2009, 119, 2399–2411. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wu, X.; Wu, L.; Wang, X.; Liu, Z. The anticancer functions of RIG-I–like receptors, RIG-I and MDA5, and their applications in cancer therapy. Transl. Res. 2017, 190, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Xia, M.; Meng, G.; Xu, C.; Song, Y.; Wei, J. Anti-angiogenic efficacy of 5’-triphosphate siRNA combining VEGF silencing and RIG-I activation in NSCLCs. Oncotarget 2015, 6, 29664–29674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poeck, H.; Besch, R.; Maihoefer, C.; Renn, M.; Tormo, D.; Morskaya, S.S.; Kirschnek, S.; Gaffal, E.; Landsberg, J.; Hellmuth, J.; et al. 5’-triphosphate-siRNA: Turning gene silencing and Rig-I activation against melanoma. Nat. Med. 2008, 14, 1256–1263. [Google Scholar] [CrossRef]
- Li, D.; Gale, R.P.; Liu, Y.; Lei, B.; Wang, Y.; Diao, D.; Zhang, M. 5′-Triphosphate siRNA targeting MDR1 reverses multi-drug resistance and activates RIG-I-induced immune-stimulatory and apoptotic effects against human myeloid leukaemia cells. Leuk. Res. 2017, 58, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, H.; Iwabuchi, M.; Kazama, Y.; Furukawa, M.; Kashiwakura, I. Effects of retinoic acid-inducible gene-I-like receptors activations and ionizing radiation cotreatment on cytotoxicity against human non-small cell lung cancer in vitro. Oncol. Lett. 2018, 15, 4697–4705. [Google Scholar] [CrossRef] [Green Version]
- Bo, T.; Yamamori, T.; Yamamoto, K.; Fujimoto, M.; Yasui, H.; Inanami, O. Mitochondrial fission promotes radiation-induced increase in intracellular Ca2+ level leading to mitotic catastrophe in mouse breast cancer EMT6 cells. Biochem. Biophys. Res. Commun. 2020, 522, 144–150. [Google Scholar] [CrossRef]
- Kim, S.J.; Syed, G.H.; Khan, M.; Chiu, W.W.; Sohail, M.A.; Gish, R.G.; Siddiqui, A. Hepatitis C virus triggers mitochondrial fission and attenuates apoptosis to promote viral persistence. Proc. Natl. Acad. Sci. USA 2014, 111, 6413–6418. [Google Scholar] [CrossRef] [Green Version]
- Kissil, J.L.; Cohen, O.; Raveh, T.; Kimchi, A. Structure-function analysis of an evolutionary conserved protein, DAP3, which mediates TNF-alpha- and Fas-induced cell death. EMBO J. 1999, 18, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Henning, K.A. The Molecular Genetics of Human Diseases with Defective DNA Damaging Processing. Ph.D. Thesis, Stanford University, Palo Alto, CA, USA, 1992. [Google Scholar]
- Ishihara, N.; Fujita, Y.; Oka, T.; Mihara, K. Regulation of Mitochondrial Morphology through Proteolytic Cleavage of OPA1. EMBO J. 2006, 25, 2966–2977. [Google Scholar] [CrossRef]
- García, M.A.; Meurs, E.F.; Esteban, M. The dsRNA protein kinase PKR: Virus and cell control. Biochimie 2007, 89, 799–811. [Google Scholar] [CrossRef]
- Elion, D.L.; Cook, R.S. Harnessing RIG-I and intrinsic immunity in the tumor microenvironment for therapeutic cancer treatment. Oncotarget 2018, 9, 29007–29017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monlun, M.; Hyernard, C.; Blanco, P.; Lartigue, L.; Faustin, B. Mitochondria as molecular platforms integrating multiple innate immune signalings. J. Mol. Biol. 2017, 429, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Ahn, D.G.; Syed, G.H.; Siddiqui, A. The essential role of mitochondrial dynamics in antiviral immunity. Mitochondrion 2018, 41, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Barbier, V.; Lang, D.; Valois, S.; Rothman, A.L.; Medin, C.L. Dengue virus induces mitochondrial elongation through impairment of Drp1-triggered mitochondrial fission. Virology 2017, 500, 149–160. [Google Scholar] [CrossRef]
- Fields, J.A.; Serger, E.; Campos, S.; Divakaruni, A.S.; Kim, C.; Smith, K.; Trejo, M.; Adame, A.; Spencer, B.; Rockenstein, E.; et al. HIV alters neuronal mitochondrial fission/fusion in the brain during HIV-associated neurocognitive disorders. Neurobiol. Dis. 2016, 86, 154–169. [Google Scholar] [CrossRef] [Green Version]
- Berg, R.K.; Melchjorsen, J.; Rintahaka, J.; Diget, E.; Søby, S.; Horan, K.A.; Gorelick, R.J.; Matikainen, S.; Larsen, C.S.; Ostergaard, L.; et al. Genomic HIV RNA Induces Innate Immune Responses through RIG-I-Dependent Sensing of Secondary-Structured RNA. PLoS ONE 2012, 7, e29291. [Google Scholar] [CrossRef] [Green Version]
- Sprokholt, J.K.; Kaptein, T.M.; van Hamme, J.L.; Overmars, R.J.; Gringhuis, S.I.; Geijtenbeek, T.B.H. RIG-I-like receptor activation by dengue virus drives follicular T helper cell formation and antibody production. PLoS Pathog. 2017, 13, e1006738. [Google Scholar] [CrossRef] [Green Version]
- Saita, S.; Ishihara, T.; Maeda, M.; Iemura, S.; Natsume, T.; Mihara, K.; Ishihara, N. Distinct types of protease systems are involved in homeostasis regulation of mitochondrial morphology via balanced fusion and fission. Genes Cells 2016, 21, 408–424. [Google Scholar] [CrossRef] [Green Version]
- Kobashigawa, S.; Kashino, G.; Suzuki, K.; Yamashita, S.; Mori, H. Ionizing radiation-induced cell death is partly caused by increase of mitochondrial reactive oxygen species in normal human fibroblast cells. Radiat. Res. 2015, 183, 455–464. [Google Scholar] [CrossRef]
- Xu, W.; Jing, L.; Wang, Q.; Lin, C.C.; Chen, X.; Diao, J.; Liu, Y.; Sun, X. Bax-PGAM5L-Drp1 complex is required for intrinsic apoptosis execution. Oncotarget 2015, 6, 30017–30034. [Google Scholar] [CrossRef] [Green Version]
- Rong, R.; Xia, X.; Peng, H.; Li, H.; You, M.; Liang, Z.; Yao, F.; Yao, X.; Xiong, K.; Huang, J.; et al. Cdk5-mediated Drp1 phosphorylation drives mitochondrial defects and neuronal apoptosis in radiation-induced optic neuropathy. Cell Death Dis. 2020, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Inoue-Yamauchi, A.; Oda, H. Depletion of mitochondrial fission factor DRP1 causes increased apoptosis in human colon cancer cells. Biochem. Biophys. Res. Commun. 2012, 421, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.Y.; Chow, K.C.; Chiao, M.T.; Yang, Y.C.; Shen, C.C. Reducing expression of dynamin-related protein 1 increases radiation sensitivity of glioblastoma cells. BioRxiv 2019, 688861. [Google Scholar] [CrossRef]
- Akita, M.; Suzuki-Karasaki, M.; Fujiwara, K.; Nakagawa, C.; Soma, M.; Yoshida, Y.; Ochiai, T.; Tokuhashi, Y.; Suzuki-Karasaki, Y. Mitochondrial division inhibitor-1 induces mitochondrial hyperfusion and sensitizes human cancer cells to TRAIL-induced apoptosis. Int. J. Oncol. 2014, 45, 1901–1912. [Google Scholar] [CrossRef]
- Kissil, J.L.; Deiss, L.P.; Bayewitch, M.; Raveh, T.; Khaspekov, G.; Kimchi, A. Isolation of DAP3, a novel mediator of interferon-gamma-induced cell death. J. Biol. Chem. 1995, 270, 27932–27936. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, T.; Shen, M.; Fujikura, D.; Tosa, N.; Kim, H.R.; Kon, S.; Uede, T.; Reed, J.C. Functional role of death-associated protein 3 (DAP3) in anoikis. J. Biol. Chem. 2004, 279, 44667–44672. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, T.; Reed, J.C. A GTP-binding adapter protein couples TRAIL receptors to apoptosis-inducing proteins. Nat. Immunol. 2001, 2, 493–500. [Google Scholar] [CrossRef]
- Xiao, L.; Xian, H.; Lee, K.Y.; Xiao, B.; Wang, H.; Yu, F.; Shen, H.M.; Liou, Y.C. Death-associated protein 3 regulates mitochondrial-encoded protein synthesis and mitochondrial dynamics. J. Biol. Chem. 2015, 290, 24961–24974. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.X.; Li, Q.; Li, P.F. Apoptosis Repressor with Caspase Recruitment Domain Contributes to Chemotherapy Resistance by Abolishing Mitochondrial Fission Mediated by Dynamin-Related Protein-1. Cancer Res. 2009, 69, 492–500. [Google Scholar] [CrossRef] [Green Version]
- Landes, T.; Martinou, J.C. Mitochondrial outer membrane permeabilization during apoptosis: The role of mitochondrial fission. Biochim. Biophys. Acta. 2011, 1813, 540–545. [Google Scholar] [CrossRef]
- Tang, L.; Wei, F.; Wu, Y.; He, Y.; Shi, L.; Xiong, F.; Gong, Z.; Guo, C.; Li, X.; Deng, H.; et al. Role of metabolism in cancer cell radioresistance and radiosensitization methods. J. Exp. Clin. Cancer Res. 2018, 37, 87. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hou, Q.; Xiao, G.; Yang, S.; Di, C.; Si, J.; Zhou, R.; Ye, Y.; Zhang, Y.; Zhang, H. Selective ATP hydrolysis inhibition in F1Fo ATP synthase enhances radiosensitivity in non-small-cell lung cancer cells (A549). Oncotarget 2017, 8, 53602–53612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacques, C.; Chevrollier, A.; Loiseau, D.; Lagoutte, L.; Savagner, F.; Malthièry, Y.; Reynier, P. mtDNA controls expression of the death associated protein 3. Exp. Cell Res. 2006, 312, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Casula, M.; Bosboom-Dobbelaer, I.; Smolders, K.; Otto, S.; Bakker, M.; de Baar, M.P.; Reiss, P.; de Ronde, A. Infection with HIV-1 induces a decrease in mtDNA. J. Infect. Dis. 2005, 191, 1468–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshino, H.; Kumai, Y.; Kashiwakura, I. Effects of endoplasmic reticulum stress on apoptosis induction in radioresistant macrophages. Mol. Med. Rep. 2017, 15, 2867–2872. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Yoshino, H.; Kazama, Y.; Kashiwakura, I. Involvement of caspase-8 in apoptosis enhancement by cotreatment with retinoic acid-inducible gene-I-like receptor agonist and ionizing radiation in human non-small cell lung cancer. Mol. Med. Rep. 2018, 18, 5286–5294. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, H.; Kashiwakura, I. Involvement of reactive oxygen species in ionizing radiation–induced upregulation of cell surface Toll-like receptor 2 and 4 expression in human monocytic cells. J. Radiat. Res. 2017, 58, 626–635. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, H.; Konno, H.; Ogura, K.; Sato, Y.; Kashiwakura, I. Relationship between the regulation of caspase-8-mediated apoptosis and radioresistance in human THP-1-derived macrophages. Int. J. Mol. Sci. 2018, 19, 3154. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, Y.; Yoshino, H.; Kashiwakura, I.; Tsuruga, E. DAP3 Is Involved in Modulation of Cellular Radiation Response by RIG-I-Like Receptor Agonist in Human Lung Adenocarcinoma Cells. Int. J. Mol. Sci. 2021, 22, 420. https://doi.org/10.3390/ijms22010420
Sato Y, Yoshino H, Kashiwakura I, Tsuruga E. DAP3 Is Involved in Modulation of Cellular Radiation Response by RIG-I-Like Receptor Agonist in Human Lung Adenocarcinoma Cells. International Journal of Molecular Sciences. 2021; 22(1):420. https://doi.org/10.3390/ijms22010420
Chicago/Turabian StyleSato, Yoshiaki, Hironori Yoshino, Ikuo Kashiwakura, and Eichi Tsuruga. 2021. "DAP3 Is Involved in Modulation of Cellular Radiation Response by RIG-I-Like Receptor Agonist in Human Lung Adenocarcinoma Cells" International Journal of Molecular Sciences 22, no. 1: 420. https://doi.org/10.3390/ijms22010420