Cell Sheet Comprised of Mesenchymal Stromal Cells Overexpressing Stem Cell Factor Promotes Epicardium Activation and Heart Function Improvement in a Rat Model of Myocardium Infarction

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
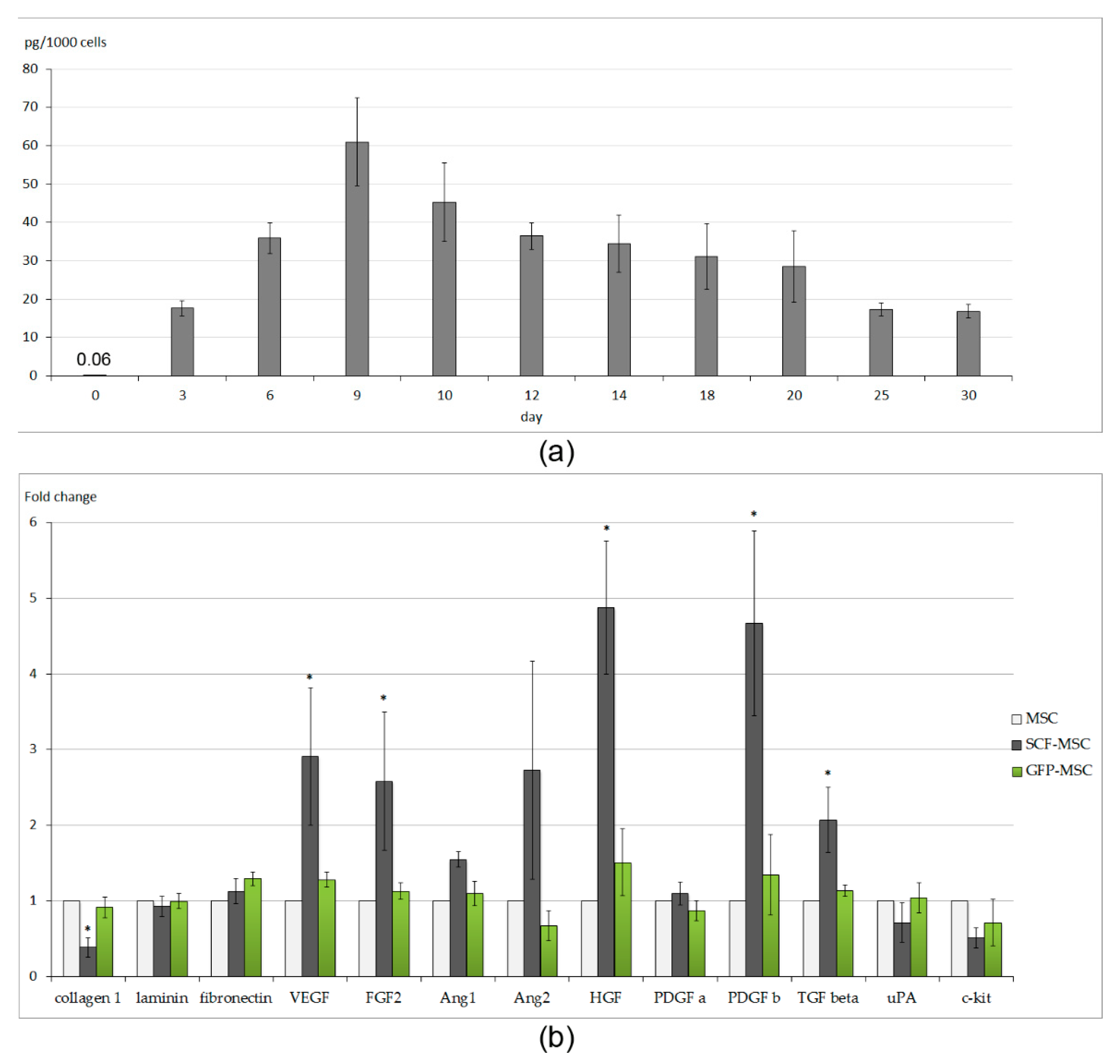
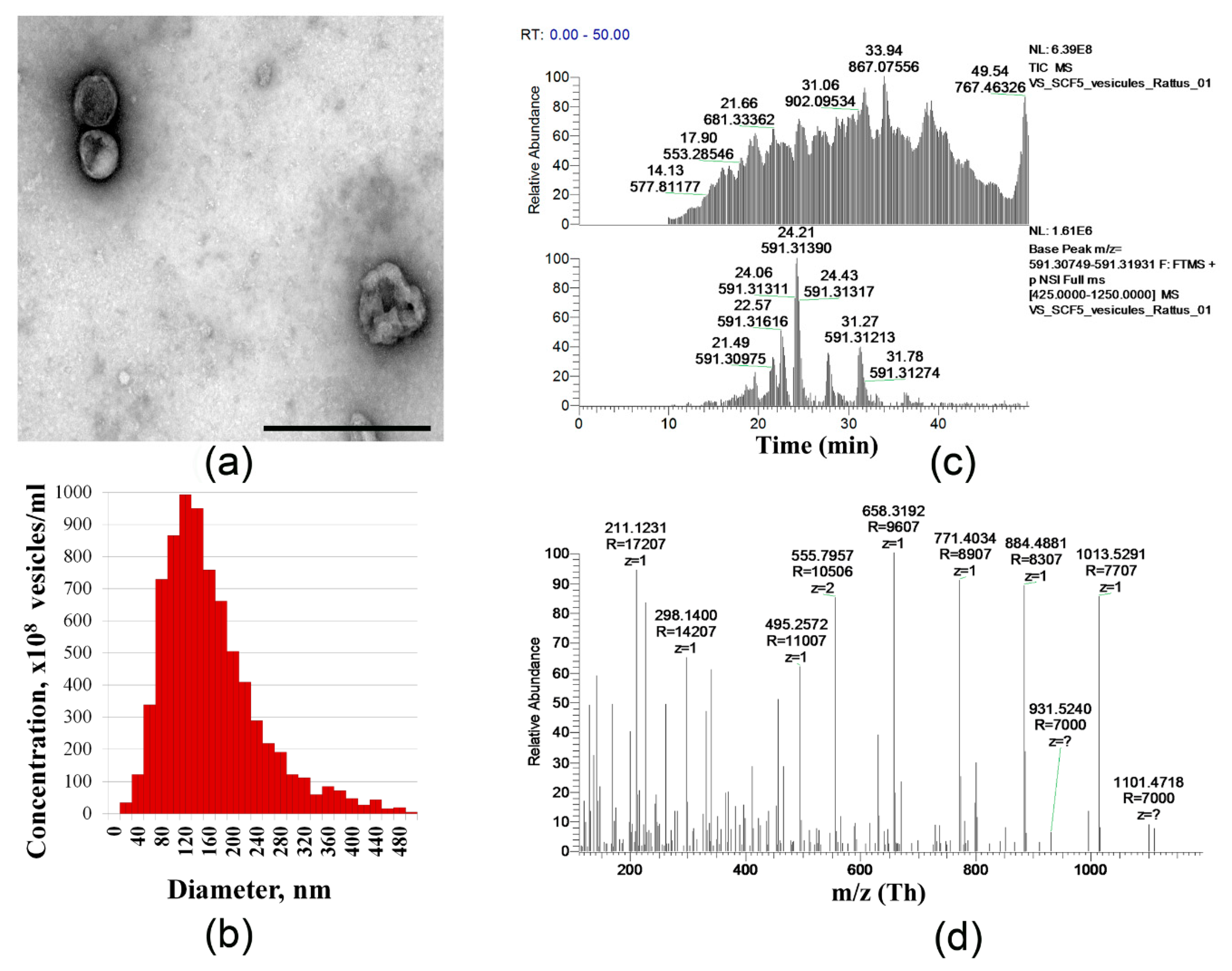
2.1. Characteristics of Cell Sheets Comprised of SCF-Producing MSCs
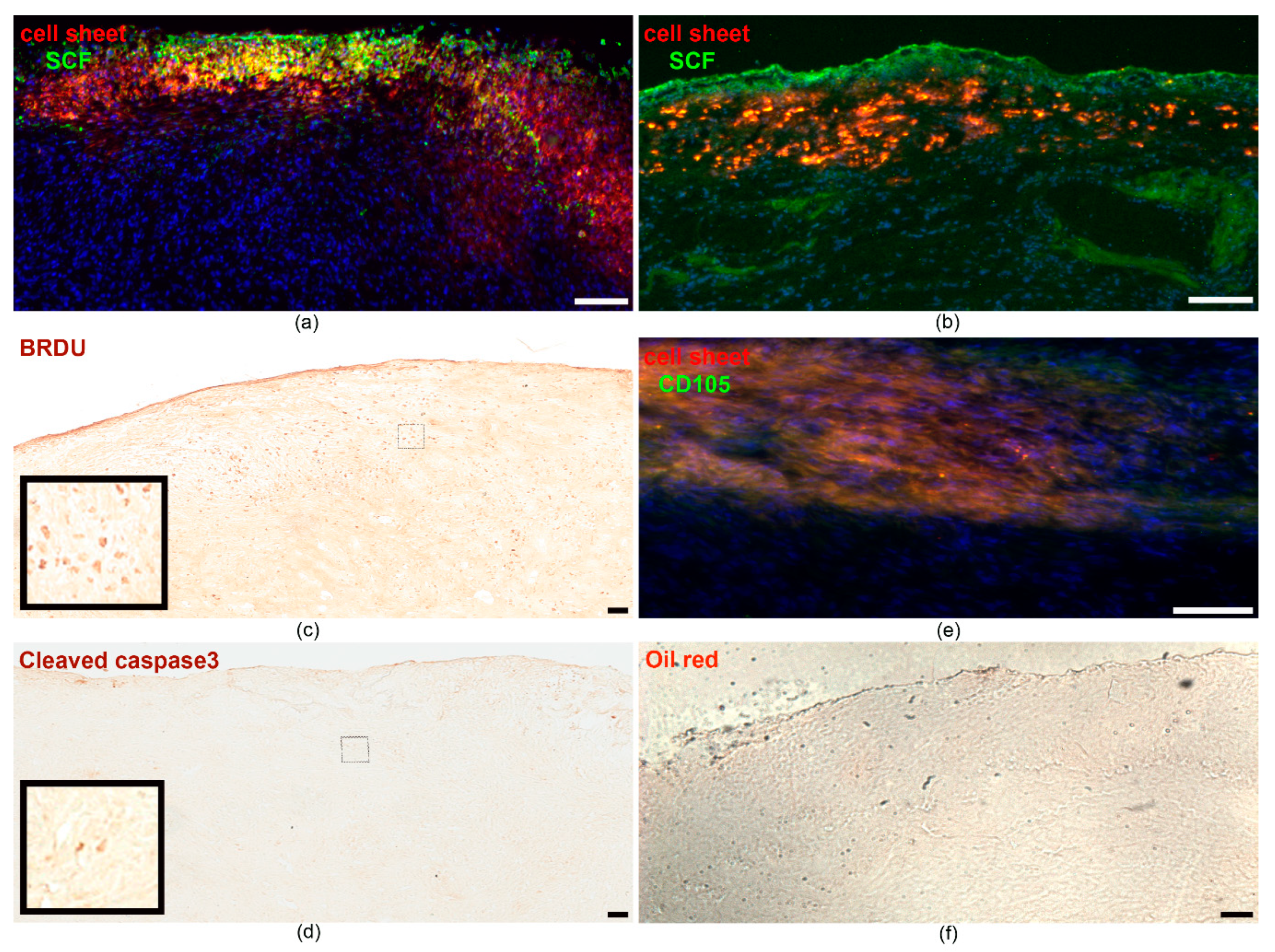
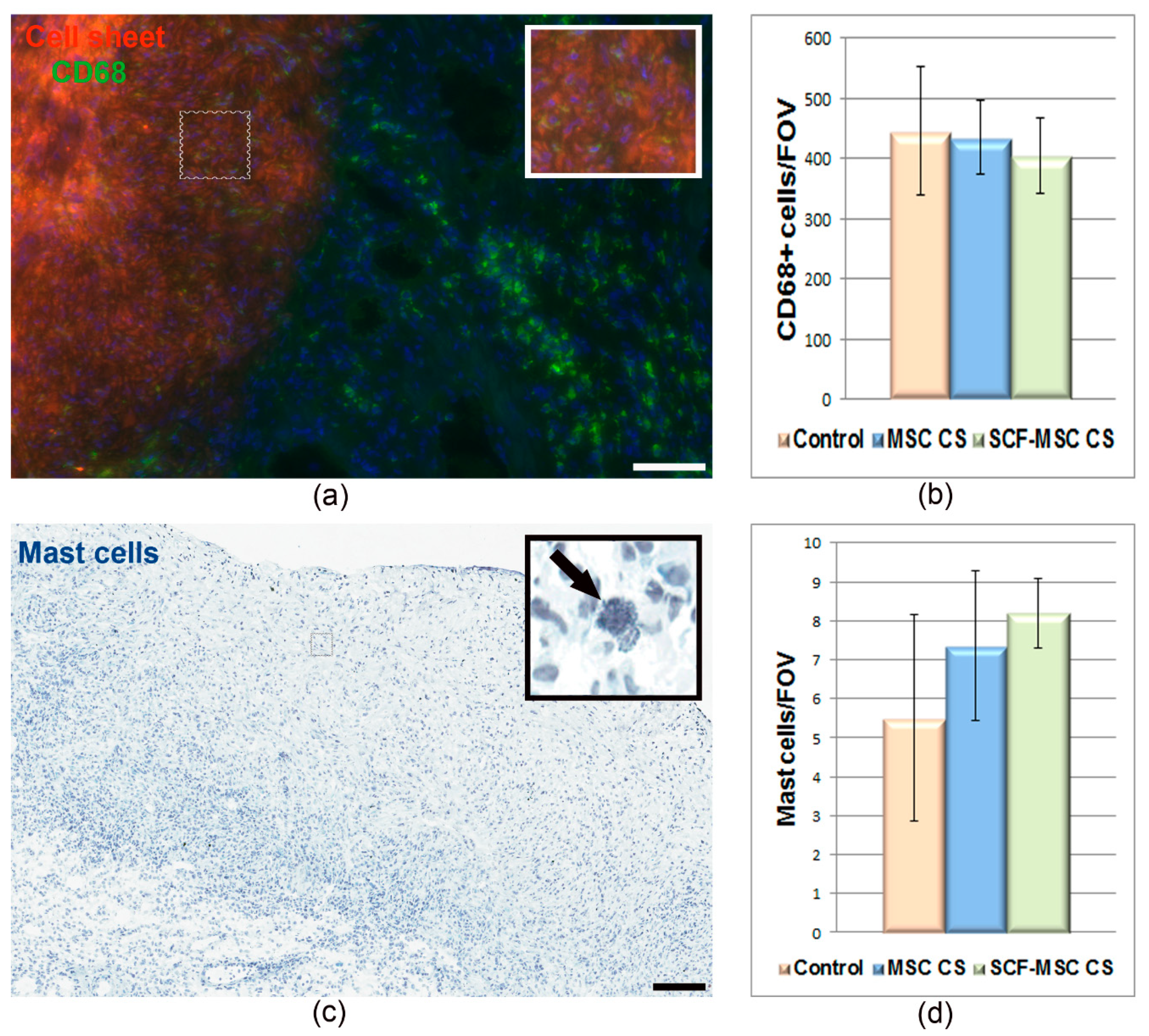
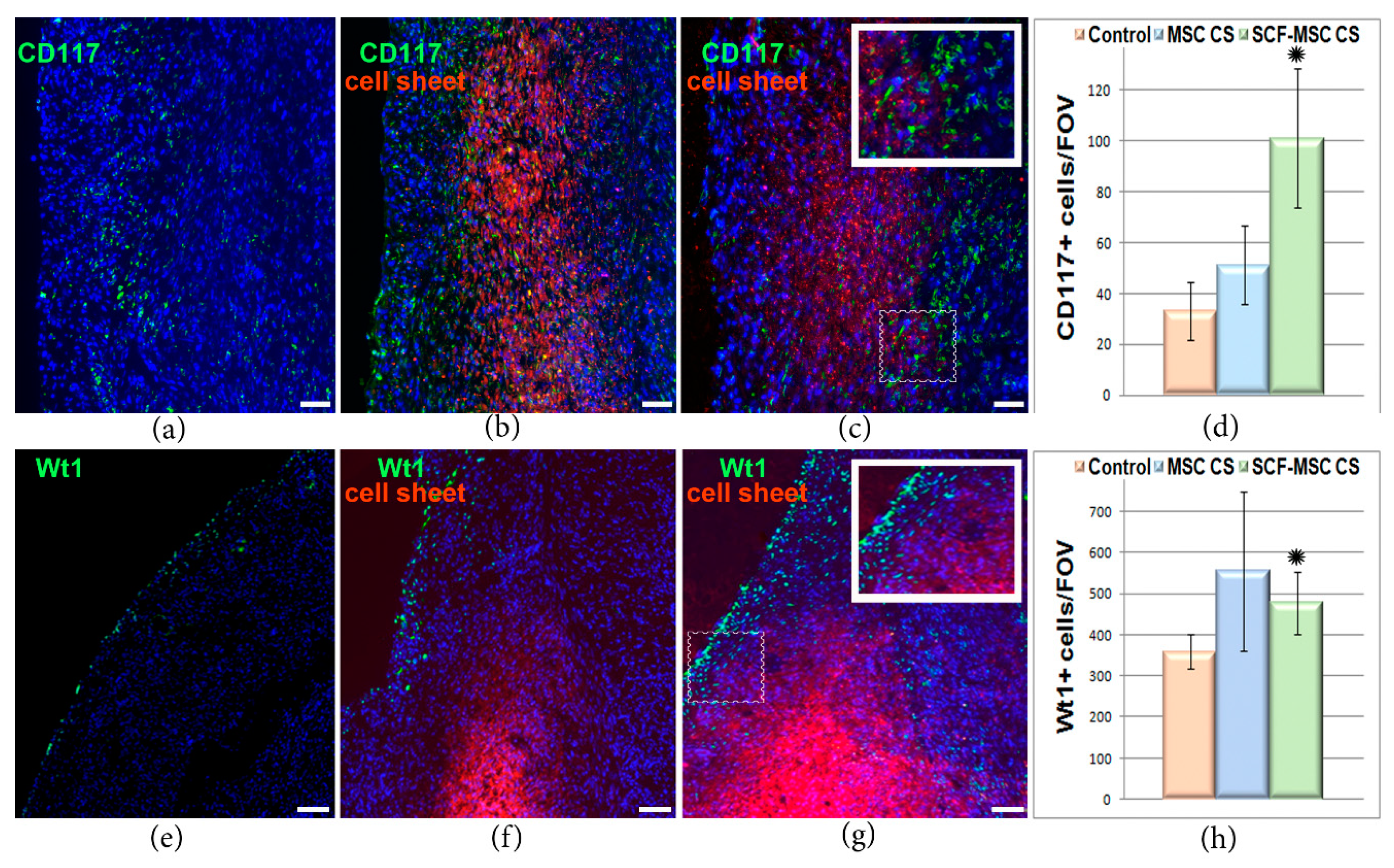
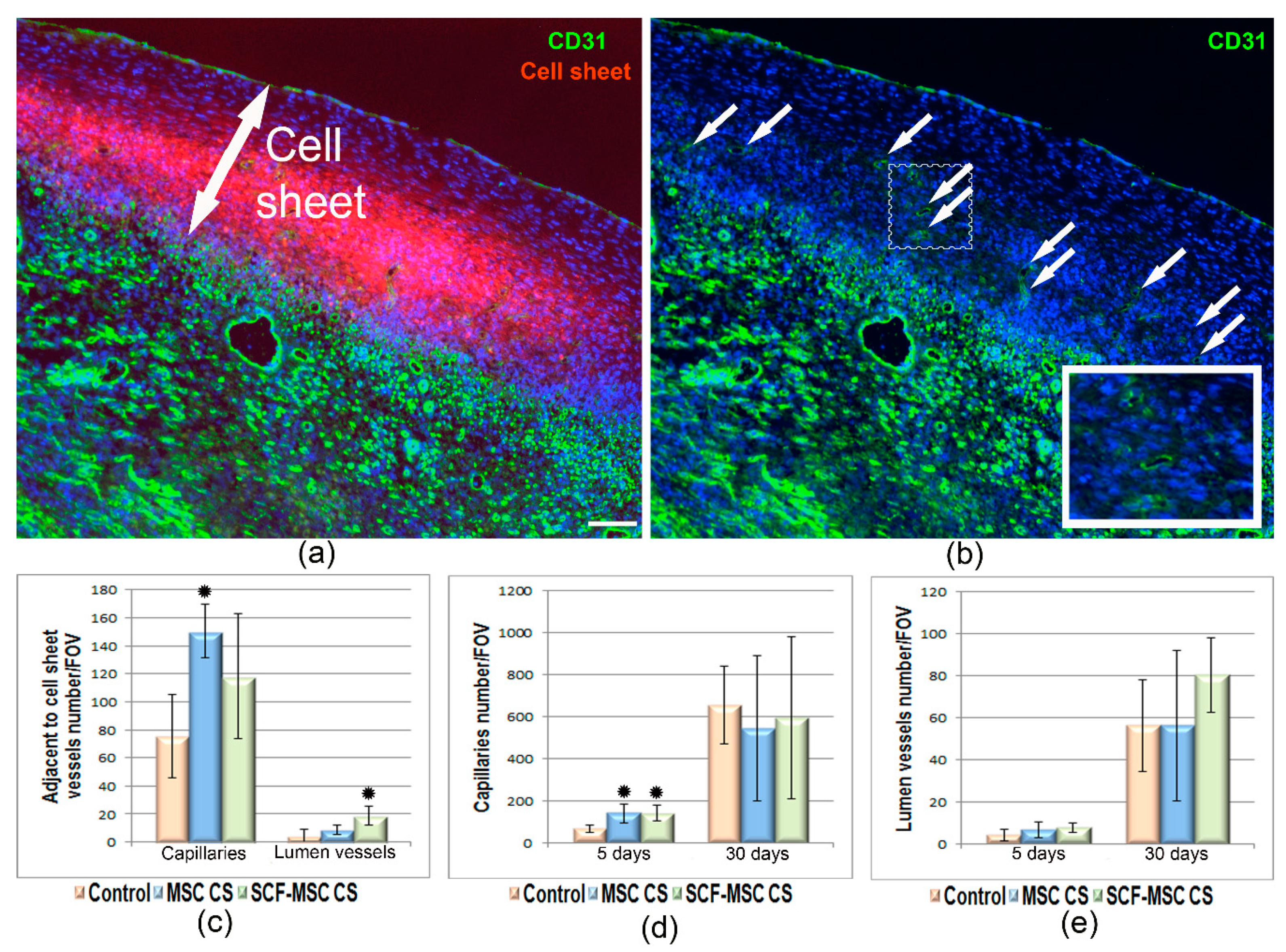
2.2. SCF-MSC CS Transplantation Prevents Tissue Fibrosis, Stimulates Amplification of C-Kit and WT1 Positive Cells in Post-Infarcted Myocardium and Improves Left Ventricle Function
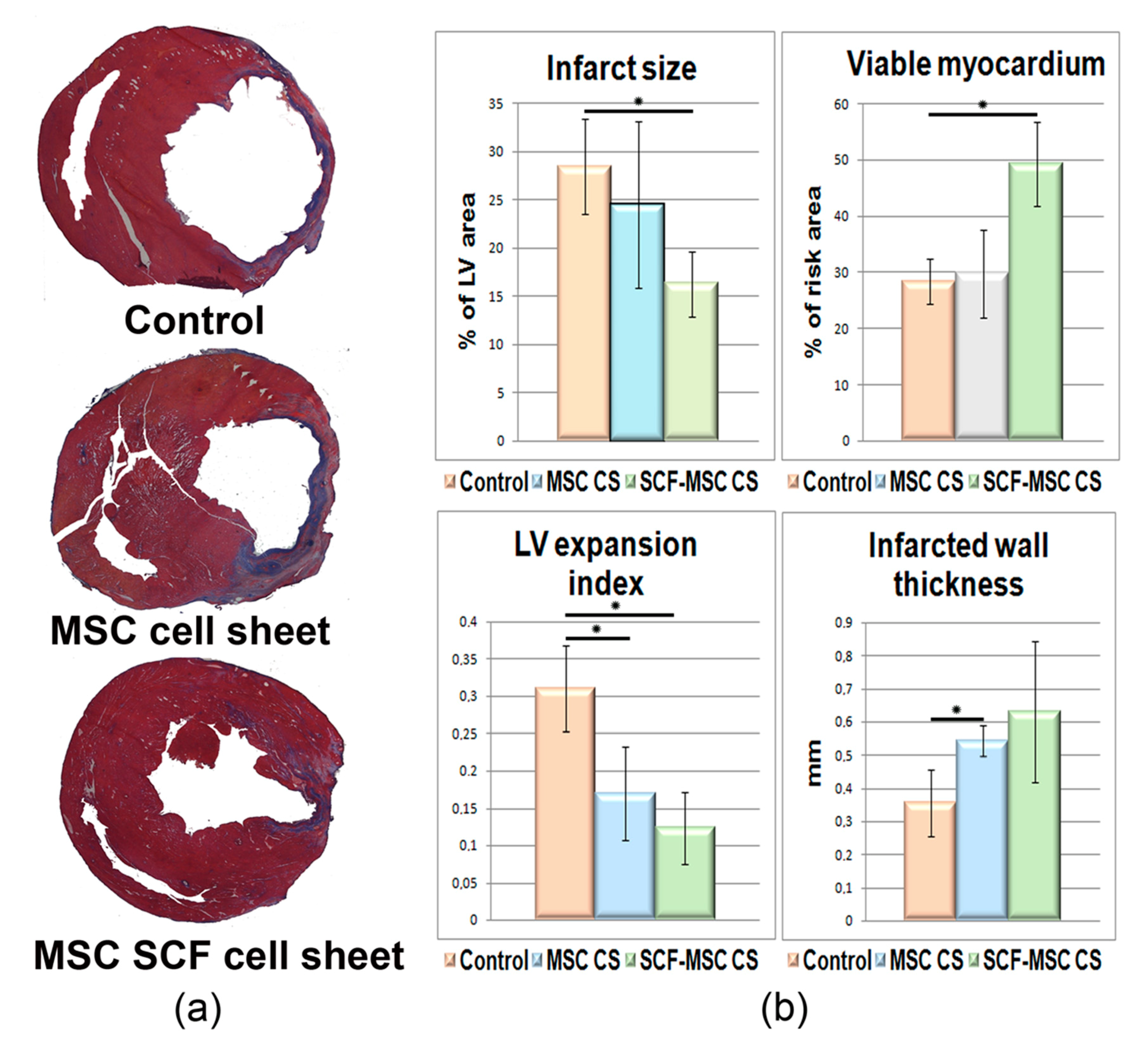
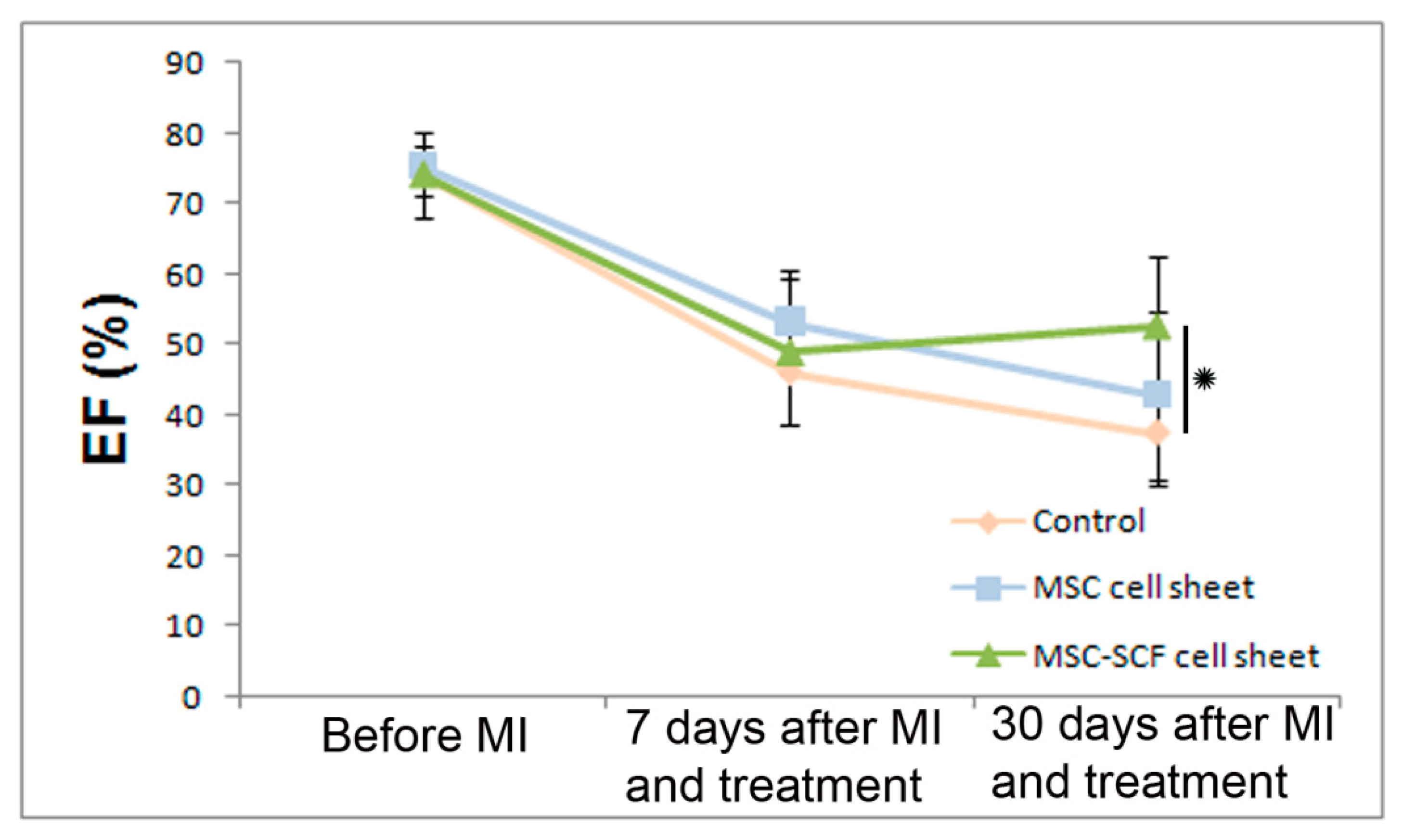
2.3. SCF-MSC CS Transplantation Prevents Tissue Fibrosis and Improves Left Ventricle Function
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Cultures
4.3. Viral Vectors and Cell Transduction
4.4. Cell Sheet Formation
4.5. Real-Time PCR
4.6. Extracellular Vesicles Isolation and Proteomics
4.7. MI Modelling and CS Transplantation
4.8. Histological Analysis
4.9. Morphometric Assay G-5562-2016
4.10. Echocardiography
4.11. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MI | Myocardium infarction |
| MSCs | Mesenchymal stromal cells MSCs |
| SCF | Stem cell factor |
| CS | Cell sheet |
| AAV | Adeno-associated virus |
| EM | Extracellular matrix |
| VEGF | Vascular endothelial growth factor |
| FGF2 | Fibroblast growth factor-2 |
| TGF-b | Tumor growth factor-b |
| HGF | Hepatocyte growth factor |
| CPC | Cardiac progenitor cells |
References
- Nguyen, P.K.; Rhee, J.-W.; Wu, J.C. Adult Stem Cell Therapy and Heart Failure, 2000 to 2016: A Systematic Review. JAMA Cardiol. 2016, 1, 831–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalu, M.M.; Mazzarello, S.; Zlepnig, J.; Dong, Y.Y.; Montroy, J.; McIntyre, L.; Devereaux, P.J.; Stewart, D.J.; Mazer, C.D.; Barron, C.C.; et al. Safety and Efficacy of Adult Stem Cell Therapy for Acute Myocardial Infarction and Ischemic Heart Failure (SafeCell Heart): A Systematic Review and Meta-Analysis. Stem Cells Transl. Med. 2018, 7, 857–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rikhtegar, R.; Pezeshkian, M.; Dolati, S.; Safaie, N.; Rad, A.A.; Mahdipour, M.; Nouri, M.; Jodati, A.R.; Yousefi, M. Stem cells as therapy for heart disease: iPSCs, ESCs, CSCs, and skeletal myoblasts. Biomed. Pharmacother. 2019, 109, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Li, C. Stem/Progenitor Cells and Their Therapeutic Application in Cardiovascular Disease. Front. Cell Dev. Biol. 2018, 6, 139. [Google Scholar] [CrossRef] [PubMed]
- Prasad, M.; Corban, M.T.; Henry, T.D.; Dietz, A.B.; Lerman, L.O.; Lerman, A. Promise of autologous CD34+ stem/progenitor cell therapy for treatment of cardiovascular disease. Cardiovasc. Res. 2020, 116, 1424–1433. [Google Scholar] [CrossRef]
- Nasser, M.I.; Qi, X.; Zhu, S.; He, Y.; Zhao, M.; Guo, H.; Zhu, P. Current situation and future of stem cells in cardiovascular medicine. Biomed. Pharmacother. 2020, 132, 110813. [Google Scholar] [CrossRef]
- Samak, M.; Hinkel, R. Stem Cells in Cardiovascular Medicine: Historical Overview and Future Prospects. Cells 2019, 8, 1530. [Google Scholar] [CrossRef] [Green Version]
- Goradel, N.H.; Hour, F.G.; Negahdari, B.; Malekshahi, Z.V.; Hashemzehi, M.; Masoudifar, A.; Mirzaei, H. Stem Cell Therapy: A New Therapeutic Option for Cardiovascular Diseases. J. Cell. Biochem. 2018, 119, 95–104. [Google Scholar] [CrossRef]
- Müller, P.; Lemcke, H.; David, R. Stem Cell Therapy in Heart Diseases-Cell Types, Mechanisms and Improvement Strategies. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 48, 2607–2655. [Google Scholar] [CrossRef]
- Vagnozzi, R.J.; Maillet, M.; Sargent, M.A.; Khalil, H.; Johansen, A.K.Z.; Schwanekamp, J.A.; York, A.J.; Huang, V.; Nahrendorf, M.; Sadayappan, S.; et al. An acute immune response underlies the benefit of cardiac stem cell therapy. Nature 2020, 577, 405–409. [Google Scholar] [CrossRef]
- Santini, M.P.; Forte, E.; Harvey, R.P.; Kovacic, J.C. Developmental origin and lineage plasticity of endogenous cardiac stem cells. Development (Camb. Engl.) 2016, 143, 1242–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witman, N.; Sahara, M. Cardiac Progenitor Cells in Basic Biology and Regenerative Medicine. Stem Cells Int. 2018, 2018, 8283648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultana, N.; Zhang, L.; Yan, J.; Chen, J.; Cai, W.; Razzaque, S.; Jeong, D.; Sheng, W.; Bu, L.; Xu, M.; et al. Resident c-kit(+) cells in the heart are not cardiac stem cells. Nat. Commun. 2015, 6, 8701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elhelaly, W.M.; Cardoso, A.C.; Pereira, A.H.M.; Elnawasany, A.; Ebrahimi, S.; Nakada, Y.; Sadek, H.A. C-Kit Cells Do Not Significantly Contribute to Cardiomyogenesis During Neonatal Heart Regeneration. Circulation 2019, 139, 559–561. [Google Scholar] [CrossRef]
- Fan, X.L.; Zhang, Y.; Li, X.; Fu, Q.L. Mechanisms underlying the protective effects of mesenchymal stem cell-based therapy. Cell. Mol. Life Sci. CMLS 2020, 77, 2771–2794. [Google Scholar] [CrossRef] [Green Version]
- Bagno, L.; Hatzistergos, K.E.; Balkan, W.; Hare, J.M. Mesenchymal Stem Cell-Based Therapy for Cardiovascular Disease: Progress and Challenges. Mol. Ther. J. Am. Soc. Gene Ther. 2018, 26, 1610–1623. [Google Scholar] [CrossRef] [Green Version]
- Lemcke, H.; Voronina, N.; Steinhoff, G.; David, R. Recent Progress in Stem Cell Modification for Cardiac Regeneration. Stem Cells Int. 2018, 2018, 1909346. [Google Scholar] [CrossRef]
- Wei, W.; Huang, Y.; Li, D.; Gou, H.-F.; Wang, W. Improved therapeutic potential of MSCs by genetic modification. Gene Ther. 2018, 25, 538–547. [Google Scholar] [CrossRef]
- Liang, Y.; Lin, Q.; Zhu, J.; Li, X.; Fu, Y.; Zou, X.; Liu, X.; Tan, H.; Deng, C.; Yu, X.; et al. The caspase-8 shRNA-modified mesenchymal stem cells improve the function of infarcted heart. Mol. Cell Biochem. 2014, 397, 7–16. [Google Scholar] [CrossRef]
- Gnecchi, M.; He, H.; Liang, O.D.; Melo, L.G.; Morello, F.; Mu, H.; Noiseux, N.; Zhang, L.; Pratt, R.E.; Ingwall, J.S.; et al. Paracrine action accounts for marked protection of ischemic heart by Akt-modified mesenchymal stem cells. Nat. Med. 2005, 11, 367–368. [Google Scholar] [CrossRef]
- He, X.; Dziak, R.; Mao, K.; Genco, R.; Swihart, M.; Li, C.; Yang, S. Integration of a novel injectable nano calcium sulfate/alginate scaffold and BMP2 gene-modified mesenchymal stem cells for bone regeneration. Tissue Eng. Part A 2013, 19, 508–518. [Google Scholar] [CrossRef] [Green Version]
- Shevchenko, E.K.; Makarevich, P.I.; Tsokolaeva, Z.I.; Boldyreva, M.A.; Sysoeva, V.Y.; Tkachuk, V.A.; Parfyonova, Y.V. Transplantation of modified human adipose derived stromal cells expressing VEGF165 results in more efficient angiogenic response in ischemic skeletal muscle. J. Transl. Med. 2013, 11, 138. [Google Scholar] [CrossRef] [Green Version]
- Boldyreva, M.A.; Shevchenko, E.K.; Molokotina, Y.D.; Makarevich, P.I.; Beloglazova, I.B.; Zubkova, E.S.; Dergilev, K.V.; Tsokolaeva, Z.I.; Penkov, D.; Hsu, M.N.; et al. Transplantation of Adipose Stromal Cell Sheet Producing Hepatocyte Growth Factor Induces Pleiotropic Effect in Ischemic Skeletal Muscle. Int. J. Mol. Sci. 2019, 20, 3088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, D.; Zhao, X.; Xiao, G.; Ni, J.; Feng, Y.; Wu, R.; Wang, G. Stem cell factor/c-kit signaling mediated cardiac stem cell migration via activation of p38 MAPK. Basic Res. Cardiol. 2008, 103, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Park, S.N.; Suh, H. Generation of insulin-producing human mesenchymal stem cells using recombinant adeno-associated virus. Yonsei Med. J. 2007, 48, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Hermanns, J.; Schulze, A.; Jansen-Db1urr, P.; Kleinschmidt, J.A.; Schmidt, R.; zur Hausen, H. Infection of primary cells by adeno-associated virus type 2 results in a modulation of cell cycle-regulating proteins. J. Virol. 1997, 71, 6020–6027. [Google Scholar] [CrossRef] [Green Version]
- Saudan, P.; Vlach, J.; Beard, P. Inhibition of S-phase progression by adeno-associated virus Rep78 protein is mediated by hypophosphorylated pRb. EMBO J. 2000, 19, 4351–4361. [Google Scholar] [CrossRef] [Green Version]
- Blazquez-Martinez, A.; Chiesa, M.; Arnalich, F.; Fernandez-Delgado, J.; Nistal, M.; De Miguel, M.P. c-Kit identifies a subpopulation of mesenchymal stem cells in adipose tissue with higher telomerase expression and differentiation potential. Differentiation 2014, 87, 147–160. [Google Scholar] [CrossRef]
- He, N.; Zhang, Y.; Zhang, S.; Wang, D.; Ye, H. Exosomes: Cell-Free Therapy for Cardiovascular Diseases. J. Cardiovasc. Transl. Res. 2020, 13, 713–721. [Google Scholar] [CrossRef]
- Tikhomirov, R.; Donnell, B.R.; Catapano, F.; Faggian, G.; Gorelik, J.; Martelli, F.; Emanueli, C. Exosomes: From Potential Culprits to New Therapeutic Promise in the Setting of Cardiac Fibrosis. Cells 2020, 9, 592. [Google Scholar] [CrossRef] [Green Version]
- Kovacic, J.C.; Mercader, N.; Torres, M.; Boehm, M.; Fuster, V. Epithelial-to-mesenchymal and endothelial-to-mesenchymal transition: From cardiovascular development to disease. Circulation 2012, 125, 1795–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacic, J.C.; Dimmeler, S.; Harvey, R.P.; Finkel, T.; Aikawa, E.; Krenning, G.; Baker, A.H. Endothelial to Mesenchymal Transition in Cardiovascular Disease: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2019, 73, 190–209. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.L.; Fang, W.W.; Ye, F.; Liu, Y.H.; Qian, J.; Shan, S.J.; Zhang, J.J.; Chunhua, R.Z.; Liao, L.M.; Lin, S.; et al. Effect on left ventricular function of intracoronary transplantation of autologous bone marrow mesenchymal stem cell in patients with acute myocardial infarction. Am. J. Cardiol. 2004, 94, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Chullikana, A.; Majumdar, A.S.; Gottipamula, S.; Krishnamurthy, S.; Kumar, A.S.; Prakash, V.S.; Gupta, P.K. Randomized, double-blind, phase I/II study of intravenous allogeneic mesenchymal stromal cells in acute myocardial infarction. Cytotherapy 2015, 17, 250–261. [Google Scholar] [CrossRef]
- Florea, V.; Rieger, A.C.; DiFede, D.L.; El-Khorazaty, J.; Natsumeda, M.; Banerjee, M.N.; Tompkins, B.A.; Khan, A.; Schulman, I.H.; Landin, A.M.; et al. Dose Comparison Study of Allogeneic Mesenchymal Stem Cells in Patients with Ischemic Cardiomyopathy (The TRIDENT Study). Circ. Res. 2017, 121, 1279–1290. [Google Scholar] [CrossRef]
- Kang, W.J.; Kang, H.J.; Kim, H.S.; Chung, J.K.; Lee, M.C.; Lee, D.S. Tissue distribution of 18F-FDG-labeled peripheral hematopoietic stem cells after intracoronary administration in patients with myocardial infarction. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2006, 47, 1295–1301. [Google Scholar]
- Sawa, Y.; Miyagawa, S. Present and future perspectives on cell sheet-based myocardial regeneration therapy. Biomed. Res. Int. 2013, 2013, 583912. [Google Scholar] [CrossRef]
- Makarevich, P.I.; Boldyreva, M.A.; Gluhanyuk, E.V.; Efimenko, A.Y.; Dergilev, K.V.; Shevchenko, E.K.; Sharonov, G.V.; Gallinger, J.O.; Rodina, P.A.; Sarkisyan, S.S.; et al. Enhanced angiogenesis in ischemic skeletal muscle after transplantation of cell sheets from baculovirus-transduced adipose-derived stromal cells expressing VEGF165. Stem Cell Res. Ther. 2015, 6, 204. [Google Scholar] [CrossRef] [Green Version]
- Dergilev, K.; Tsokolaeva, Z.; Makarevich, P.; Beloglazova, I.; Zubkova, E.; Boldyreva, M.; Ratner, E.; Dyikanov, D.; Menshikov, M.; Ovchinnikov, A.; et al. C-Kit Cardiac Progenitor Cell Based Cell Sheet Improves Vascularization and Attenuates Cardiac Remodeling following Myocardial Infarction in Rats. Biomed. Res. Int. 2018, 2018, 3536854. [Google Scholar] [CrossRef]
- Li, Z.; MacDougald, O.A. Stem cell factor: The bridge between bone marrow adipocytes and hematopoietic cells. Haematologica 2019, 104, 1689–1691. [Google Scholar] [CrossRef]
- Fazel, S.; Cimini, M.; Chen, L.; Li, S.; Angoulvant, D.; Fedak, P.; Verma, S.; Weisel, R.D.; Keating, A.; Li, R.-K. Cardioprotective c-kit+ cells are from the bone marrow and regulate the myocardial balance of angiogenic cytokines. J. Clin. Investig. 2006, 116, 1865–1877. [Google Scholar] [CrossRef] [PubMed]
- Hatzistergos, K.E.; Saur, D.; Seidler, B.; Balkan, W.; Breton, M.; Valasaki, K.; Takeuchi, L.M.; Landin, A.M.; Khan, A.; Hare, J.M. Stimulatory effects of MSCs on cKit+ Cardiac Stem Cells are Mediated by SDF1/CXCR4 and SCF/cKit Signaling Pathways. Circ. Res. 2016, 119, 921–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vajravelu, B.N.; Hong, K.U.; Al-Maqtari, T.; Cao, P.; Keith, M.C.L.; Wysoczynski, M.; John, Z.; Moore, J.B., IV; Bolli, R. C-Kit Promotes Growth and Migration of Human Cardiac Progenitor Cells via the PI3K-AKT and MEK-ERK Pathways. PLoS ONE 2015, 10, e0140798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Jie, W.; Shen, Z.; Li, M.; Lan, Y.; Kong, Y.; Guo, S.; Li, T.; Zheng, S. SCF increases cardiac stem cell migration through PI3K/AKT and MMP-2/-9 signaling. Int. J. Mol. Med. 2014, 34, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Vicinanza, C.; Aquila, I.; Scalise, M.; Cristiano, F.; Marino, F.; Cianflone, E.; Mancuso, T.; Marotta, P.; Sacco, W.; Lewis, F.C.; et al. Adult cardiac stem cells are multipotent and robustly myogenic: C-kit expression is necessary but not sufficient for their identification. Cell Death Differ. 2017, 24, 2101–2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Drummond, C.A.; Fan, X.; Haller, S.T.; Liu, J.; Malhotra, D.; Tian, J. Hiding inside? Intracellular expression of non-glycosylated c-kit protein in cardiac progenitor cells. Stem Cell Res. 2016, 16, 795–806. [Google Scholar] [CrossRef] [Green Version]
- Chimenti, I.; Gaetani, R.; Barile, L.; Frati, G.; Messina, E.; Giacomello, A. c-kit cardiac progenitor cells: What is their potential? Proc. Natl. Acad. Sci. USA 2009, 106, E78. [Google Scholar] [CrossRef] [Green Version]
- Stastna, M.; Chimenti, I.; Marbán, E.; Van Eyk, J.E. Identification and functionality of proteomes secreted by rat cardiac stem cells and neonatal cardiomyocytes. Proteomics 2010, 10, 245–253. [Google Scholar] [CrossRef]
- Smits, A.M.; Dronkers, E.; Goumans, M.-J. The epicardium as a source of multipotent adult cardiac progenitor cells: Their origin, role and fate. Pharmacol. Res. 2018, 127, 129–140. [Google Scholar] [CrossRef]
- Velecela, V.; Torres-Cano, A.; García-Melero, A.; Ramiro-Pareta, M.; Müller-Sánchez, C.; Segarra-Mondejar, M.; Chau, Y.-Y.; Campos-Bonilla, B.; Reina, M.; Soriano, F.X.; et al. Epicardial cell shape and maturation are regulated by Wt1 via transcriptional control of Bmp4. Development 2019, 146, dev178723. [Google Scholar] [CrossRef] [Green Version]
- Gittenberger-de Groot, A.C.; Winter, E.M.; Poelmann, R.E. Epicardium-derived cells (EPDCs) in development, cardiac disease and repair of ischemia. J. Cell Mol. Med. 2010, 14, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Traktuev, D.O.; Tsokolaeva, Z.I.; Shevelev, A.A.; Tkachuk, V.A.; March, K.L.; Parfyonova, Y.V. Urokinase Gene Transfer Augments Angiogenesis in Ischemic Skeletal and Myocardial Muscle. Mol. Ther. 2007, 15, 1939–1946. [Google Scholar] [CrossRef] [PubMed]
- Hochman, J.S.; Choo, H. Limitation of myocardial infarct expansion by reperfusion independent of myocardial salvage. Circulation 1987, 75, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Forward | Primer Reverse |
|---|---|---|
| B-Actin | AGCCATGTACGTAGCCATCC | CTCTCAGCTGTGGTGGTGAA |
| VEGF | ACTGGACCCTGGCTTTACTG | ACGCACTCCAGGGCTTCATC |
| Angiopoietin1 | GTGGCTGGAAAAACTTGAGA | TGGATTTCAAGACGGGATGT |
| bFGF | AAGCGGCTCTACTGCAAG | AGCCAGACATTGGAAGAAACA |
| PDGFa | CCTGTGCCATCCGCAGGAAGAGA | TTGGCCACCTTGACGCTGCGGTG |
| PDGFb | GATCCGCTCCTTTGATGATC | GTCTCACACTTGCATGCCAG |
| HGF | CCAGCTAGAAACAAAGACTTGAAAGA | GAAATGTTTAAGATCTGTTTGCGTT |
| FGF2 | ACGGCGTCCGGGAGAA | ACACTCCCTTGATGGACACAACT |
| TGF-β1 | TGCTTCAGCTCCACAGAGAA | TGGTTGTAGAGGGCAAGGAC |
| c-kit | CACTCACGGGCGGATCAC | TCCGGTGCCATCCACTTCAC |
| Collagen I | CAACCTCAAGAAGTCCCTGC | AGGTGAATCGACTGTTGCCT |
| Fibronectin | CCTTAAGCCTTCTGCTCTGG | CGGCAAAAGAAAGCAGAACT |
| uPA | CTGACCCAGAGTGGAAAACAG | CGGCCATCGATGTTACAGAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dergilev, K.V.; Shevchenko, E.K.; Tsokolaeva, Z.I.; Beloglazova, I.B.; Zubkova, E.S.; Boldyreva, M.A.; Menshikov, M.Y.; Ratner, E.I.; Penkov, D.; Parfyonova, Y.V. Cell Sheet Comprised of Mesenchymal Stromal Cells Overexpressing Stem Cell Factor Promotes Epicardium Activation and Heart Function Improvement in a Rat Model of Myocardium Infarction. Int. J. Mol. Sci. 2020, 21, 9603. https://doi.org/10.3390/ijms21249603
Dergilev KV, Shevchenko EK, Tsokolaeva ZI, Beloglazova IB, Zubkova ES, Boldyreva MA, Menshikov MY, Ratner EI, Penkov D, Parfyonova YV. Cell Sheet Comprised of Mesenchymal Stromal Cells Overexpressing Stem Cell Factor Promotes Epicardium Activation and Heart Function Improvement in a Rat Model of Myocardium Infarction. International Journal of Molecular Sciences. 2020; 21(24):9603. https://doi.org/10.3390/ijms21249603
Chicago/Turabian StyleDergilev, Konstantin V., Evgeny K. Shevchenko, Zoya I. Tsokolaeva, Irina B. Beloglazova, Ekaterina S. Zubkova, Maria A. Boldyreva, Mikhail Yu. Menshikov, Elizaveta I. Ratner, Dmitry Penkov, and Yelena V. Parfyonova. 2020. "Cell Sheet Comprised of Mesenchymal Stromal Cells Overexpressing Stem Cell Factor Promotes Epicardium Activation and Heart Function Improvement in a Rat Model of Myocardium Infarction" International Journal of Molecular Sciences 21, no. 24: 9603. https://doi.org/10.3390/ijms21249603