Distinctive Features of Orbital Adipose Tissue (OAT) in Graves’ Orbitopathy

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
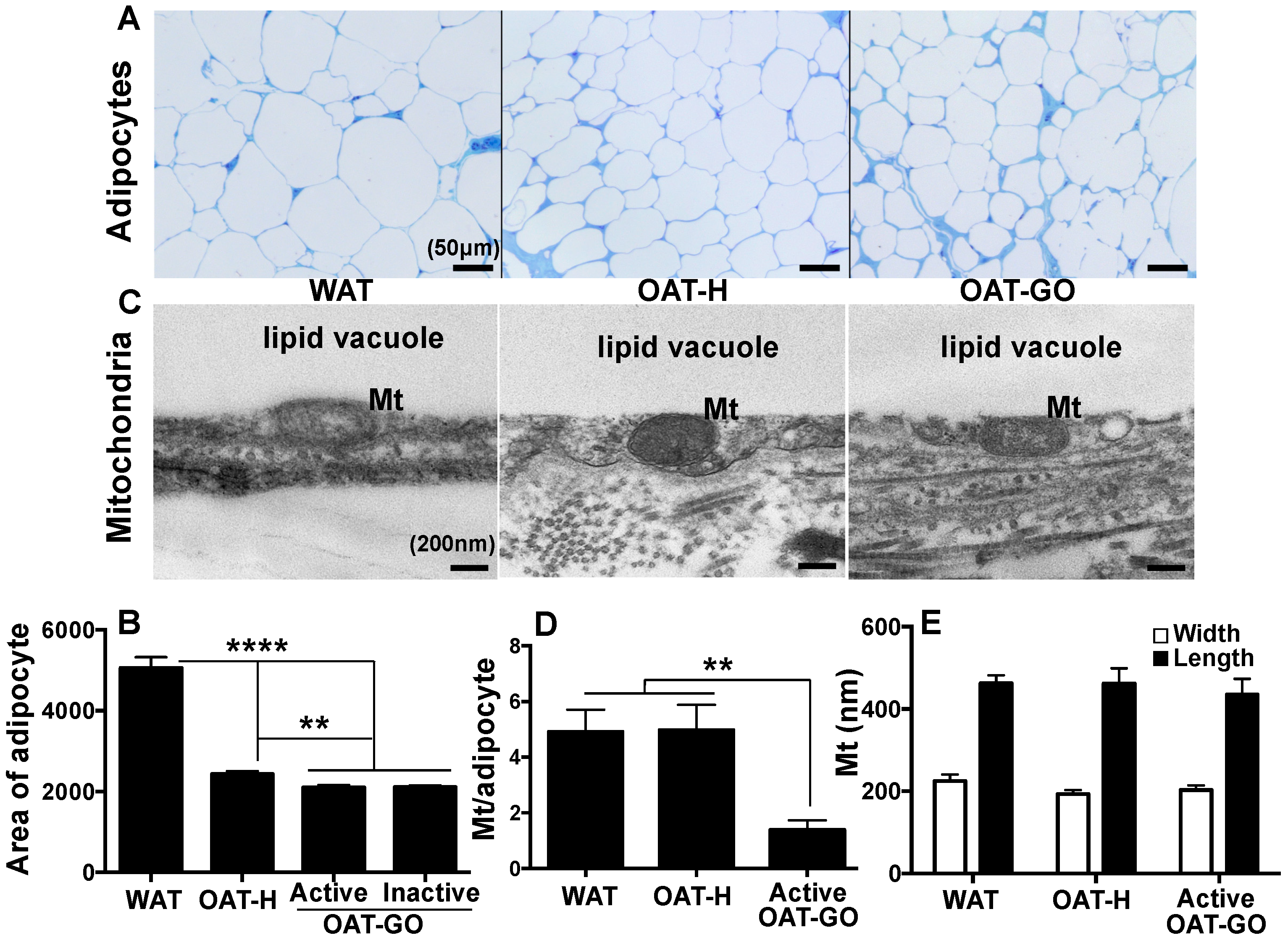
2.1. Adipocytes Size and Mitochondria Count per Adipocyte
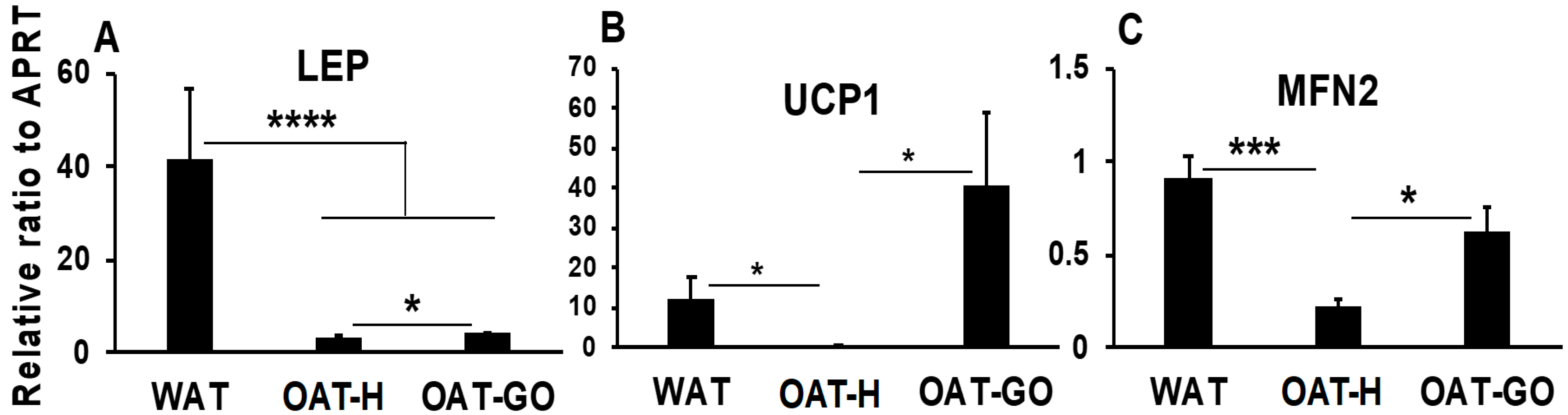
2.2. Differences in OAT versus WAT or BAT/BRITE in Marker Genes
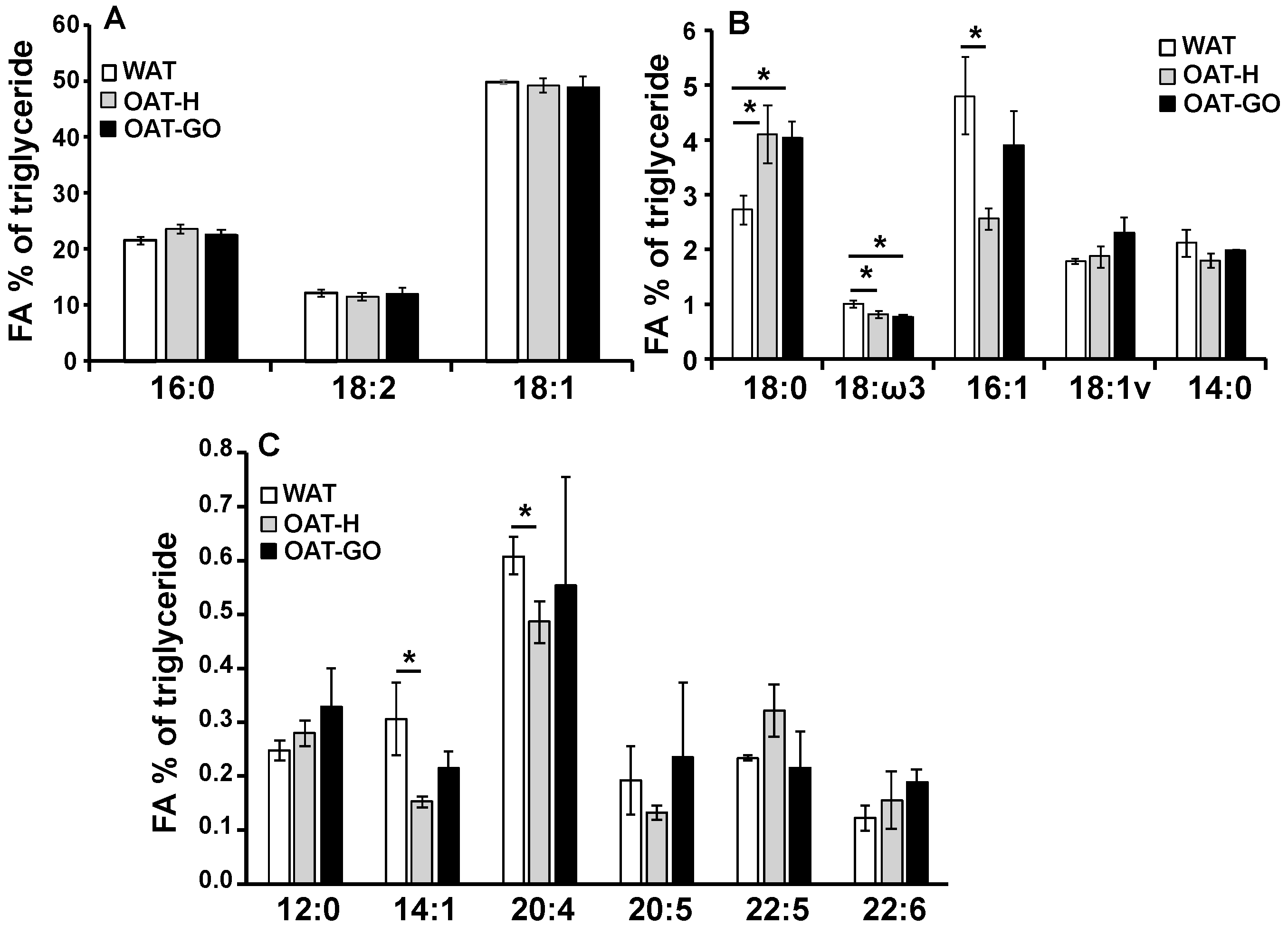
2.3. FA Composition of Triglyceride Comparing OAT and WAT
2.4. Whole Transcriptome Analysis of Lipid Metabolisms in OAT Compared to WAT
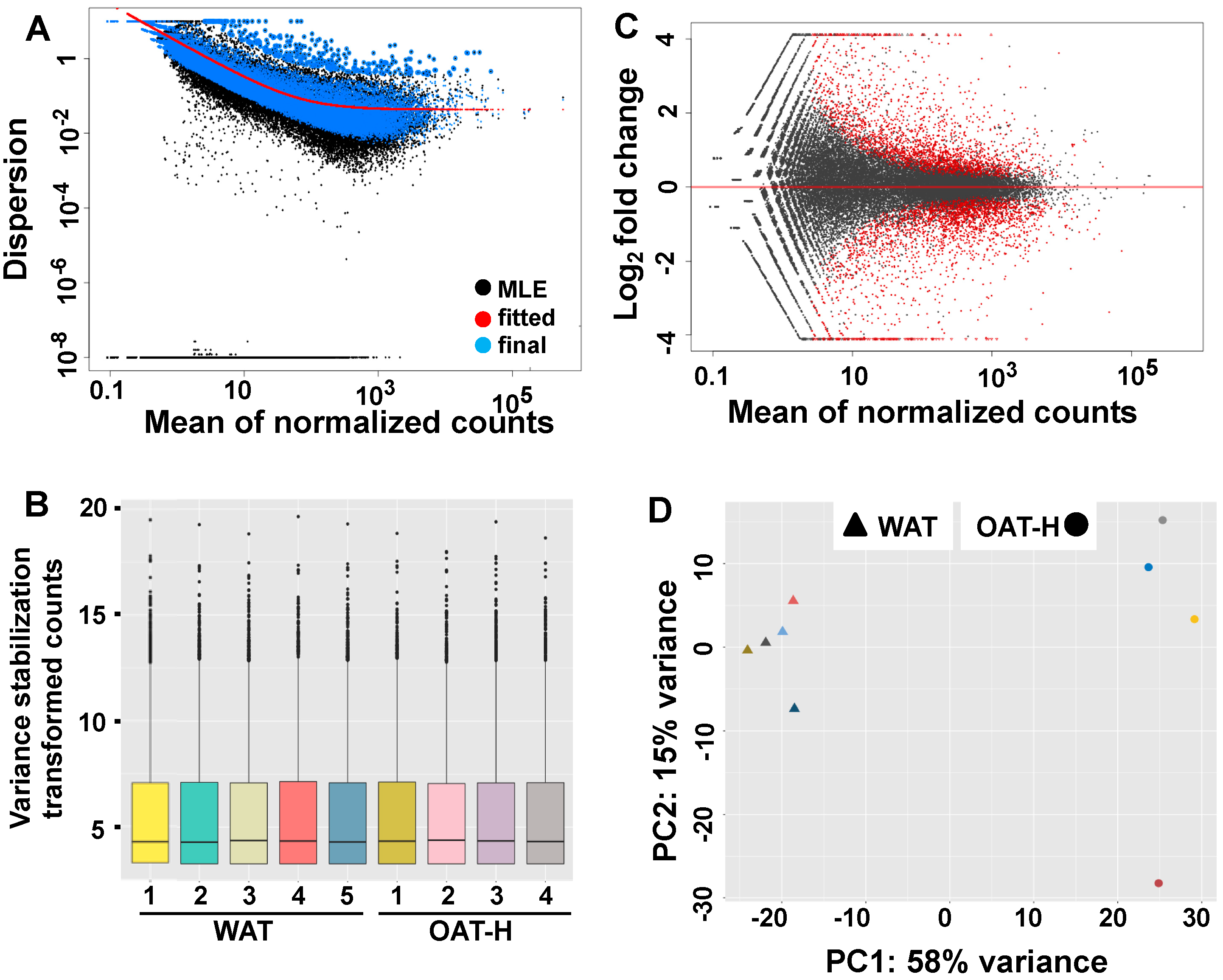
- To better understand OAT, transcriptional profiles were analyzed by RNA sequencing (RNAseq) in ex vivo fat tissue samples of OAT-H (n = 4) compared to WAT (n = 5). A total of 60,668 genes were in the genome build and 23,766 genes (total counts across all samples > 1) were analyzed (Table S1A). Prior to differential gene expression (DGE) analysis, a good fit normalized count of genes from all samples for DESeq2 model had been shown by shrinkage estimation of dispersion plots (Figure 4A), in which the majority of genes were scattered around the curve of expected dispersion value. At sample-level, Figure 4B showed similar distribution patterns across all samples at lower, median and upper quantiles and counts for outlying features.In DGE analysis comparing OAT-H vs. WAT, we obtained 3790 genes with FDR-p significant (<0.05), which has 1746 genes (7.35%) up-regulated and 2044 genes (8.6%) down-regulated. A summarized log2fold change of genes over the mean of normalized counts is shown in the MvA plot (Figure 4C). A PCA-plot at sample-level (Figure 4D) showed a clear separation of samples from OAT-H or WAT that used the top 500 genes (FDR < 0.05) with highest variance (DGE data in Table S1B).
- DGE analysis comparing OAT-H with WAT confirmed substantially lower expression levels of MFN2 and LEP, as well as a significantly increased expression of LEP receptor (LEPR) (Table S1A). Lower expression level of ADIPOQ (adiponectin) was seen in OAT-H compared with WAT with no difference in ADIPOQ receptors (Table S1A).In further analysis, we used a setting of log2fold > 0.5 or <−0.5; FDR-p < 0.05, and obtained 3277 genes (1495 up and 1782 down-regulated) for IPA® analysis. The up- or down-regulated gene set of OAT-H versus WAT was also subject to IPA® analysis. Pathway analysis suggested substantially reduced lipid metabolic activity (overlap-p < 0.01; z-score < −2) including FA metabolism, lipid uptake, uFAs conversion, lipolysis, lipid synthesis, and oxidation of lipid (e.g., ACADM, ECHS1) compared with WAT (Table S2). This evidence for reduced lipid metabolic activity was further supported by IPA® analysis of down-regulated genes. Furthermore, we identified significantly increased expression of the FA transporter, SLC27A6, in OAT-H compared with WAT.
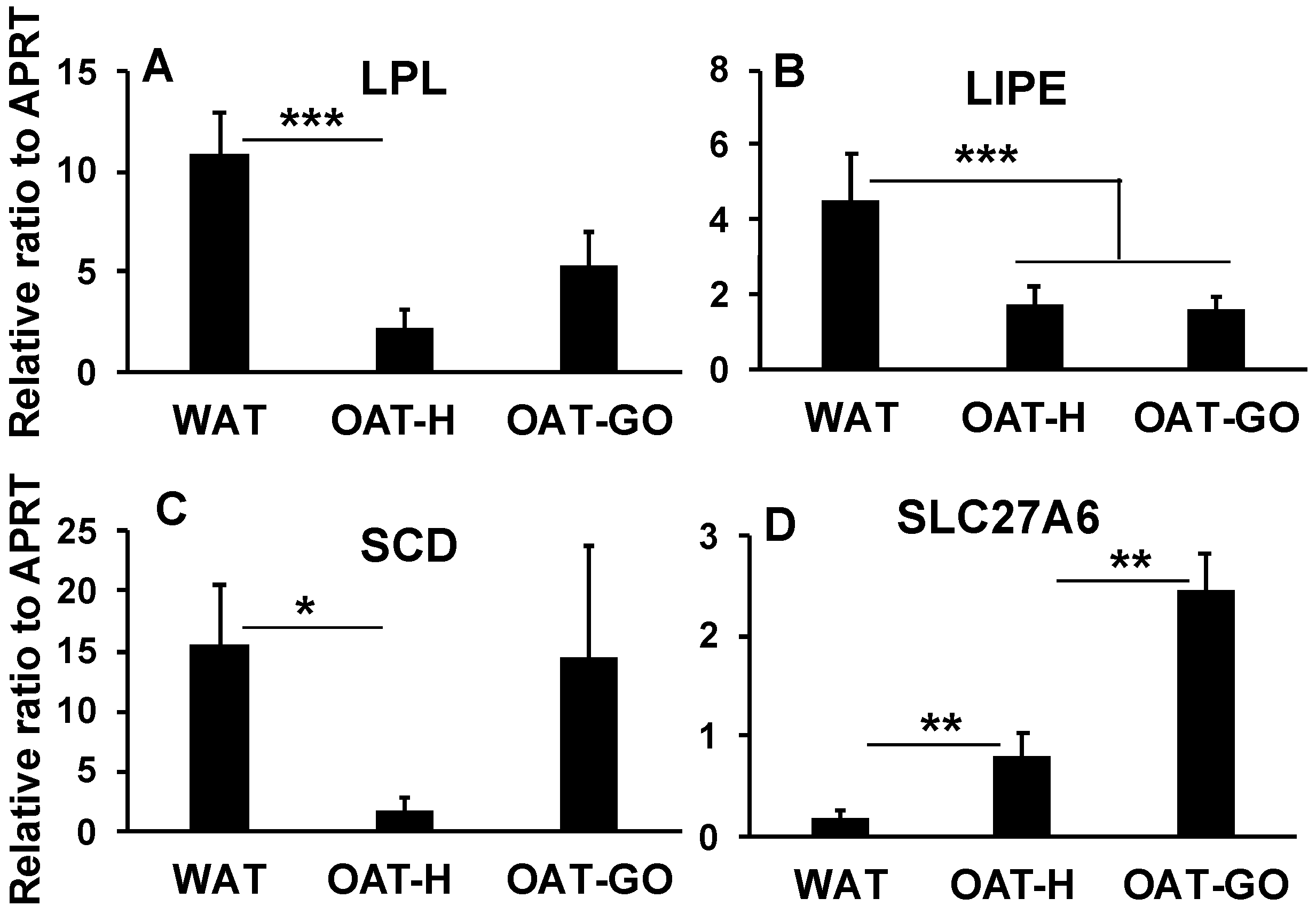
- Targeted QPCR analysis of genes highlighted in the whole transcriptome analysis confirmed substantially lower levels of elements of lipid metabolism including lipid uptake (LPL), lipolysis (LIPE) and uFAs conversion (SCD), and substantially up-regulated SLC27A6 (FA transporter) in OAT-H, when compared with WAT (Figure 5A–D). SLC27A6 has been reported as a transporter for long chain FAs uptake (>14 carbons) [27,28].In OAT-GO samples, LIPE transcripts (lipolysis) remained low, while variable LPL (lipid uptake) and SCD (uFAs conversion) expression levels were detected, not significantly different compared to WAT by QPCR analysis (Figure 5A–C). The lack of significant difference of FASN (fatty acid synthase) mRNA transcripts between OAT-H, OAT-GO and WAT was confirmed in QPCR analysis (Figure S1), as well as the further increase in SLC27A6 expression in OAT-GO compared to OAT (Figure 5D).
2.5. Signatures of Biological and Signaling Pathways in OAT by IPA® Analysis
- Substantially lower expression of the Homeobox (HOX) families (A, B, C, D) were observed in OAT-H compared with WAT (sub-grouped in Table S1A). HOXA5, HOXA6, HOXC8 and T-box transcription factor 5 (TBX5) were among the top most lowly expressed genes in OAT-H and relate to embryonic development of WAT/BAT (highlighted in Table S1B) [9,10]. Upregulation in the Iroquois homeobox gene family (IRX3 and IRX5), which play a role in an early step of neural and adipose tissue development, was observed in OAT-H [29,30]. The top ten up-regulated genes in OAT-H are shown in Table S1B.
- In IPA®, in addition to the changes in lipid metabolisms detailed above, substantially lower activity (overlap-p < 0.01; z-score < −2) was seen in pathways of cell movement and molecule transport in OAT-H compared with WAT (Table S2). In agreement with this, signaling pathways showed substantially lower activity in OAT-H of oxidative phosphorylation (OXPHOS), glycolysis, gluconeogenesis, glycogen degradation and cAMP-mediated signaling (Table S3). Furthermore, IPA® of down-regulated genes further supported the observed lower metabolic activity and signaling pathways of OAT-H compared with WAT (Supplementary Materials Tables S2 and S3).
- Certain signaling pathways were substantially activated (overlap-p < 0.01; z-score > 2, IPA® analysis) in OAT-H compared with WAT. (A) Activation of Wnt/Ca+ signaling pathway [31] was observed, as indicated by 11 up-regulated genes including Wnt family members (Wnt5A and Wnt5B) (Table S3). (B) Components of fibroblast growth factor (FGF) signaling [32] were significantly increased in OAT-H compared to WAT including FGF family and FGF-receptor 2 (FGFR2) (Table S3). Furthermore, we observed significantly increased expression of IGF2, IGF1R, IGF-binding proteins (IGFBP4&6, IGF2BP2) but down-regulated IGF1, IGFBP3 and Akt1 expression [33] (Table S1A). (C) Activation of Sirtuin signaling pathway [34] was observed as indicated by 11 up- and 64 down-regulated genes in IPA® analysis (Table S3).
- Additional up-regulated genes in OAT-H compared to WAT included expression of molecules (e.g., neuronal biomarkers, NRGN [35] and SNAP25 [36]) associated with neurogenesis (see Table S2) including development of neurons, proliferation of neuronal cells, differentiation of neurons, outgrowth of neuritis, neurotransmission and neuritogenesis.
- Up- and down-regulated expression of growth factors of interest in OAT-H compared with WAT are summarized in Table 1. The up-regulated (range of ~2 to 45-fold, Table S4) growth factors interfere with signaling pathways including TGFβ/Wnt/Ca+/FGF/Notch/MAPK/PI3K, potentially relevant to a wide range of biological processes of OAT [31,37,38].
3. Discussion
3.1. OAT Is Distinct from WAT and BAT/BRITE
3.2. Expression of a Distinct FA Transporter in OAT
3.3. OAT Is not Implicated in Maintenance of Energy Balance
3.4. RNAseq Data Provide Additional Insight into Role of IGF-1 Signaling in GO
3.5. Hypothesis of Dysfunction of Mitochondria in OAT Expansion in GO
3.6. Weaknesses and Strengths
3.7. Speculations from Current Study
3.8. Clinical Impact from Current Study
3.9. Conclusions
4. Materials and Methods
4.1. Adipose Tissue Collection and Preparation
4.2. Adipocytes Analysis in Ex Vivo Adipose Tissues by Light Microscopy
4.3. Electronic Microscopy Analysis of Adipocytes and Mitochondria in Ex Vivo Tissues
4.4. QPCR for Markers of Adipose Tissues of WAT, BAT/BRITE and OAT
4.5. RNA-Seq Sample Preparation and Sequencing
4.6. RNA-seq Read Alignment and Differential Expression Analysis
4.7. Ingenuity® Pathway Analysis (IPA®, Qiagen)
4.8. Gas Chromatography (GC) Analysis of Triglyceride FAs
4.9. Statistical and Bioinformatics Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GO | Graves’ Orbitopathy |
| OAT | Orbital adipose tissue |
| WAT | White adipose tissue |
| WAT/BRITE | Brown adipose tissue/BRown in white |
| PFs | preadipocytes/fibroblasts |
| FA | fatty acid |
| LEP | Leptin |
| LPL | lipoprotein lipase |
| LIPE | hormone-sensitive lipase |
| SCD | stearoyl-CoA desaturase |
| FASN | fatty acid synthase |
| UCP1 | uncoupling protein 1 |
| MFN2 | mitofusin 2 |
| TSHR | thyrotropin receptor |
| IGF | insulin like growth factor |
| TGFβ | transforming growth factor betta |
| FGF | fibroblast growth factor |
| NRGN | neurogranin |
| SNAP25 | synaptosomal-associated protein |
| MSC | mesenchymal stem cell |
References
- Bahn, R.S. Graves’ ophthalmopathy. N. Engl. J. Med. 2010, 362, 726–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, P.N.; Zhang, L.; Lee, R.W.J.; Muller, I.; Ezra, D.G.; Dayan, C.M.; Kahaly, G.J.; Ludgate, M. New insights into the pathogenesis and nonsurgical management of Graves orbitopathy. Nat. Rev. Endocrinol. 2020, 16, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Wolfram-Gabel, R.; Kahn, J.L. Adipose body of the orbit. Clin. Anat. 2002, 15, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Crisp, M.S.; Lane, C.; Halliwell, M.; Wynford-Thomas, D.; Ludgate, M. Thyrotropin receptor transcripts in human adipose tissue. J. Clin. Endocrinol. Metab. 1997, 82, 2003–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, R.S.; Kahaly, G.J.; Patel, A.; Sile, S.; Thompson, E.H.Z.; Perdok, R.; Fleming, J.C.; Fowler, B.T.; Marcocci, C.; Marino, M.; et al. Teprotumumab for the Treatment of Active Thyroid Eye Disease. N. Engl. J. Med. 2020, 382, 341–352. [Google Scholar] [CrossRef]
- Krieger, C.C.; Neumann, S.; Place, R.F.; Marcus-Samuels, B.; Gershengorn, M.C. Bidirectional TSH and IGF-1 receptor cross-talk mediates stimulation of hyaluronan secretion by Graves’ disease immunoglobins. J. Clin. Endocrinol. Metab. 2014. [Google Scholar] [CrossRef]
- Smith, T.J.; Kahaly, G.J.; Ezra, D.G.; Fleming, J.C.; Dailey, R.A.; Tang, R.A.; Harris, G.J.; Antonelli, A.; Salvi, M.; Goldberg, R.A.; et al. Teprotumumab for Thyroid-Associated Ophthalmopathy. N. Engl. J. Med. 2017, 376, 1748–1761. [Google Scholar] [CrossRef]
- Zhang, L.; Grennan-Jones, F.; Draman, M.S.; Lane, C.; Morris, D.; Dayan, C.M.; Tee, A.R.; Ludgate, M. Possible targets for nonimmunosuppressive therapy of Graves’ orbitopathy. J. Clin. Endocrinol. Metab. 2014, 99, E1183–E1190. [Google Scholar] [CrossRef]
- Billon, N.; Dani, C. Developmental origins of the adipocyte lineage: New insights from genetics and genomics studies. Stem. Cell Rev. 2012, 8, 55–66. [Google Scholar] [CrossRef]
- Gesta, S.; Tseng, Y.H.; Kahn, C.R. Developmental origin of fat: Tracking obesity to its source. Cell 2007, 131, 242–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolders, M.H.; Graniewski-Wijnands, H.S.; Meinders, A.E.; Fogteloo, A.J.; Pijl, H.; de Keizer, R.J. Exophthalmos in obesity. Ophthalmic. Res. 2004, 36, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Coenen, M.J.; Scherer, P.E.; Bahn, R.S. Evidence for enhanced adipogenesis in the orbits of patients with Graves’ ophthalmopathy. J. Clin. Endocrinol. Metab. 2004, 89, 930–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeGroot, L.J. Graves’ Disease and the Manifestations of Thyrotoxicosis. In Endotext [internet]; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., Dungan, K., Grossman, A., Hershman, J.M., Kaltsas, G., Koch, C., Kopp, P., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar] [PubMed]
- Zhang, L.; Grennan-Jones, F.; Lane, C.; Rees, D.A.; Dayan, C.M.; Ludgate, M. Adipose tissue depot-specific differences in the regulation of hyaluronan production of relevance to Graves’ orbitopathy. J. Clin. Endocrinol. Metab. 2012, 97, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ji, Q.H.; Ruge, F.; Lane, C.; Morris, D.; Tee, A.R.; Dayan, C.M.; Ludgate, M. Reversal of Pathological Features of Graves’ Orbitopathy by Activation of Forkhead Transcription Factors, FOXOs. J. Clin. Endocrinol. Metab. 2016, 101, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Coelho, M.; Oliveira, T.; Fernandes, R. Biochemistry of adipose tissue: An endocrine organ. Arch. Med. Sci. 2013, 9, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Spiegelman, B.M. Adipocytes as regulators of energy balance and glucose homeostasis. Nature 2006, 444, 847–853. [Google Scholar] [CrossRef] [Green Version]
- Benador, I.Y.; Veliova, M.; Mahdaviani, K.; Petcherski, A.; Wikstrom, J.D.; Assali, E.A.; Acin-Perez, R.; Shum, M.; Oliveira, M.F.; Cinti, S.; et al. Mitochondria Bound to Lipid Droplets Have Unique Bioenergetics, Composition, and Dynamics that Support Lipid Droplet Expansion. Cell. Metab. 2018, 27, 869–885.e6. [Google Scholar] [CrossRef] [Green Version]
- Draman, M.S.; Stechman, M.; Scott-Coombes, D.; Dayan, C.M.; Rees, D.A.; Ludgate, M.; Zhang, L. The Role of Thyrotropin Receptor Activation in Adipogenesis and Modulation of Fat Phenotype. Front. Endocrinol. (Lausanne) 2017, 8, 83. [Google Scholar] [CrossRef] [Green Version]
- Jespersen, N.Z.; Larsen, T.J.; Peijs, L.; Daugaard, S.; Homoe, P.; Loft, A.; de Jong, J.; Mathur, N.; Cannon, B.; Nedergaard, J.; et al. A classical brown adipose tissue mRNA signature partly overlaps with brite in the supraclavicular region of adult humans. Cell Metab. 2013, 17, 798–805. [Google Scholar] [CrossRef] [Green Version]
- Bukowiecki, L.; Collet, A.J.; Follea, N.; Guay, G.; Jahjah, L. Brown adipose tissue hyperplasia: A fundamental mechanism of adaptation to cold and hyperphagia. Am. J. Physiol. 1982, 242, E353–E539. [Google Scholar] [CrossRef]
- Karpe, F.; Pinnick, K.E. Biology of upper-body and lower-body adipose tissue—link to whole-body phenotypes. Nat. Rev. Endocrinol. 2015, 11, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Combs, T.P.; Pajvani, U.B.; Berg, A.H.; Lin, Y.; Jelicks, L.A.; Laplante, M.; Nawrocki, A.R.; Rajala, M.W.; Parlow, A.F.; Cheeseboro, L.; et al. A transgenic mouse with a deletion in the collagenous domain of adiponectin displays elevated circulating adiponectin and improved insulin sensitivity. Endocrinology 2004, 145, 367–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schluter, A.; Horstmann, M.; Diaz-Cano, S.; Plohn, S.; Stahr, K.; Mattheis, S.; Oeverhaus, M.; Lang, S.; Flogel, U.; Berchner-Pfannschmidt, U.; et al. Genetic immunization with mouse thyrotrophin hormone receptor plasmid breaks self-tolerance for a murine model of autoimmune thyroid disease and Graves’ orbitopathy. Clin. Exp. Immunol. 2018, 191, 255–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Baker, G.; Janus, D.; Paddon, C.A.; Fuhrer, D.; Ludgate, M. Biological effects of thyrotropin receptor activation on human orbital preadipocytes. Investig. Ophthalmol. Vis. Sci. 2006, 47, 5197–5203. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Gimeno, R.E.; Ortegon, A.M.; Patel, S.; Punreddy, S.; Ge, P.; Sun, Y.; Lodish, H.F.; Stahl, A. Characterization of a heart-specific fatty acid transport protein. J. Biol. Chem. 2003, 278, 16039–16044. [Google Scholar] [CrossRef] [Green Version]
- Yen, M.C.; Chou, S.K.; Kan, J.Y.; Kuo, P.L.; Hou, M.F.; Hsu, Y.L. New Insight on Solute Carrier Family 27 Member 6 (SLC27A6) in Tumoral and Non-Tumoral Breast Cells. Int. J. Med. Sci. 2019, 16, 366–375. [Google Scholar] [CrossRef] [Green Version]
- Bosse, A.; Zulch, A.; Becker, M.B.; Torres, M.; Gomez-Skarmeta, J.L.; Modolell, J.; Gruss, P. Identification of the vertebrate Iroquois homeobox gene family with overlapping expression during early development of the nervous system. Mech. Dev. 1997, 69, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Claussnitzer, M.; Dankel, S.N.; Kim, K.H.; Quon, G.; Meuleman, W.; Haugen, C.; Glunk, V.; Sousa, I.S.; Beaudry, J.L.; Puviindran, V.; et al. FTO Obesity Variant Circuitry and Adipocyte Browning in Humans. N. Engl. J. Med. 2015, 373, 895–907. [Google Scholar] [CrossRef] [Green Version]
- De, A. Wnt/Ca2+ signaling pathway: A brief overview. Acta Biochim. Biophys. Sin. (Shanghai) 2011, 43, 745–756. [Google Scholar] [CrossRef]
- Schrijver, B.; Kooiman, M.A.; Kasteleijn, E.; van Holten-Neelen, C.; Virakul, S.; Paridaens, D.; Peeters, R.P.; van Hagen, P.M.; Dalm, V.; Dik, W.A. Basic Fibroblast Growth Factor Induces Adipogenesis in Orbital Fibroblasts: Implications for the Pathogenesis of Graves’ Orbitopathy. Thyroid 2019, 29, 395–404. [Google Scholar] [CrossRef]
- Bendall, S.C.; Stewart, M.H.; Menendez, P.; George, D.; Vijayaragavan, K.; Werbowetski-Ogilvie, T.; Ramos-Mejia, V.; Rouleau, A.; Yang, J.; Bosse, M.; et al. IGF and FGF cooperatively establish the regulatory stem cell niche of pluripotent human cells in vitro. Nature 2007, 448, 1015–1021. [Google Scholar] [CrossRef]
- Verdin, E.; Hirschey, M.D.; Finley, L.W.; Haigis, M.C. Sirtuin regulation of mitochondria: Energy production, apoptosis, and signaling. Trends Biochem. Sci. 2010, 35, 669–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peacock, W., IV; Van Meter, T.E.; Mirshahi, N.; Ferber, K.; Gerwien, R.; Rao, V.; Sair, H.I.; Diaz-Arrastia, R.; Korley, F.K. Derivation of a Three Biomarker Panel to Improve Diagnosis in Patients with Mild Traumatic Brain Injury. Front. Neurol. 2017, 8, 641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmalm, A.; Brinkmalm, G.; Honer, W.G.; Frolich, L.; Hausner, L.; Minthon, L.; Hansson, O.; Wallin, A.; Zetterberg, H.; Blennow, K.; et al. SNAP-25 is a promising novel cerebrospinal fluid biomarker for synapse degeneration in Alzheimer’s disease. Mol. Neurodegener. 2014, 9, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-beta and the TGF-beta Family: Context-Dependent Roles in Cell and Tissue Physiology. Cold Spring Harb. Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, B.J.; Tripathi, R.C.; Livingston, A.M.; Borisuth, N.S. The role of growth factors in the embryogenesis and differentiation of the eye. Am. J. Anat. 1991, 192, 442–471. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Cheng, Y.; Su, G. A review of the multifunctionality of angiopoietin-like 4 in eye disease. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef]
- Tanaka, K.; Matsumoto, E.; Higashimaki, Y.; Katagiri, T.; Sugimoto, T.; Seino, S.; Kaji, H. Role of osteoglycin in the linkage between muscle and bone. J. Biol. Chem. 2012, 287, 11616–11628. [Google Scholar] [CrossRef] [Green Version]
- Pakvasa, M.; Alverdy, A.; Mostafa, S.; Wang, E.; Fu, L.; Li, A.; Oliveira, L.; Athiviraham, A.; Lee, M.J.; Wolf, J.M.; et al. Neural EGF-like protein 1 (NELL-1): Signaling crosstalk in mesenchymal stem cells and applications in regenerative medicine. Genes Dis. 2017, 4, 127–137. [Google Scholar] [CrossRef]
- Massague, J. TGFbeta signalling in context. Nat. Rev. Mol. Cell Biol. 2012, 13, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Fiuza, U.M.; Arias, A.M. Cell and molecular biology of Notch. J. Endocrinol. 2007, 194, 459–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Zhu, S.; Wu, M.; Han, W.; Yu, Y. Functional receptors and intracellular signal pathways of midkine (MK) and pleiotrophin (PTN). Biol. Pharm. Bull. 2014, 37, 511–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, J.; Harikumar, K.G.; Erice, C.; Chen, C.; Gu, X.; Wang, L.; Parker, N.; Cheng, Z.; Xu, W.; Williams, B.O.; et al. Structure and function of Norrin in assembly and activation of a Frizzled 4-Lrp5/6 complex. Genes Dev. 2013, 27, 2305–2319. [Google Scholar] [CrossRef] [Green Version]
- Kato, T. Biological roles of hepatocyte growth factor-Met signaling from genetically modified animals. Biomed. Rep. 2017, 7, 495–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouret, S.G. Neurodevelopmental actions of triglyceridetin. Brain Res. 2010, 1350, 2–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, R.B. Direct and indirect effects of triglyceridetin on adipocyte metabolism. Biochim. Biophys. Acta 2014, 1842, 414–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazak, L.; Chouchani, E.T.; Stavrovskaya, I.G.; Lu, G.Z.; Jedrychowski, M.P.; Egan, D.F.; Kumari, M.; Kong, X.; Erickson, B.K.; Szpyt, J.; et al. UCP1 deficiency causes brown fat respiratory chain depletion and sensitizes mitochondria to calcium overload-induced dysfunction. Proc. Natl. Acad. Sci. USA 2017, 114, 7981–7986. [Google Scholar] [CrossRef] [Green Version]
- Kusminski, C.M.; Scherer, P.E. Mitochondrial dysfunction in white adipose tissue. Trends Endocrinol. Metab. 2012, 23, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Sawyer, S.L.; Cheuk-Him Ng, A.; Innes, A.M.; Wagner, J.D.; Dyment, D.A.; Tetreault, M.; Care4Rare Canada, C.; Majewski, J.; Boycott, K.M.; Screaton, R.A.; et al. Homozygous mutations in MFN2 cause multiple symmetric lipomatosis associated with neuropathy. Hum. Mol. Genet. 2015, 24, 5109–5114. [Google Scholar] [CrossRef] [Green Version]
- Bartness, T.J.; Liu, Y.; Shrestha, Y.B.; Ryu, V. Neural innervation of white adipose tissue and the control of lipolysis. Front Neuroendocrinol. 2014, 35, 473–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaszkiewicz, M.; Willows, J.W.; Johnson, C.P.; Townsend, K.L. The Importance of Peripheral Nerves in Adipose Tissue for the Regulation of Energy Balance. Biology 2019, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozdon, K.; Fitchett, C.; Rose, G.E.; Ezra, D.G.; Bailly, M. Mesenchymal Stem Cell-Like Properties of Orbital Fibroblasts in Graves’ Orbitopathy. Investig. Ophthalmol. Vis. Sci. 2015, 56, 5743–5750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotte, G.J.; Schaafsma, P.J.; Botha, C.P.; Wielopolski, P.A.; Simonsz, H.J. Visualization of Sliding and Deformation of Orbital Fat During Eye Rotation. Trans. Vis. Sci. Technol. 2016, 5, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draman, M.S.; Grennan-Jones, F.; Zhang, L.; Taylor, P.N.; Tun, T.K.; McDermott, J.; Moriarty, P.; Morris, D.; Lane, C.; Sreenan, S.; et al. Effects of prostaglandin F(2alpha) on adipocyte biology relevant to graves’ orbitopathy. Thyroid 2013, 23, 1600–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer zu Horste, M.; Stroher, E.; Berchner-Pfannschmidt, U.; Schmitz-Spanke, S.; Pink, M.; Gothert, J.R.; Fischer, J.W.; Gulbins, E.; Eckstein, A.K. A novel mechanism involved in the pathogenesis of Graves ophthalmopathy (GO): Clathrin is a possible targeting molecule for inhibiting local immune response in the orbit. J. Clin. Endocrinol. Metab. 2011, 96, E1727–E1736. [Google Scholar] [CrossRef] [Green Version]
- Ortigosa, J.L.; Mendoza, F.; Argote, R.M.; Garcia, G.; Cervantes, C.; Parra, A. Propranolol Effect on Plasma Glucose, Free Fatty-Acid, Insulin, and Growth-Hormone in Graves-Disease. Metab. Clin. Exp. 1976, 25, 1201–1207. [Google Scholar] [CrossRef]
- Gagnon, A.; Antunes, T.T.; Ly, T.; Pongsuwan, P.; Gavin, C.; Lochnan, H.A.; Sorisky, A. Thyroid-stimulating hormone stimulates lipolysis in adipocytes in culture and raises serum free fatty acid levels in vivo. Metabolism 2010, 59, 547–553. [Google Scholar] [CrossRef]
- Haluzik, M.; Nedvidkova, J.; Bartak, V.; Dostalova, I.; Vlcek, P.; Racek, P.; Taus, M.; Svacina, S.; Alesci, S.; Pacak, K. Effects of hypo- and hyperthyroidism on noradrenergic activity and glycerol concentrations in human subcutaneous abdominal adipose tissue assessed with microdialysis. J. Clin. Endocrinol. Metab. 2003, 88, 5605–5608. [Google Scholar] [CrossRef]
- Marcus, C.; Ehren, H.; Bolme, P.; Arner, P. Regulation of lipolysis during the neonatal period. Importance of thyrotropin. J. Clin. Investig. 1988, 82, 1793–1797. [Google Scholar]
- Rodbell, M. Metabolism of Isolated Fat Cells. I. Effects of Hormones on Glucose Metabolism and Lipolysis. J. Biol. Chem. 1964, 239, 375–380. [Google Scholar]
- Stein, J.D.; Childers, D.; Gupta, S.; Talwar, N.; Nan, B.; Lee, B.J.; Smith, T.J.; Douglas, R. Risk factors for developing thyroid-associated ophthalmopathy among individuals with Graves disease. JAMA Ophthalmol. 2015, 133, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar, A.; Simental-Mendia, L.E.; Pedone, C.; Ferretti, G.; Nachtigal, P.; Bo, S.; Derosa, G.; Maffioli, P.; Watts, G.F. Statin therapy and plasma free fatty acids: A systematic review and meta-analysis of controlled clinical trials. Br. J. Clin. Pharmacol. 2016, 81, 807–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kershbaum, A.; Bellet, S.; Caplan, R.F.; Feinberg, L.J. Effect of Cigarette Smoking on Free Fatty Acids in Patients with Myocardial Infarction. Circulation 1961, 24, 970. [Google Scholar] [CrossRef]
- Prummel, M.F.; Wiersinga, W.M. Smoking and Risk of Graves-Disease. JAMA-J. Am. Med. Assoc. 1993, 269, 479–482. [Google Scholar] [CrossRef]
- Bartalena, L.; Baldeschi, L.; Boboridis, K.; Eckstein, A.; Kahaly, G.J.; Marcocci, C.; Perros, P.; Salvi, M.; Wiersinga, W.M.; European Group on Graves’ Orbitopathy (EUGOGO). The 2016 European Thyroid Association/European Group on Graves’ Orbitopathy Guidelines for the Management of Graves’ Orbitopathy. Eur. Thyroid J. 2016, 5, 9–26. [Google Scholar] [CrossRef] [Green Version]
- Newman, G.R.; Hobot, J.A. Resin Microscopy and On-Section Immunocytochemistry; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar] [CrossRef]
- Zhang, L.; Bowen, T.; Grennan-Jones, F.; Paddon, C.; Giles, P.; Webber, J.; Steadman, R.; Ludgate, M. Thyrotropin receptor activation increases hyaluronan production in preadipocyte fibroblasts: Contributory role in hyaluronan accumulation in thyroid dysfunction. J. Biol. Chem. 2009, 284, 26447–26455. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Frankish, A.; Diekhans, M.; Ferreira, A.M.; Johnson, R.; Jungreis, I.; Loveland, J.; Mudge, J.M.; Sisu, C.; Wright, J.; Armstrong, J.; et al. GENCODE reference annotation for the human and mouse genomes. Nucleic Acids Res. 2019, 47, D766–D773. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Gingeras, T.R. Mapping RNA-seq Reads with STAR. Curr. Protoc. Bioinf. 2015, 51. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorcevic, M.; Hilton, C.; McNeil, C.; Christodoulides, C.; Hodson, L.; Karpe, F.; Pinnick, K.E. A cellular model for the investigation of depot specific human adipocyte biology. Adipocyte 2017, 6, 40–55. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Up-Regulated Growth Factor (Multi-Function 1) | Known Function/Pathway |
|---|---|
| Angiopoietin (ANGPT4) [39] | angiogenesis |
| Osteoglycin (OGN) [40] | bone formation |
| Neural EGF-like protein (NELL1) [41] | bone formation |
| TGFβ family 1 [42] (TGFB2, BMP3, GDF-6,7,11) and Latent TGFβ binding protein 1 (LTBP4) | TGFβ signaling pathway |
| Jagged 1 protein (JAG1) 1 [43] | Notch signaling pathway |
| Midkine (MDK) 1, Pleiotrophin (PTN) 1 [44] | multi-signaling pathway |
| Norrin cystine knot growth factor (NDP) 1 [45] | Wnt-signaling pathway |
| Hepatocyte growth factor (HGF) 1 [46] | HGF/Met signaling pathway |
| FGF-(7,9,10,17&18)1 via FGFR [32] | multi-signaling pathway |
| IGF21 via IGF1R/IGF2R [33] | multi-signaling pathway |
| Down-regulated growth factors as below | |
| TGFA, BMP6, GDF10, IGF1, FGF-2,8&13, VEGF-B&D, GMFG, JAG2, LEFTY2, LEP and PDGF-B&C | |
| OAT-H | OAT-GO | WAT | BAT/BRITE [17,20] | |
|---|---|---|---|---|
| Hypertrophy (Figure 1) | - | - | +++ | - |
| LEP (WAT marker) (Figure 2) | --- | --- | +++ | --- |
| Hyperplasia (Figure 1) | +++ | +++ | - | +++ |
| Lipid vacuole (Figure 1) | - | - | - | +++ |
| Mt number/adipocyte (Figure 1) | - | --- | - | +++ |
| UCP1 (BAT marker) (Figure 2) | --- | - | - | +++ |
| MFN2 (Mt marker) (Figure 2) | --- | - | - | +++ |
| LIPE (Lipolysis, Figure 5) | - | - | +++ | |
| LPL (FA uptake, Figure 5) | - | +++ | +++ | |
| SCD (uFAs conversion, Figure 5) | --- | - | - | |
| FASN (FA synthesis, Figure S1) | - | - | - | |
| Linoleic acid (essential FA, Figure 3) | +++ | +++ | +++ | |
| de novo lipogenesis (SCD, Linoleic acid and FASN) | - | - | - | |
| Stearic acid (Figure 3) | +++ | +++ | - | |
| SLC27A6 (transporter) (Figure 5) | +++ | +++ | - |
| ID | Sex | Age | GO Status | Analysis |
|---|---|---|---|---|
| OAT-H1 | F | 51 | no | RNAseq(1.06) |
| OAT-H2 | F | 64 | no | RNAseq(0.97) |
| OAT-H3 | F | 64 | no | RNAseq(0.87)/Lipidomic/adipocyte |
| OAT-H4 | F | 76 | no | RNAseq(1.09)/adipocyte |
| OAT-H5 | F | 37 | no | Lipidomic |
| OAT-H6 | F | 46 | no | Lipidomic |
| OAT-GO1 | F | 47 | active | Lipidomic/QPCR/adipocyte |
| OAT-GO2 | M | 49 | active | QPCR/adipocyte |
| OAT-GO3 | M | 78 | active | QPCR |
| OAT-GO4 | F | 16 | inactive | Lipidomic/QPCR |
| OAT-GO5 | F | 56 | inactive | Lipidomic/QPCR |
| OAT-GO6 | M | 45 | inactive | QPCR |
| OAT-GO7 | M | 60 | inactive | adipocyte |
| OAT-GO8 | F | 55 | inactive | adipocyte |
| OAT-GO9 | M | 29 | inactive | adipocyte/PCR |
| OAT-GO10 | F | 47 | inactive | QPCR |
| WAT1 | F | 72 | no | RNAseq(1.02)/QPCR |
| WAT2 | F | 72 | no | RNAseq(0.97)/QPCR |
| WAT3 | F | 44 | no | RNAseq(1.00)/QPCR |
| WAT4 | F | 62 | no | RNAseq(1.22)/QPCR |
| WAT5 | F | 50 | no | RNAseq(0.95)/QPCR |
| WAT6 | F | 37 | no | Lipidomic/QPCR/adipocyte |
| WAT7 | F | 64 | no | Lipidomic/QPCR/adipocyte |
| WAT8 | F | 46 | no | Lipidomic |
| WAT9 | M | 49 | no | QPCR |
| WAT10 | M | 54 | no | QPCR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Evans, A.; von Ruhland, C.; Draman, M.S.; Edkins, S.; Vincent, A.E.; Berlinguer-Palmini, R.; Rees, D.A.; Haridas, A.S.; Morris, D.; et al. Distinctive Features of Orbital Adipose Tissue (OAT) in Graves’ Orbitopathy. Int. J. Mol. Sci. 2020, 21, 9145. https://doi.org/10.3390/ijms21239145
Zhang L, Evans A, von Ruhland C, Draman MS, Edkins S, Vincent AE, Berlinguer-Palmini R, Rees DA, Haridas AS, Morris D, et al. Distinctive Features of Orbital Adipose Tissue (OAT) in Graves’ Orbitopathy. International Journal of Molecular Sciences. 2020; 21(23):9145. https://doi.org/10.3390/ijms21239145
Chicago/Turabian StyleZhang, Lei, Anna Evans, Chris von Ruhland, Mohd Shazli Draman, Sarah Edkins, Amy E. Vincent, Rolando Berlinguer-Palmini, D. Aled Rees, Anjana S Haridas, Dan Morris, and et al. 2020. "Distinctive Features of Orbital Adipose Tissue (OAT) in Graves’ Orbitopathy" International Journal of Molecular Sciences 21, no. 23: 9145. https://doi.org/10.3390/ijms21239145