CaSBP11 Participates in the Defense Response of Pepper to Phytophthora capsici through Regulating the Expression of Defense-Related Genes


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. CaSBP11 Protein Localizes to the Nucleus
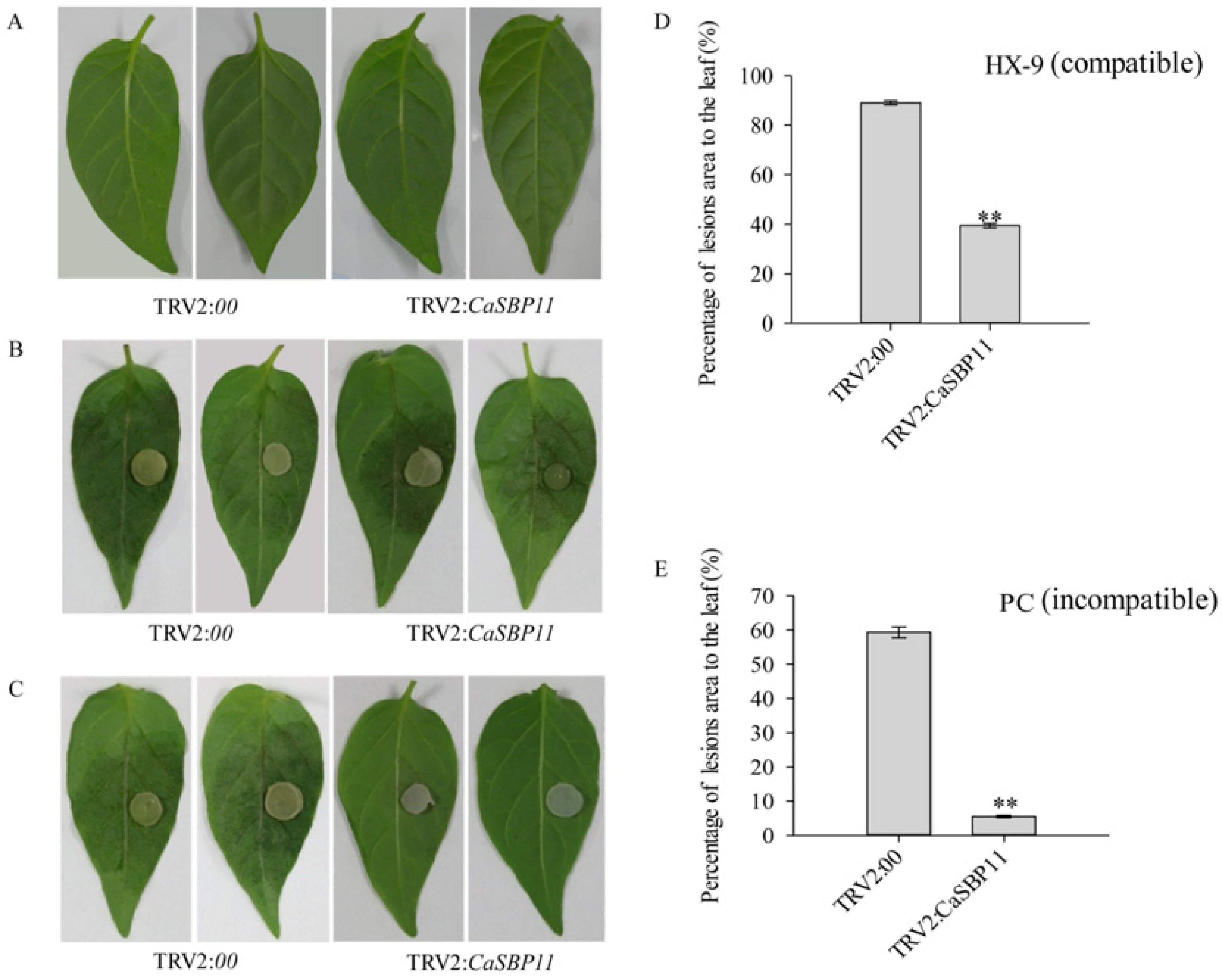
2.2. Silencing of CaSBP11 Enhanced Pepper Resistance to P. capsici Infection
2.3. CaSBP11 Is Involved in Pepper Defense Response to P. capsici through Inducing Defense-Related Genes
2.4. Transient Expression of CaSBP11 in Pepper
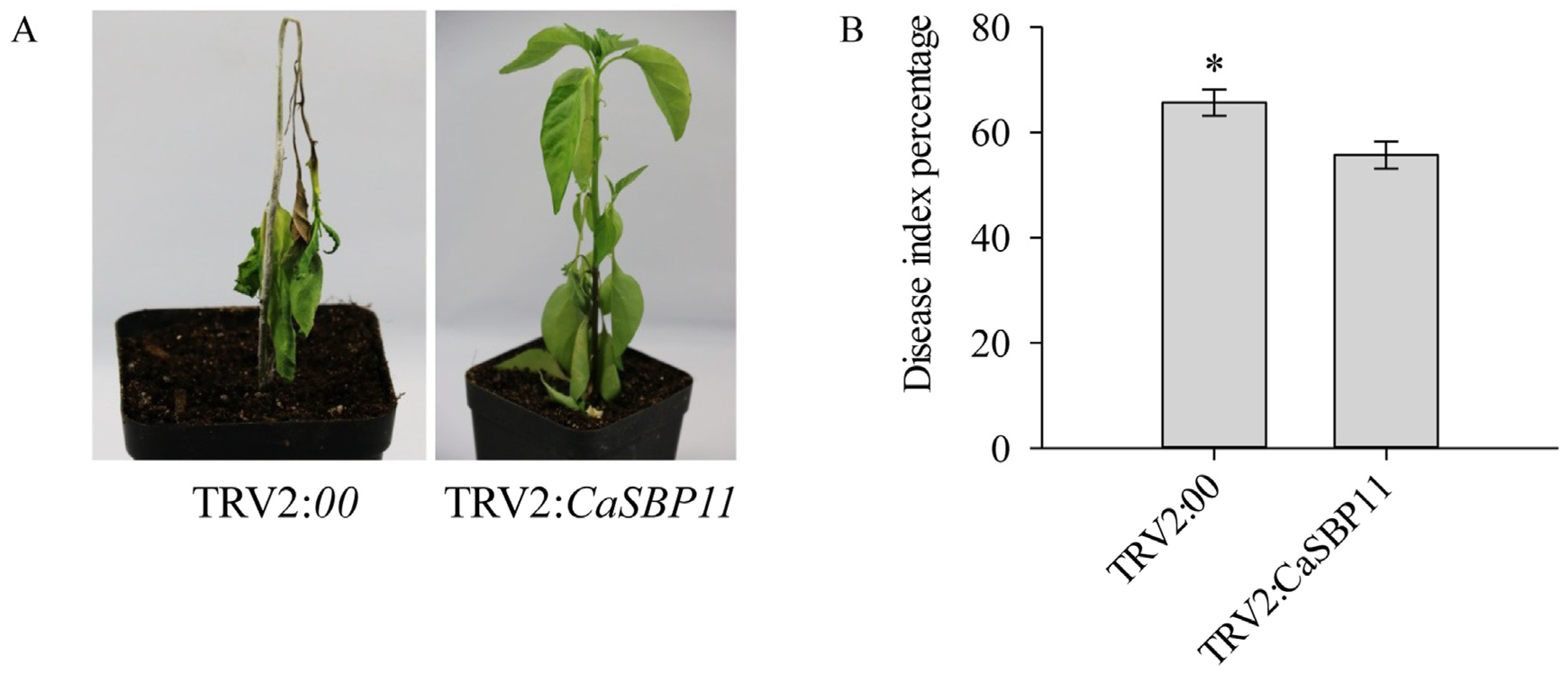
2.5. Overexpression of CaSBP11 in N. benthamiana Increased Susceptibility to P. capsici Infection
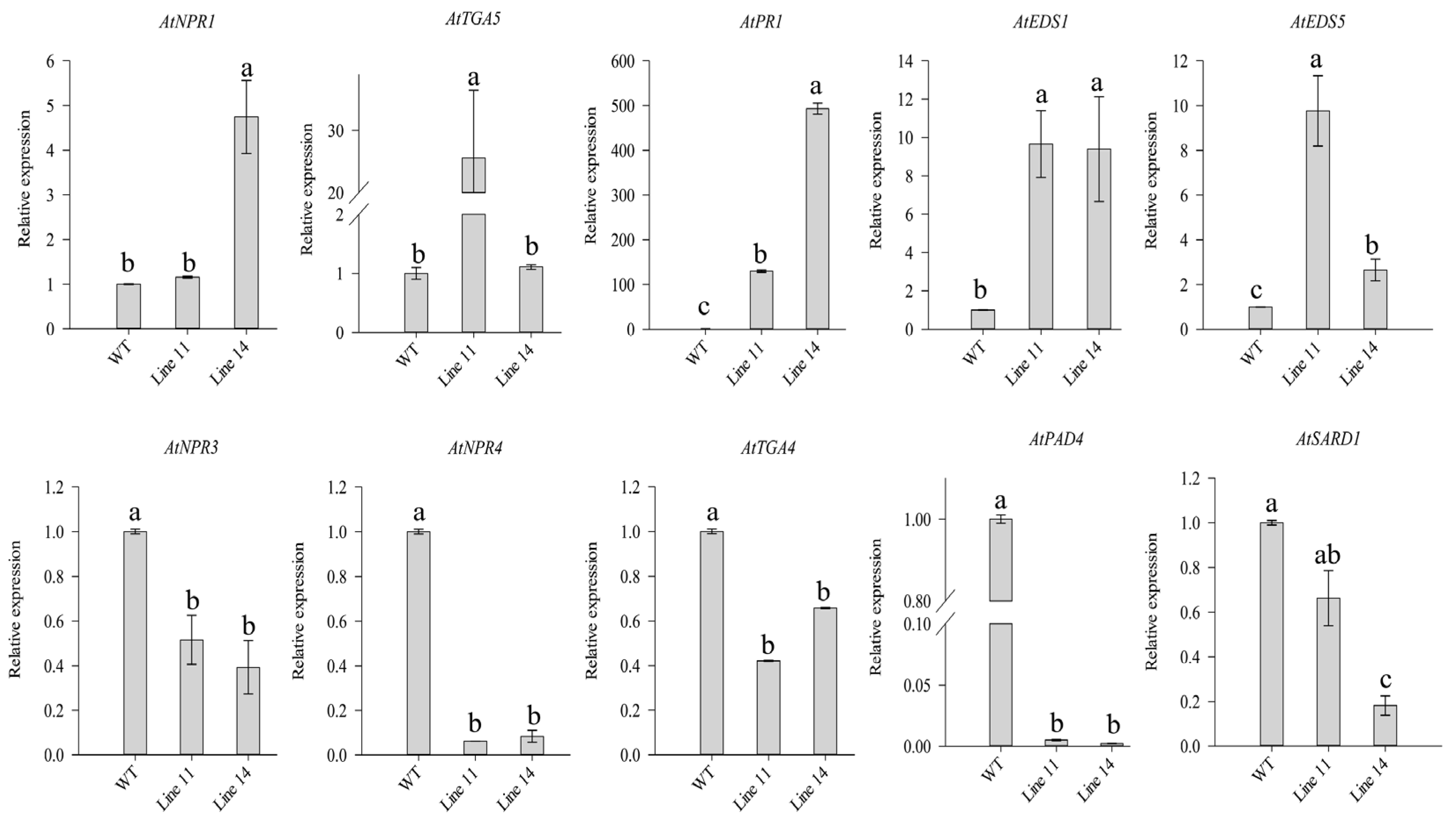
2.6. Overexpression of CaSBP11 in WT and Mutant Arabidopsis thaliana
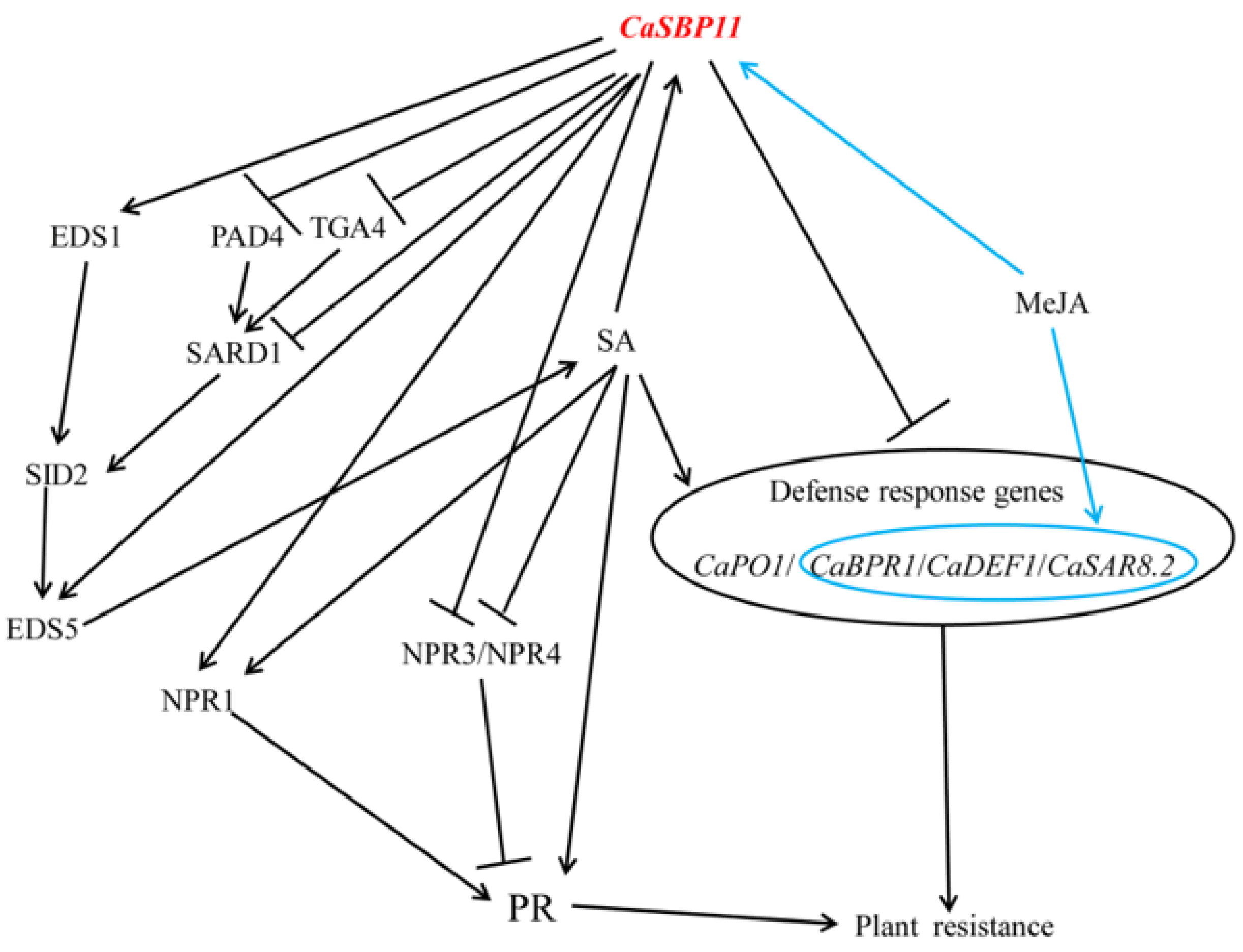
3. Discussion
4. Materials and Methods
4.1. Plant Material and Pathogen Preparation
4.2. Subcellular Localization of CaSBP11Protein in Pepper
4.3. Virus Induced Gene Silencing (VIGS) of CaSBP11 in Pepper
4.4. Transient Expression of CaSBP11 in Pepper
4.5. Overexpression of CaSBP11 in N. benthamiana
4.6. Overexpression of CaSBP11 in WT and Mutant Arabidopsis thaliana
4.7. Disease Index Percentage Statistics
4.8. RNA Extraction and Quantitative Real-Time PCR
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barchenger, D.W.; Lamour, K.H.; Bosland, P.W. Challenges and strategies for breeding resistance in Capsicum annuum to the multifarious pathogen. Phytophthora capsici. Front. Plant Sci. 2018, 9, 628. [Google Scholar] [CrossRef]
- Liu, K.K. Studies on the Resistance and Its Mechanisms of Pepper to Phytophthora Capsici; Institute of Horticulture, Northwest A&F University: Yangling, China, 2009. [Google Scholar]
- Hausbeck, M.K.; Lamour, K.H. Phytophthora capsici on vegetable crops: Research progress and management challenges. Plant Dis. 2004, 88, 1292–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granke, L.L.; Quesada-ocampo, L.; Hausbeck, M.K. Advances in research on Phytophthora capsici on vegetable crops in the United States. Plant Dis. 2012, 95, 1588–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.M.; Luo, D.X.; Khan, A.; Liu, K.K.; Arisha, M.H.; Zhang, H.X.; Cheng, G.X.; Ma, X.; Gong, Z.H. CanTF, a novel transcription factor in pepper, is involved in resistance to Phytophthora capsici as well as abiotic stresses. Plant Mol. Biol. Rep. 2018, 36, 776–789. [Google Scholar] [CrossRef]
- Ng, D.W.K.; Abeysinghe, J.K.; Kamali, M. Regulating the regulators: The control of transcription factors in plant defense signaling. Int. J. Mol. Sci. 2018, 19, 3737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, G.; Meng, X.; Liu, Y.; Zheng, Z.; Chen, Z.; Zhang, S. Phosphorylation of a WRKY transcription factor by two pathogen-responsive MAPKs drives phytoalexin biosynthesis in Arabidopsis. Plant Cell 2011, 23, 1639–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, Z. Potentiation of developmentally regulated plant defense response by AtWRKY18, a pathogen-induced Arabidopsis transcription factor. Plant Physiol. 2002, 129, 706–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Zhang, H.; Ali, M.; Wei, A.; Luo, D.; Gong, Z. The CaAP2/ERF064 regulates dual functions in pepper: Plant cell death and resistance to Phytophthora capsici. Genes 2019, 10, 541. [Google Scholar] [CrossRef] [Green Version]
- Klein, J.; Saedler, H.; Huijser, P. A new family of DNA binding proteins includes putative transcriptional regulators of the Antirrhinum majus floral meristem identity gene SQUAMOSA. Mol. Gen. Genet. 1996, 250, 7–16. [Google Scholar] [CrossRef]
- Yamasaki, K.; Kigawa, T.; Inoue, M.; Tateno, M.; Yamasaki, T.; Yabuki, T.; Aoki, M.; Seki, E.; Matsuda, T.; Nunokawa, E.; et al. A novel zinc-binding motif revealed by solution structures of DNA-binding domains of Arabidopsis SBP-family transcription factors. J. Mol. Biol. 2004, 337, 49–63. [Google Scholar] [CrossRef]
- Cardon, G.; Höhmann, S.; Klein, J.; Nettesheim, K.; Saedler, H.; Huijser, P. Molecular characterisation of the Arabidopsis SBP-box genes. Gene 1999, 237, 91–104. [Google Scholar] [CrossRef]
- Lännenpää, M.; Jänönen, I.; Hölttä-Vuori, M.; Gardemeister, M.; Porali, I.; Sopanen, T. A new SBP-box gene BpSPL1 in silver birch (Betula pendula). Physiol. Plant 2004, 120, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Poole, M.; Hong, Y.; Thompson, A.J.; King, G.J.; Manning, K.; To, M.; Giovannoni, J.J.; Seymour, G.B. A naturally occurring epigenetic mutation in a gene encoding an SBP-box transcription factor inhibits tomato fruit ripening. Nat Genet. 2006, 38, 948–952. [Google Scholar] [CrossRef]
- Li, J.; Hou, H.; Li, X.; Xiang, J.; Yin, X.; Gao, H.; Zheng, Y.; Bassett, C.L.; Wang, X. Genome-wide identification and analysis of the SBP-box family genes in apple (Malus×domestica Borkh.). Plant Physiol. Biochem. 2013, 70, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, B.; Zhao, D.; Li, C.; Shao, F.; Lu, S. Genome-wide analysis and molecular dissection of the SPL gene family in Salvia miltiorrhiza. J. Integr. Plant Biol. 2014, 56, 38–50. [Google Scholar] [CrossRef]
- Zhang, X.; Dou, L.; Pang, C.; Song, M.; Wei, H.; Fan, S.; Wang, C.; Yu, S. Genomic organization, differential expression, and functional analysis of the SPL gene family in Gossypium hirsutum. Mol. Genet Genom. 2015, 290, 115–126. [Google Scholar] [CrossRef]
- Zhang, H.-X.; Jin, J.-H.; He, Y.-M.; Lu, B.-Y.; Li, D.-W.; Chai, W.-G.; Khan, A.; Gong, Z.-H. Genome-wide identification and analysis of the SBP-Box family genes under Phytophthora capsici stress in pepper (Capsicum annuum L.). Front. Plant Sci. 2016, 7, 504. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Zhou, H.; Sheng, S.; Cao, M.; Li, Y.; Pang, X. Genome-wide organization and expression profiling of the SBP-box gene family in Chinese jujube (Ziziphus jujuba mill.). Int. J. Mol. Sci. 2017, 18, 1734. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Zhang, S.; Chen, F.; Liu, B.; Wu, L.; Li, F.; Zhang, J.; Bao, M.; Liu, G. Genome-wide identification and characterization of the SBP-box gene family in Petunia. BMC Genom. 2018, 19, 1–18. [Google Scholar] [CrossRef]
- Li, J.; Gao, X.; Sang, S.; Liu, C. Genome-wide identification, phylogeny, and expression analysis of the SBP-box gene family in Euphorbiaceae. BMC Genom. 2019, 20, 912. [Google Scholar] [CrossRef]
- Hou, H.M.; Li, J.; Gao, M.; Singer, S.D.; Wang, H.; Mao, L.Y.; Fei, Z.J.; Wang, X.P. Genomic organization, phylogenetic comparison and differential expression of the SBP-box family genes in grape. PLoS ONE 2013, 8, e59358. [Google Scholar] [CrossRef]
- Zhang, S.D.; Ling, L.Z. Genome-wide identification and evolutionary analysis of the SBP-box gene family in castor bean. PLoS ONE 2014, 9, e86688. [Google Scholar] [CrossRef]
- Ma, Y.; Guo, J.W.; Bade, R.; Men, Z.H.; Hasi, A. Genome-wide identification and phylogenetic analysis of the SBP-box gene family in melons. Genet. Mol. Res. 2014, 13, 8794–8806. [Google Scholar] [CrossRef]
- Xu, Z.; Sun, L.; Zhou, Y.; Yang, W.; Cheng, T.; Wang, J.; Zhang, Q. Identification and expression analysis of the SQUAMOSA promoter-binding protein (SBP)-box gene family in Prunus mume. Mol. Genet. Genom. 2015, 290, 1701–1715. [Google Scholar] [CrossRef]
- Wu, Z.; Cao, Y.; Yang, R.; Qi, T.; Hang, Y.; Lin, H.; Zhou, G.; Wang, Z.Y.; Fu, C. Switchgrass SBP-box transcription factors PvSPL1 and 2 function redundantly to initiate side tillers and affect biomass yield of energy crop. Biotechnol. Biofuels 2016, 9, 101. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Han, Z.; Li, J.; Qin, H.; Zhou, L.; Wang, Y.; Zhu, X.; Ma, Y.; Fang, W. Genome-wide analysis of the SBP-box gene family transcription factors and their responses to abiotic stresses in tea (Camellia sinensis). Genomics 2019, 112, 2194–2202. [Google Scholar] [CrossRef]
- Unte, U.S.; Sorensen, A.; Pesaresi, P.; Gandikota, M.; Leister, D.; Saedler, H.; Huijser, P. SPL8, an SBP-Box gene that affects pollen sac development in Arabidopsis. Plant Cell 2003, 15, 1009–1019. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Xu, W.; Liu, X.; Mao, X.; Li, A.; Wang, J.; Chang, X.; Zhang, X.; Jing, R. Functional conservation and divergence among homoeologs of TaSPL20 and TaSPL21, two SBP-Box genes governing yield-related traits in Hexaploid Wheat. Plant Physiol. 2017, 174, 1177–1191. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Gruber, M.Y.; Amyot, L.; Hannoufa, A. SPL13 regulates shoot branching and flowering time in Medicago sativa. Plant Mol. Biol. 2018, 96, 119–133. [Google Scholar] [CrossRef]
- Stone, J.M.; Liang, X.; Nekl, E.R.; Stiers, J.J. Arabidopsis AtSPL14, a plant-specific SBP-domain transcription factor, participates in plant development and sensitivity to fumonisin B1. Plant J. 2005, 1, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Padmanabhan, M.S.; Ma, S.; Burch-Smith, T.M.; Czymmek, K.; Huijser, P.; Dinesh-Kumar, S.P. Novel positive regulatory role for the SPL6 transcription factor in the N TIR-NB-LRR receptor-mediated plant innate immunity. PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef] [Green Version]
- Mao, Y.B.; Liu, Y.Q.; Chen, D.Y.; Chen, F.Y.; Fang, X.; Hong, G.J.; Wang, L.J.; Wang, J.W.; Chen, X.Y. Jasmonate response decay and defense metabolite accumulation contributes to age-regulated dynamics of plant insect resistance. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Jung, C.; Yeu, S.Y.; Koo, Y.J.; Kim, M.; Choi, Y.D.; Cheong, J.J. Transcript profile of transgenic Arabidopsis constitutively producing methyl jasmonate. J. Plant Biol. 2007, 50, 12–17. [Google Scholar] [CrossRef]
- Yao, S.; Yang, Z.; Yang, R.; Huang, Y.; Guo, G.; Kong, X.; Lan, Y.; Zhou, T.; Wang, H.; Wang, W.; et al. Transcriptional regulation of miR528 by OsSPL9 orchestrates antiviral response in rice. Mol. Plant 2019, 12, 1114–1122. [Google Scholar] [CrossRef]
- Hou, H.; Yan, Q.; Wang, X.; Xu, H. A SBP-Box gene VpSBP5 from chinese wild vitis species responds to Erysiphe necator and defense signaling molecules. Plant Mol. Biol. Rep. 2013, 31, 1261–1270. [Google Scholar] [CrossRef]
- Alonso, J.M.; Stepanova, A.N.; Leisse, T.J.; Kim, C.J.; Chen, H.; Shinn, P.; Stevenson, D.K.; Zimmerman, J.; Barajas, P.; Cheuk, R.; et al. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 2003, 301, 653–657. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Chung, E.H.; Hubert, D.A.; Tornero, P.; Dangl, J.L. Specific missense alleles of the Arabidopsis jasmonic acid co-receptor COI1 regulate innate immune receptor accumulation and function. PLoS Genet. 2012, 8, e1003018. [Google Scholar] [CrossRef] [Green Version]
- Hubert, D.A.; He, Y.; McNulty, B.C.; Tornero, P.; Dangl, J.L. Specific Arabidopsis HSP90.2 alleles recapitulate RAR1 cochaperone function in plant NB-LRR disease resistance protein regulation. Proc. Natl. Acad. Sci. USA 2009, 106, 9556–9563. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.W.; Kim, Y.J.; Hwang, B.K. The hypersensitive induced reaction and leucine-rich repeat proteins regulate plant cell death associated with disease and plant immunity. Mol. Plant Microbe Interact. 2011, 24, 68–78. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Hwang, B.K. Identification of the pepper SAR8.2 gene as a molecular marker for pathogen infection, abiotic elicitors and environmental stresses in Capsicum annuum. Planta 2003, 216, 387–396. [Google Scholar] [CrossRef]
- Kim, Y.J.; Hwang, B.K. Pepper gene encoding a basic pathogenesis-related 1 protein is pathogen and ethylene inducible. Physiol. Plant 2000, 108, 51–60. [Google Scholar] [CrossRef]
- Do, H.M.; Lee, S.C.; Jung, H.W.; Sohn, K.H.; Hwang, B.K. Differential expression and in situ localization of a pepper defensin (CADEF1) gene in response to pathogen infection, abiotic elicitors and environmental stresses in Capsicum annuum. Plant Sci. 2004, 166, 1297–1305. [Google Scholar] [CrossRef]
- Do, H.M.; Hong, J.K.; Jung, H.W.; Kim, S.H.; Ham, J.H.; Hwang, B.K. Expression of peroxidase-like genes, H2O2 production, and peroxidase activity during the hypersensitive response to Xanthomonas campestris pv. vesicatoria in Capsicum annuum. Mol. Plant Microbe Interact. 2003, 16, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.L. Defence Responses of Pepper (Capsicum Annuum L.) and Grafting Seedings to Phytophthora Capsici Leonia and the Function Analysis of CaRGA2 Gene; Institute of Horticulture, Northwest A&F University: Yangling, China, 2013. [Google Scholar]
- van Huijsduijnen, R.A.M.H.; Cornelissen, B.J.C.; van Loon, L.C.; van Boom, J.H.; Tromp, M.; Bol, J.F. Virus-induced synthesis of messenger RNAs for precursors of pathogenesis-related proteins in tobacco. EMBO J. 1985, 4, 2167–2171. [Google Scholar] [CrossRef]
- Cheol Song, G.; Sim, H.-J.; Kim, S.-G.; Ryu, C.-M. Root-mediated signal transmission of systemic acquired resistance against above-ground and below-ground pathogens. Ann. Bot. 2016, 118, 821–831. [Google Scholar] [CrossRef] [Green Version]
- Sohn, S.I.; Kim, Y.H.; Kim, B.R.; Lee, S.Y.; Lim, C.K.; Hur, J.H.; Lee, J.Y. Transgenic tobacco expressing the hrpNEP gene from Erwinia pyrifoliae triggers defense responses against Botrytis cinerea. Mol. Cells 2007, 24, 232–239. [Google Scholar] [CrossRef] [Green Version]
- Qiu, A.; Lei, Y.; Yang, S.; Wu, J.; Li, J.; Bao, B.; Cai, Y.; Wang, S.; Lin, J.; Wang, Y.; et al. CaC3H14 encoding a tandem CCCH zinc finger protein is directly targeted by CaWRKY40 and positively regulates the response of pepper to inoculation by Ralstonia solanacearum. Mol. Plant Pathol. 2018, 19, 2221–2235. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.A.N.; Bouwmeester, K.; Van de Mortel, J.E.; Shan, W.; Govers, F. A novel Arabidopsis–oomycete pathosystem: Differential interactions with Phytophthora capsici reveal a role for camalexin, indole glucosinolates and salicylic acid in defence. Plant Cell Environ. 2013, 1192–1203. [Google Scholar] [CrossRef]
- Wang, L.; Tsuda, K.; Truman, W.; Sato, M.; Nguyen, L.V.; Katagiri, F.; Glazebrook, J. CBP60g and SARD1 play partially redundant critical roles in salicylic acid signaling. Plant J. 2011, 67, 1029–1041. [Google Scholar] [CrossRef]
- Sun, T.; Busta, L.; Zhang, Q.; Ding, P.; Jetter, R.; Zhang, Y. TGACG-BINDING FACTOR 1 (TGA1) and TGA4 regulate salicylic acid and pipecolic acid biosynthesis by modulating the expression of SYSTEMIC ACQUIRED RESISTANCE DEFICIENT 1 (SARD1) and CALMODULIN-BINDING PROTEIN 60g (CBP60g). New Phytol. 2018, 217, 344–354. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Sun, T.; Ao, K.; Peng, Y.; Zhang, Y.; Li, X.; Zhang, Y. Opposite roles of salicylic acid receptors NPR1 and NPR3/NPR4 in transcriptional regulation of plant immunity. Cell 2018, 173, 1454–1467. [Google Scholar] [CrossRef]
- Zhang, Y.; Tessaro, M.J.; Lassner, M.; Li, X. Knockout analysis of Arabidopsis transcription factors TGA2, TGA5, and TGA6 reveals their redundant and essential roles in systemic acquired resistance. Plant Cell 2003, 15, 2647–2653. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; et al. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228–232. [Google Scholar] [CrossRef] [Green Version]
- Petersen, M.; Brodersen, P.; Naested, H.; Andreasson, E.; Lindhart, U.; Johansen, B.; Nielsen, H.B.; Lacy, M.; Austin, M.J.; Parker, J.E.; et al. Arabidopsis MAP kinase 4 negatively regulates systemic acquired resistance. Cell 2000, 103, 1111–1120. [Google Scholar] [CrossRef] [Green Version]
- Brodersen, P.; Petersen, M.; Nielsen, H.B.; Zhu, S.; Newman, M.A.; Shokat, K.M.; Rietz, S.; Parker, J.; Mundy, J. Arabidopsis MAP kinase 4 regulates salicylic acid- and jasmonic acid/ethylene-dependent responses via EDS1 and PAD4. Plant J. 2006, 47, 532–546. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Meng, Y.; Zhang, M.; Tong, X.; Wang, Q.; Sun, Y.; Quan, J.; Govers, F.; Shan, W. Infection of Arabidopsis thaliana by Phytophthora parasitica and identification of variation in host specificity. Mol. Plant Pathol. 2011, 12, 187–201. [Google Scholar] [CrossRef]
- Jin, J.-H.; Zhang, H.-X.; Tan, J.-Y.; Yan, M.-J.; Li, D.-W.; Khan, A.; Gong, Z.-H. A new ethylene-responsive factor CaPTI1 gene of pepper (Capsicum annuum L.) involved in the regulation of defense response to Phytophthora capsici. Front. Plant Sci. 2016, 6, 1217. [Google Scholar] [CrossRef]
- Wang, J.E. Expression Analysis and Functional Identification of CaRGA1 and CaPOD Genes Induced by Phytophthora Capsici in Pepper; Institute of Horticulture, Northwest A&F University: Yangling, China, 2013. [Google Scholar]
- Zhang, Z.; Li, D.-W.; Jin, J.-H.; Yin, Y.-X.; Zhang, H.-X.; Chai, W.-G.; Gong, Z.-H. VIGS approach reveals the modulation of anthocyanin biosynthetic genes by CaMYB in chili pepper leaves. Front. Plant Sci. 2015, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.S.; Gong, Z.; Ji, J.; Yin, Y.; Xiao, H.; Khan, M.A. Construction of the intermediate vector pVBG2307 by incorporating vital elements of expression vectors pBI121 and pBI221. Genet. Mol. Res. 2012, 11, 3091–3104. [Google Scholar] [CrossRef]
- Mou, S.L.; Liu, Z.Q.; Gao, F.; Yang, S.; Su, M.X.; Shen, L.; Wu, Y.; He, S.L. CaHDZ27, a homeodomain-leucine zipper I (HD-Zip I) protein, positively regulates the resistance to Ralstonia solanacearum infection in pepper. Mol. Plant-Microbe Interact. 2017, 30, 960–973. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.L.; Jia, Q.L.; Li, D.W.; Wang, J.E.; Yin, Y.X.; Gong, Z.H. Characteristic of the Pepper CaRGA2 Gene in Defense Responses against Phytophthora capsici Leonian. Int. J. Mol. Sci. 2013, 14, 8985–9004. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.K.; Jeong, M.P.; Young, H.J.; Lee, S.; Chung, E.; Kim, S.Y.; Seung, H.Y.; Choi, D. A plant EPF-type zinc-finger protein, CaPIF1, involved in defence against pathogens. Mol. Plant Pathol. 2005, 6, 269–285. [Google Scholar] [CrossRef]
- Li, S.; Zhao, H.; Li, Y.; Niu, S.; Cai, B. Complete genome sequence of the naphthalene-degrading Pseudomonas putida strain ND6. J. Bacteriol. 2012, 194, 5154–5155. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, D.; Li, Y.; Cai, B. Overexpression, purification and characterization of a new salicylate hydroxylase from naphthalene-degrading Pseudomonas sp. strain ND6. Microbiol. Res. 2005, 160, 307–313. [Google Scholar] [CrossRef]
- Zhang, Y.L. Identification of Physiological Race of Phytophthora Capsici and Analysis of the Efficiency of Chemical Control; Institute of Horticulture, Northwest A&F University: Yangling, China, 2009. [Google Scholar]
- Zhang, H.; Ali, M.; Feng, X.; Jin, J.; Huang, L.; Khan, A.; Lv, J.; Gao, S.; Luo, D.; Gong, Z. A novel transcription factor CaSBP12 gene negatively regulates the defense response against Phytophthora capsici in Pepper (Capsicum annuum L.). Int. J. Mol. Sci. 2018, 20, 48. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.L.; Chen, R.G.; Gong, Z.H.; Yin, Y.X.; Ahmed, S.S.; He, Y.M. Exogenous abscisic acid increases antioxidant enzymes and related gene expression in pepper (Capsicum annuum) leaves subjected to chilling stress. Genet. Mol. Res. 2012, 11, 4063–4080. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Yin, Y.X.; Wang, S.B.; Zhang, H.X.; Xiao, H.J.; Jin, J.H.; Ji, J.J.; Jing, H.; Chen, R.G.; Arisha, M.H.; Gong, Z.H. Cloning and expression analysis of CaPIP1-1 gene in pepper (Capsicum annuum L.). Gene 2015, 563, 87–93. [Google Scholar] [CrossRef]
- Du, Y.; Mpina, M.H.; Birch, P.R.J.; Bouwmeester, K.; Govers, F. Phytophthora infestans RXLR effector AVR1 interacts with exocyst component Sec5 to manipulate plant immunity. Plant Physiol. 2015, 169, 01169. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.H.; Zhang, H.X.; Ali, M.; Gai, W.X.; Cheng, G.X.; Yu, Q.H.; Yang, S.B.; Li, X.X.; Gong, Z.H. A small heat shock protein CaHsp25.9 positively regulates heat, salt, and drought stress tolerance in pepper (Capsicum annuum L.). Plant Physiol. Biochem. 2019, 142, 151–162. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.-X.; Feng, X.-H.; Jin, J.-H.; Khan, A.; Guo, W.-L.; Du, X.-H.; Gong, Z.-H. CaSBP11 Participates in the Defense Response of Pepper to Phytophthora capsici through Regulating the Expression of Defense-Related Genes. Int. J. Mol. Sci. 2020, 21, 9065. https://doi.org/10.3390/ijms21239065
Zhang H-X, Feng X-H, Jin J-H, Khan A, Guo W-L, Du X-H, Gong Z-H. CaSBP11 Participates in the Defense Response of Pepper to Phytophthora capsici through Regulating the Expression of Defense-Related Genes. International Journal of Molecular Sciences. 2020; 21(23):9065. https://doi.org/10.3390/ijms21239065
Chicago/Turabian StyleZhang, Huai-Xia, Xiao-Hui Feng, Jing-Hao Jin, Abid Khan, Wei-Li Guo, Xiao-Hua Du, and Zhen-Hui Gong. 2020. "CaSBP11 Participates in the Defense Response of Pepper to Phytophthora capsici through Regulating the Expression of Defense-Related Genes" International Journal of Molecular Sciences 21, no. 23: 9065. https://doi.org/10.3390/ijms21239065