Sulfur Homeostasis in Plants

State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, The Chinese Academy of Sciences, Beijing 100093, China
*
Author to whom correspondence should be addressed.
†
Current address: College of Horticulture, Northwest A&F University, Yangling 712100, Shaanxi, China.
‡
Current address: Duolun County Grassland Management Station, Inner Mongolia Autonomous Region, Duolun 027300, Inner Mongolia, China.
§
These authors contributed equally to this work.
Int. J. Mol. Sci. 2020, 21(23), 8926; https://doi.org/10.3390/ijms21238926
Submission received: 2 November 2020
/
Revised: 15 November 2020
/
Accepted: 20 November 2020
/
Published: 25 November 2020
(This article belongs to the Special Issue Iron and Sulfur in Plants 2.0)
{kind=link}
{kind=link}
Abstract
:Sulfur (S) is an essential macronutrient for plant growth and development. S is majorly absorbed as sulfate from soil, and is then translocated to plastids in leaves, where it is assimilated into organic products. Cysteine (Cys) is the first organic product generated from S, and it is used as a precursor to synthesize many S-containing metabolites with important biological functions, such as glutathione (GSH) and methionine (Met). The reduction of sulfate takes place in a two-step reaction involving a variety of enzymes. Sulfate transporters (SULTRs) are responsible for the absorption of SO42− from the soil and the transport of SO42− in plants. There are 12–16 members in the S transporter family, which is divided into five categories based on coding sequence homology and biochemical functions. When exposed to S deficiency, plants will alter a series of morphological and physiological processes. Adaptive strategies, including cis-acting elements, transcription factors, non-coding microRNAs, and phytohormones, have evolved in plants to respond to S deficiency. In addition, there is crosstalk between S and other nutrients in plants. In this review, we summarize the recent progress in understanding the mechanisms underlying S homeostasis in plants.
1. Introduction
Sulfur (S) is one of the essential elements for growth and development, and is considered to be the fourth most important nutrient element after nitrogen (N), phosphorus (P), and potassium (K) in plants. S is a constituent of amino acids, chloroplasts, sulfatides, vitamins, coenzymes, and prosthetic groups (iron–S clusters, lipoic acid, thiamine, coenzyme A, etc.) [1,2,3]. Therefore, S plays an important role in photosynthesis, respiration, and the formation of cell membrane structures in plants. Because animals cannot synthesize S-containing amino acids, the circulation of S between plants and the environment is of great significance to the nutrition and health of humans and animals [1,2,4].
In recent years, with the gradual recognition of the importance of S, research on plant S nutrition has also received more and more attention. Indeed, there has been a systematic exploration of the absorption of S and the metabolic processes of S assimilation in plants [2,5,6,7]. Here, the recent progress associated with studies of S homeostasis in plants is summarized.
2. The Physiological Functions of S in Plants
S plays an important role in the process of plant resistance to biotic and abiotic stress [8,9]. There are many S-containing substances related to stress resistance in plants, including glutathione (GSH), S-containing proteins, phytochelatins, and glucosinolates. These S-containing compounds can enhance plant resistance under various stresses. For example, glutathione (GSH) is an important antioxidant substance. The redox system, composed of its reduced state and oxidized state, can eliminate the reactive oxygen species produced by oxidative stress, thereby improving the stress tolerance of plants [2,4,6,10,11]. Glucosinolate is an important secondary metabolite in plants, and its degradation products are important substances in plants for resistance to pests and herbivores [12,13,14].
S is closely related to crop yield and quality [11,15,16]. The content of S-containing amino acids in plants is an important index for evaluating crop quality. S deficiency reduces the proportion of S-containing amino acids in crop grains, whereas S application increases the content of S-containing proteins, thereby increasing the nutritional value of grains [16,17]. Furthermore, S supply levels affect wheat flour extraction rates, gluten quality, and baking quality [18]. S nutrition also affects the yield and quality of forage grass, and the application of S fertilizer can improve these factors, as well as the nutritional status of herbivorous livestock, increasing the yield and quality of wool and milk production and quality in dairy cows [19]. In addition, S nutrition also affects N fixation in legumes. For example, the application of S fertilizers could increase the capacity of N fixation in peas (Pisum sativum L.) and alfalfa (Medicago sativa L.) [20,21].
3. Response of Plants to S Deficiency
In recent years, due to the widespread application of high-purity S-free or low-S fertilizers, the control of industrial S-containing waste gas emissions, and intensive agriculture, S levels in the soil have been reduced, and the problem of soil S deficiency has gradually emerged [22].
Due to the poor mobility of S in plants, the symptoms of S deficiency first appear in the young parts of the plant [18]. Different types of plants have different symptoms of S deficiency, but they are all characterized by the reduced height and chlorosis of leaves (especially new leaves) [18]. The responses of plant root morphology to S-deficient conditions have also been studied [23,24,25]. Under S-deficient conditions, the root development in Arabidopsis was enhanced, leading to more lateral roots and higher root hair density [26]. Dan et al. found that S deficiency had little effect on primary root elongation in Arabidopsis [27]. However, lateral root development in Arabidopsis was significantly inhibited, including a decrease in the number of lateral roots, as well as the density of lateral root primordia and lateral roots. In addition, S application can increase the root length and root surface area of alfalfa grown in soil with low available S [28].
Many studies have been carried out on the response of plants to S nutrition at the physiological level [29,30,31]. Since S is a component of proteins, chloroplasts, and some important enzymes and coenzymes, S deficiency stress decreases S content and S-containing amino acids, leading to reduced metabolic activity in plants [29,30,31]. S deficiency leads to a hindrance of the synthesis of key enzymes in the process of carbon © metabolism, slows the rate of photosynthesis, and results in accumulation of more reactive oxygen species in plants [32].
Plants might reuse S nutrients under S-deficient conditions. Oilseed rape (Brassica napus L.) can transfer S from leaves to the root system during short-term S deficiency, thereby enhancing the recycling of S [33]. In Medicago truncatula plants, the distribution of photosynthetic products can be adjusted to avoid the impact of S deficiency on the next generation [34].
4. S Absorption in Plants
Although plant leaves can absorb gaseous SO2 and H2S, the main source of available S for plants is sulfate (SO42−) in the soil [35]. Plant roots actively absorb SO42− and transport it to the aboveground parts via the xylem. Most SO42− is assimilated into reduced organic S in plastids (mainly in chloroplasts), while the excess SO42− is transported to vacuoles for storage [2,10,36]. Plant sulfate transporters (SULTRs) are responsible for the absorption of SO42− from the soil and the transport of SO42− in plants [37,38].
The plant S transporter family generally includes 12–16 genes, which are divided into five categories according to their coding sequence homology, biochemical properties, and physiological functions [39]. Mutant analysis proved that the two high-affinity S transporters, SULTR1.1 and SULTR1.2, in Arabidopsis are mainly located in the root hairs, root epidermis, and cortex, and they are upregulated by S deficiency. Therefore, they are considered to be mainly responsible for the absorption of S from soil [40]. Another high-affinity S transporter, SULTR1.3 in Arabidopsis, is located in the phloem, and is responsible for the transport of S between source and sink tissues, with its expression in roots and leaves being enhanced under S-deficient conditions [41]. In addition, the high-affinity S transporter HVST1, cloned in barley, has been shown to be expressed in the pericycle and xylem parenchyma cells, and may be involved in S transport in vascular tissues [42].
The second category of S transporters is mainly distributed in the xylem and phloem, and is responsible for the transport of SO42− between tissues. SULTR2.1, encoding a low-affinity S transporter in Arabidopsis, is expressed in the xylem parenchyma and pericycle cells, and is upregulated under S-deficient conditions [43]. SULTR2.1 is mainly responsible for the transport of S from the root system to the shoot. SULTR2.1 may mediate SO42− movement into the xylem parenchyma cells, and then increase the concentration of S in the symplast, thereby facilitating the unloading of S from the xylem [43]. SULTR2.1 has also been shown to be involved in the transfer of S to developing seeds [44]. A low-affinity transporter SULTR2.2 is expressed in the phloem, and is responsible for the transfer of SO42− into the companion cell, which may control the S concentration in the phloem sap [45].
At present, little is known about the function of the third category of S transporter. Arabidopsis SULTR3.5 has been shown to assist SULTR2.1 during the transport of SO42− from roots to the shoots, but it cannot function as an S transporter by itself [46]. Unlike in Arabidopsis, SULTR3.5 can function as an S transporter in yeast [46].
SULTR4.1 and SULTR4.2 in Arabidopsis belong to the fourth category of S transporters. They are located in the vacuole membrane and are responsible for the transfer of SO42− from the vacuole. Under S-deficient conditions, the expression of SULTR4.1 and SULTR4.2 is upregulated, and the SO42− stored in the vacuole is transferred out to meet the plant’s demand for S [47]. In addition, the S transporter SULTR5.2 has been defined as a member of category 5, and is quite different from other members of the S transporter family. Nevertheless, Tomatsu et al. indicated that SULTR5.2 had been identified as a molybdenum transporter [48]. Finally, the important process of SO42− transportation to its assimilation site (i.e., plastids) has not yet been determined [49].
5. S Assimilation and Metabolism in Plants
Sulfate cannot be directly used by plants. It needs to be activated into adenosine 5′-phosphosulfate (APS) and 3′-phosphoadenosine-5′-phosphosulfate (PAPS) [2,50]. ATP Sylase (ATPS) catalyzes SO42− to APS, with APS then phosphorylated to the storage form PAPS by APS kinase (APK). The reduction of sulfate takes place exclusively in the plastid in a two-step reaction. First, adenosine 5′-phosphosulfate reductase (APR) catalyzes the formation of sulfite from APS. Then, the sulfite reductase (SIR) catalyzes sulfite to divalent sulfide (S2−). Under the action of serine acetyltransferases (SAT) and O-acetylserine(thiol)lyase (OASTL), sulfide is integrated into the C scheleton of O-acetylserine (OAS), leading to the formation of the first important S-containing organic compound cysteine (Cys) [10]. More specifically, SAT catalyzes the acetylation reaction of serine and acetyl-CoA to generate OAS, while OASTL catalyzes OAS and S2− to generate Cys. It is generally believed that OASTL is not the rate-limiting step in the synthesis of Cys. As a limiting factor for the generation of OAS, SAT may indirectly affect the synthesis of Cys (Figure 1) [1,2,10].
The synthesis of Cys is a key link in the process of S assimilation and metabolism, and is also the main coordination link between S, C, and N nutrition. Unlike sulfate, which is specifically reduced in plastids, Cys synthesis can be performed in plastids, mitochondria, and the cytoplasmic matrix. Cys is used as a precursor to synthesize many S-containing metabolites with important biological functions in plants [2,10,51,52,53].
5.1. Glutathione
The content of GSH is highest among the sulfhydryl compounds in plants. It participates in the regulation of redox and plays a role in plant resistance to adversity stress [54,55]. It is also the main form of storage and transportation of organic S in plants. Plants use Cys as a synthetic precursor to synthesize GSH through a two-step reaction. First, γ-glutamylcysteine (γ-EC) can be generated from Cys and glutamate under the action of γ-glutamylcysteine synthetase (γ-ECS), which is the rate-limiting step for the synthesis of GSH. Secondly, γ-EC interacts with glycine (or alanine) to produce GSH under the action of glutathione synthetase (GSHS) [56,57]. GSH plays a variety of important roles in maintaining the normal metabolism of plants, including being an important antioxidant, removing active oxygen and heterologous harmful substances, and protecting sulfhydryl groups in proteins and enzyme molecules [2,4,6,10,11]. GSH is also the synthetic precursor of phytochelatins, which play an important role in the process of plant resistance to heavy metal stress [58].
5.2. Methionine
Met is another S-containing amino acid with important biological functions, and it plays a linking role in the process of primary and secondary metabolism in plants [59,60]. Met is synthesized by a multi-step enzymatic reaction, using Cys as the synthetic precursor. Met is synthesized to S-adenosyl methionine (SAM) under the catalysis of S-adenosylmethionine synthase (SAMS), and SAM is the precursor for the synthesis of ethylene, polyamines, vitamins, coenzymes, nicotianamine (NA), and mugineic acid (MA) [6,59]. Among these derivatives, ethylene is an important hormone in plants, and NA and MA play a key role in the absorption and transport of iron and other metal ions in plants [6,59].
5.3. PAPS
5.4. Glucosinolate
Glucosinolates are important, S-containing, secondary metabolites that are mostly found in cruciferous plants [12,13,61]. Their synthesis requires the participation of Cys, PAPS, and sulfotransferase (SOT) [62]. Glucosinolates themselves do not have physiological activity. When plants are damaged, glucosinolates are hydrolyzed by myrosinase in the cells to produce nitriles, thiocyanates, and isothiocyanates with different biological activities [63,64]. These hydrolysates have different physiological functions, including the production of peculiar aroma components in cruciferous vegetables, inhibition of microbial growth, resistance to insects and herbivores, and inactivation of cancer genes [12,13,14].
6. Molecular Regulatory Mechanisms of S Absorption and Assimilation in Plants
Compared with research into other macroelements, research on the regulation mechanisms of S assimilation and absorption in plants is lagging behind, and is still poorly understood [65,66]. In recent years, with the development of genomic, transcriptomic, and metabolomic research, understanding of the physiological and molecular regulatory mechanisms underlying responses to S-deficiency stress in plants has been enhanced [5,6,30,65,67,68,69].
A number of studies have shown that S transporters and the S assimilation enzyme APR play a key role in S nutrition regulation [37,70]. The regulation of S absorption, transport, and assimilation in plants always follows the negative feedback mechanism driven by demand [71]. For example, the expression of genes responsible for S absorption and assimilation is upregulated under S-deficient conditions, and is reduced when S is supplied [72]. In addition, S assimilation products are involved in regulating S absorption and metabolism in plants. Cys and GSH are considered as playing a negative regulation role in S absorption and assimilation, while OAS is believed to play a positive regulation role (Figure 2) [68,73].
6.1. The cis-Acting Element SURE (Sulfur-Responsive Element)
Maruyama-Nakashita et al. first revealed the regulatory mechanism of S absorption and assimilation by plant at the molecular level [74]. By using various inhibitors, the regulation of SULTR1.1 was found to occur at the transcriptional level, and required the participation of protein phosphorylase under S-deficient conditions, which suggests that the promoter of SULTR1.1 may have a cis-acting element in response to S deficiency [74]. Later, a 16-base, cis-acting element was found in the promoter region of the S transporter SULTR1.1 in Arabidopsis. The core region of this element, GAGAC, plays a key role in the response to S deficiency, and was named SURE [75]. Homologous sequences of the core region of SURE have also been found in the promoters of other S-deficiency-inducible genes, including Arabidopsis NIT3 (NITRILASE3) and soybean seed storage protein b-conglycinin, which implies a general regulatory mechanism for S deficiency-induced gene expression [26,76]. However, the promoter region of another high-affinity S transporter SULTR1.2 does not contain this element, indicating that the regulatory pathways of S transporters are diverse [75]. Recent studies have also shown that SULTR1.1 is more specifically regulated by S deficiency, while SULTR1.2 seems to be affected by the plant’s overall metabolic requirements [77]. This result also implies that SURE has specificity and limitations in regulating the response to S deficiency. In addition, the SURE element contains an auxin response factor (ARF) binding sequence (GAGACA), but SURE only plays a role in the response to S deficiency and does not respond to auxin signals [75]. Recently, the sdi-1 gene, which is only induced by S deficiency, has been cloned in wheat. Its promoter region also contains a six-base, S deficiency response, cis-acting element. The position of this element in wheat sdi-1 is the same as that of SURE in SULTR1.1 in Arabidopsis [78].
6.2. SLIM1 Transcription Factor
SLIM1 is the first transcription factor known to be involved in the regulation of S absorption, and is responsible for the up-regulation of S transporters under S-deficient conditions [79,80]. These authors isolated the Arabidopsis mutant slim1 (sulfur limitation 1) by screening mutants with a green fluorescent reporter gene. The SLIM1 gene was identified as a member of the transcription factor EIL (ETHYLENE-INSENSITIVE3-LIKE) family, with a specific role in the regulation of S nutrition. SULTR1.2 in the slim1 mutant cannot be upregulated by S deficiency, indicating that SLIM1 is involved in the regulation of S transporters under S-deficient conditions. Transcriptome analysis of slim1 mutants has shown that SLIM1 regulates the S transporters SULTR1.2, SULTR1.1, SULTR3.4, and SULTR4.2, as well as a myrosinase gene. However, SLIM1 does not affect the expression of the key enzyme APR, which is induced by S deficiency [79]. In addition, the regulation of S transporters by SLIM1 does not require the participation of SURE elements. Therefore, the interaction between transcription factors and cis-acting elements in regulating the S deficiency response needs further study [79]. In addition, SLIM1 is located in the vascular tissue, but the nutrient absorption site is on the surface of the root system. Therefore, the regulation of S absorption by SLIM1 may involve long-distance signal transduction between tissues.
6.3. miR395
miR395 is involved in the regulation of S deficiency signals in plants, and its target genes include the low-affinity S transporter SULTR2.1 and the three homologous genes APS1, APS3, and APS4 encoding the APS enzyme [2,81]. S deficiency induces the up-regulation of miR395, and this process depends on SLIM1 [2,81]. Strangely, S deficiency enhances the expression of miR395 and its target gene SULTR2.1 at the same time, but tissue localization studies have shown that the expression sites of the two genes are different. miR395 is located in the phloem companion cell, and SULTR2.1 is located in the xylem parenchyma. It seems that SULTR2.1 only transports S in the xylem. Similarly, due to constraints of spatial location, the expression of SULTR2.1 may not be fully regulated by miR395 [2,81]. In addition, miR395 is also involved in regulating the accumulation of Sin leaves and transporting it from old leaves to new leaves [81,82].
7. The Role of Hormones in the Regulation of S Nutrition in Plants
Phytohormones are involved in the regulation of S absorption, transport, and assimilation metabolism in plants.
7.1. Cytokinins
Cytokinins (CTKs) inhibit the expression of S transporters SULTR1.1 and SULTR1.2, thereby negatively regulating S uptake in Arabidopsis. This process is dependent on cytokinin receptors CRE1/WOL/AHK4 [83]. However, CTKs do not affect the induction of S absorption under S-deficient conditions, indicating that the negative regulation of cytokinins and the signal pathway of S absorption under S-deficient conditions are two independent pathways [83]. In addition, studies have reported that exogenous treatment with cytokinin induces the expression of the S transporter SULTR2.2 and the key S assimilation enzyme APR [84]. These results suggest that the role of cytokinins in regulating the plant’s response to S deficiency is not very clearly understood.
7.2. Auxin
Some progress has been made on the role of auxin in the regulation of root morphology under S-deficient conditions [26,85]. As mentioned earlier, glucosinolates are decomposed into SO42− and indole-3-acetonitrile (IAN) by myrosinase. IAN can be converted into indole acetic acid (IAA) under the action of nitrilase (NIT). There are four nitrilase genes in Arabidopsis, three of which (NIT1, NIT2, and NIT3) can convert IAN to IAA [86]. S deficiency enhances the expression of the nitrilase-encoding gene NIT3 in Arabidopsis, and then promotes the conversion of glucosinolates to IAN. Therefore, it has been speculated that the synthesis of auxin is enhanced to promote root development in Arabidopsis under S-deficient conditions [26]. However, there is no difference in auxin content in plants growing under S-deficient and S-sufficient conditions [26]. In addition, transcriptome studies have shown that S deficiency can induce IAA synthesis-related genes, which may increase the synthesis of auxin, thereby enhancing Arabidopsis root development under S-deficient conditions [85]. By using the DR5::GUS reporter strain, the inhibition of lateral root development in Arabidopsis by S deficiency was found to be due to reduced auxin synthesis or auxin sensitivity, which indicates that auxin negatively regulates the response of Arabidopsis to S deficiency [27]. Therefore, auxin may regulate root morphology under S-deficient conditions through both positive and negative feedback pathways [87,88]. First, S deficiency upregulates the NIT3 and myrosinase-related genes, thereby increasing the synthesis of auxin to enhance root growth [26,85]. On the other hand, the increased auxin contents change cell calcium ion concentrations and then upregulate the expression level of calmodulin. Calmodulin acts on the auxin-related transcription factor IAA28, which may inhibit the expression of auxin-induced genes [89]. In addition to the positive regulation pathway, in which S deficiency enhances root growth by increasing auxin synthesis, there is an IAA28-mediated negative feedback regulation pathway that prevents the enhancement of plant root development under S-deficient conditions, and in turn initiates a negative feedback regulation mechanism for restricting root growth [87,88]. Although this evidence indicates that auxin is involved in regulating the response of root morphology to S deficiency, its mechanism remains to be clarified.
Furthermore, auxin has no effect on the upregulation of high-affinity plant S transporter expression under S-deficient conditions [74]. SURE, the key regulatory element of the S deficiency response, contains a binding sequence (GAGACA) for the auxin response factor. However, there is no evidence to indicate that SURE is related to auxin signaling [75]. Recent studies have shown that the overexpression of auxin-related genes in response to S deficiency changes many metabolic processes in plants without affecting S metabolism [90].
7.3. Other Hormones
The ethylene signal pathway may be related to the response of tobacco LSU (response to low sulfur) family of proteins under S-deficient conditions [91,92]. Transcriptome analysis has shown that jasmonic acid (JA) is related to the expression of genes involved in S metabolism, and JA synthesis-related genes are induced by S deficiency [93,94]. These results imply that JA plays a positive role in the regulation of S metabolism. Recently, gibberellin has been implicated in the regulation of APR transcription and enzyme activity in Arabidopsis. JA, salicylic acid, ethylene, and CKs only regulate APR enzyme activity, whereas ABA (abscisic acid) does not affect the upregulation of APR under salt stress [95].
8. Systemic Signals Involved in Regulation of Plant S Nutrition
Systemic signals (i.e., long-distance signals) are messengers that connect plant roots and shoots. They can regulate the overall relationship between nutrient requirements in plants and nutrient supply in soil. Several S-deficiency-responsive genes (such as APR3) have been shown to be regulated by systemic signals, and are believed to be related to the decrease in OAS content in Arabidopsis [96]. However, systemic signals that are involved in regulation of plant S nutrition are relatively elusive [97]. In the process of regulating the expression of S transporters and S absorption activity, no systemic signals from the shoots to the roots were found in rape plants under S-deficient conditions [98]. In Arabidopsis, SULTR1.1, and SULTR1.2 are only regulated by local S deficiency, but not affected by systemic signals. In addition, the expression of SULTR1.1 and SULTR1.2 does not depend on OAS or GSH [77]. Overall, there are conflicting conclusions about whether plants have a systemic response to S-deficiency stress. Specific systemic signal molecules have not been determined. Further study is needed to elucidate the regulation mechanism of systemic signals under S-deficient conditions.
9. Crosstalk between S and Other Nutrition in Plants
There is a tendency for the balance of nutrient metabolism to be maintained in the plant body, so the interaction between nutrient elements will affect the plant response to the stress of mineral nutrient deficiency [99]. However, there are relatively few reports on the interaction between S and other nutrients [8]. Therefore, more attention needs to be paid to the mechanism of crosstalk between S and other forms of nutrition in plants [100].
9.1. S and N
There is an interaction between the regulation of S and N nutrition metabolism in plants [101]. N deficiency inhibits the response of SULTR1.1 and SULTR1.2 to S deficiency in Arabidopsis [74]. N deficiency downregulates expression of the key enzyme APR during S assimilation, and the Cys precursor OAS may be a node that connects N and S metabolism [102,103]. Indeed, N supply can induce the upregulation of genes responsible for S absorption and assimilation, thereby increasing the plant’s S absorption efficiency [104].
A lack of S nutrition affects the utilization of N in plants. S deficiency inhibits the uptake of nitrate and reduces the activity of nitrate reductase, leading to the accumulation of nitrate and a reduction in N utilization in plants [105]. S-deficiency stress leads to a decrease in the content of S and S-containing amino acids in plants, which may be the cause of hindered protein synthesis and the accumulation of non-protein forms of N [106]. However, in a study of rapeseed plants under short-term S deficiency, the total N content and N absorption were not affected [33].
9.2. S and C
The absorption and metabolism of S are also affected by C metabolism in plants [101]. Sugar can upregulate the expression of the S transporters SULTR1.1 and SULTR1.2, and increase the activity and transcription level of APR [107,108]. Under CO2-free conditions, the absorption and transport of S are inhibited, while the activity and transcription level of APR are decreased in plants [107].
9.3. S and P
There is a correlation between S and P nutrition metabolism in plants [109]. Sulfolipids are quickly synthesized to replace phospholipids under P-deficient conditions, and phospholipids can also replace sulfolipids under S-deficient conditions [110]. S and P transporters have similar topological structures and molecular regulation mechanisms [111]. For example, the absorption and transport of S and P in plants are both negatively regulated by cytokinins [83,112]. miR395, which is involved in the regulation of the S deficiency response [81], is also upregulated under P-deficient conditions [113]. Transcriptome analysis has shown that P deficiency upregulates the expression of S transporters in Arabidopsis [114]. PHR1 (Phosphate Response 1), which is a key transcription factor in the P-deficiency signaling pathway, may play an important role in the P and S nutrient interaction in Arabidopsis [115]. SULTR1.3, which is responsible for the transport of S in shoots, is upregulated by P deficiency, and its promoter region contains a PHR1 binding sequence [115]. By using phr mutants, the regulation of SULTR1.3 by P deficiency was proven to depend on PHR1 [115]. PHR1 also inhibits the expression of other S transporters (SULTR2.1 and SULTR3.4) [115]. Furthermore, the PHR1 homologous gene PSR1 in Chlamydomonas reinhardtii also negatively regulates S transporters [116]. Nevertheless, P deficiency has not shown any effect on the expression of SULTR1.1 and SULTR1.2 in Arabidopsis [74].
Overall, the regulation of a plant’s response to S deficiency is interrelated with the metabolism of N, C, and P; however, the specific regulation mechanisms are still not very well known.
10. Conclusions
S nutrition plays an important role in the growth and development of plants. It is closely related to the response of plants to biotic and abiotic stresses, as well as the yield and quality of crops. Soil S deficiency has gradually become one of the main factors limiting plant growth and crop yields. However, compared with studies on other macronutrient elements, there is not enough research on plant S nutrition, and a mechanistic understanding of the absorption, metabolism, and regulation of S nutrition remains elusive. Therefore, more effort is needed to understand the plant’s response to soil S deficiency stress and its regulatory mechanisms, and this will form the basis for improving S nutrient utilization efficiency in crops.
Author Contributions
Conceptualization, A.Y. and Y.G.; writing, Q.L., Y.G., and A.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China (2016YFC0500706) and the Natural Science Basic Research Plan in Shaanxi Province of China (Program No. 2020JQ-250).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nakai, Y.; Maruyama-Nakashita, A. Biosynthesis of Sulfur-Containing Small Biomolecules in Plants. Int. J. Mol. Sci. 2020, 21, 3470. [Google Scholar]
- Maruyama-Nakashita, A. Metabolic changes sustain the plant life in low-sulfur environments. Curr. Opin. Plant Biol. 2017, 39, 144–151. [Google Scholar] [PubMed]
- Yoshimoto, N.; Saito, K. S-Alk(en)ylcysteine sulfoxides in the genus Allium: Proposed biosynthesis, chemical conversion, and bioactivities. J. Exp. Bot. 2019, 70, 4123–4137. [Google Scholar] [PubMed]
- Kopriva, S.; Malagoli, M.; Takahashi, H. Sulfur nutrition: Impacts on plant development, metabolism, and stress responses. J. Exp. Bot. 2019, 70, 4069–4073. [Google Scholar]
- Watanabe, M.; Hoefgen, R. Sulphur systems biology-making sense of omics data. J. Exp. Bot. 2019, 70, 4155–4170. [Google Scholar]
- Aarabi, F.; Naake, T.; Fernie, A.R.; Hoefgen, R. Coordinating Sulfur Pools under Sulfate Deprivation. Trends Plant Sci. 2020, 25, 1227–1239. [Google Scholar]
- Bouranis, D.L.; Malagoli, M.; Avice, J.C.; Bloem, E. Advances in Plant Sulfur Research. Plants 2020, 9, 256. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Mahmud, J.A.; Nahar, K.; Mohsin, S.M.; Parvin, K.; Fujita, M. Interaction of sulfur with phytohormones and signaling molecules in conferring abiotic stress tolerance to plants. Plant Signal. Behav. 2018, 13, e1477905. [Google Scholar]
- Capaldi, F.R.; Gratão, P.L.; Reis, A.R.; Lima, L.W.; Azevedo, R.A. Sulfur Metabolism and Stress Defense Responses in Plants. Trop. Plant Biol. 2015, 8, 60–73. [Google Scholar]
- Chan, K.X.; Phua, S.Y.; Van Breusegem, F. Secondary sulfur metabolism in cellular signalling and oxidative stress responses. J. Exp. Bot. 2019, 70, 4237–4250. [Google Scholar]
- Henriet, C.; Aime, D.; Terezol, M.; Kilandamoko, A.; Rossin, N.; Combes-Soia, L.; Labas, V.; Serre, R.-F.; Prudent, M.; Kreplak, J.; et al. Water stress combined with sulfur deficiency in pea affects yield components but mitigates the effect of deficiency on seed globulin composition. J. Exp. Bot. 2019, 70, 4287–4303. [Google Scholar] [PubMed]
- Aarabi, F.; Kusajima, M.; Tohge, T.; Konishi, T.; Gigolashvili, T.; Takamune, M.; Sasazaki, Y.; Watanabe, M.; Nakashita, H.; Fernie, A.R.; et al. Sulfur deficiency-induced repressor proteins optimize glucosinolate biosynthesis in plants. Sci. Adv. 2016, 2, e1601087. [Google Scholar] [PubMed] [Green Version]
- Zhang, L.; Kawaguchi, R.; Morikawa-Ichinose, T.; Allahham, A.; Kim, S.-J.; Maruyama-Nakashita, A. Sulfur Deficiency-Induced Glucosinolate Catabolism Attributed to Two β-Glucosidases, BGLU28 and BGLU30, is Required for Plant Growth Maintenance under Sulfur Deficiency. Plant Cell Physiol. 2020, 61, 803–813. [Google Scholar]
- Traka, M.H. Chapter Nine—Health Benefits of Glucosinolates. In Advances in Botanical Research; Kopriva, S., Ed.; Academic Press: Cambridge, MA, USA, 2016; Volume 80, pp. 247–279. [Google Scholar]
- Fuentes-Lara, L.O.; Medrano-Macías, J.; Pérez-Labrada, F.; Rivas-Martínez, E.N.; García-Enciso, E.L.; González-Morales, S.; Juárez-Maldonado, A.; Rincón-Sánchez, F.; Benavides-Mendoza, A. From Elemental Sulfur to Hydrogen Sulfide in Agricultural Soils and Plants. Molecules 2019, 24, 2282. [Google Scholar]
- Yu, Z.; Juhasz, A.; Islam, S.; Diepeveen, D.; Zhang, J.; Wang, P.; Ma, W. Impact of mid-season sulphur deficiency on wheat nitrogen metabolism and biosynthesis of grain protein. Sci. Rep. 2018, 8, 2499. [Google Scholar] [PubMed] [Green Version]
- Blake-Kalff, M.M.A.; Hawkesford, M.J.; Zhao, F.J.; McGrath, S.P. Diagnosing sulfur deficiency in field-grown oilseed rape (Brassica napus L.) and wheat (Triticum aestivum L.). Plant Soil. 2000, 225, 95–107. [Google Scholar]
- Scherer, H.W. Sulphur in crop production—Invited paper. Eur. J. Agron. 2001, 14, 81–111. [Google Scholar]
- Wang, S.P.; Wang, Y.F.; Cui, X.Y.; Chen, Z.Z.; Chi, H.K.; Schnug, E.; Haneklaus, S.; Fleckenstein, J. Influence of sulphur fertilizer on sulphur cycling and its implications on sulphur fertilizer requirement of grazed pasture in warm seasonal rangeland of Nei Monggol steppe of China. Acta Bot. Sin. 2002, 44, 204–211. [Google Scholar]
- Scherer, H.W.; Pacyna, S.; Spoth, K.R.; Schulz, M. Low levels of ferredoxin, ATP and leghemoglobin contribute to limited N2 fixation of peas (Pisum sativum L.) and alfalfa (Medicago sativa L.) under S deficiency conditions. Biol. Fert. Soils 2008, 44, 909–916. [Google Scholar]
- Schneider, S.; Schintlmeister, A.; Becana, M.; Wagner, M.; Woebken, D.; Wienkoop, S. Sulfate is transported at significant rates through the symbiosome membrane and is crucial for nitrogenase biosynthesis. Plant Cell Environ. 2019, 42, 1180–1189. [Google Scholar]
- Gao, Y.; Li, X.; Tian, Q.-Y.; Wang, B.-L.; Zhang, W.-H. Sulfur deficiency had different effects on Medicago truncatula ecotypes A17 and R108 in terms of growth, root morphology and nutrient contents. J. Plant Nutr. 2016, 39, 301–314. [Google Scholar]
- Kimura, Y.; Ushiwatari, T.; Suyama, A.; Tominaga-Wada, R.; Wada, T.; Maruyama-Nakashita, A. Contribution of Root Hair Development to Sulfate Uptake in Arabidopsis. Plants 2019, 8, 106. [Google Scholar]
- Joshi, N.C.; Meyer, A.J.; Bangash, S.A.K.; Zheng, Z.-L.; Leustek, T. Arabidopsis γ-glutamylcyclotransferase affects glutathione content and root system architecture during sulfur starvation. New Phytol. 2019, 221, 1387–1397. [Google Scholar] [PubMed] [Green Version]
- Dong, W.; Wang, Y.; Takahashi, H. CLE-CLAVATA1 Signaling Pathway Modulates Lateral Root Development under Sulfur Deficiency. Plants 2019, 8, 103. [Google Scholar]
- Kutz, A.; Muller, A.; Hennig, P.; Kaiser, W.M.; Piotrowski, M.; Weiler, E.W. A role for nitrilase 3 in the regulation of root morphology in sulphur-starving Arabidopsis thaliana. Plant J. 2002, 30, 95–106. [Google Scholar]
- Dan, H.; Yang, G.; Zheng, Z.-L. A negative regulatory role for auxin in sulphate deficiency response in Arabidopsis thaliana. Plant Mol. Biol. 2007, 63, 221–235. [Google Scholar]
- Wang, Y.F.; Wang, S.P.; Cui, X.Y.; Chen, Z.Z.; Schnug, E.; Haneklau, S. Effects of sulphur supply on the morphology of shoots and roots of alfalfa (Medicago sativa L.). Grass Forage Sci. 2003, 58, 160–167. [Google Scholar]
- Forieri, I.; Sticht, C.; Reichelt, M.; Grerz, N.; Hawkesford, M.J.; Malagoli, M.; Wirtz, M.; Hell, R. System analysis of metabolism and the transcriptome in Arabidopsis thaliana roots reveals differential co-regulation upon iron, sulfur and potassium deficiency. Plant Cell Environ. 2017, 40, 95–107. [Google Scholar]
- Bielecka, M.; Watanabe, M.; Morcuende, R.; Scheible, W.-R.; Hawkesford, M.J.; Hesse, H.; Hoefgen, R. Transcriptome and metabolome analysis of plant sulfate starvation and resupply provides novel information on transcriptional regulation of metabolism associated with sulfur, nitrogen and phosphorus nutritional responses in Arabidopsis. Front. Plant Sci. 2015, 5, 805. [Google Scholar]
- Sung, J.; Baek, S.; Kim, J.; Kim, Y.; Lee, Y.; Lee, S.; Lee, D.; Jung, H.-I. Responses of Primary Metabolites and Glucosinolates in Sulfur Deficient-Cabbage (Brassica rapa L. ssp. Pekinensis). J. Plant Biochem. Physiol. 2018, 6, 223. [Google Scholar]
- Lunde, C.; Zygadlo, A.; Simonsen, H.T.; Nielsen, P.L.; Blennow, A.; Haldrup, A. Sulfur starvation in rice: The effect on photosynthesis, carbohydrate metabolism, and oxidative stress protective pathways. Physiol. Plant. 2008, 134, 508–521. [Google Scholar] [PubMed]
- Abdallah, M.; Dubousset, L.; Meuriot, F.; Etienne, P.; Avice, J.C.; Ourry, A. Effect of mineral sulphur availability on nitrogen and sulphur uptake and remobilization during the vegetative growth of Brassica napus L. J. Exp. Bot. 2010, 61, 2635–2646. [Google Scholar] [PubMed]
- Zuber, H.; Poignavent, G.; Le Signor, C.; Aime, D.; Vieren, E.; Tadla, C.; Lugan, R.; Belghazi, M.; Labas, V.; Santoni, A.-L.; et al. Legume adaptation to sulfur deficiency revealed by comparing nutrient allocation and seed traits in Medicago truncatula. Plant J. 2013, 76, 982–996. [Google Scholar] [PubMed]
- Telman, W.; Dietz, K.-J. Thiol redox-regulation for efficient adjustment of sulfur metabolism in acclimation to abiotic stress. J. Exp. Bot. 2019, 70, 4223–4236. [Google Scholar]
- Huang, Q.; Wang, M.; Xia, Z. The SULTR gene family in maize (Zea mays L.): Gene cloning and expression analyses under sulfate starvation and abiotic stress. J. Plant Physiol. 2018, 220, 24–33. [Google Scholar]
- Takahashi, H. Sulfate transport systems in plants: Functional diversity and molecular mechanisms underlying regulatory coordination. J. Exp. Bot. 2019, 70, 4075–4087. [Google Scholar]
- Vatansever, R.; Koc, I.; Ozyigit, I.I.; Sen, U.; Uras, M.E.; Anjum, N.A.; Pereira, E.; Filiz, E. Genome-wide identification and expression analysis of sulfate transporter (SULTR) genes in potato (Solanum tuberosum L.). Planta 2016, 244, 1167–1183. [Google Scholar]
- Takahashi, H.; Kopriva, S.; Giordano, M.; Saito, K.; Hell, R. Sulfur Assimilation in Photosynthetic Organisms: Molecular Functions and Regulations of Transporters and Assimilatory Enzymes. Annu. Rev. Plant Biol. 2011, 62, 157–184. [Google Scholar]
- Yoshimoto, N.; Takahashi, H.; Smith, F.W.; Yamaya, T.; Saito, K. Two distinct high-affinity sulfate transporters with different inducibilities mediate uptake of sulfate in Arabidopsis roots. Plant J. 2002, 29, 465–473. [Google Scholar]
- Yoshimoto, N.; Inoue, E.; Saito, K.; Yamaya, T.; Takahashi, H. Phloem-localizing sulfate transporter, Sultr1;3, mediates re-distribution of sulfur from source to sink organs in Arabidopsis. Plant Physiol. 2003, 131, 1511–1517. [Google Scholar]
- Rae, A.L.; Smith, F.W. Localisation of expression of a high-affinity sulfate transporter in barley roots. Planta 2002, 215, 565–568. [Google Scholar] [PubMed]
- Maruyama-Nakashita, A.; Watanabe-Takahashi, A.; Inoue, E.; Yamaya, T.; Saito, K.; Takahashi, H. Sulfur-Responsive Elements in the 3′-Nontranscribed Intergenic Region Are Essential for the Induction of SULFATE TRANSPORTER 2;1 Gene Expression in Arabidopsis Roots under Sulfur Deficiency. Plant Cell 2015, 27, 1279–1296. [Google Scholar]
- Awazuhara, M.; Fujiwara, T.; Hayashi, H.; Watanabe-Takahashi, A.; Takahashi, H.; Saito, K. The function of SULTR2;1 sulfate transporter during seed development in Arabidopsis thaliana. Physiol. Plant. 2005, 125, 95–105. [Google Scholar]
- Kirschner, S.; Woodfield, H.; Prusko, K.; Koczor, M.; Gowik, U.; Hibberd, J.M.; Westhoff, P. Expression of SULTR2;2, encoding a low-affinity sulphur transporter, in the Arabidopsis bundle sheath and vein cells is mediated by a positive regulator. J. Exp. Bot. 2018, 69, 4897–4906. [Google Scholar]
- Kataoka, T.; Hayashi, N.; Yamaya, T.; Takahashi, H. Root-to-shoot transport of sulfate in Arabidopsis. Evidence for the role of SULTR3;5 as a component of low-affinity sulfate transport system in the root vasculature. Plant Physiol. 2004, 136, 4198–4204. [Google Scholar]
- Kataoka, T.; Watanabe-Takahashi, A.; Hayashi, N.; Ohnishi, M.; Mimura, T.; Buchner, P.; Hawkesford, M.J.; Yamaya, T.; Takahashi, H. Vacuolar sulfate transporters are essential determinants controlling internal distribution of sulfate in Arabidopsis. Plant Cell 2004, 16, 2693–2704. [Google Scholar]
- Tomatsu, H.; Takano, J.; Takahashi, H.; Watanabe-Takahashi, A.; Shibagaki, N.; Fujiwara, T. An Arabidopsis thaliana high-affinity molybdate transporter required for efficient uptake of molybdate from soil. Proc. Natl. Acad. Sci. USA 2007, 104, 18807–18812. [Google Scholar]
- Chen, Z.; Zhao, P.-X.; Miao, Z.-Q.; Qi, G.-F.; Wang, Z.; Yuan, Y.; Ahmad, N.; Cao, M.-J.; Hell, R.; Wirtz, M.; et al. SULTR3s Function in Chloroplast Sulfate Uptake and Affect ABA Biosynthesis and the Stress Response. Plant Physiol. 2019, 180, 593–604. [Google Scholar]
- Kaufmann, C.; Sauter, M. Sulfated plant peptide hormones. J. Exp. Bot. 2019, 70, 4267–4277. [Google Scholar]
- Joshi, J.; Saboori-Robat, E.; Solouki, M.; Mohsenpour, M.; Marsolais, F. Distribution and possible biosynthetic pathway of non-protein sulfur amino acids in legumes. J. Exp. Bot. 2019, 70, 4115–4121. [Google Scholar]
- Nakabayashi, R.; Saito, K. Ultrahigh resolution metabolomics for S-containing metabolites. Curr. Opin. Biotechnol. 2017, 43, 8–16. [Google Scholar]
- Günal, S.; Hardman, R.; Kopriva, S.; Mueller, J.W. Sulfation pathways from red to green. J. Biol. Chem. 2019, 294, 12293–12312. [Google Scholar]
- Cheng, M.C.; Ko, K.; Chang, W.L.; Kuo, W.C.; Chen, G.H.; Lin, T.P. Increased glutathione contributes to stress tolerance and global translational changes in Arabidopsis. Plant J. 2015, 83, 926–939. [Google Scholar]
- Speiser, A.; Silbermann, M.; Dong, Y.H.; Haberland, S.; Uslu, V.V.; Wang, S.S.; Bangash, S.A.K.; Reichelt, M.; Meyer, A.J.; Wirtz, M.; et al. Sulfur Partitioning between Glutathione and Protein Synthesis Determines Plant Growth. Plant Physiol. 2018, 177, 927–937. [Google Scholar]
- Hendrix, S.; Jozefczak, M.; Wójcik, M.; Deckers, J.; Vangronsveld, J.; Cuypers, A. Glutathione: A key player in metal chelation, nutrient homeostasis, cell cycle regulation and the DNA damage response in cadmium-exposed Arabidopsis thaliana. Plant Physiol. Biochem. 2020, 154, 498–507. [Google Scholar]
- Jez, J.M. Structural biology of plant sulfur metabolism: From sulfate to glutathione. J. Exp. Bot. 2019, 70, 4089–4103. [Google Scholar]
- Maruyama-Nakashita, A.; Ohkama-Ohtsu, N. Sulfur Assimilation and Glutathione Metabolism in Plants. In Glutathione in Plant Growth, Development, and Stress Tolerance; Hossain, M.A., Mostofa, M.G., Diaz-Vivancos, P., Burritt, D.J., Fujita, M., Tran, L.-S.P., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 287–308. [Google Scholar]
- Wawrzynska, A.; Moniuszko, G.; Sirko, A. Links Between Ethylene and Sulfur Nutrition-A Regulatory Interplay or Just Metabolite Association? Front. Plant Sci. 2015, 6, 1053. [Google Scholar]
- Amir, R.; Cohen, H.; Hacham, Y. Revisiting the attempts to fortify methionine content in plant seeds. J. Exp. Bot. 2019, 70, 4105–4114. [Google Scholar]
- Fu, L.; Wang, M.; Han, B.; Tan, D.; Sun, X.; Zhang, J. Arabidopsis Myrosinase Genes AtTGG4 and AtTGG5 Are Root-Tip Specific and Contribute to Auxin Biosynthesis and Root-Growth Regulation. Int. J. Mol. Sci. 2016, 17, 892. [Google Scholar]
- Kopriva, S.; Gigolashvili, T. Glucosinolate Synthesis in the Context of Plant Metabolism. In Advances in Botanical Research; Kopriva, S., Ed.; Academic Press: Cambridge, MA, USA, 2016; Volume 80, pp. 99–124. [Google Scholar]
- Sugiyama, R.; Hirai, M.Y. Atypical Myrosinase as a Mediator of Glucosinolate Functions in Plants. Front. Plant Sci. 2019, 10, 1008. [Google Scholar]
- Jeschke, V.; Weber, K.; Moore, S.S.; Burow, M. Coordination of Glucosinolate Biosynthesis and Turnover Under Different Nutrient Conditions. Front. Plant Sci. 2019, 10, 1560. [Google Scholar] [PubMed] [Green Version]
- Kopriva, S. Plant sulfur nutrition: From Sachs to Big Data. Plant Signal. Behav. 2015, 10, e1055436. [Google Scholar] [PubMed] [Green Version]
- Huang, X.-Y.; Chao, D.-Y.; Koprivova, A.; Danku, J.; Wirtz, M.; Mueller, S.; Sandoval, F.J.; Bauwe, H.; Roje, S.; Dilkes, B.; et al. Nuclear Localised MORE SULPHUR ACCUMULATION1 Epigenetically Regulates Sulphur Homeostasis in Arabidopsis thaliana. PLoS Genet. 2016, 12, e1006298. [Google Scholar]
- Canales, J.; Uribe, F.; Henríquez-Valencia, C.; Lovazzano, C.; Medina, J.; Vidal, E.A. Transcriptomic analysis at organ and time scale reveals gene regulatory networks controlling the sulfate starvation response of Solanum lycopersicum. BMC Plant Biol. 2020, 20, 385. [Google Scholar]
- Huang, X.Y.; Li, M.; Luo, R.; Zhao, F.J.; Salt, D.E. Epigenetic regulation of sulfur homeostasis in plants. J. Exp. Bot. 2019, 70, 4171–4182. [Google Scholar]
- Bonnot, T.; Martre, P.; Hatte, V.; Dardevet, M.; Leroy, P.; Bénard, C.; Falagán, N.; Martin-Magniette, M.-L.; Deborde, C.; Moing, A.; et al. Omics Data Reveal Putative Regulators of Einkorn Grain Protein Composition under Sulfur Deficiency. Plant Physiol. 2020, 183, 501–516. [Google Scholar]
- Scheerer, U.; Haensch, R.; Mendel, R.R.; Kopriva, S.; Rennenberg, H.; Herschbach, C. Sulphur flux through the sulphate assimilation pathway is differently controlled by adenosine 5′-phosphosulphate reductase under stress and in transgenic poplar plants overexpressing gamma-ECS, SO, or APR. J. Exp. Bot. 2010, 61, 609–622. [Google Scholar]
- Kopriva, S. Regulation of sulfate assimilation in Arabidopsis and beyond. Ann. Bot. 2006, 97, 479–495. [Google Scholar]
- Maruyama-Nakashita, A. Transcriptional regulation of genes involved in sulfur assimilation in plants: Understanding from the analysis of high-affinity sulfate transporters. Plant Biotechnol. 2008, 25, 323–328. [Google Scholar]
- Hubberten, H.-M.; Klie, S.; Caldana, C.; Degenkolbe, T.; Willmitzer, L.; Hoefgen, R. Additional role of O-acetylserine as a sulfur status-independent regulator during plant growth. Plant J. 2012, 70, 666–677. [Google Scholar]
- Maruyama-Nakashita, A.; Nakamura, Y.; Watanabe-Takahashi, A.; Yamaya, T.; Takahashi, H. Induction of SULTR1;1 sulfate transporter in Arabidopsis roots involves protein phosphorylation/dephosphorylation circuit for transcriptional regulation. Plant Cell Physiol. 2004, 45, 340–345. [Google Scholar] [PubMed] [Green Version]
- Maruyama-Nakashita, A.; Nakamura, Y.; Watanabe-Takahashi, A.; Inoue, E.; Yamaya, T.; Takahashi, H. Identification of a novel cis-acting element conferring sulfur deficiency response in Arabidopsis roots. Plant J. 2005, 42, 305–314. [Google Scholar] [PubMed]
- Awazuhara, M.; Kim, H.; Goto, D.B.; Matsui, A.; Hayashi, H.; Chino, M.; Kim, S.G.; Naito, S.; Fujiwara, T. A 235-bp region from a nutritionally regulated soybean seed-specific gene promoter can confer its sulfur and nitrogen response to a constitutive promoter in aerial tissues of Arabidopsis thaliana. Plant Sci. 2002, 163, 75–82. [Google Scholar]
- Rouached, H.; Wirtz, M.; Alary, R.; Hell, R.; Arpat, A.B.; Davidian, J.C.; Fourcroy, P.; Berthomieu, P. Differential regulation of the expression of two high-affinity sulfate transporters, SULTR1.1 and SULTR1.2, in Arabidopsis. Plant Physiol. 2008, 147, 897–911. [Google Scholar]
- Howarth, J.R.; Parmar, S.; Barraclough, P.B.; Hawkesford, M.J. A sulphur deficiency-induced gene, sdi1, involved in the utilization of stored sulphate pools under sulphur-limiting conditions has potential as a diagnostic indicator of sulphur nutritional status. Plant Biotechnol. J. 2009, 7, 200–209. [Google Scholar]
- Maruyama-Nakashita, A.; Nakamura, Y.; Tohge, T.; Saito, K.; Takahashi, H. Arabidopsis SLIM1 Is a Central Transcriptional Regulator of Plant Sulfur Response and Metabolism. Plant Cell 2006, 18, 3235–3251. [Google Scholar]
- Wawrzyńska, A.; Sirko, A. EIN3 interferes with the sulfur deficiency signaling in Arabidopsis thaliana through direct interaction with the SLIM1 transcription factor. Plant Sci. 2016, 253, 50–57. [Google Scholar]
- Kawashima, C.G.; Matthewman, C.A.; Huang, S.; Lee, B.-R.; Yoshimoto, N.; Koprivova, A.; Rubio-Somoza, I.; Todesco, M.; Rathjen, T.; Saito, K.; et al. Interplay of SLIM1 and miR395 in the regulation of sulfate assimilation in Arabidopsis. Plant J. 2011, 66, 863–876. [Google Scholar]
- Liang, G.; Yang, F.; Yu, D. MicroRNA395 mediates regulation of sulfate accumulation and allocation in Arabidopsis thaliana. Plant J. 2010, 62, 1046–1057. [Google Scholar]
- Werner, T.; Nehnevajova, E.; Koellmer, I.; Novak, O.; Strnad, M.; Kraemer, U.; Schmuelling, T. Root-Specific Reduction of Cytokinin Causes Enhanced Root Growth, Drought Tolerance, and Leaf Mineral Enrichment in Arabidopsis and Tobacco. Plant Cell 2010, 22, 3905–3920. [Google Scholar]
- Ohkama, N.; Takei, K.; Sakakibara, H.; Hayashi, H.; Yoneyama, T.; Fujiwara, T. Regulation of sulfur-responsive gene expression by exogenously applied cytokinins in Arabidopsis thaliana. Plant Cell Physiol. 2002, 43, 1493–1501. [Google Scholar]
- Nikiforova, V.; Freitag, J.; Kempa, S.; Adamik, M.; Hesse, H.; Hoefgen, R. Transcriptome analysis of sulfur depletion in Arabidopsis thaliana: Interlacing of biosynthetic pathways provides response specificity. Plant J. 2003, 33, 633–650. [Google Scholar]
- Vorwerk, S.; Biernacki, S.; Hillebrand, H.; Janzik, I.; Müller, A.; Weiler, E.W.; Piotrowski, M. Enzymatic characterization of the recombinant Arabidopsis thaliana nitrilase subfamily encoded by the NIT2/NIT1/NIT3-gene cluster. Planta 2001, 212, 508–516. [Google Scholar]
- Nikiforova, V.J.; Daub, C.O.; Hesse, H.; Willmitzer, L.; Hoefgen, R. Integrative gene-metabolite network with implemented causality deciphers informational fluxes of sulphur stress response. J. Exp. Bot. 2005, 56, 1887–1896. [Google Scholar]
- Hoefgen, R.; Nikiforova, V.J. Metabolomics integrated with transcriptomics: Assessing systems response to sulfur-deficiency stress. Physiol. Plant. 2008, 132, 190–198. [Google Scholar]
- Rogg, L.E.; Lasswell, J.; Bartel, B. A gain-of-function mutation in IAA28 suppresses lateral root development. Plant Cell 2001, 13, 465–480. [Google Scholar]
- Falkenberg, B.; Witt, I.; Zanor, M.I.; Steinhauser, D.; Mueller-Roeber, B.; Hesse, H.; Hoefgen, R. Transcription factors relevant to auxin signalling coordinate broad-spectrum metabolic shifts including sulphur metabolism. J. Exp. Bot. 2008, 59, 2831–2846. [Google Scholar]
- Moniuszko, G.; Skoneczny, M.; Zientara-Rytter, K.; Wawrzynska, A.; Glow, D.; Cristescu, S.M.; Harren, F.J.M.; Sirko, A. Tobacco LSU-like protein couples sulphur-deficiency response with ethylene signalling pathway. J. Exp. Bot. 2013, 64, 5173–5182. [Google Scholar]
- Sirko, A.; Wawrzynska, A.; Rodriguez, M.C.; Sektas, P. The family of LSU-like proteins. Front. Plant Sci. 2015, 5, 774. [Google Scholar]
- Maruyama-Nakashita, A.; Inoue, E.; Watanabe-Takahashi, A.; Yarnaya, T.; Takahashi, H. Transcriptome profiling of sulfur-responsive genes in Arabidopsis reveals global effects of sulfur nutrition on multiple metabolic pathways. Plant Physiol. 2003, 132, 597–605. [Google Scholar]
- Jost, R.; Altschmied, L.; Bloem, E.; Bogs, J.; Gershenzon, J.; Hahnel, U.; Hansch, R.; Hartmann, T.; Kopriva, S.; Kruse, C.; et al. Expression profiling of metabolic genes in response to methyl jasmonate reveals regulation of genes of primary and secondary sulfur-related pathways in Arabidopsis thaliana. Photosynth. Res. 2005, 86, 491–508. [Google Scholar] [PubMed]
- Koprivova, A.; North, K.A.; Kopriva, S. Complex signaling network in regulation of adenosine 5′-phosphosulfate reductase by salt stress in Arabidopsis roots. Plant Physiol. 2008, 146, 1408–1420. [Google Scholar] [PubMed] [Green Version]
- Hubberten, H.M.; Drozd, A.; Tran, B.V.; Hesse, H.; Hoefgen, R. Local and systemic regulation of sulfur homeostasis in roots of Arabidopsis thaliana. Plant J. 2012, 72, 625–635. [Google Scholar] [PubMed]
- Hubberten, H.-M.; Watanabe, M.; Bielecka, M.; Heyneke, E.; Aarabi, F.; Hoefgen, R. More Than a Substrate: The O-Acetylserine Responsive Transcriptome. In Molecular Physiology and Ecophysiology of Sulfur; De Kok, L.J., Hawkesford, M.J., Rennenberg, H., Saito, K., Schnug, E., Eds.; Springer: Cham, Switzerland, 2015; pp. 133–143. [Google Scholar]
- Buchner, P.; Stuiver, C.E.E.; Westerman, S.; Wirtz, M.; Hell, R.; Hawkesford, M.J.; De Kok, L.J. Regulation of sulfate uptake and expression of sulfate transporter genes in Brassica oleracea as affected by atmospheric H2S and pedospheric sulfate nutrition. Plant Physiol. 2004, 136, 3396–3408. [Google Scholar] [PubMed] [Green Version]
- Dong, Y.; Silbermann, M.; Speiser, A.; Forieri, I.; Linster, E.; Poschet, G.; Samami, A.A.; Wanatabe, M.; Sticht, C.; Teleman, A.A.; et al. Sulfur availability regulates plant growth via glucose-TOR signaling. Nature Commun. 2017, 8, 1174. [Google Scholar]
- Courbet, G.; Gallardo, K.; Vigani, G.; Brunel-Muguet, S.; Trouverie, J.; Salon, C.; Ourry, A. Disentangling the complexity and diversity of crosstalk between sulfur and other mineral nutrients in cultivated plants. J. Exp. Bot. 2019, 70, 4183–4196. [Google Scholar]
- Jobe, T.O.; Zenzen, I.; Rahimzadeh Karvansara, P.; Kopriva, S. Integration of sulfate assimilation with carbon and nitrogen metabolism in transition from C3 to C4 photosynthesis. J. Exp. Bot. 2019, 70, 4211–4221. [Google Scholar]
- Koprivova, A.; Suter, M.; Op den Camp, R.; Brunold, C.; Kopriva, S. Regulation of sulfate assimilation by nitrogen in Arabidopsis. Plant Physiol. 2000, 122, 737–746. [Google Scholar]
- Kopriva, S. Nitrogen and Sulfur Metabolism in C4 Plants. In C4 Photosynthesis and Related CO2 Concentrating Mechanisms; Raghavendra, A., Sage, R., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 32, pp. 109–128. [Google Scholar]
- Hesse, H.; Nikiforova, V.; Gakiere, B.; Hoefgen, R. Molecular analysis and control of cysteine biosynthesis: Integration of nitrogen and sulphur metabolism. J. Exp. Bot. 2004, 55, 1283–1292. [Google Scholar]
- Prosser, I.M.; Purves, J.V.; Saker, L.R.; Clarkson, D.T. Rapid disruption of nitrogen metabolism and nitrate transport in spinach plants deprived of sulphate. J. Exp. Bot. 2001, 52, 113–121. [Google Scholar]
- Hawkesford, M.J.; De Kok, L.J. Managing sulphur metabolism in plants. Plant Cell Environ. 2006, 29, 382–395. [Google Scholar] [PubMed]
- Kopriva, S.; Suter, M.; von Ballmoos, P.; Hesse, H.; Krahenbuhl, U.; Rennenberg, H.; Brunold, C. Interaction of sulfate assimilation with carbon and nitrogen metabolism in Lemna minor. Plant Physiol. 2002, 130, 1406–1413. [Google Scholar] [PubMed] [Green Version]
- Hesse, H.; Trachsel, N.; Suter, M.; Kopriva, S.; von Ballmoos, P.; Rennenberg, H.; Brunold, C. Effect of glucose on assimilatory sulphate reduction in Arabidopsis thaliana roots. J. Exp. Bot. 2003, 54, 1701–1709. [Google Scholar] [PubMed] [Green Version]
- Allahham, A.; Kanno, S.; Zhang, L.; Maruyama-Nakashita, A. Sulfur Deficiency Increases Phosphate Accumulation, Uptake, and Transport in Arabidopsis thaliana. Int. J. Mol. Sci. 2020, 21, 2971. [Google Scholar]
- Sugimoto, K.; Sato, N.; Tsuzuki, M. Utilization of a chloroplast membrane sulfolipid as a major internal sulfur source for protein synthesis in the early phase of sulfur starvation in Chlamydomonas reinhardtii. FEBS Lett. 2007, 581, 4519–4522. [Google Scholar] [PubMed] [Green Version]
- Smith, F.W.; Rae, A.L.; Hawkesford, M.J. Molecular mechanisms of phosphate and sulphate transport in plants. Biochim. Biophys. Acta Biomembr. 2000, 1465, 236–245. [Google Scholar]
- Franco-Zorrilla, J.M.; Martin, A.C.; Leyva, A.; Par-Ares, J.P. Interaction between phosphate-starvation, sugar, and cytokinin signaling in Arabidopsis and the roles of cytokinin receptors CRE1/AHK4 and AHK3. Plant Physiol. 2005, 138, 847–857. [Google Scholar]
- Hsieh, L.-C.; Lin, S.-I.; Shih, A.C.-C.; Chen, J.-W.; Lin, W.-Y.; Tseng, C.-Y.; Li, W.-H.; Chiou, T.-J. Uncovering Small RNA-Mediated Responses to Phosphate Deficiency in Arabidopsis by Deep Sequencing. Plant Physiol. 2009, 151, 2120–2132. [Google Scholar]
- Misson, J.; Raghothama, K.G.; Jain, A.; Jouhet, J.; Block, M.A.; Bligny, R.; Ortet, P.; Creff, A.; Somerville, S.; Rolland, N.; et al. A genome-wide transcriptional analysis using Arabidopsis thaliana Affymetrix gene chips determined plant responses to phosphate deprivation. Proc. Natl. Acad. Sci. USA 2005, 102, 11934–11939. [Google Scholar]
- Rouached, H.; Secco, D.; Arpat, B.; Poirier, Y. The transcription factor PHR1 plays a key role in the regulation of sulfate shoot-to-root flux upon phosphate starvation in Arabidopsis. BMC Plant Biol. 2011, 11, 19. [Google Scholar]
- Moseley, J.L.; Gonzalez-Ballester, D.; Pootakham, W.; Bailey, S.; Grossman, A.R. Genetic Interactions Between Regulators of Chlamydomonas Phosphorus and Sulfur Deprivation Responses. Genetics 2009, 181, 889–905. [Google Scholar] [PubMed] [Green Version]
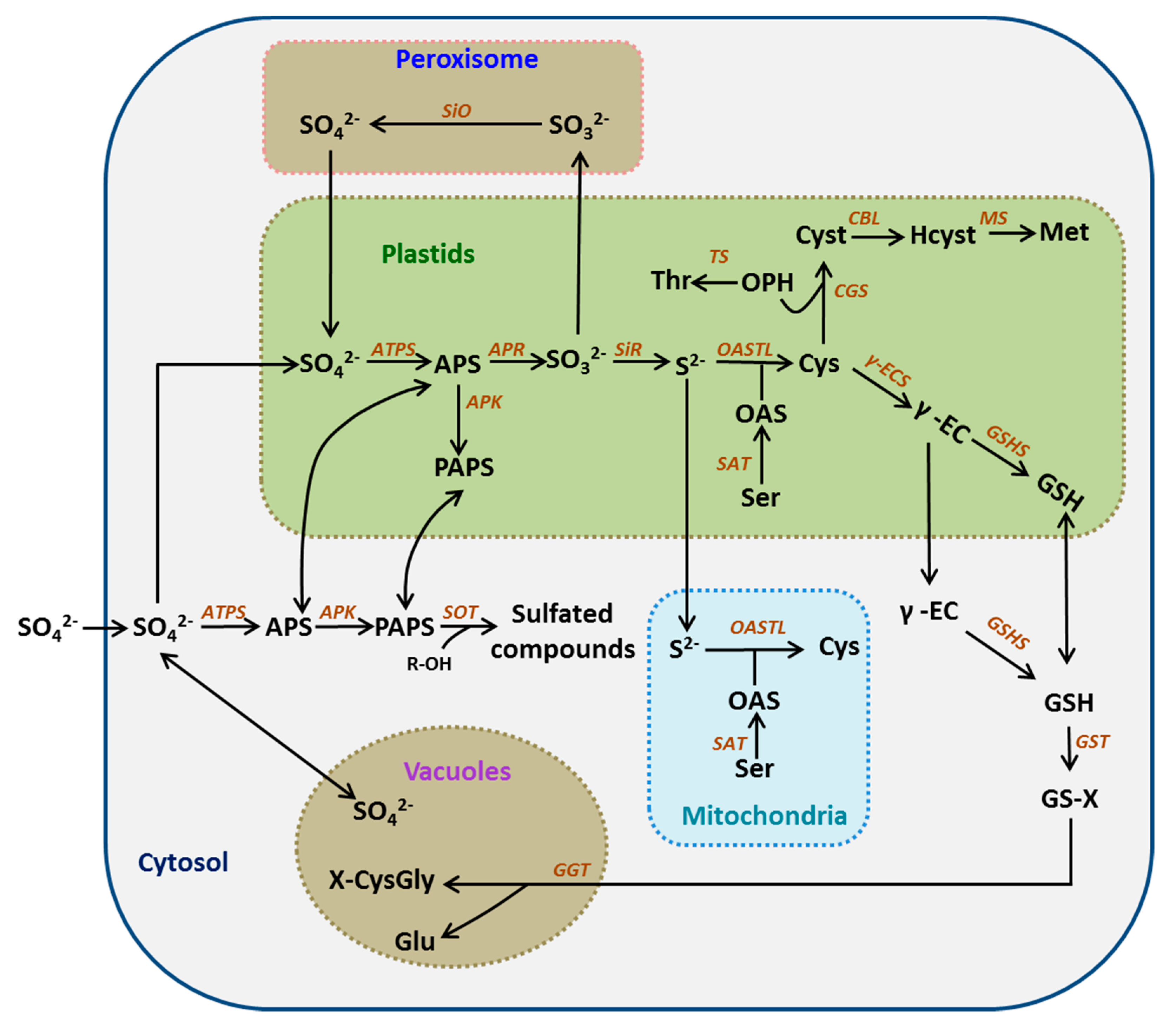
Figure 1.
The sulfate assimilation and metabolism pathway. Enzymes are indicated in orange letters. Abbreviations of metabolites: APS, adenosine 5′-phosphosulfate; PAPS, 3′-phosphoadenosine 5′-phosphosulfate; R-OH, hydroxylated precursor; Ser, serine; OAS, O-acetylserine; Cys, cysteine; OPH, O-phosphohomoserine; Thr, threonine; Cyst, cystathionine; Hcyst, homocysteine; Met, methionine; γ-EC, γ-glutamylcysteine; GSH, glutathione; GS-X, glutathione conjugate; Glu, glutamate; X-CysGly, cysteinylglycine conjugate. Abbreviations of enzymes: ATPS, ATP sufurylase; APK, APS kinase; SOT, sulfotransferase; APR, APS reductase; SiO, sulphite oxidase; SiR, sulfite reductase; SAT, serine acetyltransferase; OAS-TL, OAS(thiol)lyase; CGS, cystathionine γ-synthase; TS, threonine synthase; CBL, cystathionine β-lyase; MS, methionine synthase; γ-ECS, γ-glutamylcysteine synthetase; GSHS, glutathione synthetase; GST, glutathione-S-transferase; GGT, γ-glutamyltransferase.
Figure 1.
The sulfate assimilation and metabolism pathway. Enzymes are indicated in orange letters. Abbreviations of metabolites: APS, adenosine 5′-phosphosulfate; PAPS, 3′-phosphoadenosine 5′-phosphosulfate; R-OH, hydroxylated precursor; Ser, serine; OAS, O-acetylserine; Cys, cysteine; OPH, O-phosphohomoserine; Thr, threonine; Cyst, cystathionine; Hcyst, homocysteine; Met, methionine; γ-EC, γ-glutamylcysteine; GSH, glutathione; GS-X, glutathione conjugate; Glu, glutamate; X-CysGly, cysteinylglycine conjugate. Abbreviations of enzymes: ATPS, ATP sufurylase; APK, APS kinase; SOT, sulfotransferase; APR, APS reductase; SiO, sulphite oxidase; SiR, sulfite reductase; SAT, serine acetyltransferase; OAS-TL, OAS(thiol)lyase; CGS, cystathionine γ-synthase; TS, threonine synthase; CBL, cystathionine β-lyase; MS, methionine synthase; γ-ECS, γ-glutamylcysteine synthetase; GSHS, glutathione synthetase; GST, glutathione-S-transferase; GGT, γ-glutamyltransferase.

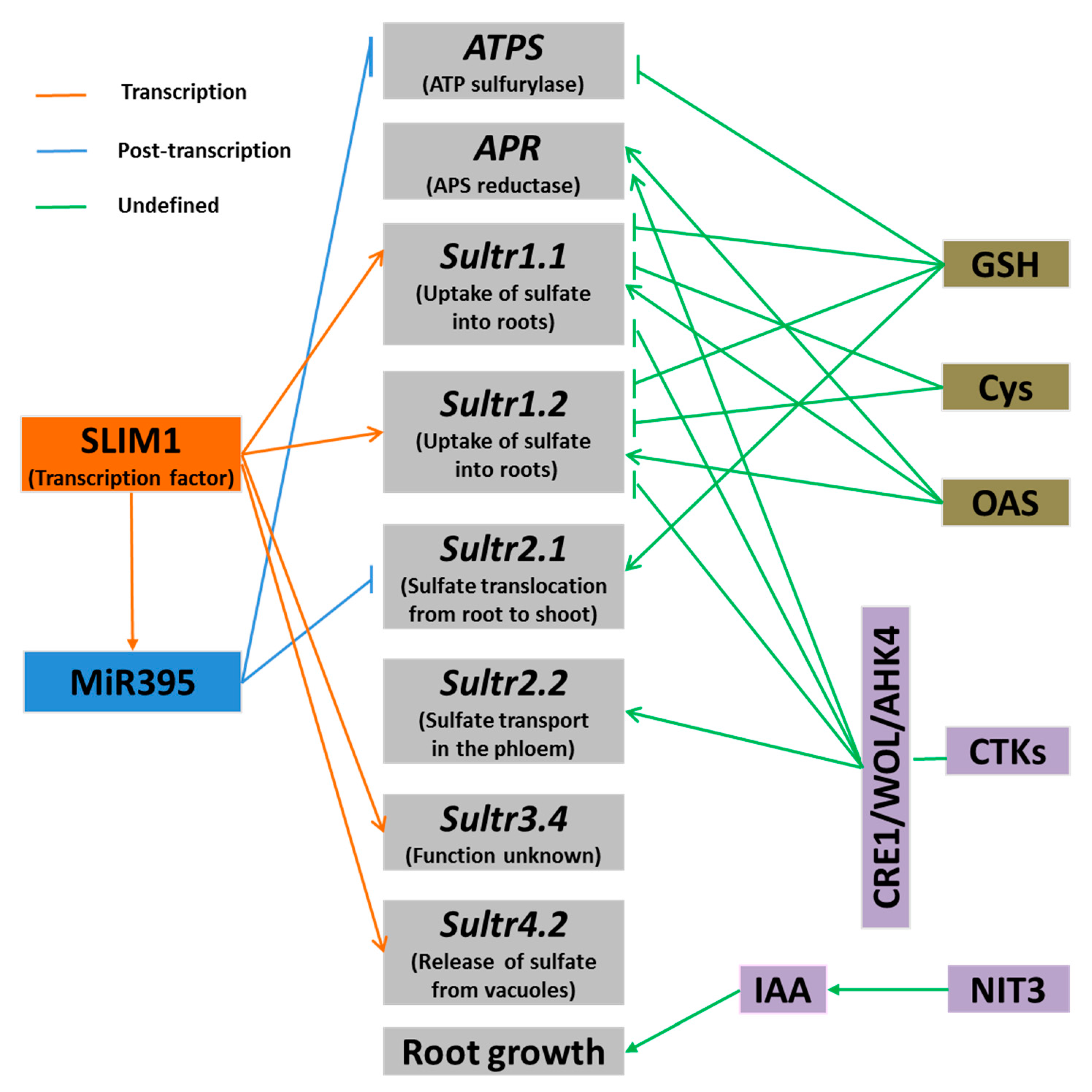
Figure 2.
Regulatory pathways of plant’s response to sulfur deficiency. Arrows denote positive effects, whereas lines ending with a short bar indicate negative effects.
Figure 2.
Regulatory pathways of plant’s response to sulfur deficiency. Arrows denote positive effects, whereas lines ending with a short bar indicate negative effects.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, Q.; Gao, Y.; Yang, A. Sulfur Homeostasis in Plants. Int. J. Mol. Sci. 2020, 21, 8926. https://doi.org/10.3390/ijms21238926
AMA Style
Li Q, Gao Y, Yang A. Sulfur Homeostasis in Plants. International Journal of Molecular Sciences. 2020; 21(23):8926. https://doi.org/10.3390/ijms21238926
Chicago/Turabian StyleLi, Qian, Yan Gao, and An Yang. 2020. "Sulfur Homeostasis in Plants" International Journal of Molecular Sciences 21, no. 23: 8926. https://doi.org/10.3390/ijms21238926
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.