Magnetic Resonance Imaging Correlates of White Matter Gliosis and Injury in Preterm Fetal Sheep Exposed to Progressive Systemic Inflammation

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
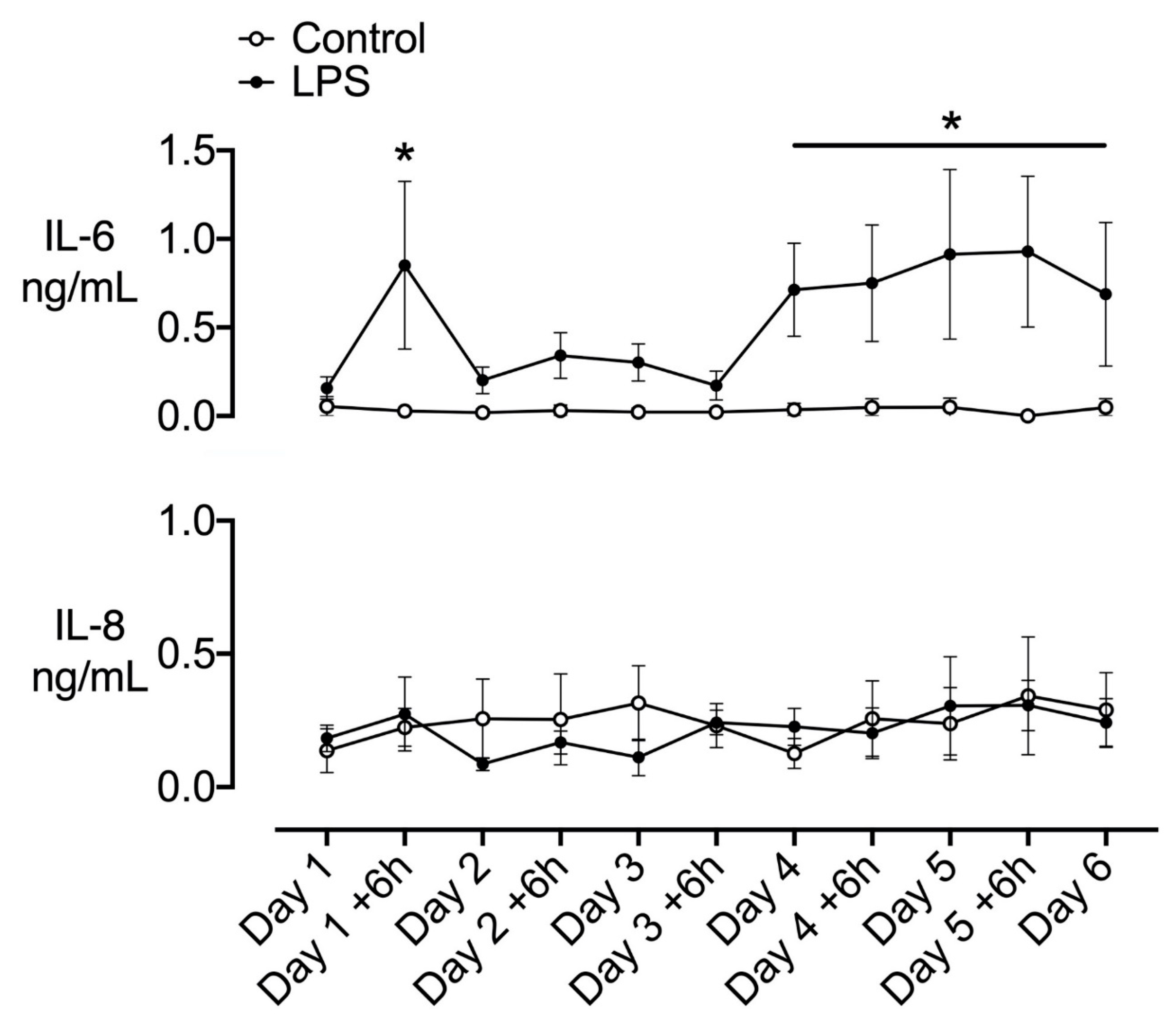
2.1. Plasma Cytokines
2.2. Postmortem Findings
2.3. Histopathology
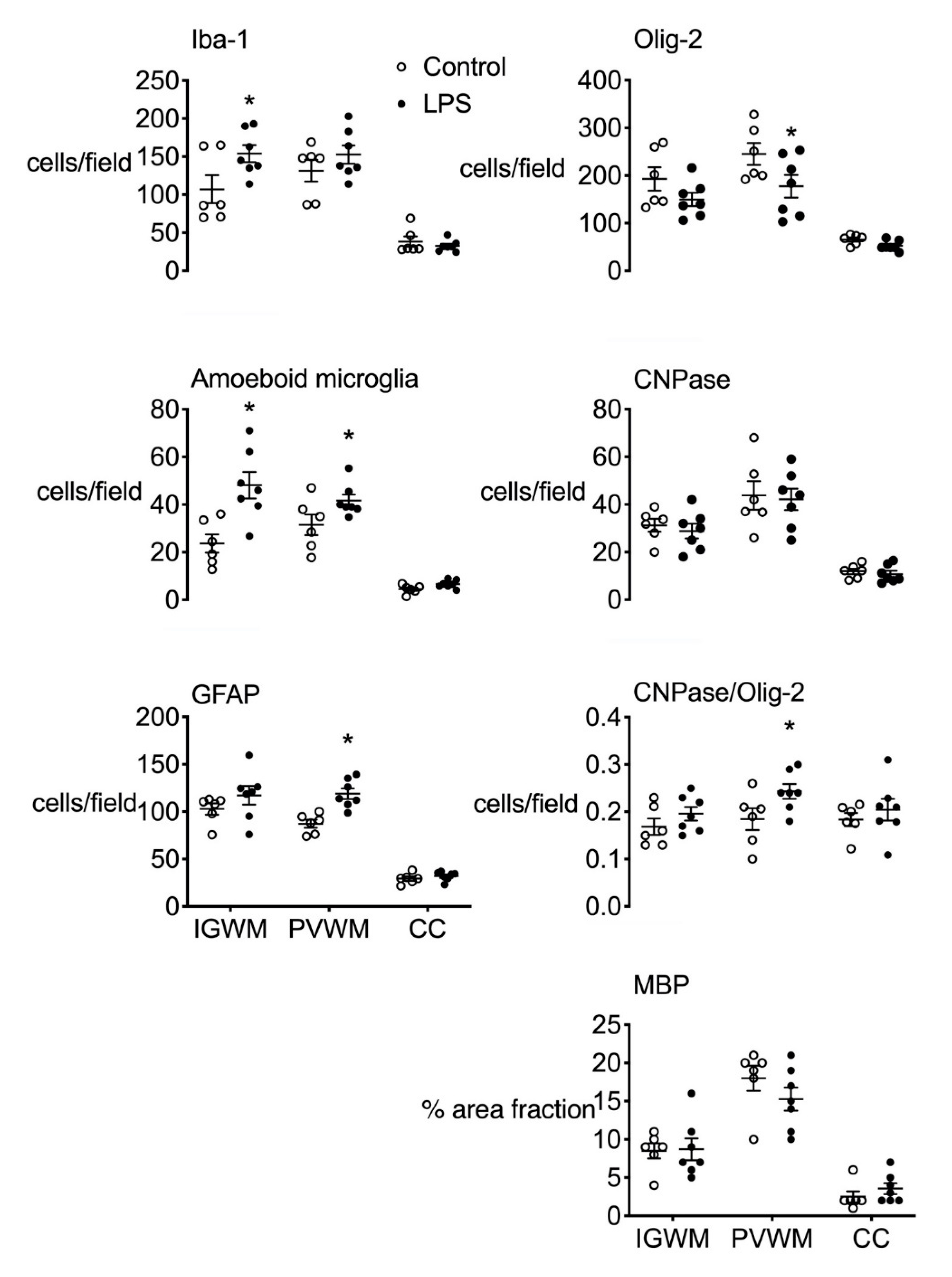
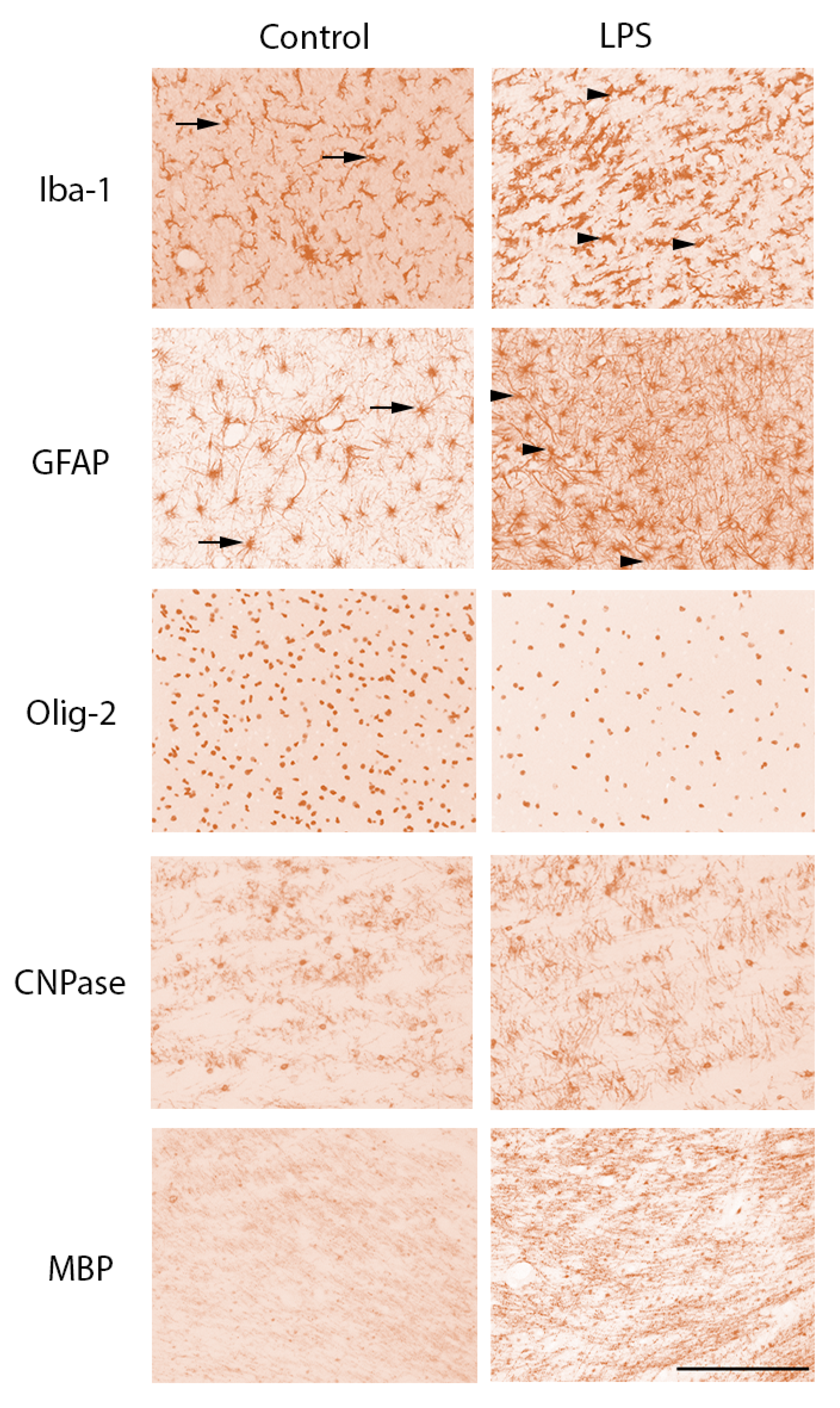
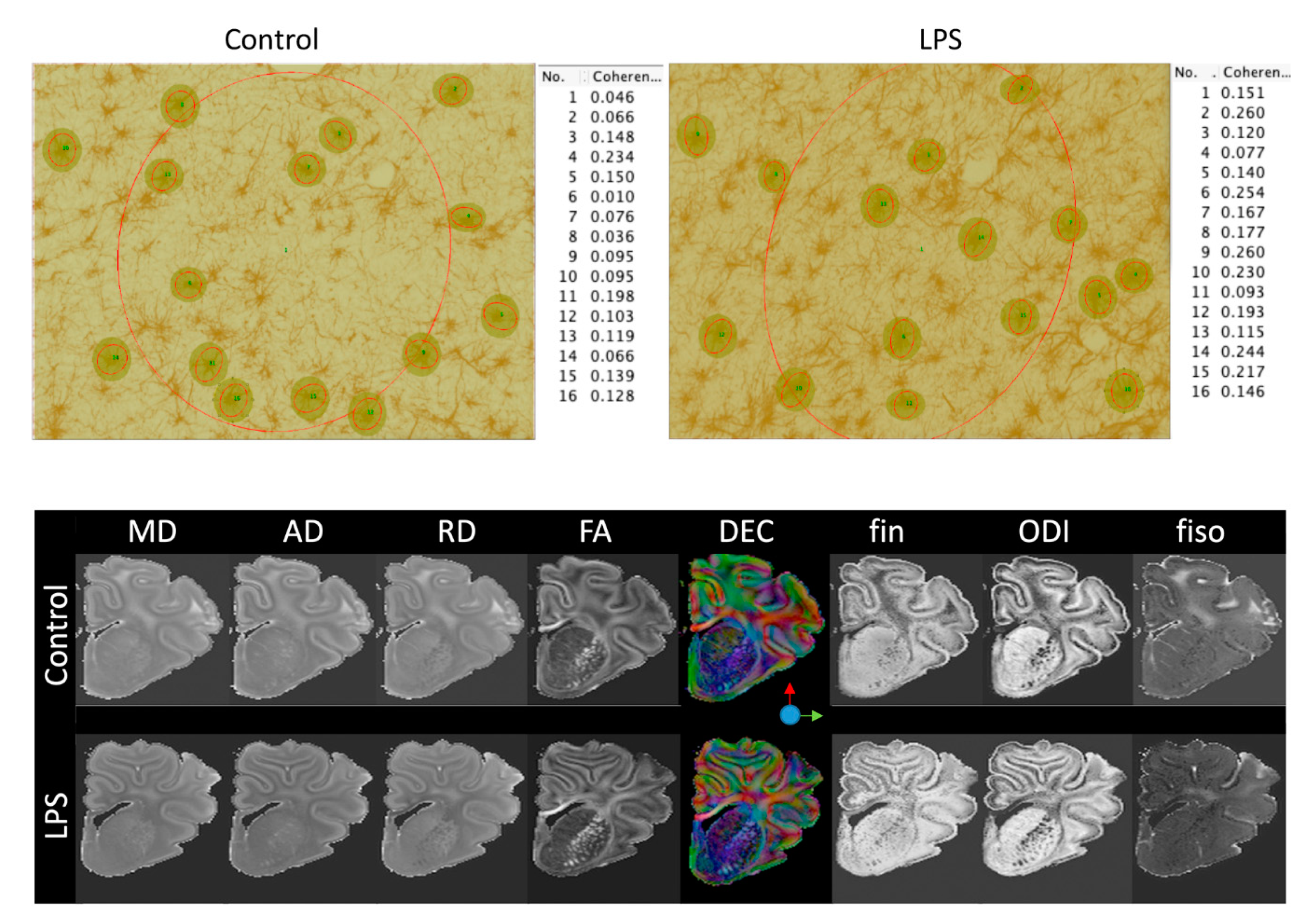
2.3.1. Gliosis
2.3.2. White Matter Development
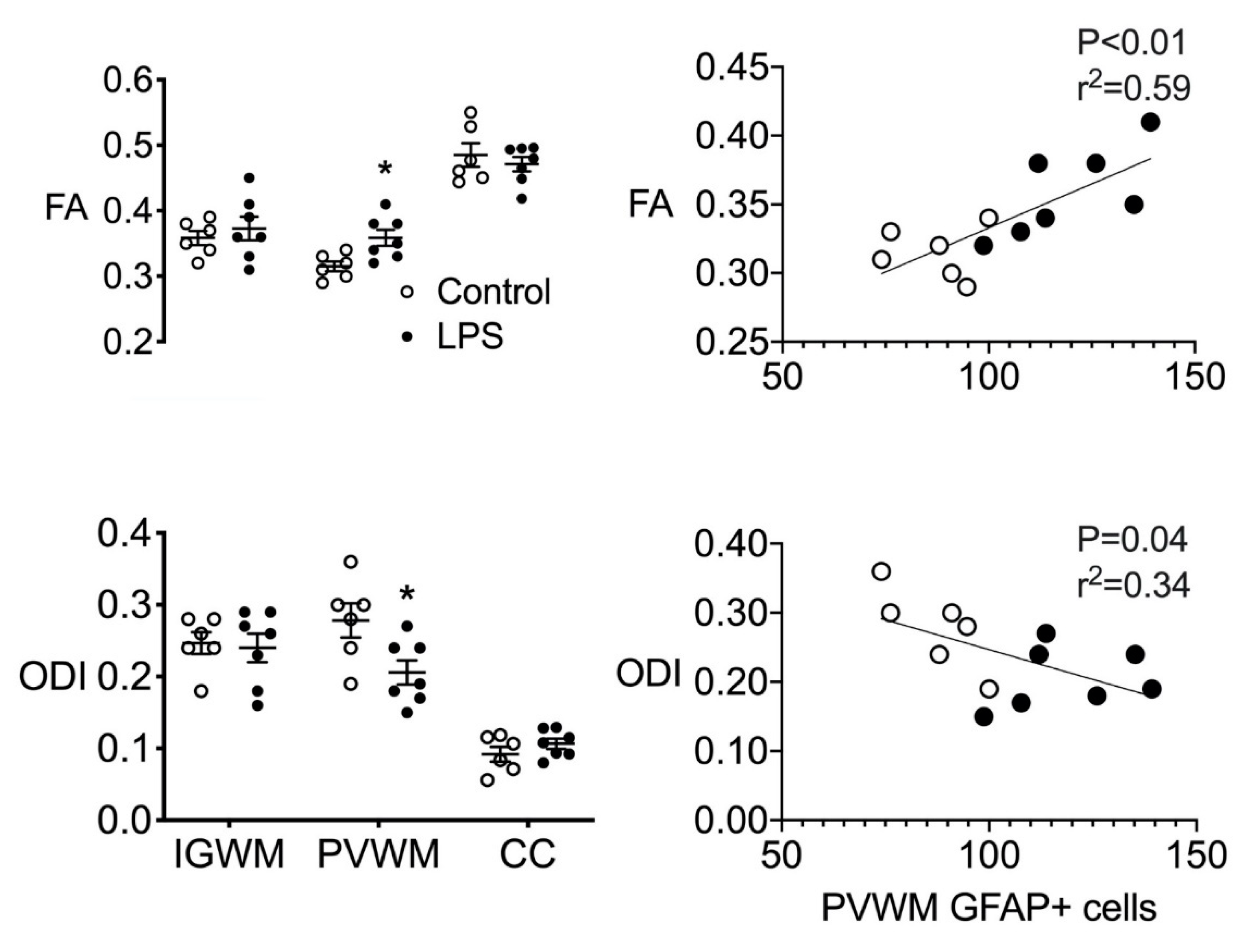
2.4. Ex Vivo MRI
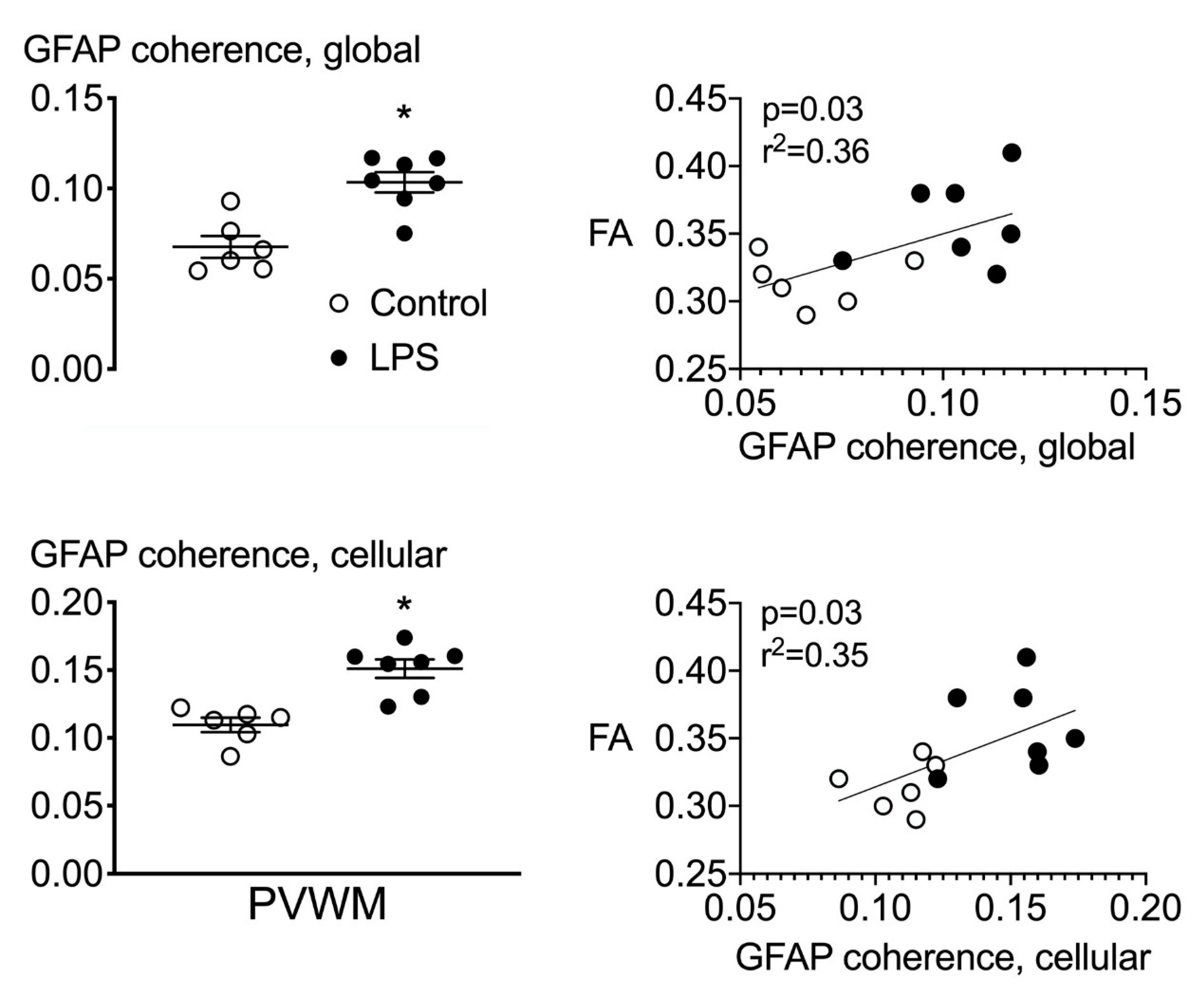
2.5. Astrocyte Coherence Analysis
2.6. Correlations
2.7. Fetal Biochemical and Physiological Outcomes
2.7.1. Baseline Period
2.7.2. Fetal Biochemistry
2.7.3. Carotid and Femoral Arterial Blood Flows and Vascular Conductance
2.7.4. EEG Activity and Fetal Movement
2.7.5. Fetal Heart Rate and Mean Arterial Pressure
3. Discussion
4. Materials and Methods
4.1. Instrumentation
4.2. Experimental Recordings
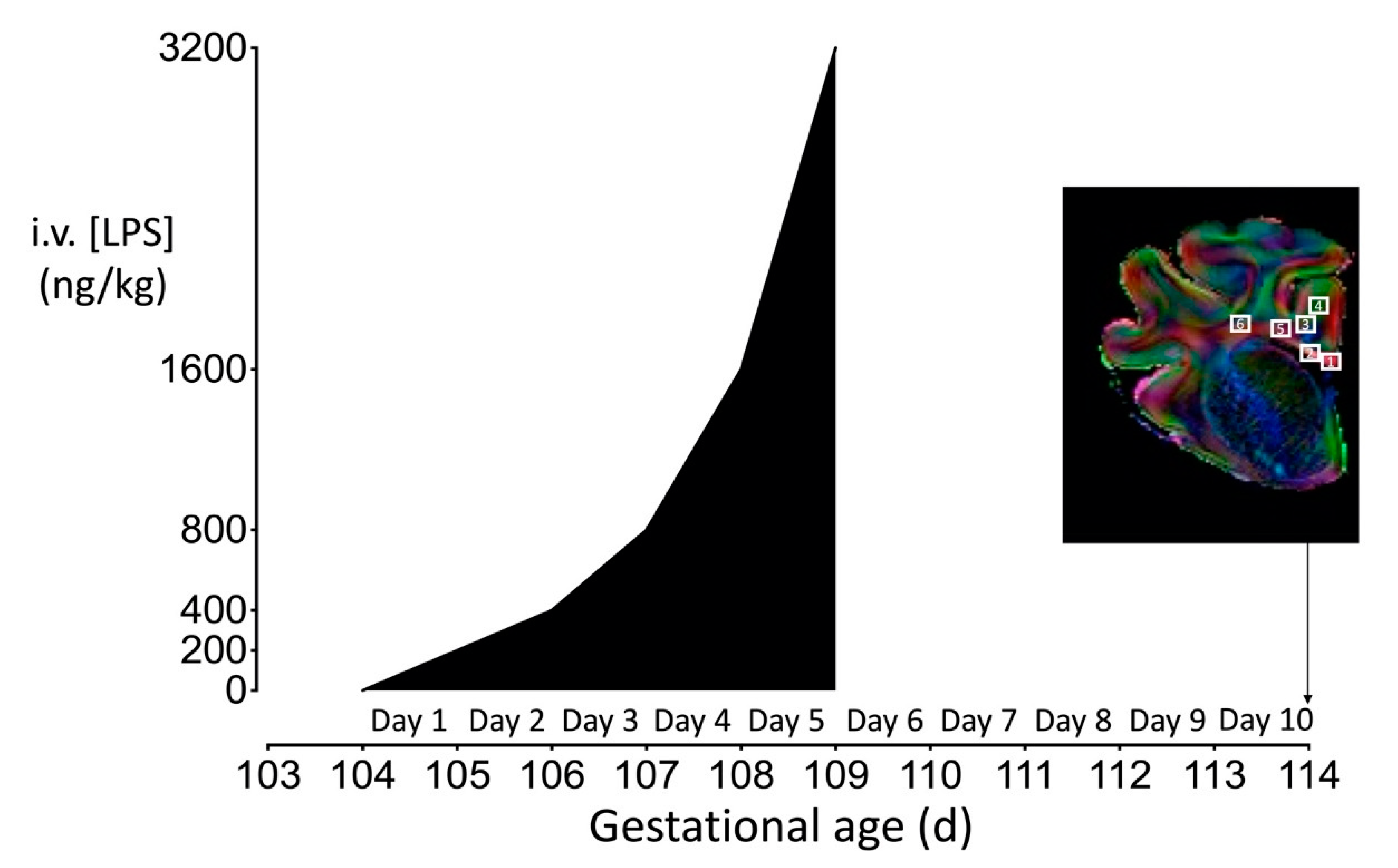
4.3. Experimental Protocol
4.4. Fetal Cytokine Measurements
4.5. Histopathology
4.6. Ex Vivo Magnetic Resonance Imaging
4.7. Data Analysis and Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CaBF | carotid arterial blood flow |
| CaVC | carotid arterial vascular conductance |
| CNPase | 2′,-3′-cyclic-nuceotide 3′-phosphodiesterase |
| EEG | electroencephalogram |
| FBF | femoral arterial blood flow |
| FHR | fetal heart rate |
| FVC | femoral arterial vascular conductance |
| GFAP | glial fibrillary acidic protein |
| Iba-1 | ionized calcium-binding adapter molecule 1 |
| IGWM | intragyral white matter |
| IL | interleukin |
| LPS | lipopolysaccharide |
| MAP | mean arterial blood pressure |
| MBP | myelin basic protein |
| Olig-2 | oligodendrocyte transcription factor 2 |
| PBS | phosphate-buffered saline |
| PVWM | periventricular white matter |
References
- Honeycutt, A.; Dunlap, L.; Chen, H.; Al Homsi, G.; Grosse, S.; Schendel, D. Economic costs associated with mental retardation, cerebral palsy, hearing loss, and vision impairment: United States, 2003. MMWR Morb. Mortal. Wkly. Rep. 2004, 53, 57–59. [Google Scholar]
- Korvenranta, E.; Lehtonen, L.; Rautava, L.; Häkkinen, U.; Andersson, S.; Gissler, M.; Hallman, M.; Leipälä, J.; Peltola, M.; Tammela, O.; et al. Impact of very preterm birth on health care costs at five years of age. Pediatrics 2010, 125, e1109–e1114. [Google Scholar] [CrossRef] [PubMed]
- Shih, S.T.F.; Tonmukayakul, U.; Imms, C.; Reddihough, D.; Graham, H.K.; Cox, L.; Carter, R. Economic evaluation and cost of interventions for cerebral palsy: A systematic review. Dev. Med. Child. Neurol. 2018, 60, 543–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Vliet, E.O.; de Kieviet, J.F.; Oosterlaan, J.; van Elburg, R.M. Perinatal infections and neurodevelopmental outcome in very preterm and very low-birth-weight infants: A meta-analysis. JAMA Pediatr. 2013, 167, 662–668. [Google Scholar] [CrossRef]
- Kuban, K.C.; Joseph, R.M.; O’Shea, T.M.; Heeren, T.; Fichorova, R.N.; Douglass, L.; Jara, H.; Frazier, J.A.; Hirtz, D.; Rollins, J.V.; et al. Extremely Low Gestational Age Newborns (ELGAN) Study Investigators, Circulating inflammatory-associated proteins in the first month of life and cognitive impairment at age 10 years in children born extremely preterm. J. Pediatr. 2017, 180, 116–123.e1. [Google Scholar] [CrossRef] [Green Version]
- Galinsky, R.; Lear, C.A.; Dean, J.M.; Wassink, G.; Dhillon, S.K.; Fraser, M.; Davidson, J.O.; Bennet, L.; Gunn, A.J. Complex interactions between hypoxia-ischemia and inflammation in preterm brain injury. Dev. Med. Child. Neurol. 2018, 60, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Riddle, A.; Dean, J.; Buser, J.R.; Gong, X.; Maire, J.; Chen, K.; Ahmad, T.; Cai, V.; Nguyen, T.; Kroenke, C.D.; et al. Histopathological correlates of magnetic resonance imaging-defined chronic perinatal white matter injury. Ann. Neurol. 2011, 70, 493–507. [Google Scholar] [CrossRef]
- Buser, J.R.; Maire, J.; Riddle, A.; Gong, X.; Nguyen, T.; Nelson, K.; Luo, N.L.; Ren, J.; Struve, J.; Sherman, L.S.; et al. Arrested preoligodendrocyte maturation contributes to myelination failure in premature infants. Ann. Neurol. 2012, 71, 93–109. [Google Scholar] [CrossRef]
- Rathbone, R.; Counsell, S.J.; Kapellou, O.; Dyet, L.; Kennea, N.; Hajnal, J.; Allsop, J.M.; Cowan, F.; Edwards, A.D. Perinatal cortical growth and childhood neurocognitive abilities. Neurology 2011, 77, 1510–1517. [Google Scholar] [CrossRef] [Green Version]
- Meng, C.; Bauml, J.G.; Daamen, M.; Jaekel, J.; Neitzel, J.; Scheef, L.; Busch, B.; Baumann, N.; Boecker, H.; Zimmer, C.; et al. Extensive and interrelated subcortical white and gray matter alterations in preterm-born adults. Brain Struct. Funct. 2016, 221, 2109–2121. [Google Scholar] [CrossRef]
- Hagberg, H.; Mallard, C.; Ferriero, D.M.; Vannucci, S.J.; Levison, S.W.; Vexler, Z.S.; Gressens, P. The role of inflammation in perinatal brain injury. Nat. Rev. Neurol. 2015, 11, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Galinsky, R.; Polglase, G.R.; Hooper, S.B.; Black, M.J.; Moss, T.J. The consequences of chorioamnionitis: Preterm birth and effects on development. J. Pregnancy 2013, 2013, 412831. [Google Scholar] [CrossRef] [Green Version]
- Soraisham, A.S.; Trevenen, C.; Wood, S.; Singhal, N.; Sauve, R. Histological chorioamnionitis and neurodevelopmental outcome in preterm infants. J. Perinatol. 2013, 33, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Anblagan, D.; Pataky, R.; Evans, M.J.; Telford, E.J.; Serag, A.; Sparrow, S.; Piyasena, C.; Semple, S.I.; Wilkinson, A.G.; Bastin, M.E.; et al. Association between preterm brain injury and exposure to chorioamnionitis during fetal life. Sci. Rep. 2016, 6, 37932. [Google Scholar] [CrossRef] [PubMed]
- Kapellou, O.; Counsell, S.J.; Kennea, N.; Dyet, L.; Saeed, N.; Stark, J.; Maalouf, E.; Duggan, P.; Ajayi-Obe, M.; Hajnal, J.; et al. Abnormal cortical development after premature birth shown by altered allometric scaling of brain growth. PLoS Med. 2006, 3, e265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomason, M.E.; Scheinost, D.; Manning, J.H.; Grove, L.E.; Hect, J.; Marshall, N.; Hernandez-Andrade, E.; Berman, S.; Pappas, A.; Yeo, L.; et al. Weak functional connectivity in the human fetal brain prior to preterm birth. Sci. Rep. 2017, 7, 39286. [Google Scholar] [CrossRef] [Green Version]
- Keogh, M.J.; Bennet, L.; Drury, P.P.; Booth, L.C.; Mathai, S.; Naylor, A.S.; Fraser, M.; Gunn, A.J. Subclinical exposure to low-dose endotoxin impairs EEG maturation in preterm fetal sheep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R270–R278. [Google Scholar] [CrossRef] [Green Version]
- Favrais, G.; van de Looij, Y.; Fleiss, B.; Ramanantsoa, N.; Bonnin, P.; Stoltenburg-Didinger, G.; Lacaud, A.; Saliba, E.; Dammann, O.; Gallego, J.; et al. Systemic inflammation disrupts the developmental program of white matter. Ann. Neurol. 2011, 70, 550–565. [Google Scholar] [CrossRef]
- Sweeney, E.L.; Dando, S.J.; Kallapur, S.G.; Knox, C.L. The Human Ureaplasma Species as Causative Agents of Chorioamnionitis. Clin. Microbiol. Rev. 2017, 30, 349–379. [Google Scholar] [CrossRef] [Green Version]
- Buhimschi, C.S.; Abdel-Razeq, S.; Cackovic, M.; Pettker, C.M.; Dulay, A.T.; Bahtiyar, M.O.; Zambrano, E.; Martin, R.; Norwitz, E.R.; Bhandari, V.; et al. Fetal heart rate monitoring patterns in women with amniotic fluid proteomic profiles indicative of inflammation. Am. J. Perinatol. 2008, 25, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Moss, T.J.; Nitsos, I.; Ikegami, M.; Jobe, A.H.; Newnham, J.P. Experimental intrauterine Ureaplasma infection in sheep. Am. J. Obstet. Gynecol. 2005, 192, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.W.; Park, C.W.; Moon, K.C.; Park, J.S.; Jun, J.K. The relationship among the progression of inflammation in umbilical cord, fetal inflammatory response, early-onset neonatal sepsis, and chorioamnionitis. PLoS ONE 2019, 14, e0225328. [Google Scholar] [CrossRef] [PubMed]
- Kuban, K.C.K.; Jara, H.; O’Shea, T.M.; Heeren, T.; Joseph, R.M.; Fichorova, R.N.; Alshamrani, K.; Aakil, A.; Beaulieu, F.; Horn, M.; et al. Association of Circulating Proinflammatory and Anti-inflammatory Protein Biomarkers in Extremely Preterm Born Children with Subsequent Brain Magnetic Resonance Imaging Volumes and Cognitive Function at Age 10 Years. J. Pediatr. 2019, 210, 81–90.e3. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, G.H.; Baghurst, K.I.; Potter, B.J.; Hetzel, B.S. Foetal brain development in the sheep. Neuropathol. Appl. Neurobiol. 1979, 5, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, R.S.; Romero, R.; Hillier, S.L.; Eschenbach, D.A.; Sweet, R.L. A review of premature birth and subclinical infection. Am. J. Obstet. Gynecol. 1992, 166, 1515–1528. [Google Scholar] [CrossRef]
- Leviton, A.; Paneth, N.; Reuss, M.L.; Susser, M.; Allred, E.N.; Dammann, O.; Kuban, K.; Van Marter, L.J.; Pagano, M.; Hegyi, T.; et al. Maternal infection, fetal inflammatory response, and brain damage in very low birth weight infants. Developmental Epidemiology Network Investigators. Pediatr. Res. 1999, 46, 566–575. [Google Scholar] [CrossRef]
- Küster, H.; Weiss, M.; Willeitner, A.E.; Detlefsen, S.; Jeremias, I.; Zbojan, J.; Geiger, R.; Lipowsky, G.; Simbruner, G. Interleukin-1 receptor antagonist and interleukin-6 for early diagnosis of neonatal sepsis 2 days before clinical manifestation. Lancet 1998, 352, 1271–1277. [Google Scholar] [CrossRef]
- Dean, J.M.; Shi, Z.; Fleiss, B.; Gunn, K.C.; Groenendaal, F.; van Bel, F.; Derrick, M.; Juul, S.E.; Tan, S.; Gressens, P.; et al. A Critical Review of Models of Perinatal Infection. Dev. Neurosci. 2015, 37, 289–304. [Google Scholar] [CrossRef]
- Spittler, A.; Razenberger, M.; Kupper, H.; Kaul, M.; Hackl, W.; Boltz-Nitulescu, G.; Függer, R.; Roth, E. Relationship between interleukin-6 plasma concentration in patients with sepsis, monocyte phenotype, monocyte phagocytic properties, and cytokine production. Clin. Infect. Dis. 2000, 31, 1338–1342. [Google Scholar] [CrossRef] [Green Version]
- Diepold, M.; Noellke, P.; Duffner, U.; Kontny, U.; Berner, R. Performance of Interleukin-6 and Interleukin-8 serum levels in pediatric oncology patients with neutropenia and fever for the assessment of low-risk. BMC Infect. Dis. 2008, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Boskabadi, H.; Maamouri, G.; Tavakol Afshari, J.; Mafinejad, S.; Hosseini, G.; Mostafavi-Toroghi, H.; Saber, H.; Ghayour-Mobarhan, M.; Ferns, G. Evaluation of serum interleukins-6, 8 and 10 levels as diagnostic markers of neonatal infection and possibility of mortality. Iran. J. Basic Med. Sci 2013, 16, 1232–1237. [Google Scholar] [PubMed]
- Galinsky, R.; Dhillon, S.K.; Dean, J.M.; Davidson, J.O.; Lear, C.A.; Wassink, G.; Nott, F.; Kelly, S.B.; Fraser, M.; Yuill, C.; et al. Tumor necrosis factor inhibition attenuates white matter gliosis after systemic inflammation in preterm fetal sheep. J. Neuroinflammation 2020, 17, 92. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.M.; van de Looij, Y.; Sizonenko, S.V.; Lodygensky, G.A.; Lazeyras, F.; Bolouri, H.; Kjellmer, I.; Huppi, P.S.; Hagberg, H.; Mallard, C. Delayed cortical impairment following lipopolysaccharide exposure in preterm fetal sheep. Ann. Neurol. 2011, 70, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hagberg, H.; Zhu, C.; Jacobsson, B.; Mallard, C. Effects of intrauterine inflammation on the developing mouse brain. Brain Res. 2007, 1144, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Galinsky, R.; Davidson, J.O.; Dean, J.M.; Green, C.R.; Bennet, L.; Gunn, A.J. Glia and hemichannels: Key mediators of perinatal encephalopathy. Neural Regen. Res. 2018, 13, 181–189. [Google Scholar] [PubMed]
- Takashima, S.; Tanaka, K. Development of cerebrovascular architecture and its relationship to periventricular leukomalacia. Arch. Neurol. 1978, 35, 11–16. [Google Scholar] [CrossRef]
- Inage, Y.W.; Itoh, M.; Takashima, S. Correlation between cerebrovascular maturity and periventricular leukomalacia. Pediatr. Neurol. 2000, 22, 204–208. [Google Scholar] [CrossRef]
- Miyawaki, T.; Matsui, K.; Takashima, S. Developmental characteristics of vessel density in the human fetal and infant brains. Early Hum. Dev. 1998, 53, 65–72. [Google Scholar] [CrossRef]
- Back, S.A.; Han, B.H.; Luo, N.L.; Chricton, C.A.; Xanthoudakis, S.; Tam, J.; Arvin, K.L.; Holtzman, D.M. Selective vulnerability of late oligodendrocyte progenitors to hypoxia-ischemia. J. Neurosci. 2002, 22, 455–463. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, S.; Furube, E.; Itoh, M.; Nakashima, T.; Miyata, S. Robust increase of microglia proliferation in the fornix of hippocampal axonal pathway after a single LPS stimulation. J. Neuroimmunol. 2015, 285, 31–40. [Google Scholar] [CrossRef]
- van Tilborg, E.; Achterberg, E.J.M.; van Kammen, C.M.; van der Toorn, A.; Groenendaal, F.; Dijkhuizen, R.M.; Heijnen, C.J.; Vanderschuren, L.; Benders, M.; Nijboer, C.H.A. Combined fetal inflammation and postnatal hypoxia causes myelin deficits and autism-like behavior in a rat model of diffuse white matter injury. Glia 2018, 66, 78–93. [Google Scholar] [CrossRef] [PubMed]
- van de Looij, Y.; Dean, J.M.; Gunn, A.J.; Huppi, P.S.; Sizonenko, S.V. Advanced magnetic resonance spectroscopy and imaging techniques applied to brain development and animal models of perinatal injury. Int. J. Dev. Neurosci. 2015, 45, 29–38. [Google Scholar] [CrossRef]
- Robitaille, P.M.; Abduljalil, A.M.; Kangarlu, A.; Zhang, X.; Yu, Y.; Burgess, R.; Bair, S.; Noa, P.; Yang, L.; Zhu, H.; et al. Human magnetic resonance imaging at 8 T. NMR Biomed. 1998, 11, 263–265. [Google Scholar] [CrossRef]
- Springer, E.; Dymerska, B.; Cardoso, P.L.; Robinson, S.D.; Weisstanner, C.; Wiest, R.; Schmitt, B.; Trattnig, S. Comparison of Routine Brain Imaging at 3 T and 7 T. Invest. Radiol. 2016, 51, 469–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budde, M.D.; Janes, L.; Gold, E.; Turtzo, L.C.; Frank, J.A. The contribution of gliosis to diffusion tensor anisotropy and tractography following traumatic brain injury: Validation in the rat using Fourier analysis of stained tissue sections. Brain 2011, 134 Pt 8, 2248–2260. [Google Scholar] [CrossRef] [Green Version]
- Mollink, J.; Kleinnijenhuis, M.; Cappellen van Walsum, A.V.; Sotiropoulos, S.N.; Cottaar, M.; Mirfin, C.; Heinrich, M.P.; Jenkinson, M.; Pallebage-Gamarallage, M.; Ansorge, O.; et al. Evaluating fibre orientation dispersion in white matter: Comparison of diffusion MRI, histology and polarized light imaging. Neuroimage 2017, 157, 561–574. [Google Scholar] [CrossRef] [PubMed]
- Nemanich, S.T.; Mueller, B.A.; Gillick, B.T. Neurite orientation dispersion and density imaging quantifies corticospinal tract microstructural organization in children with unilateral cerebral palsy. Hum. Brain Mapp. 2019, 40, 4888–4900. [Google Scholar] [CrossRef] [Green Version]
- Travis, K.E.; Adams, J.N.; Ben-Shachar, M.; Feldman, H.M. Decreased and Increased Anisotropy along Major Cerebral White Matter Tracts in Preterm Children and Adolescents. PLoS ONE 2015, 10, e0142860. [Google Scholar] [CrossRef]
- Fleiss, B.; Gressens, P. Tertiary mechanisms of brain damage: A new hope for treatment of cerebral palsy? Lancet Neurol. 2012, 11, 556–566. [Google Scholar] [CrossRef]
- Rose, S.E.; Hatzigeorgiou, X.; Strudwick, M.W.; Durbridge, G.; Davies, P.S.; Colditz, P.B. Altered white matter diffusion anisotropy in normal and preterm infants at term-equivalent age. Magn. Reson. Med. 2008, 60, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Anjari, M.; Srinivasan, L.; Allsop, J.M.; Hajnal, J.V.; Rutherford, M.A.; Edwards, A.D.; Counsell, S.J. Diffusion tensor imaging with tract-based spatial statistics reveals local white matter abnormalities in preterm infants. Neuroimage 2007, 35, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Drury, P.P.; Bennet, L.; Booth, L.C.; Davidson, J.O.; Wassink, G.; Gunn, A.J. Maturation of the mitochondrial redox response to profound asphyxia in fetal sheep. PLoS ONE [Electron. Resour.] 2012, 7, e39273. [Google Scholar] [CrossRef] [Green Version]
- Stark, M.J.; Hodyl, N.A.; Belegar, V.K.; Andersen, C.C. Intrauterine inflammation, cerebral oxygen consumption and susceptibility to early brain injury in very preterm newborns. Arch. Dis. Child. Fetal Neonatal Ed. 2016, 101, F137–F142. [Google Scholar] [CrossRef]
- Mathai, S.; Booth, L.C.; Davidson, J.O.; Drury, P.P.; Fraser, M.; Jensen, E.C.; George, S.; Naylor, A.; Gunn, A.J.; Bennet, L. Acute on chronic exposure to endotoxin in preterm fetal sheep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R189–R197. [Google Scholar] [CrossRef] [Green Version]
- Galinsky, R.; Hooper, S.B.; Polglase, G.R.; Moss, T.J. Intrauterine inflammation alters fetal cardiopulmonary and cerebral haemodynamics in sheep. J. Physiol. 2013, 591 Pt 20, 5061–5070. [Google Scholar] [CrossRef]
- Galinsky, R.; Hooper, S.B.; Wallace, M.J.; Westover, A.J.; Black, M.J.; Moss, T.J.; Polglase, G.R. Intrauterine inflammation alters cardiopulmonary and cerebral haemodynamics at birth in preterm lambs. J. Physiol. 2013, 591 Pt 8, 2127–2137. [Google Scholar] [CrossRef]
- Lee, H.S.; Ghetti, A.; Pinto-Duarte, A.; Wang, X.; Dziewczapolski, G.; Galimi, F.; Huitron-Resendiz, S.; Piña-Crespo, J.C.; Roberts, A.J.; Verma, I.M.; et al. Astrocytes contribute to gamma oscillations and recognition memory. Proc. Natl. Acad. Sci. USA 2014, 111, E3343–E3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, C.D.; Read, J.; Acharjee, S.; Pittman, Q.J. Early Life Inflammation Increases CA1 Pyramidal Neuron Excitability in a Sex and Age Dependent Manner through a Chloride Homeostasis Disruption. J. Neurosci. 2019, 39, 7244–7259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vikman, K.S.; Owe-Larsson, B.; Brask, J.; Kristensson, K.S.; Hill, R.H. Interferon-gamma-induced changes in synaptic activity and AMPA receptor clustering in hippocampal cultures. Brain Res. 2001, 896, 18–29. [Google Scholar] [CrossRef]
- Didion, S.P. Cellular and Oxidative Mechanisms Associated with Interleukin-6 Signaling in the Vasculature. Int. J. Mol. Sci. 2017, 18, 2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, B.S.; Geller, D.A. Molecular regulation of the human inducible nitric oxide synthase (iNOS) gene. Shock 2000, 13, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Titheradge, M.A. Nitric oxide in septic shock. Biochim. Biophys. Acta 1999, 1411, 437–455. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.Y.; Phillips, D.J.; Stockx, E.M.; Yu, V.Y.; Walker, A.M. Endotoxin has acute and chronic effects on the cerebral circulation of fetal sheep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R640–R650. [Google Scholar] [CrossRef] [PubMed]
- Percie du Sert, N.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; Emerson, M.; et al. Reporting animal research: Explanation and elaboration for the ARRIVE guidelines 2.0. PLoS Biol. 2020, 18, e3000411. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.E.; Gluckman, P.D. Real-time spectral intensity analysis of the EEG on a common microcomputer. J. Neurosci. Methods 1990, 32, 9–13. [Google Scholar] [CrossRef]
- van den Heuij, L.G.; Mathai, S.; Davidson, J.O.; Lear, C.A.; Booth, L.C.; Fraser, M.; Gunn, A.J.; Bennet, L. Synergistic white matter protection with acute-on-chronic endotoxin and subsequent asphyxia in preterm fetal sheep. J. Neuroinflammation 2014, 11, 89. [Google Scholar] [CrossRef] [Green Version]
- Lear, C.A.; Davidson, J.O.; Booth, L.C.; Wassink, G.; Galinsky, R.; Drury, P.P.; Fraser, M.; Bennet, L.; Gunn, A.J. Biphasic changes in fetal heart rate variability in preterm fetal sheep developing hypotension after acute on chronic lipopolysaccharide exposure. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R387–R395. [Google Scholar] [CrossRef] [Green Version]
- Jakovcevski, I.; Filipovic, R.; Mo, Z.; Rakic, S.; Zecevic, N. Oligodendrocyte development and the onset of myelination in the human fetal brain. Front. Neuroanat. 2009, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Drury, P.P.; Davidson, J.O.; Bennet, L.; Booth, L.C.; Tan, S.; Fraser, M.; van den Heuij, L.G.; Gunn, A.J. Partial neural protection with prophylactic low-dose melatonin after asphyxia in preterm fetal sheep. J. Cereb. Blood Flow Metab. 2014, 34, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Nott, F.; Jane Pillow, J.; Dahl, M.; Kelly, S.B.; Melville, J.; McDonald, C.; Nitsos, I.; Lim, R.; Wallace, E.M.; Jenkin, G.; et al. Brain inflammation and injury at 48 h is not altered by human amnion epithelial cells in ventilated preterm lambs. Pediatr. Res. 2020, 88, 27–37. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Barres, B.A. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Yushkevich, P.A.; Alexander, D.C.; Gee, J.C. Deformable registration of diffusion tensor MR images with explicit orientation optimization. Med. Image Anal. 2006, 10, 764–785. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Schneider, T.; Wheeler-Kingshott, C.A.; Alexander, D.C. NODDI: Practical in vivo neurite orientation dispersion and density imaging of the human brain. Neuroimage 2012, 61, 1000–1016. [Google Scholar] [CrossRef] [PubMed]
- Levin, J.R.; Serlin, R.C.; Seaman, M.A. A controlled, powerful multiple-comparison strategy for several situations. Psychol. Bull. 1994, 115, 153–159. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight (kg) | Brain Weight (g) | Sex (M, F) | |
|---|---|---|---|
| Saline | 2.01 ± 0.18 | 32.7 ± 1.4 | 2, 4 |
| LPS | 1.94 ± 0.13 | 31.1 ± 0.8 | 3, 4 |
| CaBF (mL/min) | CaVC (mL/min/mmHg) | EEG (Hz) | EEG (dB) | Nuchal EMG (µV) | |
|---|---|---|---|---|---|
| Baseline | |||||
| Control | 28 ± 3 | 0.7 ± 0.1 | 10 ± 0 | 17 ± 1 | 2 ± 0 |
| LPS | 28 ± 4 | 0.8 ± 0.1 | 10 ± 1 | 16 ± 1 | 2 ± 0 |
| Day 1 | |||||
| Control | 31 ± 2 | 0.8 ± 0.1 | 10 ± 0 | 17 ± 1 | 2 ± 0 |
| LPS | 31 ± 4 | 0.9 ± 0.1 | 10 ± 1 | 17 ± 1 | 2 ± 0 |
| Day 2 | |||||
| Control | 28 ± 3 | 0.7 ± 0.1 | 10 ± 0 | 17 ± 1 | 2 ± 0 |
| LPS | 32 ± 5 | 0.9 ± 0.1 | 11 ± 0 | 17 ± 1 | 2 ± 0 |
| Day 3 | |||||
| Control | 29 ± 3 | 0.7 ± 0.1 | 11 ± 0 | 18 ± 1 | 2 ± 0 |
| LPS | 36 ± 5 | 1.0 ± 0.1 | 11 ± 0 | 17 ± 1 | 2 ± 0 |
| Day 4 | |||||
| Control | 29 ± 3 | 0.7 ± 0.1 | 10 ± 0 | 18 ± 1 | 2 ± 0 |
| LPS | 41 ± 5 * | 1.2 ± 0.1 * | 11 ± 0 * | 17 ± 1 | 2 ± 0 |
| Day 5 | |||||
| Control | 34 ± 2 | 0.8 ± 0.1 | 11 ± 0 | 18 ± 1 | 2 ± 0 |
| LPS | 44 ± 6 | 1.2 ± 0.2 * | 12 ± 0 * | 17 ± 1 | 2 ± 0 |
| Day 6 | |||||
| Control | 37 ± 3 | 0.9 ± 0.1 | 11 ± 0 | 19 ± 1 | 2 ± 0 |
| LPS | 44 ± 6 | 1.2 ± 0.2 | 12 ± 0 * | 17 ± 1 | 2 ± 0 |
| Day 7 | |||||
| Control | 38 ± 3 | 0.9 ± 0.1 | 11 ± 1 | 19 ± 1 | 2 ± 0 |
| LPS | 43 ± 7 | 1.1 ± 0.2 | 12 ± 0 * | 17 ± 1 | 2 ± 0 |
| Day 8 | |||||
| Control | 38 ± 3 | 1.0 ± 0.1 | 11 ± 1 | 19 ± 1 | 2 ± 0 |
| LPS | 45 ± 8 | 1.2 ± 0.2 | 12 ± 0 | 18 ± 1 | 2 ± 0 |
| Day 9 | |||||
| Control | 39 ± 3 | 1.0 ± 0.1 | 11 ± 1 | 19 ± 1 | 2 ± 0 |
| LPS | 46 ± 7 | 1.2 ± 0.2 | 12 ± 0 | 18 ± 1 | 2 ± 0 |
| Day 10 | |||||
| Control | 41 ± 3 | 1.0 ± 0.1 | 11 ± 1 | 19 ± 1 | 2 ± 0 |
| LPS | 48 ± 7 | 1.2 ± 0.2 | 12 ± 0 | 18 ± 1 | 2 ± 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galinsky, R.; van de Looij, Y.; Mitchell, N.; Dean, J.M.; Dhillon, S.K.; Yamaguchi, K.; Lear, C.A.; Wassink, G.; Davidson, J.O.; Nott, F.; et al. Magnetic Resonance Imaging Correlates of White Matter Gliosis and Injury in Preterm Fetal Sheep Exposed to Progressive Systemic Inflammation. Int. J. Mol. Sci. 2020, 21, 8891. https://doi.org/10.3390/ijms21238891
Galinsky R, van de Looij Y, Mitchell N, Dean JM, Dhillon SK, Yamaguchi K, Lear CA, Wassink G, Davidson JO, Nott F, et al. Magnetic Resonance Imaging Correlates of White Matter Gliosis and Injury in Preterm Fetal Sheep Exposed to Progressive Systemic Inflammation. International Journal of Molecular Sciences. 2020; 21(23):8891. https://doi.org/10.3390/ijms21238891
Chicago/Turabian StyleGalinsky, Robert, Yohan van de Looij, Natasha Mitchell, Justin M. Dean, Simerdeep K. Dhillon, Kyohei Yamaguchi, Christopher A. Lear, Guido Wassink, Joanne O. Davidson, Fraser Nott, and et al. 2020. "Magnetic Resonance Imaging Correlates of White Matter Gliosis and Injury in Preterm Fetal Sheep Exposed to Progressive Systemic Inflammation" International Journal of Molecular Sciences 21, no. 23: 8891. https://doi.org/10.3390/ijms21238891