Involvement of MicroRNA-1-FAM83A Axis Dysfunction in the Growth and Motility of Lung Cancer Cells

 ,
,
Abstract
:1. Introduction
2. Results
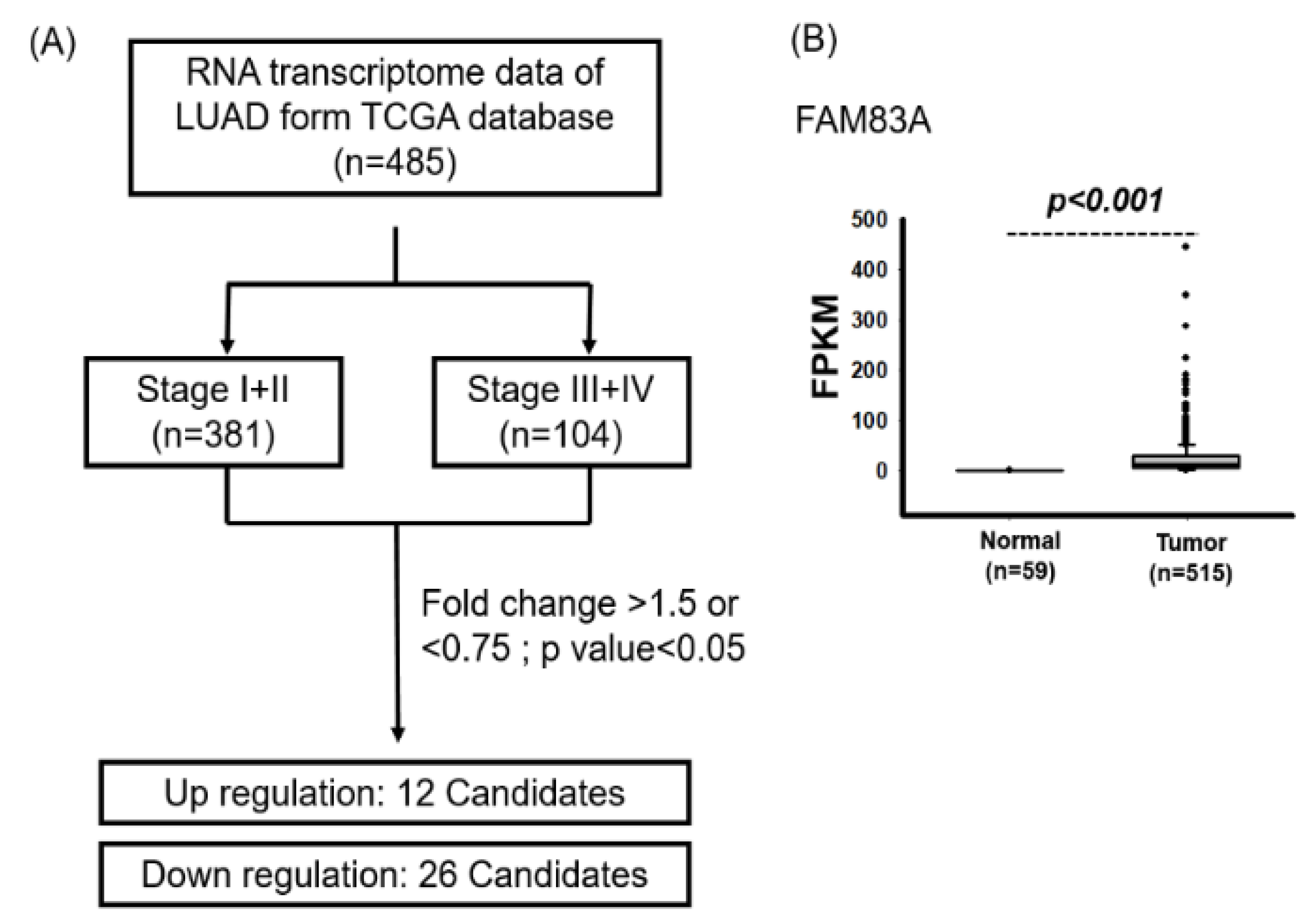
2.1. Identification of Prognostic Biomarkers for Lung Cancer
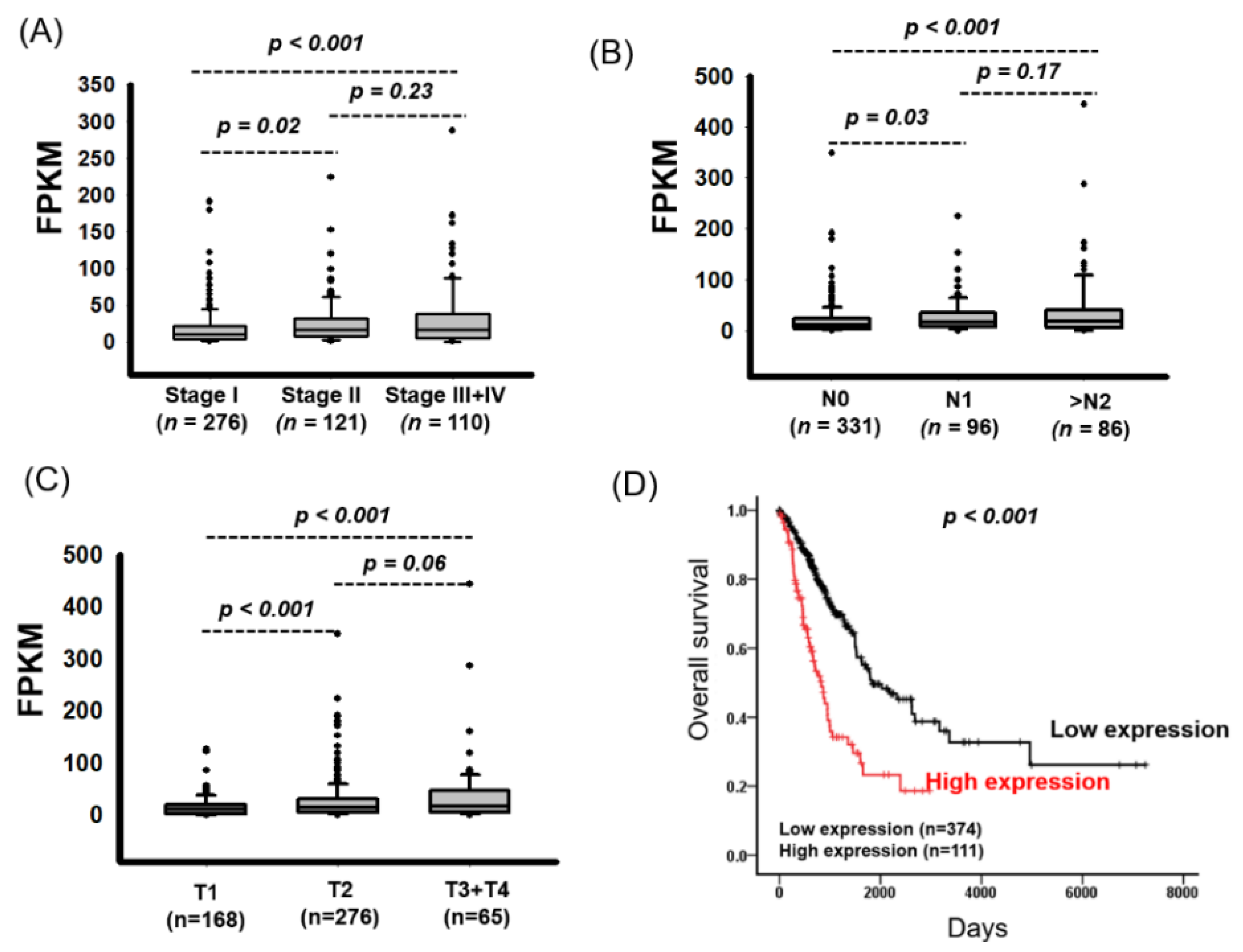
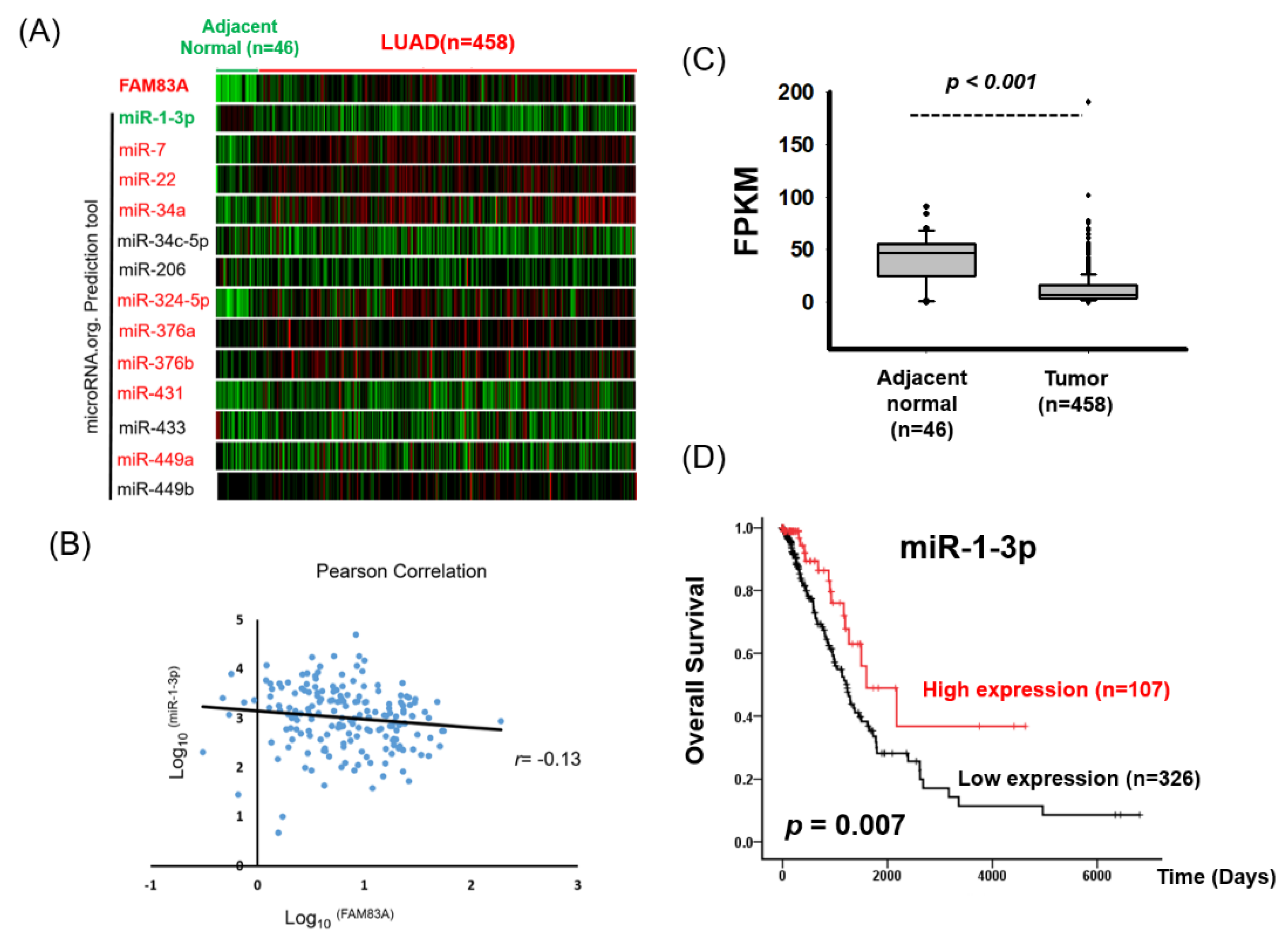
2.2. High FAM83A Expression Correlated with Worse Survival in Patients with LUAD
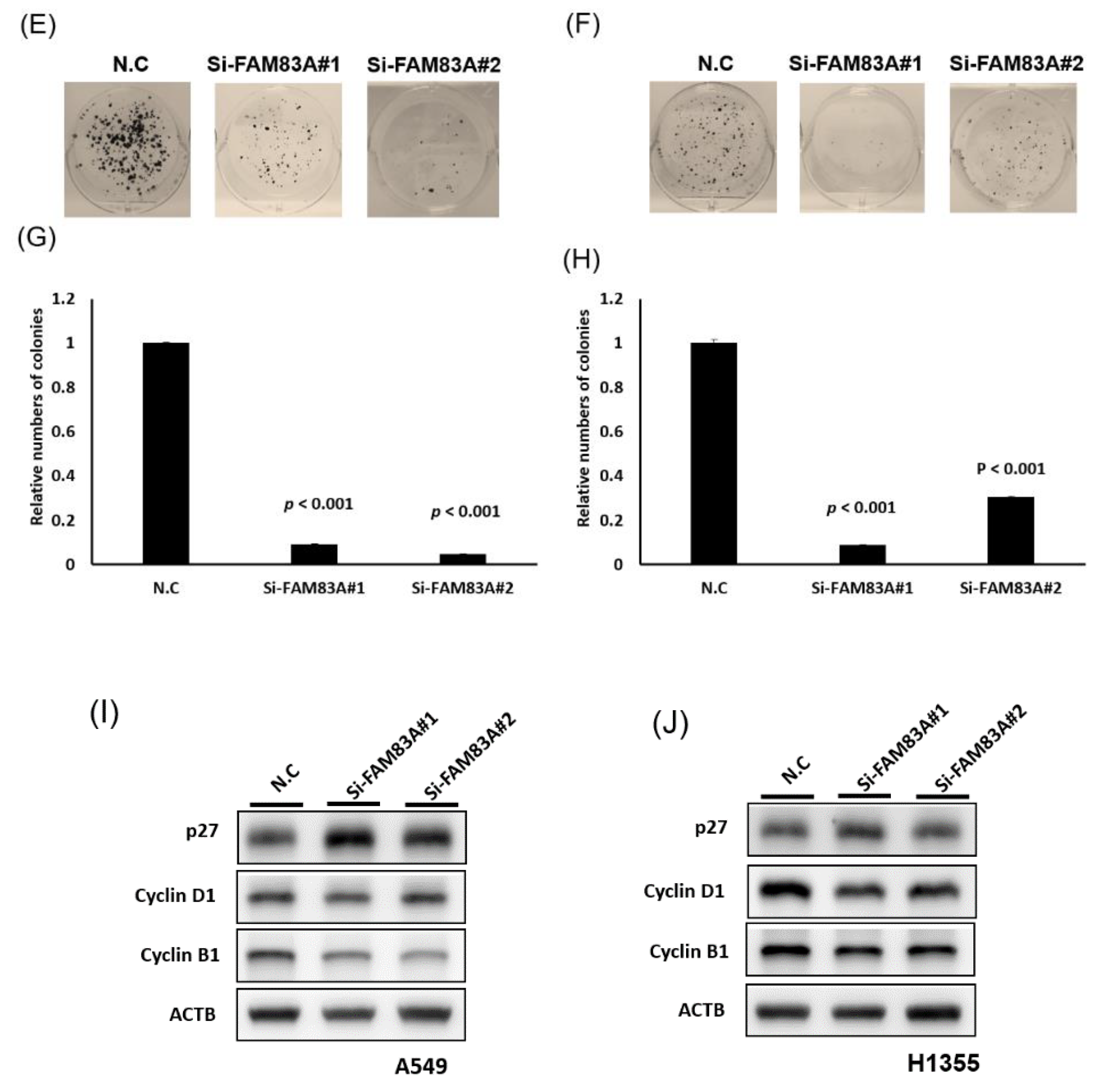
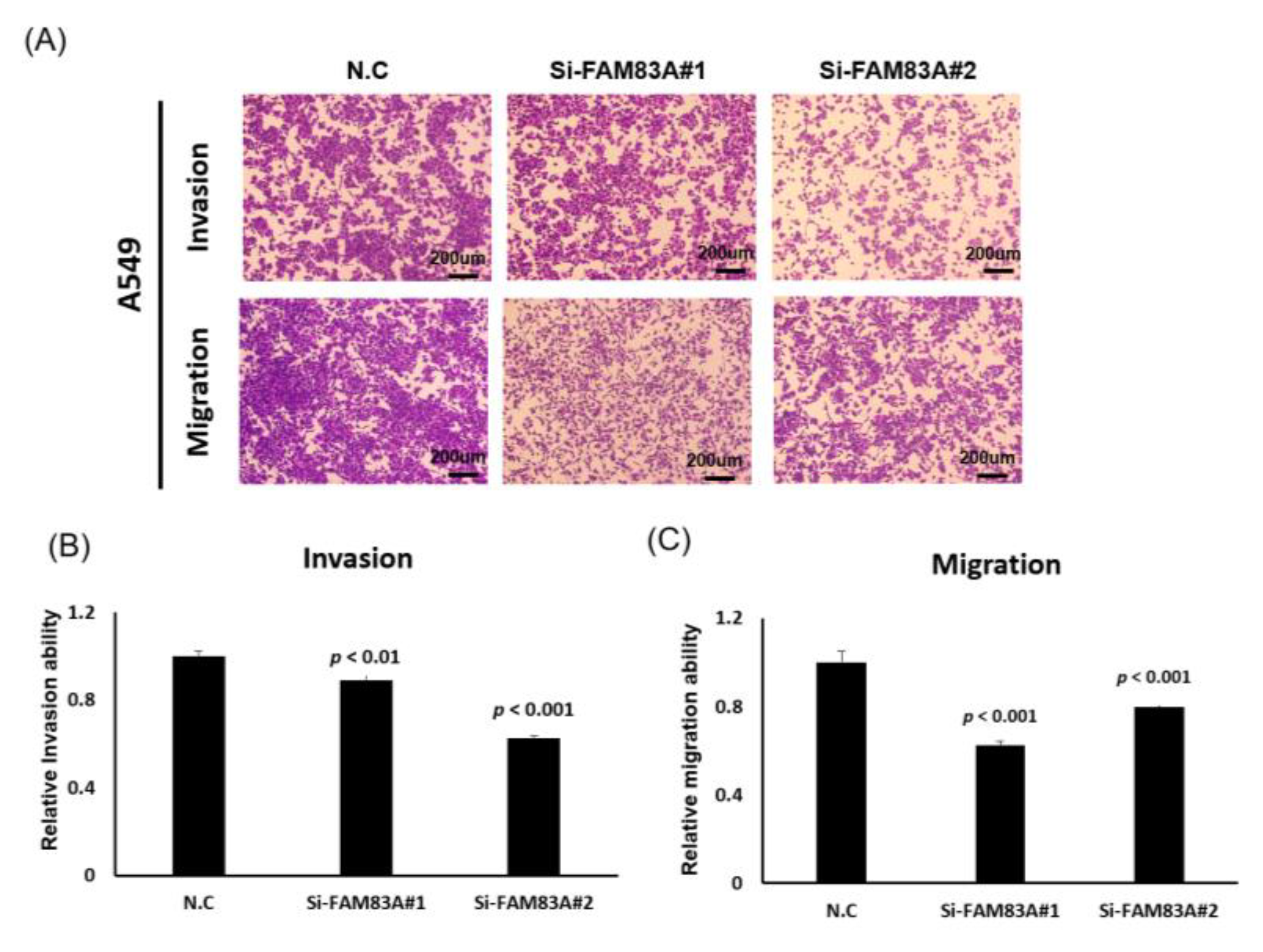
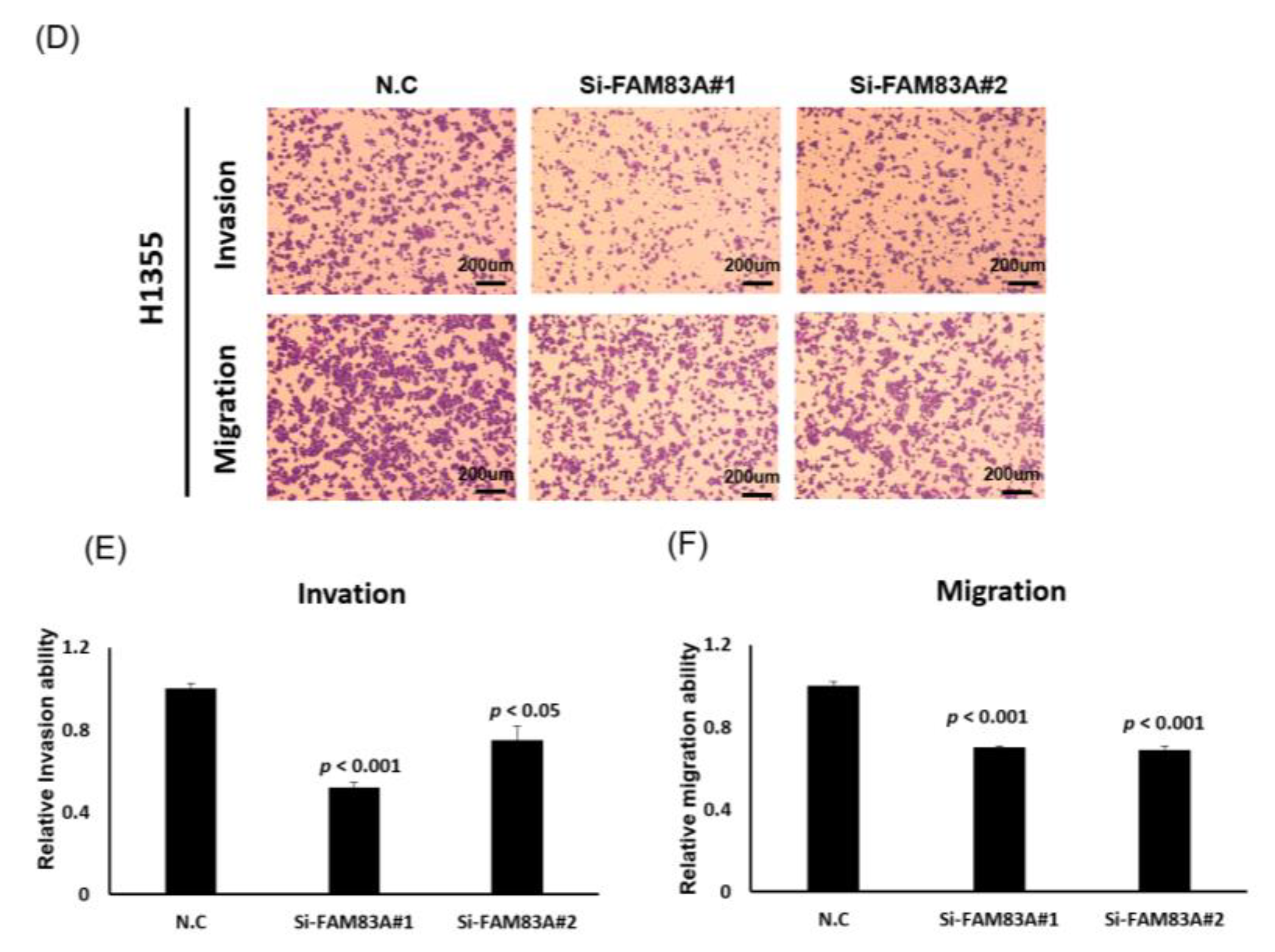
2.3. FAM83A Regulates Lung Cancer Cell Growth and Motility
2.4. Identification of FAM83A-Associated Signaling Pathways
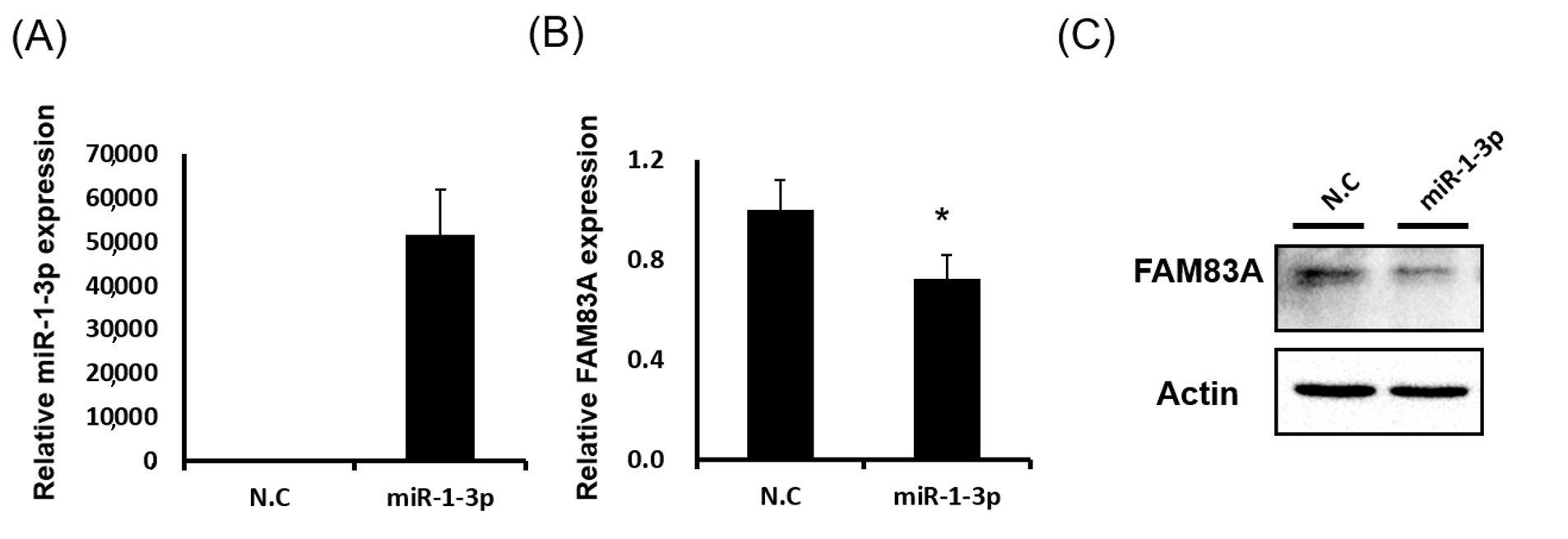
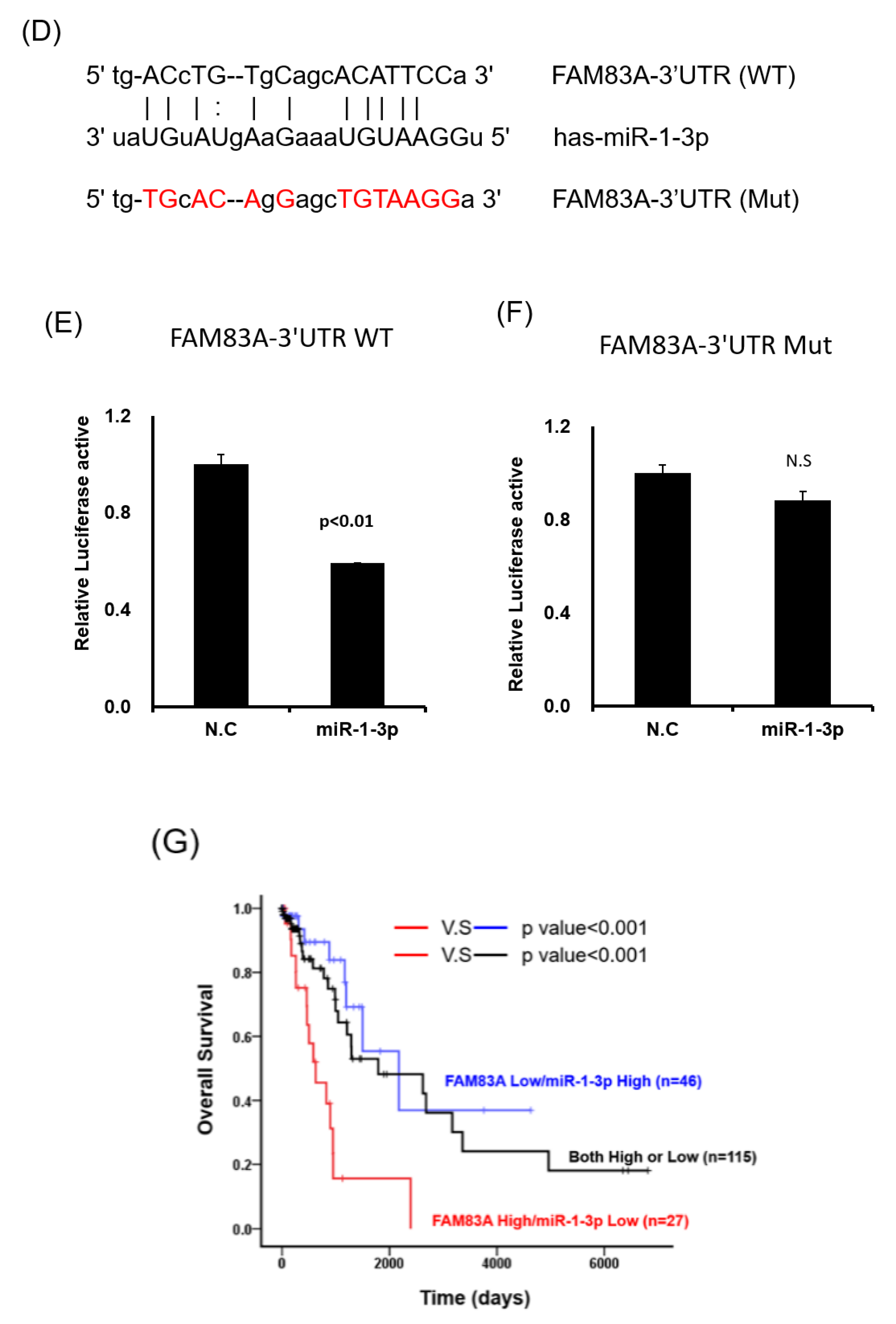
2.5. miR-1-3p Suppresses FAM83A Expression by Targeting the 3 Prime Untranslated Region of FAM83A
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Gene Expression Profiles According to Cancer Genome Atlas Data
4.3. Real-Time Reverse Transcription Polymerase Chain Reaction
- GAPDH-F: TGCACCACCAACTGCTTAGC
- GAPDH-R: GGCATGGACTGTGGTCATGAG
- FAM83A-F: CCCTATAAAGAGTGGCAACAG
- FAM83A-R: AACAGTGAGCAAACACACCG
4.4. RNA Interference Knockdown of FAM83A
4.5. Colony Formation Assay
4.6. Cell Proliferation, Migration, and Invasion Assay
4.7. Cell Migration and Invasion Ability
4.8. Microarray Data Analysis
4.9. Western Blotting
4.10. Stem–Loop Reverse Transcription PCR
- miR-1-3p-GSF: 5′-CGGCGGTGGAATGTAAAGAAGT-3′
- Universal reverse: 5′-CTGGTGTCGTGGAGTCGGCAATTC-3′
- U6-F: 5′-CTCGCTTCGGCAGCACA-3′
- U6-R: 5′-AACGCTTCACGAATTTGCGT-3
4.11. miR-1-3p Mimics Trasfection
4.12. miRNA Target Candidates and Luciferase Reporter Assay
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.D.; Alexandrov, A.; Kim, J.; Wala, J.; Berger, A.H.; Pedamallu, C.S.; Shukla, S.A.; Guo, G.; Brooks, A.N.; Murray, B.A.; et al. Distinct patterns of somatic genome alterations in lung adenocarcinomas and squamous cell carcinomas. Nat. Genet. 2016, 48, 607–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Socinski, M.A.; Crowell, R.; Hensing, T.E.; Langer, C.J.; Lilenbaum, R.; Sandler, A.B.; Morris, D.; American College of Chest, P. Treatment of non-small cell lung cancer, stage IV: ACCP evidence-based clinical practice guidelines (2nd edition). Chest 2007, 132, 277S–289S. [Google Scholar] [CrossRef] [PubMed]
- Cipriano, R.; Miskimen, K.L.; Bryson, B.L.; Foy, C.R.; Bartel, C.A.; Jackson, M.W. Conserved oncogenic behavior of the FAM83 family regulates MAPK signaling in human cancer. Mol. Cancer Res. 2014, 12, 1156–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Huang, J.; Liu, Z.; Liang, Q.; Zhang, N.; Jin, Y. FAM83A is amplified and promotes cancer stem cell-like traits and chemoresistance in pancreatic cancer. Oncogenesis 2017, 6, e300. [Google Scholar] [CrossRef]
- Parameswaran, N.; Bartel, C.A.; Hernandez-Sanchez, W.; Miskimen, K.L.; Smigiel, J.M.; Khalil, A.M.; Jackson, M.W. A FAM83A Positive Feed-back Loop Drives Survival and Tumorigenicity of Pancreatic Ductal Adenocarcinomas. Sci. Rep. 2019, 9, 13396. [Google Scholar] [CrossRef] [Green Version]
- Richtmann, S.; Wilkens, D.; Warth, A.; Lasitschka, F.; Winter, H.; Christopoulos, P.; Herth, F.J.F.; Muley, T.; Meister, M.; Schneider, M.A. FAM83A and FAM83B as Prognostic Biomarkers and Potential New Therapeutic Targets in NSCLC. Cancers 2019, 11, 652. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.; Jiao, Z.; Yu, A.; Wang, T. Long noncoding antisense RNA FAM83A-AS1 promotes lung cancer cell progression by increasing FAM83A. J. Cell. Biochem. 2019, 120, 10505–10512. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Meier, R.; Furuta, S.; Lenburg, M.E.; Kenny, P.A.; Xu, R.; Bissell, M.J. FAM83A confers EGFR-TKI resistance in breast cancer cells and in mice. J. Clin. Investig. 2012, 122, 3211–3220. [Google Scholar] [CrossRef] [Green Version]
- Bartel, C.A.; Jackson, M.W. HER2-positive breast cancer cells expressing elevated FAM83A are sensitive to FAM83A loss. PLoS ONE 2017, 12, e0176778. [Google Scholar] [CrossRef]
- Liu, C.; Peng, X.; Li, Y.; Liu, S.; Hou, R.; Zhang, Y.; Zuo, S.; Liu, Z.; Luo, R.; Li, L.; et al. Positive feedback loop of FAM83A/PI3K/AKT/c-Jun induces migration, invasion and metastasis in hepatocellular carcinoma. Biomed. Pharmacother. 2020, 123, 109780. [Google Scholar] [CrossRef] [PubMed]
- Grant, S. FAM83A and FAM83B: Candidate oncogenes and TKI resistance mediators. J. Clin. Investig. 2012, 122, 3048–3051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cipriano, R.; Graham, J.; Miskimen, K.L.; Bryson, B.L.; Bruntz, R.C.; Scott, S.A.; Brown, H.A.; Stark, G.R.; Jackson, M.W. FAM83B mediates EGFR- and RAS-driven oncogenic transformation. J. Clin. Investig. 2012, 122, 3197–3210. [Google Scholar] [CrossRef] [PubMed]
- McHugh, B.J.; Murdoch, A.; Haslett, C.; Sethi, T. Loss of the integrin-activating transmembrane protein Fam38A (Piezo1) promotes a switch to a reduced integrin-dependent mode of cell migration. PLoS ONE 2012, 7, e40346. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.N.; Lu, Y.P.; Liu, J.J.; Huang, J.K.; Liu, Y.P.; Xiao, C.X.; Jazag, A.; Ren, J.L.; Guleng, B. Piezo1 is as a novel trefoil factor family 1 binding protein that promotes gastric cancer cell mobility in vitro. Dig. Dis. Sci. 2014, 59, 1428–1435. [Google Scholar] [CrossRef]
- Etem, E.O.; Ceylan, G.G.; Ozaydin, S.; Ceylan, C.; Ozercan, I.; Kuloglu, T. The increased expression of Piezo1 and Piezo2 ion channels in human and mouse bladder carcinoma. Adv. Clin. Exp. Med. Off. Organ Wroc. Med Univ. 2018, 27, 1025–1031. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, Y.; Huang, T.; Wu, F.; Liu, L.; Kwan, J.S.H.; Cheng, A.S.L.; Yu, J.; To, K.F.; Kang, W. PIEZO1 functions as a potential oncogene by promoting cell proliferation and migration in gastric carcinogenesis. Mol. Carcinog. 2018, 57, 1144–1155. [Google Scholar] [CrossRef]
- Zhang, J.T.; Lin, Y.C.; Xiao, B.F.; Yu, B.T. Overexpression of Family with Sequence Similarity 83, Member A (FAM83A) Predicts Poor Clinical Outcomes in Lung Adenocarcinoma. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 4264–4272. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, X.; Ji, X.; Liu, B.; Amos, C.I. RNA-seq analysis of lung adenocarcinomas reveals different gene expression profiles between smoking and nonsmoking patients. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2015, 36, 8993–9003. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, N.; Leidinger, P.; Becker, K.; Backes, C.; Fehlmann, T.; Pallasch, C.; Rheinheimer, S.; Meder, B.; Stahler, C.; Meese, E.; et al. Distribution of miRNA expression across human tissues. Nucleic Acids Res. 2016, 44, 3865–3877. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, X.; Liu, Y.; Han, G.; Sun, D. Long noncoding RNA RMRP promotes proliferation and invasion via targeting miR-1-3p in non-small-cell lung cancer. J. Cell. Biochem. 2019, 120, 15170–15181. [Google Scholar] [CrossRef] [PubMed]
- Jiao, D.; Chen, J.; Li, Y.; Tang, X.; Wang, J.; Xu, W.; Song, J.; Li, Y.; Tao, H.; Chen, Q. miR-1-3p and miR-206 sensitizes HGF-induced gefitinib-resistant human lung cancer cells through inhibition of c-Met signalling and EMT. J. Cell Mol. Med. 2018, 22, 3526–3536. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Shen, Y.; Feng, G. Predicting the survival of patients with lung adenocarcinoma using a four-gene prognosis risk model. Oncol. Lett. 2019, 18, 535–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Geng, J.; Xu, S.; Meng, Q.; Chen, K.; Liu, F.; Yang, F.; Pan, B.; Yu, Y. FAM83A signaling induces epithelial-mesenchymal transition by the PI3K/AKT/Snail pathway in NSCLC. Aging 2019, 11, 6069–6088. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, R.; Zhang, D.; Lu, T.; Yu, W.; Wo, Y.; Liu, A.; Sui, T.; Cui, J.; Qin, Y.; et al. Circ-ZKSCAN1 regulates FAM83A expression and inactivates MAPK signaling by targeting miR-330-5p to promote non-small cell lung cancer progression. Transl. Lung Cancer Res. 2019, 8, 862–875. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liao, G.Q.; He, P.; Zhu, H.; Liu, P.H.; Qu, Y.M.; Song, X.M.; Xu, Q.W.; Gao, Q.; Zhang, Y.; et al. Detection of circulating cancer cells in lung cancer patients with a panel of marker genes. Biochem. Biophys. Res. Commun. 2008, 372, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.W.; Li, Z.H.; Lei, L.; Liu, C.C.; Wang, Z.; Fei, L.R.; Yang, M.Q.; Huang, W.J.; Xu, H.T. FAM83A Promotes Lung Cancer Progression by Regulating the Wnt and Hippo Signaling Pathways and Indicates Poor Prognosis. Front. Oncol. 2020, 10, 180. [Google Scholar] [CrossRef] [Green Version]
- Ramirez de Molina, A.; Gallego-Ortega, D.; Sarmentero, J.; Banez-Coronel, M.; Martin-Cantalejo, Y.; Lacal, J.C. Choline kinase is a novel oncogene that potentiates RhoA-induced carcinogenesis. Cancer Res. 2005, 65, 5647–5653. [Google Scholar] [CrossRef] [Green Version]
- Ramirez de Molina, A.; Rodriguez-Gonzalez, A.; Gutierrez, R.; Martinez-Pineiro, L.; Sanchez, J.; Bonilla, F.; Rosell, R.; Lacal, J. Overexpression of choline kinase is a frequent feature in human tumor-derived cell lines and in lung, prostate, and colorectal human cancers. Biochem. Biophys. Res. Commun. 2002, 296, 580–583. [Google Scholar] [CrossRef]
- Koshizuka, K.; Hanazawa, T.; Fukumoto, I.; Kikkawa, N.; Matsushita, R.; Mataki, H.; Mizuno, K.; Okamoto, Y.; Seki, N. Dual-receptor (EGFR and c-MET) inhibition by tumor-suppressive miR-1 and miR-206 in head and neck squamous cell carcinoma. J. Hum. Genet. 2017, 62, 113–121. [Google Scholar] [CrossRef]
- Lin, M.H.; Chen, Y.Z.; Lee, M.Y.; Weng, K.P.; Chang, H.T.; Yu, S.Y.; Dong, B.J.; Kuo, F.R.; Hung, L.T.; Liu, L.F.; et al. Comprehensive identification of microRNA arm selection preference in lung cancer: miR-324-5p and -3p serve oncogenic functions in lung cancer. Oncol. Lett. 2018, 15, 9818–9826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Tsai, K.W.; Lo, Y.H.; Liu, H.; Yeh, C.Y.; Chen, Y.Z.; Hsu, C.W.; Chen, W.S.; Wang, J.H. Linc00659, a long noncoding RNA, acts as novel oncogene in regulating cancer cell growth in colorectal cancer. Mol. Cancer 2018, 17, 72. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Wu, C.C.; Tu, Y.T.; Chen, Y.R.; Lee, M.C.; Tsai, K.W. Involvement of the MicroRNA-1-LITAF Axis in Gastric Cancer Cell Growth and Invasion. Anticancer Res. 2020, 40, 6247–6256. [Google Scholar] [CrossRef] [PubMed]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA.org resource: Targets and expression. Nucleic Acids Res. 2008, 36, D149–D153. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | No. (%) | OS | |
|---|---|---|---|
| CHR (95% CI) | p-Value | ||
| Pathology stage | |||
| I + II | 381 (78.6) | 1.00 | |
| III + IV | 104 (21.4) | 2.76 (2.00–3.80) | < 0.001 |
| pT stage (n = 482) | |||
| T1 + T2 | 423 (87.2) | 1.00 | |
| T3 + T4 | 62 (12.8) | 2.43 (1.65–3.60) | < 0.001 |
| pN stage (n = 481) | |||
| N0 | 323 (66.6) | 1.00 | |
| N1 | 89 (18.4) | 2.49 (1.73–3.58) | < 0.001 |
| >N2 | 73 (15.0) | 3.31 (2.24–4.88) | < 0.001 |
| pM stage | |||
| M0 | 460 (94.8) | 1.00 | |
| M1 | 25 (5.2) | 2.23 (1.31–3.80) | 0.003 |
| Variables | FAM83A (n = 511) | |||
|---|---|---|---|---|
| No. (%) | Mean ± SD | Median | p-Value | |
| Pathology stage | ||||
| I | 275 (53.8) | 18.05 ± 25.58 cd | 10.32 | < 0.001 a |
| II | 126 (24.7) | 24.28 ± 30.47 c | 16.16 | |
| III+ IV | 110 (21.5) | 41.67 ± 66.33 d | 20.54 | |
| pT stage (n = 508) | ||||
| T1 | 169 (33.1) | 15.29 ± 18.70 ef | 10.81 | < 0.001 a |
| T2 | 276 (54.0) | 26.81 ± 39.51 e | 14.64 | |
| T3 + T4 | 66 (12.9) | 39.07 ± 67.64 f | 17.20 | |
| pN stage (n = 507) | ||||
| N0 | 339 (66.3) | 19.33 ± 30.56 gh | 11.00 | < 0.001 a |
| N1 | 95 (18.6) | 27.61 ± 34.32 g | 16.85 | |
| >N2 | 77 (15.1) | 43.91 ± 67.96 h | 21.45 | |
| pM stage | ||||
| M0 | 485 (94.9) | 23.09 ± 32.62 | 12.72 | 0.531 b |
| M1 | 26 (5.1) | 54.10 ± 105.81 | 18.65 | |
| Characteristic | No. (%) | OS | |||
|---|---|---|---|---|---|
| CHR (95% CI) | p-Value | AHR (95% CI) | p-Value | ||
| FAM83A | (n = 485) | ||||
| Low | 374 (77.1) | 1.00 | 1.00 | ||
| High | 111 (22.9) | 2.57 (1.87–3.54) | <0.001 | 2.17 (1.57–3.01) | <0.001 |
| Variables | miR-1-3p (n = 453) | |||
|---|---|---|---|---|
| No. (%) | Mean ± SD | Median | p-Value | |
| Pathology stage | ||||
| I | 248 (54.7) | 13.24 ± 18.01 | 8.07 | 0.325 a |
| II | 115 (25.4) | 10.18 ± 12.27 | 5.19 | |
| III | 73 (16.1) | 10.88 ± 10.13 | 6.49 | |
| IV | 17 (3.8) | 12.71 ± 20.41 | 6.13 | |
| pT stage | ||||
| T1 | 157 (34.7) | 15.18 ± 16.56 e | 10.01 | 0.002 b |
| T2 | 238 (52.5) | 10.02 ± 15.63 e | 5.95 | |
| T3 | 43 (9.5) | 12.68 ± 13.92 | 6.55 | |
| T4 | 15 (3.3) | 10.20 ± 7.87 | 6.61 | |
| pN stage | ||||
| N0 | 306 (67.5) | 12.53 ± 17.29 | 6.74 | 0.772 a |
| N1 | 81 (17.9) | 11.46 ± 13.12 | 7.67 | |
| N2 | 65 (14.3) | 10.80 ± 10.48 | 6.49 | |
| N3 | 1 (0.3) | 3.03 ± | 3.03 | |
| pM stage | ||||
| M0 | 436 (96.2) | 12.04 ± 15.58 | 6.74 | 0.863 c |
| M1 | 17 (3.8) | 12.71 ± 20.41 | 6.13 | |
| Characteristic | No. (%) | CHR (95% CI) | p-Value | AHR (95% CI) | p-Value |
|---|---|---|---|---|---|
| miR-1-3p | (n = 433) | ||||
| Low | 326 (75.3) | 1.00 | 1.00 | ||
| High | 107 (24.7) | 0.48 (0.28–0.83) | 0.009 | 0.54 (0.31–0.94) | 0.029 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, P.-J.; Chen, Y.-H.; Tsai, K.-W.; Yeah, H.-Y.; Yeh, C.-Y.; Tu, Y.-T.; Yang, C.-Y. Involvement of MicroRNA-1-FAM83A Axis Dysfunction in the Growth and Motility of Lung Cancer Cells. Int. J. Mol. Sci. 2020, 21, 8833. https://doi.org/10.3390/ijms21228833
Liu P-J, Chen Y-H, Tsai K-W, Yeah H-Y, Yeh C-Y, Tu Y-T, Yang C-Y. Involvement of MicroRNA-1-FAM83A Axis Dysfunction in the Growth and Motility of Lung Cancer Cells. International Journal of Molecular Sciences. 2020; 21(22):8833. https://doi.org/10.3390/ijms21228833
Chicago/Turabian StyleLiu, Pei-Jung, Yu-Hsuan Chen, Kuo-Wang Tsai, Hui-Ying Yeah, Chung-Yu Yeh, Ya-Ting Tu, and Chih-Yun Yang. 2020. "Involvement of MicroRNA-1-FAM83A Axis Dysfunction in the Growth and Motility of Lung Cancer Cells" International Journal of Molecular Sciences 21, no. 22: 8833. https://doi.org/10.3390/ijms21228833