Identification of Anthocyanins-Related Glutathione S-Transferase (GST) Genes in the Genome of Cultivated Strawberry (Fragaria × ananassa)

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
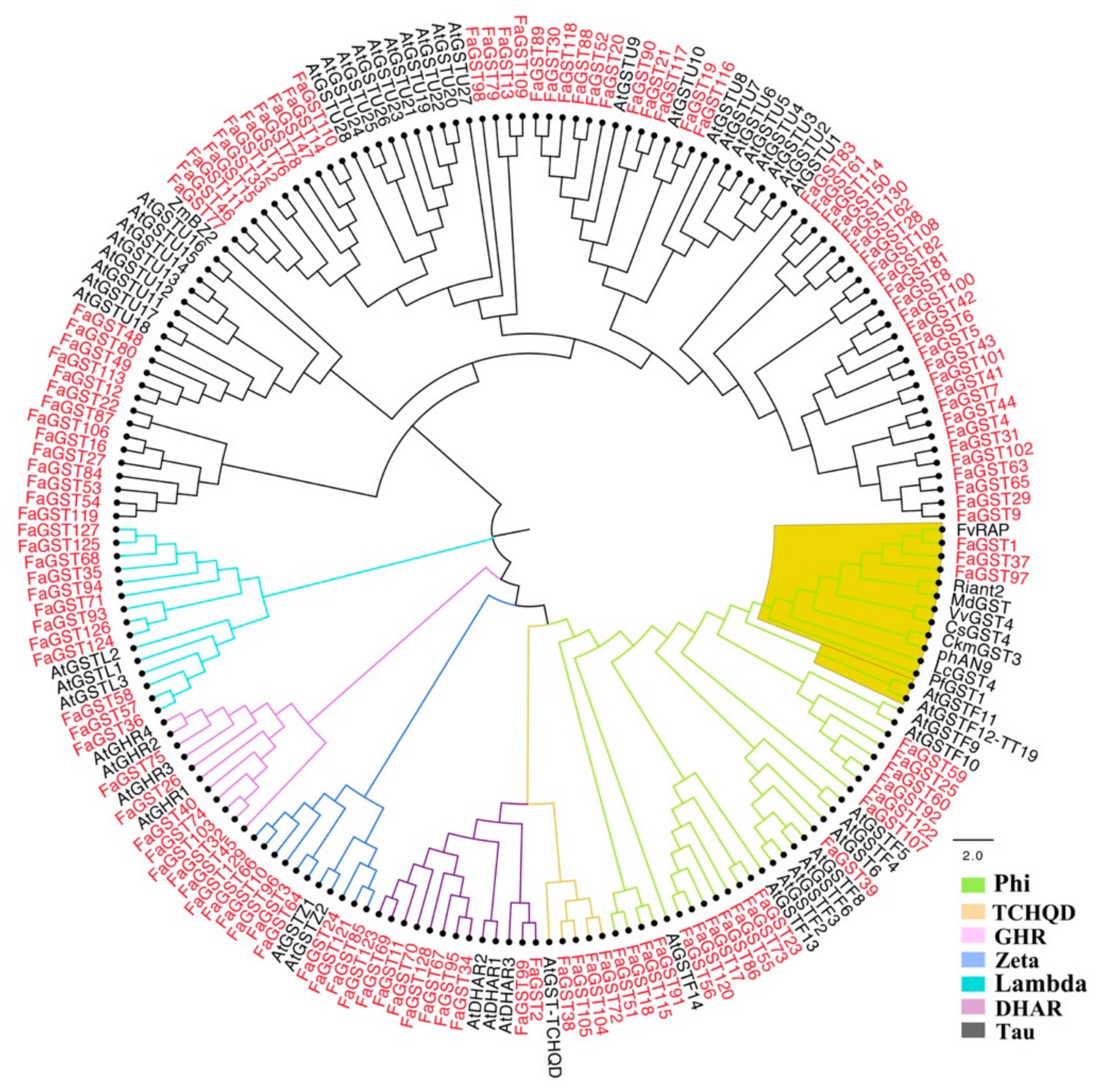
2.1. Identification, Characteristics, and Evolutionary Analysis of the Entire 130 FaGST Genes in Cultivated Strawberry
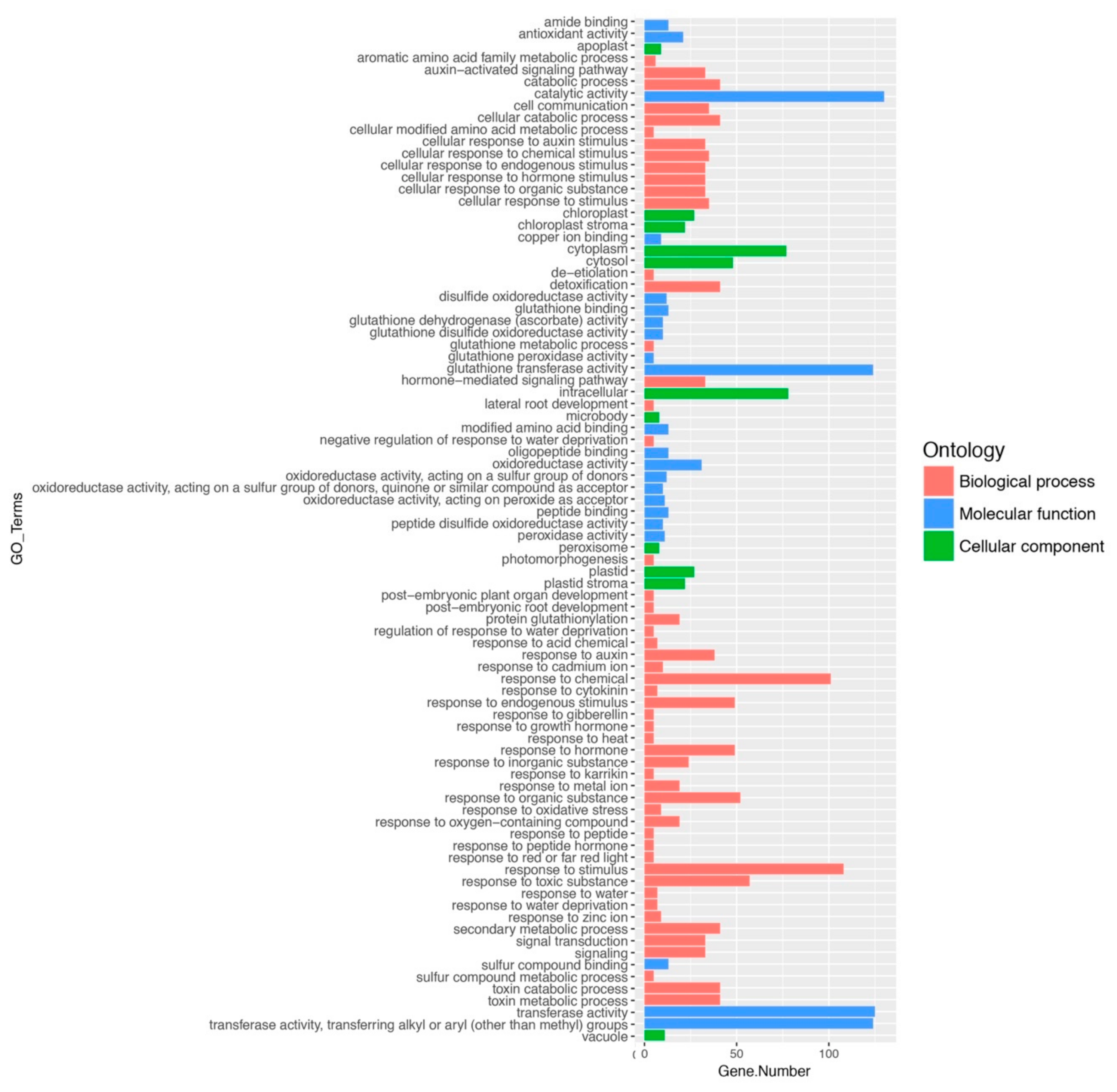
2.2. Gene Ontology and Classification of FaGST Genes
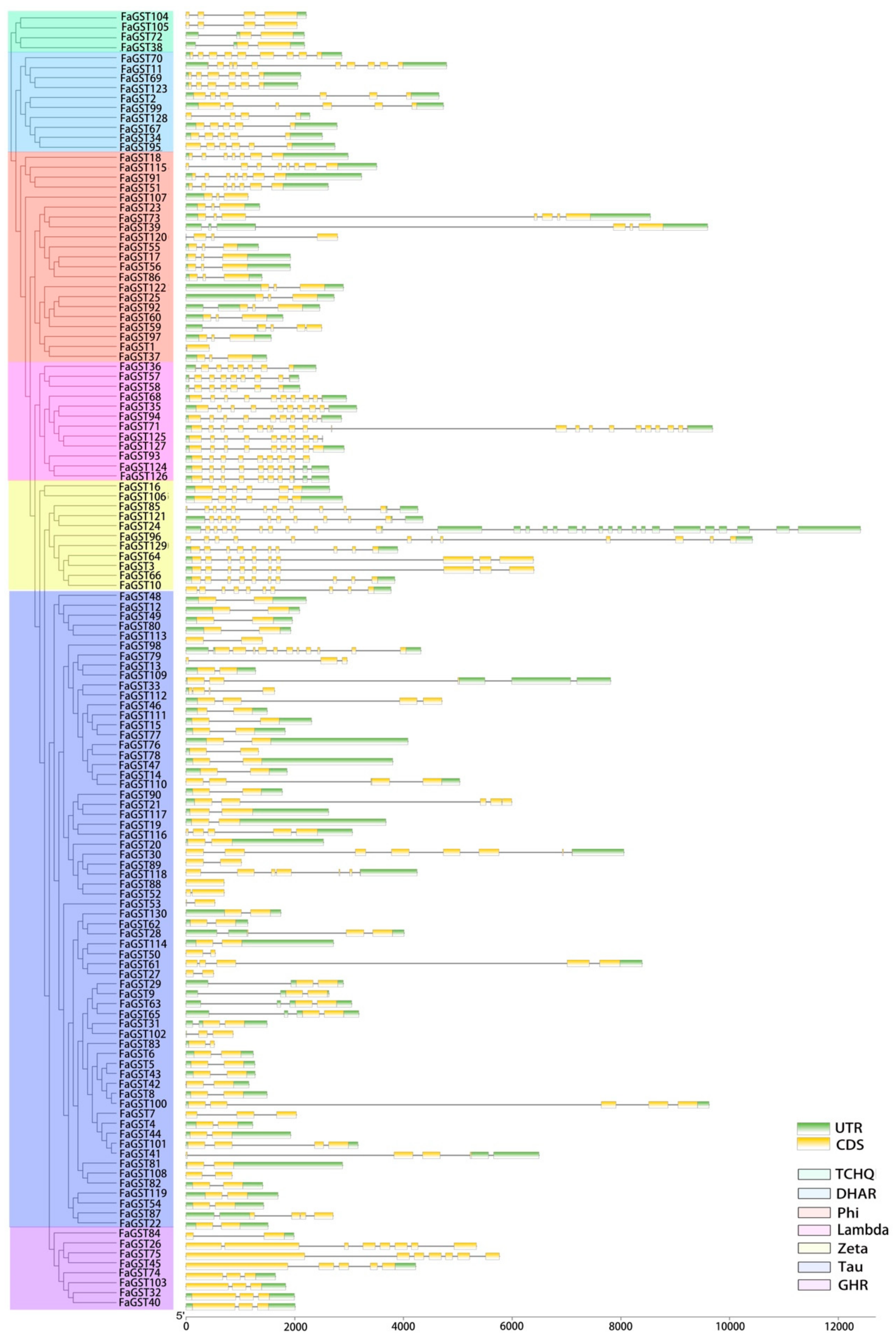
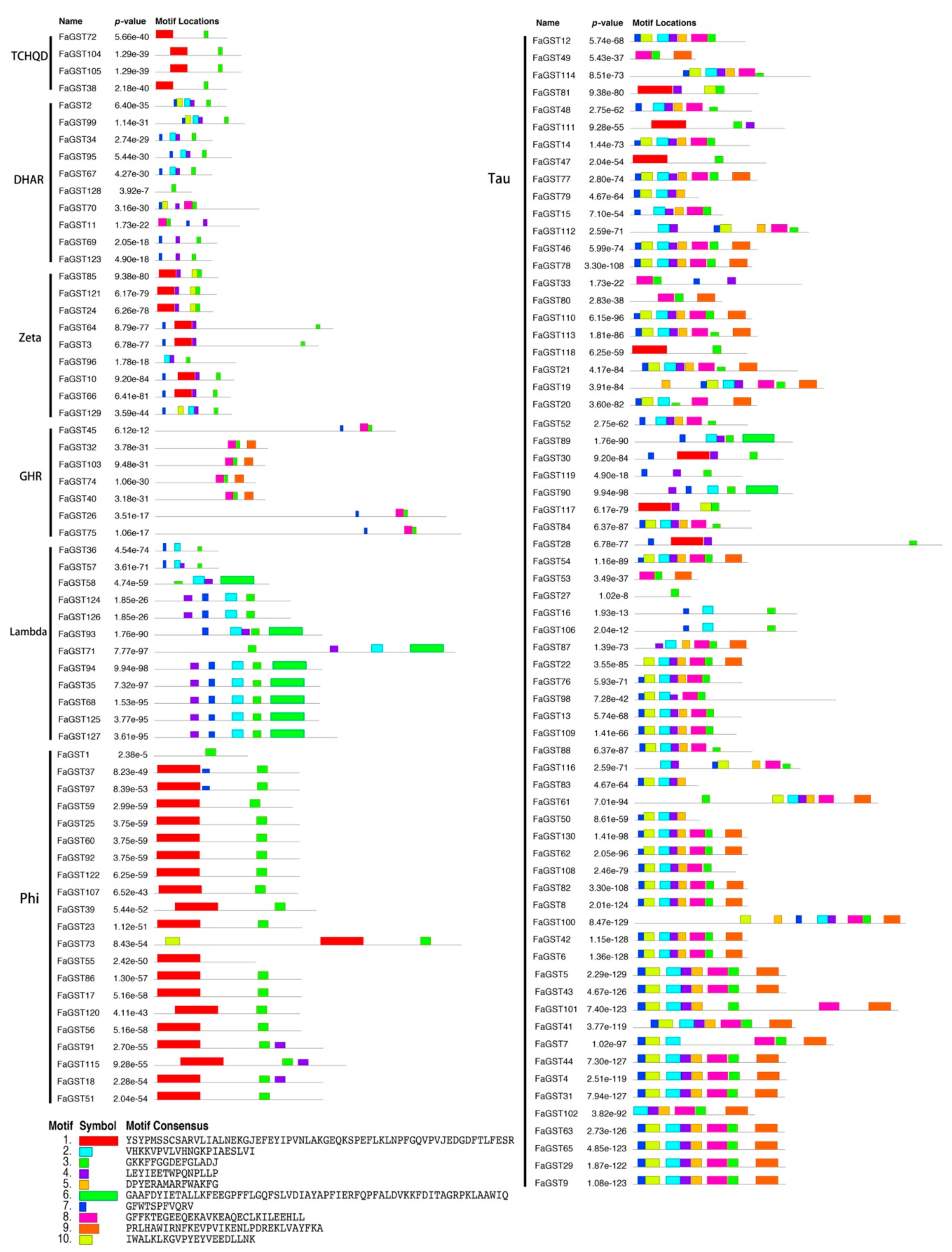
2.3. Gene Structure and Conserved Motifs Analysis
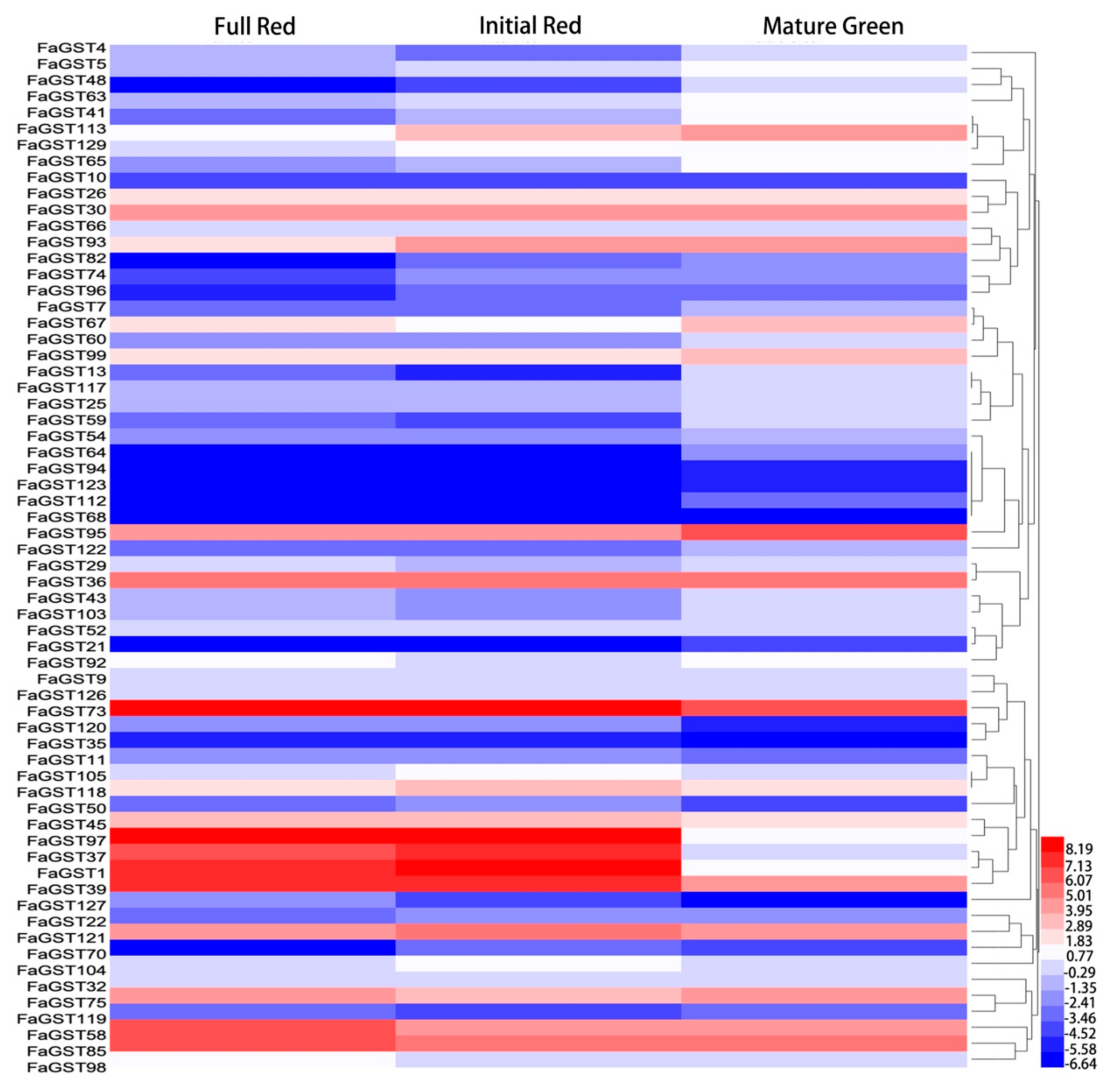
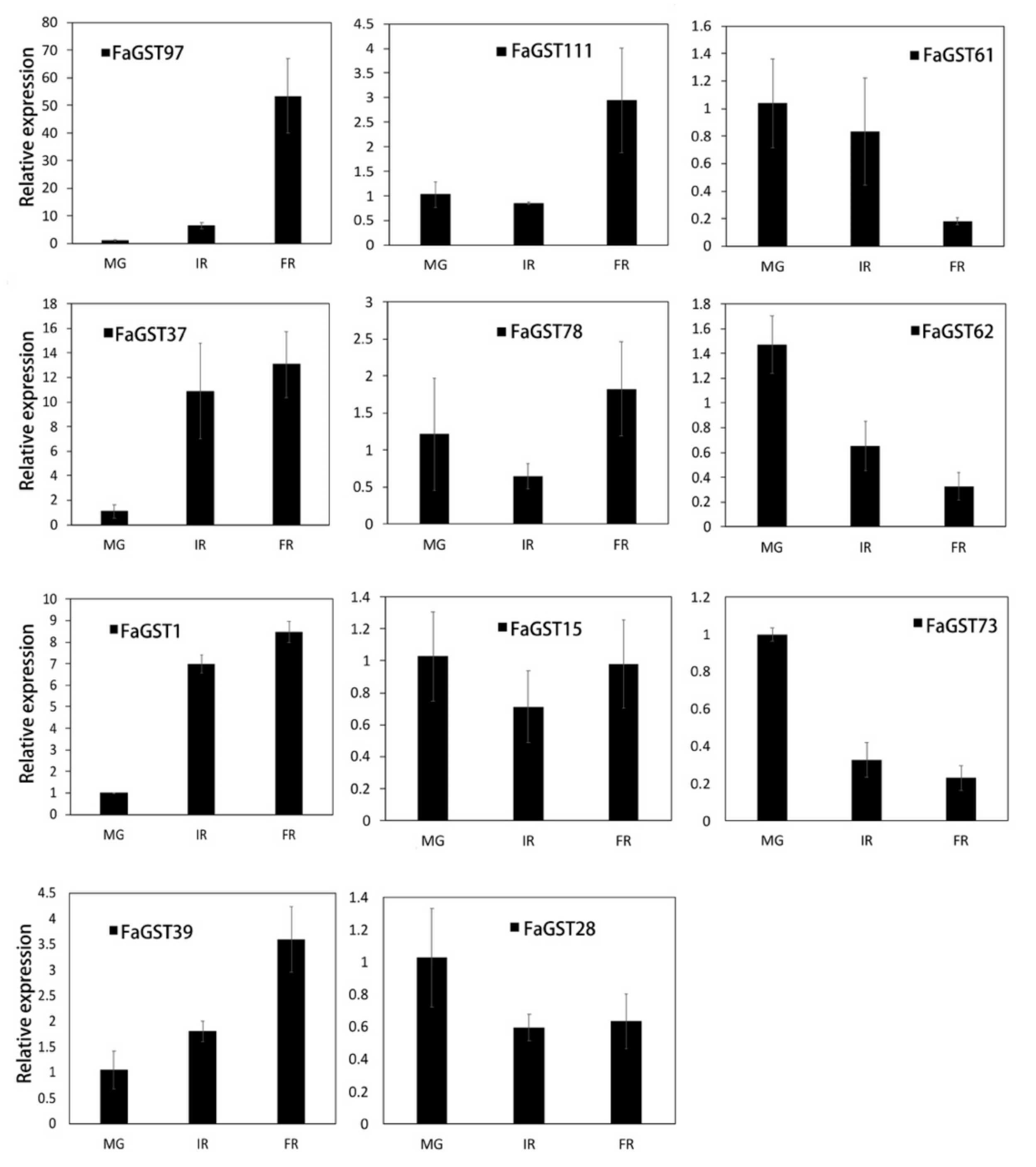
2.4. Expression Analysis of FaGST Genes in Different Developmental Stages
2.5. Anthocyanins Accumulation and Expression Analysis of Anthocyanin Biosynthetic Structural Genes
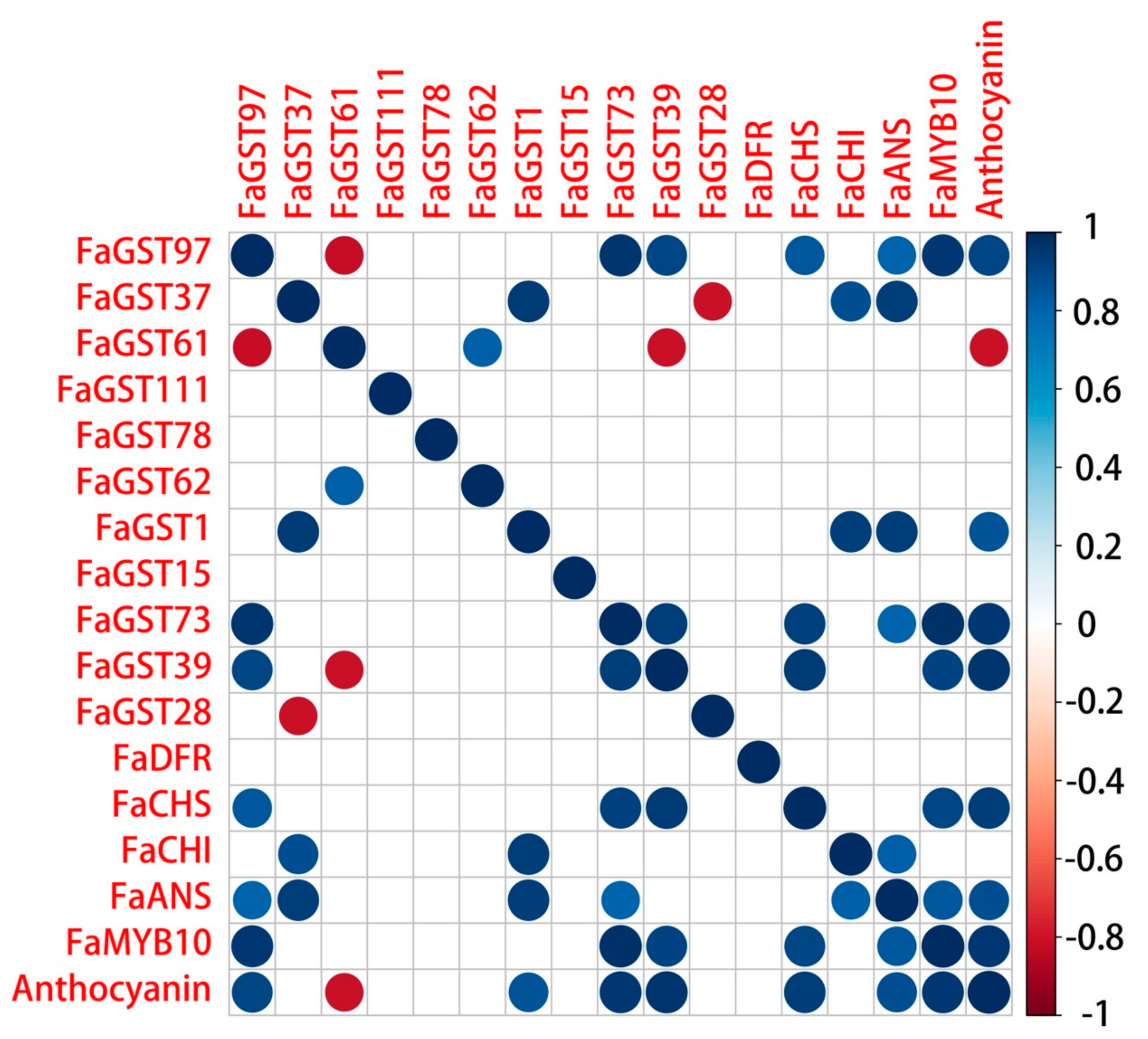
2.6. Correlation Analysis of Anthocyanin Biosynthetic Structural Genes and FaGST Genes
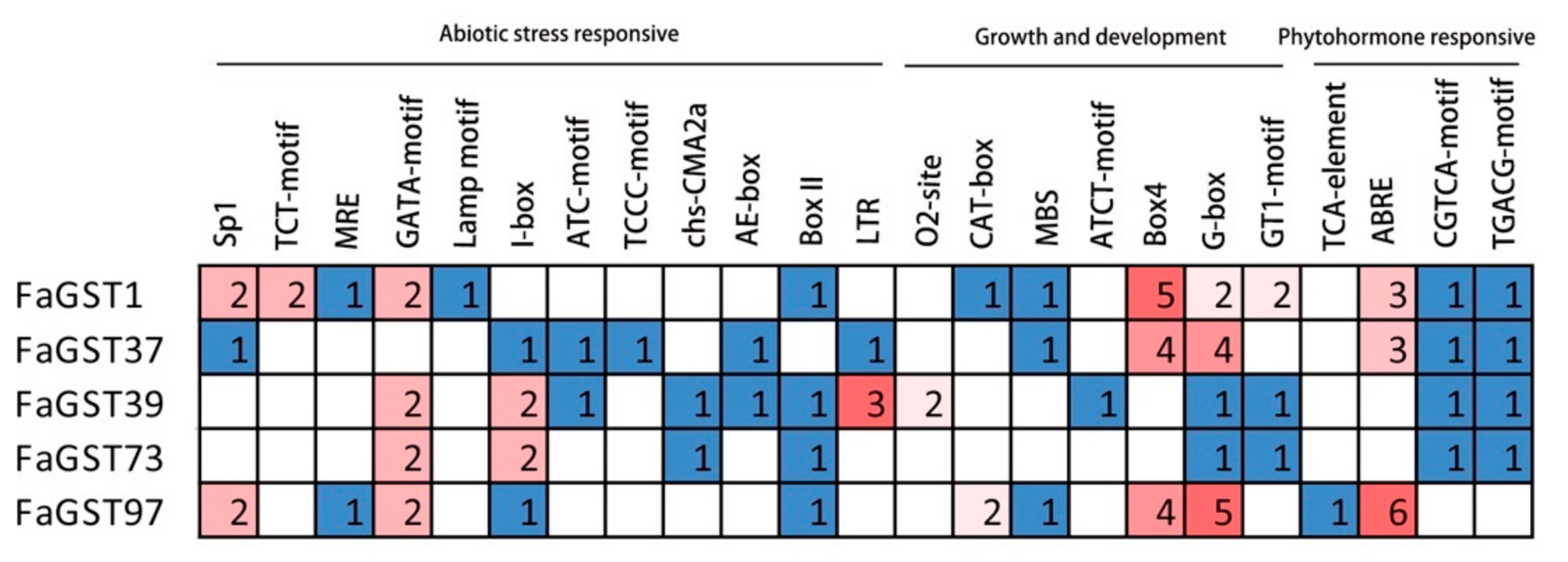
2.7. Identification of Cis-Regulatory Elements in the Promoters of Putative Anthocyanin-Related FaGST Genes
3. Discussion
3.1. Identification and Comprehensive Analysis of the Entire FaGST Genes
3.2. Gene Ontology and Classification Revealed the Putative Anthocyanin-Related FaGST Genes
3.3. Expression and Correlation Analysis Confirmed the Involvement of FaGST in Strawberry Anthocyanins Accumulation
4. Materials and Methods
4.1. Plant Materials
4.2. Identification and Comprehensive Analysis of FaGST Genes
4.3. Phylogenetic and Evolutionary Analysis of FaGST Genes in Strawberry
4.4. Expression Analysis of FaGST Genes in Strawberry
4.5. Measurement of Anthocyanins Content
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pillet, J.; Folta, K.M. Pigments in Strawberry. In Pigments in Fruits and Vegetables; Chen, C., Ed.; Springer Science & Business Media: NewYork, NY, USA, 2015; pp. 205–216. [Google Scholar]
- Peña-Sanhueza, D.; Inostroza-Blancheteau, C.; Ribera-Fonseca, A.; Reyes-Díaz, M. Anthocyanins in Berries and Their Potential Use in Human Health. In Superfood and Functional Food-The Development of Superfoods and Their Roles as Medicine; Shiomi, N., Waisundara, V., Eds.; InTechOpen: London, UK, 2017; pp. 155–172. [Google Scholar]
- He, J.; Giusti, M.M. Anthocyanins: Natural colorants with health-promoting properties. Annu. Rev. Food Sci. Technol. 2010, 1, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Winkelshirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Han, Y.; Vimolmangkang, S.; Soria-Guerra, R.E.; Rosales-Mendoza, S.; Zheng, D.; Lygin, A.V.; Korban, S.S. Ectopic expression of apple F3′H genes contributes to anthocyanin accumulation in the Arabidopsis tt7 mutant grown under nitrogen stress. Plant. Physiol 2010, 153, 806–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Xiao, M.; Luo, Y.; Wang, J.; Wang, H. The effect of RNAi-induced silencing of FaDFR on anthocyanin metabolism in strawberry (Fragaria × ananassa) fruit. Sci. Hortic. 2013, 160, 123–128. [Google Scholar] [CrossRef]
- Jiang, F.; Wang, J.-Y.; Jia, H.-F.; Jia, W.-S.; Wang, H.-Q.; Xiao, M. RNAi-mediated silencing of the flavanone 3-hydroxylase gene and its effect on flavonoid biosynthesis in strawberry fruit. J. Plant. Growth Regul. 2013, 32, 182–190. [Google Scholar] [CrossRef]
- Baudry, A.; Heim, M.A.; Dubreucq, B.; Caboche, M.; Weisshaar, B.; Lepiniec, L. TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana. Plant. J. 2004, 39, 366–380. [Google Scholar] [CrossRef]
- Xu, W.; Dubos, C.; Lepiniec, L. Transcriptional control of flavonoid biosynthesis by MYB-bHLH-WDR complexes. Trends Plant. Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef]
- Lin-Wang, K.; McGhie, T.K.; Wang, M.; Liu, Y.; Warren, B.; Storey, R.; Espley, R.V.; Allan, A.C. Engineering the anthocyanin regulatory complex of strawberry (Fragaria vesca). Front. Plant. Sci. 2014, 5, 651. [Google Scholar] [CrossRef] [Green Version]
- Medina-Puche, L.; Cumplido-Laso, G.; Amil-Ruiz, F.; Hoffmann, T.; Ring, L.; Rodríguez-Franco, A.; Caballero, J.L.; Schwab, W.; Muñoz-Blanco, J.; Blanco-Portales, R. MYB10 plays a major role in the regulation of flavonoid/phenylpropanoid metabolism during ripening of Fragaria × ananassa fruits. J. Exp. Bot. 2013, 65, 401–417. [Google Scholar] [CrossRef] [Green Version]
- Schaart, J.G.; Dubos, C.; Romero De La Fuente, I.; Houwelingen, A.M.; Vos, R.C.; Jonker, H.H.; Xu, W.; Routaboul, J.M.; Lepiniec, L.; Bovy, A.G. Identification and characterization of MYB-bHLH-WD40 regulatory complexes controlling proanthocyanidin biosynthesis in strawberry (Fragaria × ananassa) fruits. New Phytol. 2013, 197, 454–467. [Google Scholar] [CrossRef]
- Zhao, J.; Dixon, R.A. The ‘ins’ and ‘outs’ of flavonoid transport. Trends Plant. Sci. 2010, 15, 72–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, C.D.; Casati, P.; Walbot, V. A multidrug resistance-associated protein involved in anthocyanin transport in Zea mays. Plant. Cell 2004, 16, 1812–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, C.; Terrier, N.; Torregrosa, L.; Vialet, S.; Fournier-Level, A.; Verries, C.; Souquet, J.-M.; Mazauric, J.-P.; Klein, M.; Cheynier, V. Grapevine MATE-type proteins act as vacuolar H+-dependent acylated anthocyanin transporters. Plant. Physiol. 2009, 150, 402–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Huhman, D.; Shadle, G.; He, X.-Z.; Sumner, L.W.; Tang, Y.; Dixon, R.A. MATE2 mediates vacuolar sequestration of flavonoid glycosides and glycoside malonates in Medicago truncatula. Plant. Cell 2011, 23, 1536–1555. [Google Scholar] [CrossRef] [Green Version]
- Marinova, K.; Pourcel, L.; Weder, B.; Schwarz, M.; Barron, D.; Routaboul, J.-M.; Debeaujon, I.; Klein, M. The Arabidopsis MATE transporter TT12 acts as a vacuolar flavonoid/H+-antiporter active in proanthocyanidin-accumulating cells of the seed coat. Plant. Cell 2007, 19, 2023–2038. [Google Scholar] [CrossRef] [Green Version]
- Francisco, R.M.; Regalado, A.; Ageorges, A.; Burla, B.J.; Bassin, B.; Eisenach, C.; Zarrouk, O.; Vialet, S.; Marlin, T.; Chaves, M.M. ABCC1, an ATP binding cassette protein from grape berry, transports anthocyanidin 3-O-glucosides. Plant. Cell 2013, 25, 1840–1854. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, C.; Hoffmann, T.; Escobar, N.M.; Ludemann, F.; Botella, M.A.; Valpuesta, V.; Schwab, W. The strawberry fruit Fra a allergen functions in flavonoid biosynthesis. Mol. Plant. 2010, 3, 113–124. [Google Scholar] [CrossRef]
- Loyall, L.; Uchida, K.; Braun, S.; Furuya, M.; Frohnmeyer, H. Glutathione and a UV light–induced glutathione S-transferase are involved in signaling to chalcone synthase in cell cultures. Plant. Cell 2000, 12, 1939–1950. [Google Scholar]
- Agrawal, G.K.; Jwa, N.-S.; Rakwal, R. A pathogen-induced novel rice (Oryza sativa L.) gene encodes a putative protein homologous to type II glutathione S-transferases. Plant. Sci. 2002, 163, 1153–1160. [Google Scholar] [CrossRef]
- Liu, Y.-J.; Han, X.-M.; Ren, L.-L.; Yang, H.-L.; Zeng, Q.-Y. Functional divergence of the glutathione S-transferase supergene family in Physcomitrella patens reveals complex patterns of large gene family evolution in land plants. Plant. Physiol. 2013, 161, 773–786. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, S.; Shikazono, N.; Tanaka, A. TRANSPARENT TESTA 19 is involved in the accumulation of both anthocyanins and proanthocyanidins in Arabidopsis. Plant. J. 2004, 37, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Mueller, L.A.; Goodman, C.D.; Silady, R.A.; Walbot, V. AN9, a petunia glutathione S-transferase required for anthocyanin sequestration, is a flavonoid-binding protein. Plant. Physiol. 2000, 123, 1561–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Díaz, R.; Madrid-Espinoza, J.; Salinas-Cornejo, J.; González-Villanueva, E.; Ruiz-Lara, S. Differential roles for VviGST1, VviGST3, and VviGST4 in proanthocyanidin and anthocyanin transport in Vitis vinifera. Front. Plant. Sci. 2016, 7, 1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, S.; Akita, Y.; Ishizaka, H.; Narumi, I.; Tanaka, A. Molecular characterization of an anthocyanin-related glutathione S-transferase gene in cyclamen. J. Plant. Physiol. 2012, 169, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Zhao, J.; Lai, B.; Qin, Y.; Wang, H.; Hu, G. LcGST4 is an anthocyanin-related glutathioneS-transferase gene in Litchi chinensis Sonn. Plant. Cell Rep. 2016, 35, 831–843. [Google Scholar] [CrossRef]
- El-Sharkawy, I.; Liang, D.; Xu, K. Transcriptome analysis of an apple (Malus × domestica) yellow fruit somatic mutation identifies a gene network module highly associated with anthocyanin and epigenetic regulation. J. Exp. Bot. 2015, 66, 7359–7376. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Liao, L.; Zhou, H.; Gu, C.; Wang, L.; Han, Y. A small indel mutation in an anthocyanin transporter causes variegated colouration of peach flowers. J. Exp. Bot. 2015, 66, 7227–7239. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Dai, C.; Li, Y.; Feng, J.; Liu, Z.; Kang, C. Reduced Anthocyanins in Petioles codes for a GST anthocyanin transporter that is essential for the foliage and fruit coloration in strawberry. J. Exp. Bot. 2018, 69, 2595–2608. [Google Scholar] [CrossRef] [Green Version]
- Sondheimer, E.; Karash, C.B. The Major Anthocyanin Pigments of the Wild Strawberry (Fragaria vesca). Nature 1956, 178, 648–649. [Google Scholar] [CrossRef]
- Wang, S.Y.; Zheng, W.; Galletta, G.J. Cultural system affects fruit quality and antioxidant capacity in strawberries. J. Agric. Food Chem. 2002, 50, 6534–6542. [Google Scholar] [CrossRef]
- Donno, D.; Cavanna, M.; Beccaro, G.; Mellano, M.; Torello-Marinoni, D.; Cerutti, A.; Bounous, G. Currants and strawberries as bioactive compound sources: Determination of antioxidant profiles with HPLC-DAD/MS. J. Appl. Bot. Food Qual. 2013, 86, 1–10. [Google Scholar]
- Sappl, P.G.; Carroll, A.J.; Clifton, R.; Lister, R.; Whelan, J.; Harvey Millar, A.; Singh, K.B. The Arabidopsisglutathione transferase gene family displays complex stress regulation and co-silencing multiple genes results in altered metabolic sensitivity to oxidative stress. Plant. J. 2009, 58, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Licciardello, C.; D’Agostino, N.; Traini, A.; Recupero, G.R.; Frusciante, L.; Chiusano, M.L. Characterization of the glutathione S-transferase gene family through ESTs and expression analyses within common and pigmented cultivars of Citrus sinensis (L.) Osbeck. BMC Plant. Biol. 2014, 14, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, Z.; Wu, J.; Han, X.; Wang-Pruski, G.; Zhang, Z. Genome-wide identification, characterization, and expression analysis related to autotoxicity of the GST gene family in Cucumis melo L. Plant. Physiol. Biochem. 2020, 155, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Kayum, A.; Nath, U.K.; Park, J.-I.; Biswas, M.K.; Choi, E.K.; Song, J.-Y.; Kim, H.-T.; Nou, I.-S. Genome-wide identification, characterization, and expression profiling of glutathione S-transferase (GST) family in pumpkin reveals likely role in cold-stress tolerance. Genes 2018, 9, 84. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.S.; Choudhury, M.; Majlish, A.-N.K.; Islam, T.; Ghosh, A. Comprehensive genome-wide analysis of glutathione S-transferase gene family in potato (Solanum tuberosum L.) and their expression profiling in various anatomical tissues and perturbation conditions. Gene 2018, 639, 149–162. [Google Scholar] [CrossRef]
- Proulx, S.R. Multiple routes to subfunctionalization and gene duplicate specialization. Genetics 2012, 190, 737–751. [Google Scholar] [CrossRef] [Green Version]
- Magadum, S.; Murugan, P.; Gangapur, D.; Ravikesavan, R. Gene duplication as a major force in evolution. J. Genet. 2013, 92, 155–161. [Google Scholar] [CrossRef]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of Gene Duplication in Plants. Plant. Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [Green Version]
- Marques, A.C.; Vinckenbosch, N.; Brawand, D.; Kaessmann, H. Functional diversification of duplicate genes through subcellular adaptation of encoded proteins. Genome Biol. 2008, 9, R54. [Google Scholar] [CrossRef] [Green Version]
- Vaish, S.; Awasthi, P.; Tiwari, S.; Tiwari, S.K.; Gupta, D.; Basantani, M.K. In silico genome-wide identification and characterization of the glutathione S-transferase gene family in Vigna radiata. Genome 2018, 61, 311–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, M.; Deutsch, M. Association of intron phases with conservation at splice site sequences and evolution of spliceosomal introns. Mol. Biol. Evol. 1999, 16, 1528–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, Y.-H.E.; Bahn, J.H.; Lin, X.; Chan, T.-M.; Wang, R.; Xiao, X. Alternative splicing modulated by genetic variants demonstrates accelerated evolution regulated by highly conserved proteins. Genome Res. 2016, 26, 440–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labrou, N.E.; Papageorgiou, A.C.; Pavli, O.; Flemetakis, E. Plant GSTome: Structure and functional role in xenome network and plant stress response. Curr. Opin. Biotechnol. 2015, 32, 186–194. [Google Scholar] [CrossRef]
- Nishida, M.; Harada, S.; Noguchi, S.; Satow, Y.; Inoue, H.; Takahashi, K. Three-dimensional structure of Escherichia coli glutathione S-transferase complexed with glutathione sulfonate: Catalytic roles of Cys10 and His106. J. Mol. Biol 1998, 281, 135–147. [Google Scholar] [CrossRef]
- Oakley, A.J. Glutathione transferases: New functions. Curr. Opin. Struct. Biol. 2005, 15, 716–723. [Google Scholar] [CrossRef]
- Dixon, D.P.; Skipsey, M.; Edwards, R. Roles for glutathione transferases in plant secondary metabolism. Phytochemistry 2010, 71, 338–350. [Google Scholar] [CrossRef]
- Marrs, K.A.; Alfenito, M.R.; Lloyd, A.M.; Walbot, V. A glutathione S-transferase involved in vacuolar transfer encoded by the maize gene Bronze-2. Nature 1995, 375, 397–400. [Google Scholar] [CrossRef]
- Larsen, E.; Alfenito, M.; Briggs, W.; Walbot, V. A carnation anthocyanin mutant is complemented by the glutathione S-transferases encoded by maize Bz2 and petunia An9. Plant. Cell Rep. 2003, 21, 900–904. [Google Scholar] [CrossRef]
- Conn, S.; Curtin, C.; Bézier, A.; Franco, C.; Zhang, W. Purification, molecular cloning, and characterization of glutathione S-transferases (GSTs) from pigmented Vitis vinifera L. cell suspension cultures as putative anthocyanin transport proteins. J. Exp. Bot. 2008, 59, 3621–3634. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.-H.; Li, H.-L.; Guo, D.; Wang, Y.; Dai, H.-F.; Mei, W.-L.; Peng, S.-Q. Transcriptome-wide identification and expression analysis of glutathione S-transferase genes involved in flavonoids accumulation in Dracaena cambodiana. Plant. Physiol. Biochem. 2016, 104, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Lee, T.; Cheng, C.-H.; Buble, K.; Zheng, P.; Yu, J.; Humann, J.; Ficklin, S.P.; Gasic, K.; Scott, K.; et al. 15 years of GDR: New data and functionality in the Genome Database for Rosaceae. Nucleic Acids Res. 2019, 47, D1137–D1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangradorvegas, A. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Aron, M.B.; Shennan, L.; Anderson, J.B.; Farideh, C.; Derbyshire, M.K.; Carol, D.W.S.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R. CDD: A Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2011, 39, D225–D229. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Zhang, Z.; Li, J.; Zhao, X.Q.; Wang, J.; Wong, K.S.; Yu, J. KaKs_Calculator: Calculating Ka and Ks Through Model Selection and Model Averaging. Genom. Proteom. Bioinform. 2006, 4, 259–263. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene1 | Gene2 | Ka | Ks | Ka/Ks | p-Value (Fisher) |
|---|---|---|---|---|---|
| FaGST25 | FaGST93 | 0.00200317 | 0.113747 | 0.0176108 | 1.06 × 10−9 |
| FaGST2 | FaGST100 | 0.00165273 | 0.048333 | 0.0341946 | 1.84 × 10−5 |
| FaGST12 | FaGST114 | 0.00375505 | 0.0832149 | 0.0451247 | 3.23 × 10−7 |
| FaGST25 | FaGST123 | 0.00804594 | 0.0584878 | 0.137566 | 0.00106652 |
| FaGST12 | FaGST81 | 0.00375505 | 0.0246355 | 0.152424 | 0.0300419 |
| FaGST6 | FaGST42 | 0.0244688 | 0.143011 | 0.171097 | 6.23 × 10−7 |
| FaGST46 | FaGST78 | 0.00492559 | 0.02814 | 0.175039 | 0.0651319 |
| FaGST79 | FaGST111 | 0.00787615 | 0.0439795 | 0.179087 | 0.00889461 |
| FaGST34 | FaGST96 | 0.0102566 | 0.0560355 | 0.183037 | 0.00310094 |
| FaGST26 | FaGST76 | 0.0202591 | 0.109809 | 0.184494 | 6.95 × 10−19 |
| FaGST13 | FaGST47 | 0.173126 | 0.886179 | 0.195362 | 4.53 × 10−17 |
| FaGST13 | FaGST79 | 0.18648 | 0.93119 | 0.20026 | 5.53 × 10−17 |
| FaGST32 | FaGST40 | 0.00760364 | 0.0369972 | 0.205519 | 0.00105106 |
| FaGST18 | FaGST92 | 0.0104022 | 0.0497516 | 0.209083 | 0.00447566 |
| FaGST58 | FaGST94 | 0.313243 | 1.48572 | 0.210836 | 9.04 × 10−19 |
| FaGST32 | FaGST104 | 0.0164355 | 0.0760217 | 0.216195 | 5.48 × 10−6 |
| FaGST33 | FaGST113 | 0.0280173 | 0.128595 | 0.217873 | 0.000335365 |
| FaGST4 | FaGST44 | 0.0471441 | 0.216188 | 0.21807 | 2.45 × 10−7 |
| FaGST22 | FaGST54 | 0.30599 | 1.31495 | 0.232701 | 8.57 × 10−16 |
| FaGST71 | FaGST125 | 0.0254999 | 0.0973965 | 0.261815 | 0.000205114 |
| FaGST15 | FaGST112 | 0.0116752 | 0.0444317 | 0.262766 | 0.0242682 |
| FaGST29 | FaGST9 | 0.00382776 | 0.0142017 | 0.269529 | 0.340562 |
| FaGST29 | FaGST65 | 0.0333278 | 0.122567 | 0.271916 | 0.000450127 |
| FaGST18 | FaGST116 | 0.0113206 | 0.0411117 | 0.275361 | 0.0294135 |
| FaGST10 | FaGST66 | 0.00308679 | 0.0103946 | 0.29696 | 0.355313 |
| FaGST38 | FaGST105 | 0.0127675 | 0.0425923 | 0.29976 | 0.0330291 |
| FaGST36 | FaGST57 | 0.0244337 | 0.0796125 | 0.306908 | 0.00426484 |
| FaGST16 | FaGST107 | 0.0136969 | 0.0414432 | 0.330497 | 0.0194532 |
| FaGST20 | FaGST118 | 0.332674 | 0.989292 | 0.336274 | 1.33 × 10−10 |
| FaGST15 | FaGST46 | 0.00987768 | 0.0282057 | 0.350201 | 0.234251 |
| FaGST18 | FaGST51 | 0.0145916 | 0.0410609 | 0.355365 | 0.0477493 |
| FaGST34 | FaGST67 | 0.0425944 | 0.117162 | 0.363551 | 0.00353968 |
| FaGST26 | FaGST45 | 0.0827313 | 0.226323 | 0.365546 | 4.19 × 10−14 |
| FaGST1 | FaGST37 | 0.0221609 | 0.0555156 | 0.399184 | 0.124764 |
| FaGST23 | FaGST74 | 0.0150372 | 0.0375368 | 0.400598 | 0.0659631 |
| FaGST36 | FaGST58 | 0.0337835 | 0.0776749 | 0.434935 | 0.0314613 |
| FaGST38 | FaGST73 | 0.00794979 | 0.0179958 | 0.441758 | 0.281114 |
| FaGST24 | FaGST122 | 0.884215 | 1.82613 | 0.484203 | 0.000255179 |
| FaGST24 | FaGST86 | 0.919184 | 1.72577 | 0.532623 | 0.000805302 |
| FaGST11 | FaGST124 | 0.0431261 | 0.0770174 | 0.559953 | 0.117755 |
| FaGST34 | FaGST129 | 0.171667 | 0.292162 | 0.587576 | 0.0453954 |
| FaGST10 | FaGST130 | 0.0303226 | 0.0492918 | 0.615164 | 0.251029 |
| FaGST13 | FaGST110 | 0.0505542 | 0.0807071 | 0.626392 | 0.190343 |
| FaGST17 | FaGST121 | 0.0854666 | 0.134315 | 0.636314 | 0.127269 |
| FaGST11 | FaGST70 | 0.0894935 | 0.139369 | 0.642133 | 0.065153 |
| FaGST6 | FaGST101 | 0.0314943 | 0.0481056 | 0.65469 | 0.319248 |
| FaGST31 | FaGST103 | 0.0558196 | 0.084181 | 0.66309 | 0.366314 |
| FaGST19 | FaGST91 | 0.286378 | 0.421856 | 0.678852 | 0.0104161 |
| FaGST35 | FaGST95 | 0.0128558 | 0.0185847 | 0.691739 | 0.499311 |
| FaGST22 | FaGST88 | 0.0768175 | 0.106394 | 0.72201 | 0.279675 |
| FaGST30 | FaGST89 | 0.177649 | 0.243199 | 0.730469 | 0.117698 |
| FaGST35 | FaGST126 | 0.0173247 | 0.0233682 | 0.74138 | 0.562354 |
| FaGST10 | FaGST97 | 0.083706 | 0.111366 | 0.751627 | 0.358856 |
| FaGST12 | FaGST48 | 0.0844299 | 0.111337 | 0.758326 | 0.31708 |
| FaGST52 | FaGST119 | 0.249063 | 0.327341 | 0.760868 | 0.178488 |
| FaGST30 | FaGST52 | 0.220105 | 0.288024 | 0.764189 | 0.168512 |
| FaGST35 | FaGST71 | 0.0215799 | 0.0279284 | 0.772687 | 0.603323 |
| FaGST32 | FaGST75 | 0.167423 | 0.216594 | 0.772981 | 0.131165 |
| FaGST17 | FaGST87 | 0.0160792 | 0.0205336 | 0.783069 | 0.683377 |
| FaGST35 | FaGST128 | 0.0208453 | 0.0255813 | 0.814864 | 0.591106 |
| FaGST50 | FaGST115 | 0.0302207 | 0.035657 | 0.84754 | 0.701964 |
| FaGST1 | FaGST98 | 0.0286463 | 0.0326903 | 0.876291 | 0.710666 |
| FaGST27 | FaGST61 | 0.17827 | 0.198328 | 0.898862 | 0.724287 |
| FaGST35 | FaGST68 | 0.017273 | 0.0138736 | 1.24503 | 0.989446 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.; Zhang, L.; Zhang, J.; Zhang, Y.; Wang, Y.; Chen, Q.; Luo, Y.; Zhang, Y.; Li, M.; Wang, X.; et al. Identification of Anthocyanins-Related Glutathione S-Transferase (GST) Genes in the Genome of Cultivated Strawberry (Fragaria × ananassa). Int. J. Mol. Sci. 2020, 21, 8708. https://doi.org/10.3390/ijms21228708
Lin Y, Zhang L, Zhang J, Zhang Y, Wang Y, Chen Q, Luo Y, Zhang Y, Li M, Wang X, et al. Identification of Anthocyanins-Related Glutathione S-Transferase (GST) Genes in the Genome of Cultivated Strawberry (Fragaria × ananassa). International Journal of Molecular Sciences. 2020; 21(22):8708. https://doi.org/10.3390/ijms21228708
Chicago/Turabian StyleLin, Yuanxiu, Lianxi Zhang, JiaHao Zhang, Yunting Zhang, Yan Wang, Qing Chen, Ya Luo, Yong Zhang, Mengyao Li, Xiaorong Wang, and et al. 2020. "Identification of Anthocyanins-Related Glutathione S-Transferase (GST) Genes in the Genome of Cultivated Strawberry (Fragaria × ananassa)" International Journal of Molecular Sciences 21, no. 22: 8708. https://doi.org/10.3390/ijms21228708