7-HYB, a Phenolic Compound Isolated from Myristica fragrans Houtt Increases Cell Migration, Osteoblast Differentiation, and Mineralization through BMP2 and β-catenin Signaling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
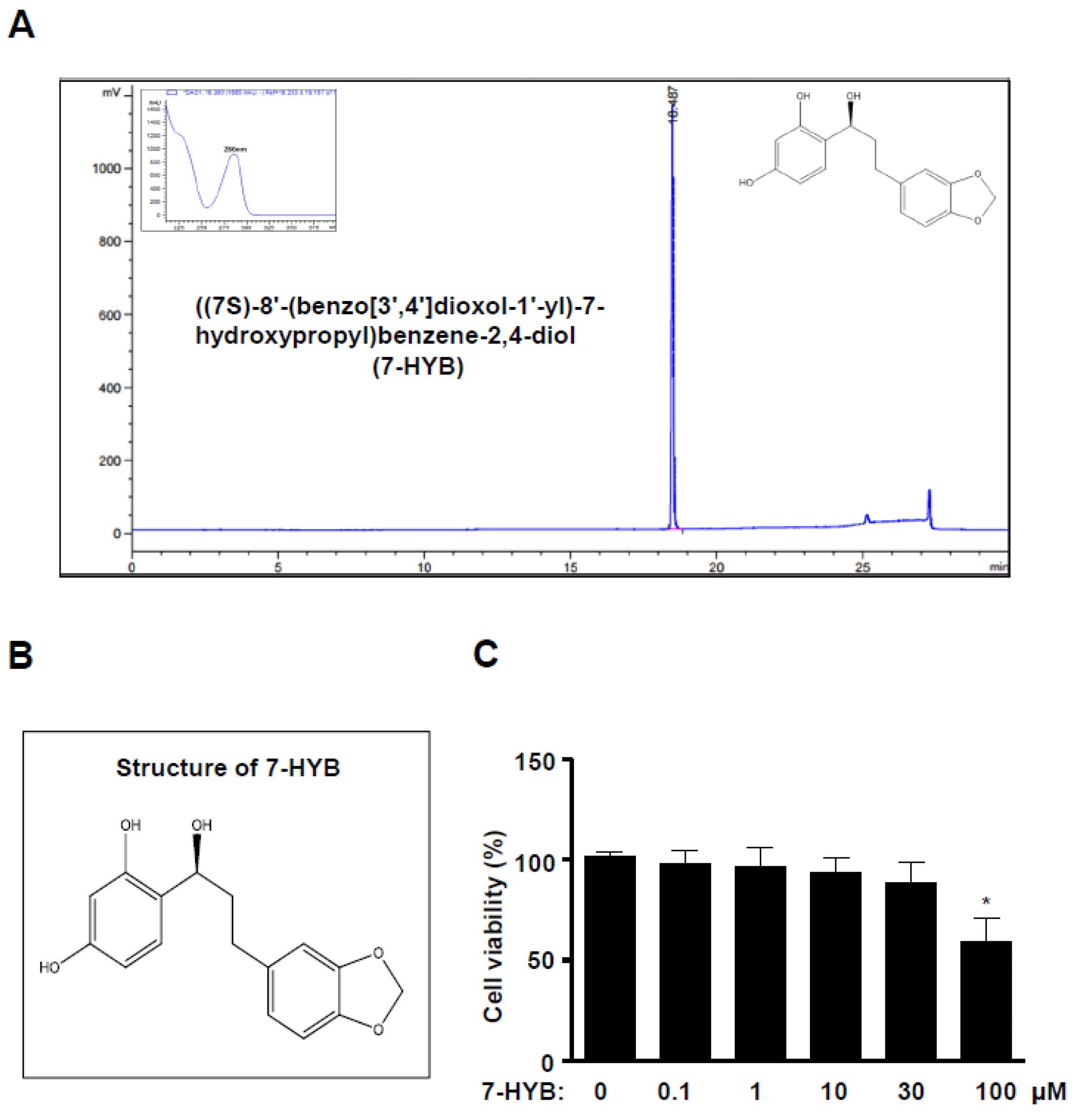
2.1. 7-HYB Has No Effect on the Cell Toxicity in Preosteoblasts
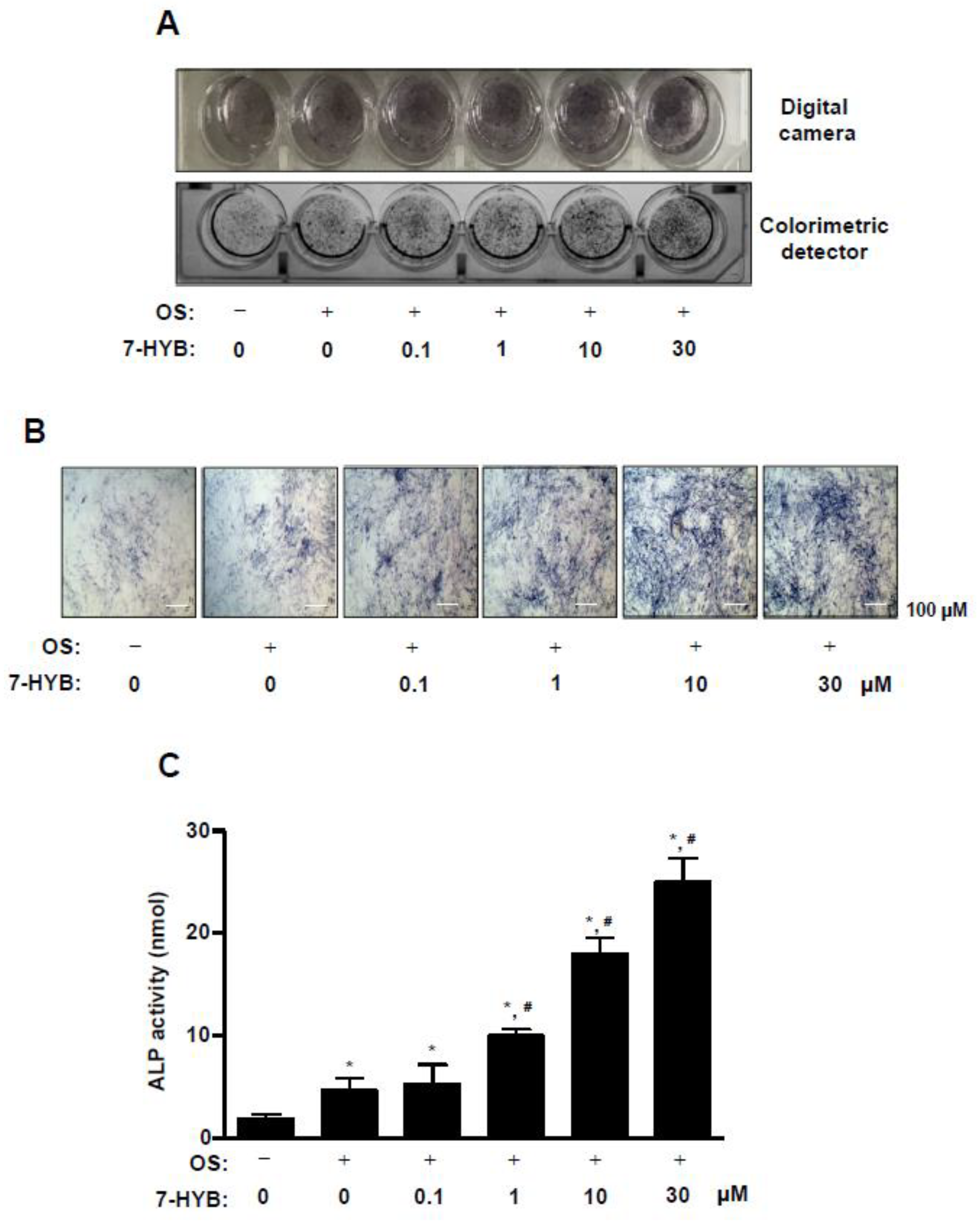
2.2. 7-HYB Promotes the Early Osteoblast Differentiation of Preosteoblasts
2.3. 7-HYB Enhances the Late Osteoblast Differentiation of Preosteoblasts
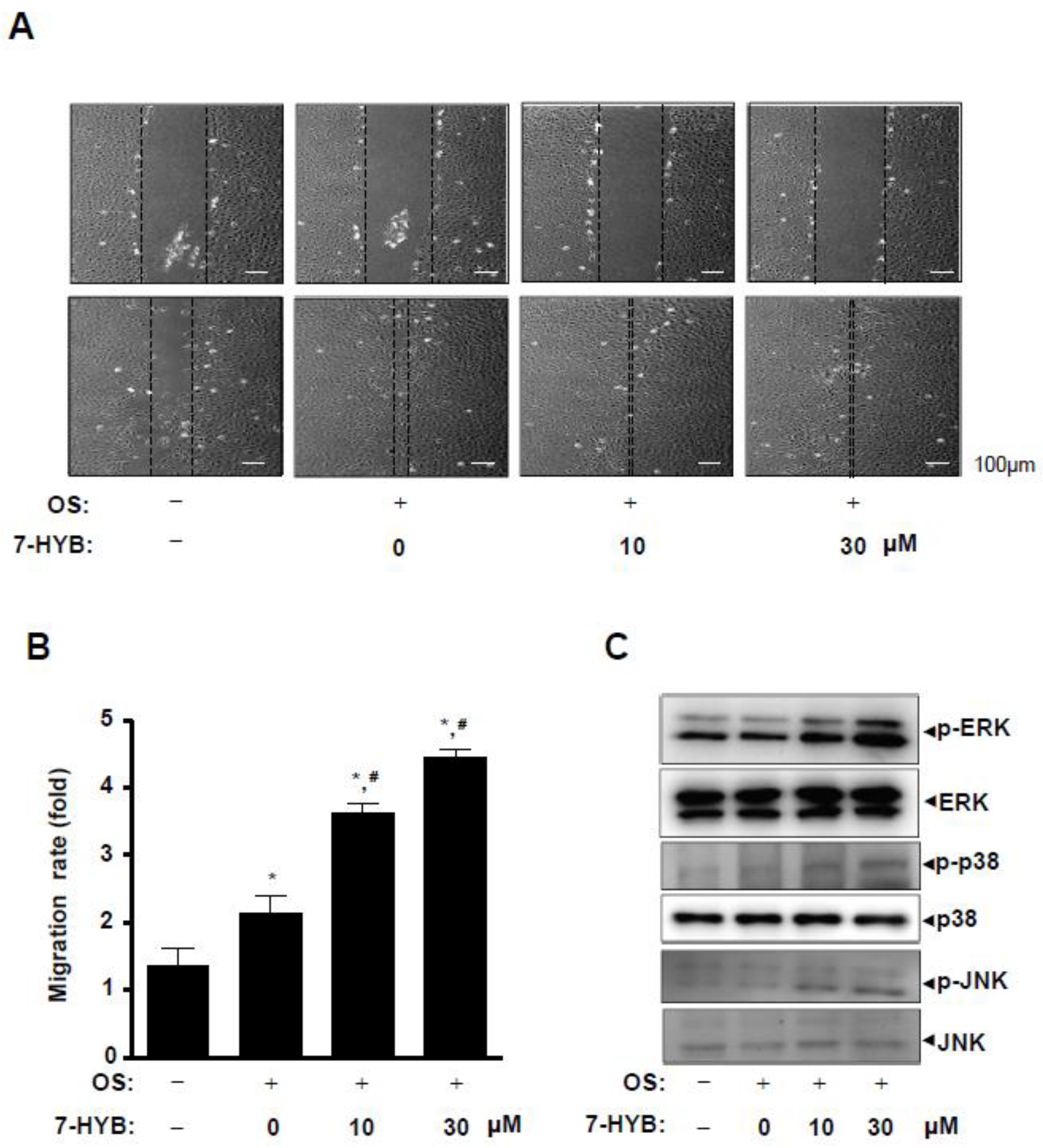
2.4. 7-HYB Increases Cell Migration in Osteoblast Differentiation of Preosteoblasts
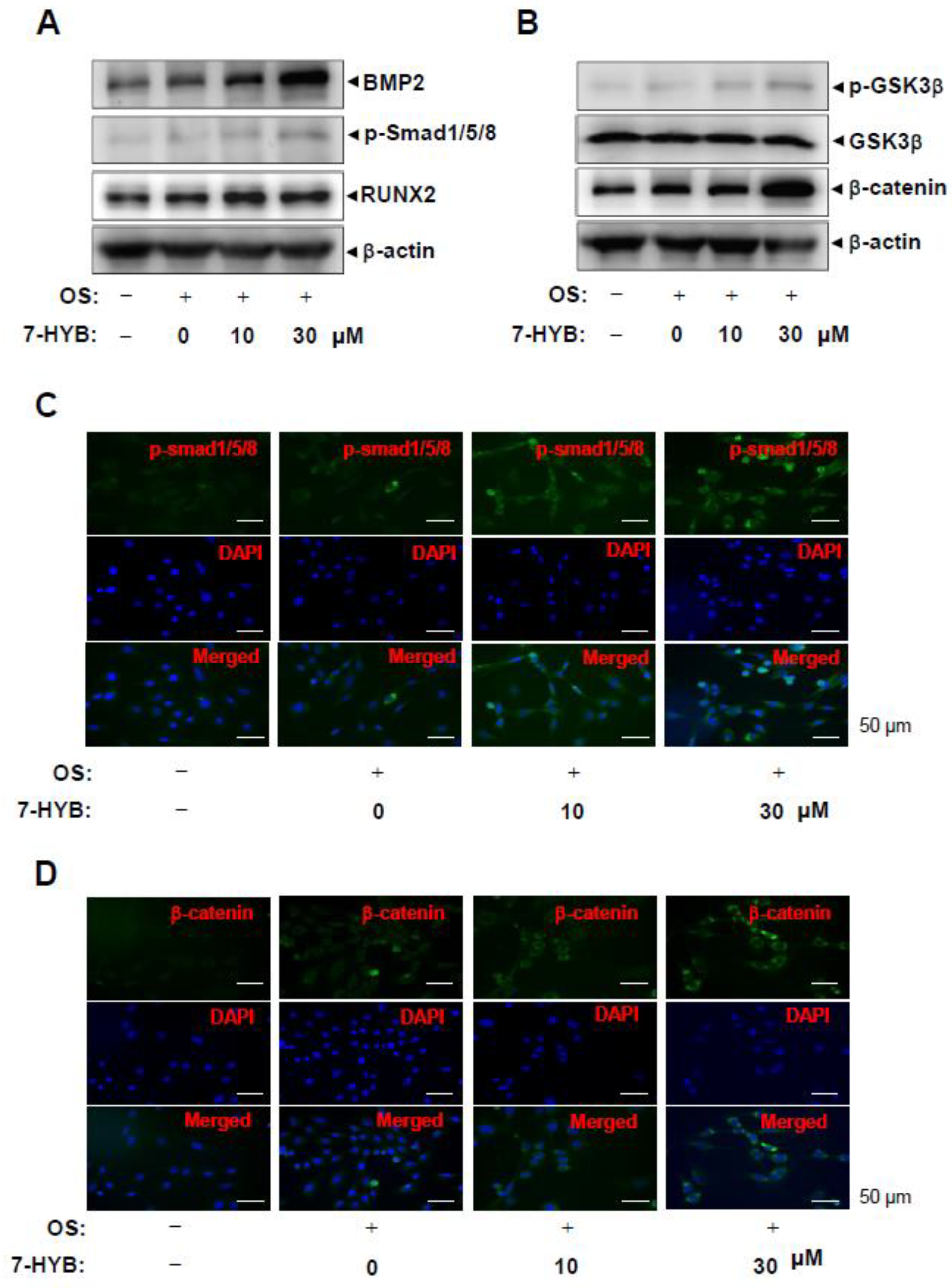
2.5. 7-HYB Stimulates BMP2-Smad1/5/8-RUNX2 and β-catenin Signaling in Osteoblast Differentiation
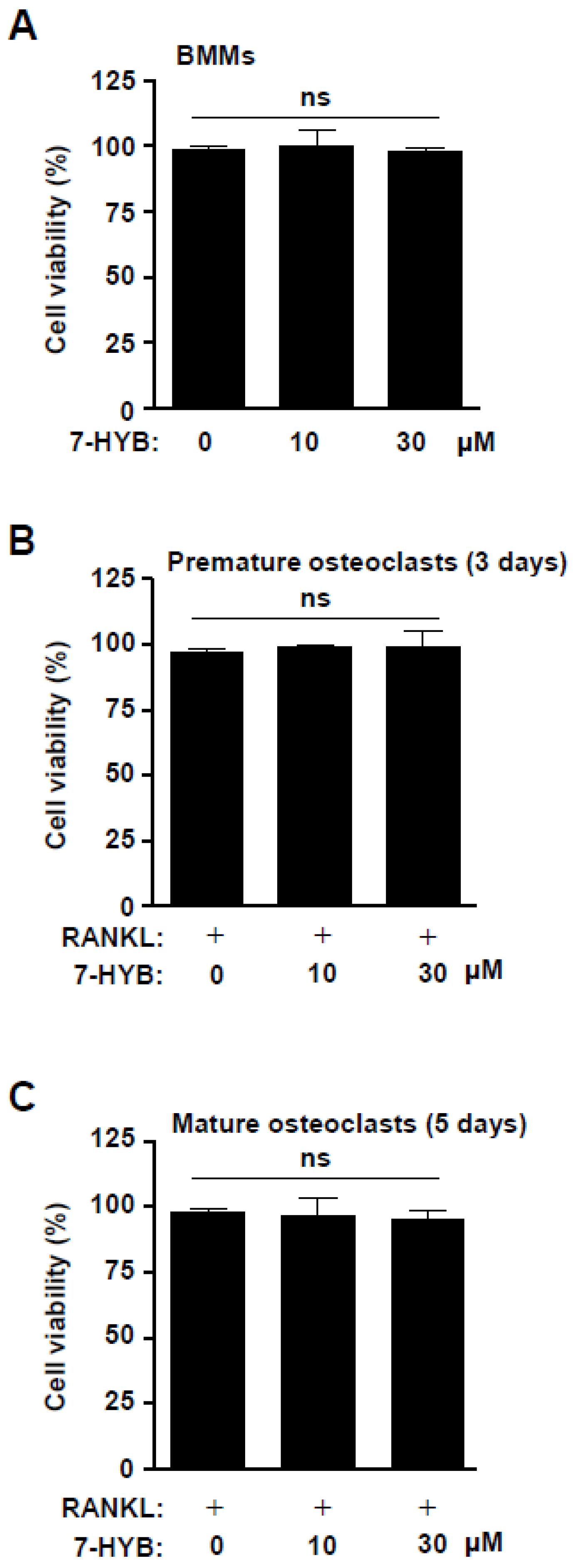
2.6. 7-HYB Does Not Affect the Cell Toxicity in Bone Marrow Macrophages (BMMs), Premature Osteoclasts, and Mature Osteoclasts
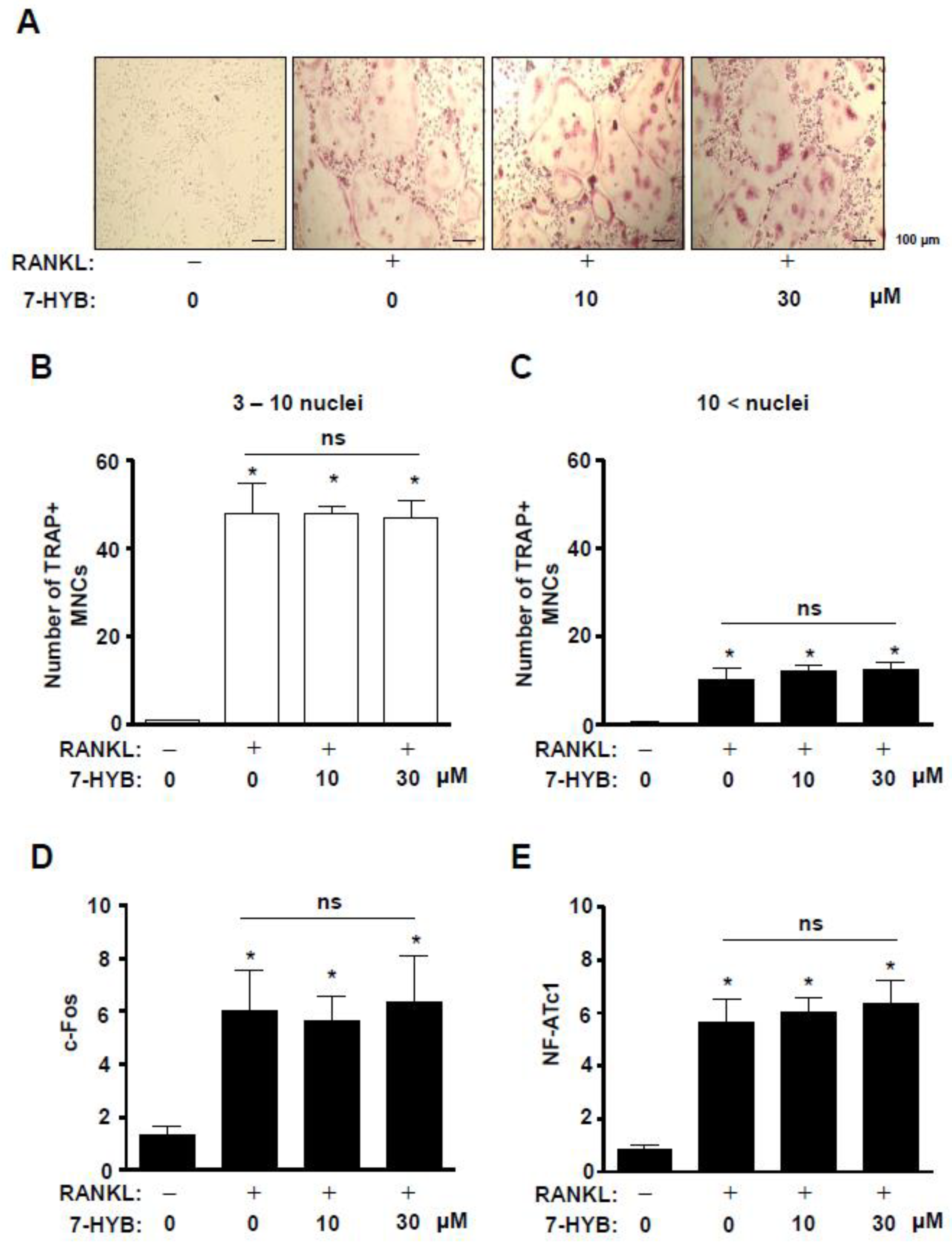
2.7. 7-HYB Has No Biological Activities on TRAP-Positive Multinucleated Osteoclasts (MNCs) and Gene Expression in RANKL-Induced Osteoclastogenesis
3. Discussion
4. Materials and Methods
4.1. Extraction and Isolation of 7-((7S)-8′-(benzo[3′,4′]dioxol-1′-yl)-7-hydroxypropyl)benzene-2,4-diol
4.2. ((7S)-8′-(benzo[3′,4′]dioxol-1′-yl)-7-hydroxypropyl)benzene-2,4-diol (7-HYB)
4.3. Nuclear Magnetic Resonance (NMR)
4.4. Culture of MC3T3E-1 Preosteoblasts, and Differentiation of Osteoblasts
4.5. MTT Assay
4.6. Western Blot Analysis
4.7. Cell Migration Assay
4.8. Alkaline Phosphatase (ALP) Staining Assay
4.9. ALP Activity Assay
4.10. Alizarin Red S (ARS) Staining
4.11. Immunocytochemistry
4.12. Live Subject Statement
4.13. Culture of Bone Marrow Macrophages, and Osteoclast Differentiation
4.14. Tartrate-Resistant Acid Phosphatase (TRAP) Staining
4.15. Quantitative Real-Time Polymerase Chain Reaction (PCR) Analysis
4.16. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Karsenty, G. The complexities of skeletal biology. Nature 2003, 423, 316–318. [Google Scholar] [CrossRef]
- An, J.; Yang, H.; Zhang, Q.; Liu, C.; Zhao, J.; Zhang, L.; Chen, B. Natural products for treatment of osteoporosis: The effects and mechanisms on promoting osteoblast-mediated bone formation. Life Sci. 2016, 147, 46–58. [Google Scholar] [CrossRef]
- Seeman, E. Pathogenesis of bone fragility in women and men. Lancet 2002, 359, 1841–1850. [Google Scholar] [CrossRef]
- Khosla, S.; Riggs, B.L. Pathophysiology of age-related bone loss and osteoporosis. Endocrinol. Metab. Clin. N. Am. 2005, 34, 1015–1030. [Google Scholar] [CrossRef]
- Marie, P.J. Osteoblast dysfunctions in bone diseases: From cellular and molecular mechanisms to therapeutic strategies. Cell. Mol. Life Sci. 2015, 72, 1347–1361. [Google Scholar] [CrossRef]
- Vondracek, S.F.; Minne, P.; McDermott, M.T. Clinical challenges in the management of osteoporosis. Clin. Interv. Aging 2008, 3, 315–329. [Google Scholar] [CrossRef] [Green Version]
- Kawai, M.; Modder, U.I.; Khosla, S.; Rosen, C.J. Emerging therapeutic opportunities for skeletal restoration. Nat. Rev. Drug Discov. 2011, 10, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Marie, P.J.; Kassem, M. Osteoblasts in osteoporosis: Past, emerging, and future anabolic targets. Eur. J. Endocrinol. 2011, 165, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Adjene, J.O.; Nwose, E.U. Histological effects of long term consumption of nutmeg on the medial geniculate body of adult Wistar rats. N. Am. J. Med. Sci. 2010, 2, 134–137. [Google Scholar] [CrossRef]
- Barceloux, D.G. Nutmeg (Myristica fragrans Houtt.). Dis. Mon. 2009, 55, 373–379. [Google Scholar] [CrossRef]
- Van Gils, C.; Cox, P.A. Ethnobotany of nutmeg in the Spice Islands. J. Ethnopharmacol. 1994, 42, 117–124. [Google Scholar] [CrossRef]
- Abourashed, E.A.; El-Alfy, A.T. Chemical diversity and pharmacological significance of the secondary metabolites of nutmeg (Myristica fragrans Houtt.). Phytochem. Rev. 2016, 15, 1035–1056. [Google Scholar] [CrossRef] [Green Version]
- Cuong, T.D.; Hung, T.M.; Na, M.; do, T.H.; Kim, J.C.; Lee, D.; Ryoo, S.; Lee, J.H.; Choi, J.S.; Min, B.S. Inhibitory effect on NO production of phenolic compounds from Myristica fragrans. Bioorg. Med. Chem. Lett. 2011, 21, 6884–6887. [Google Scholar] [CrossRef]
- Duan, L.; Tao, H.W.; Hao, X.J.; Gu, Q.Q.; Zhu, W.M. Cytotoxic and antioxidative phenolic compounds from the traditional Chinese medicinal plant, Myristica fragrans. Planta Med. 2009, 75, 1241–1245. [Google Scholar] [CrossRef] [Green Version]
- El-Alfy, A.T.; Abourashed, E.A.; Patel, C.; Mazhari, N.; An, H.; Jeon, A. Phenolic compounds from nutmeg (Myristica fragrans Houtt.) inhibit the endocannabinoid-modulating enzyme fatty acid amide hydrolase. J. Pharm. Pharmacol. 2019, 71, 1879–1889. [Google Scholar] [CrossRef]
- Guntur, A.R.; Rosen, C.J. The skeleton: A multi-functional complex organ: New insights into osteoblasts and their role in bone formation: The central role of PI3Kinase. J. Endocrinol. 2011, 211, 123–130. [Google Scholar] [CrossRef]
- Lee, H.S.; Jung, E.Y.; Bae, S.H.; Kwon, K.H.; Kim, J.M.; Suh, H.J. Stimulation of osteoblastic differentiation and mineralization in MC3T3-E1 cells by yeast hydrolysate. Phytother. Res. 2011, 25, 716–723. [Google Scholar] [CrossRef]
- Kim, M.B.; Song, Y.; Hwang, J.K. Kirenol stimulates osteoblast differentiation through activation of the BMP and Wnt/beta-catenin signaling pathways in MC3T3-E1 cells. Fitoterapia 2014, 98, 59–65. [Google Scholar] [CrossRef]
- Katagiri, T.; Yamaguchi, A.; Komaki, M.; Abe, E.; Takahashi, N.; Ikeda, T.; Rosen, V.; Wozney, J.M.; Fujisawa-Sehara, A.; Suda, T. Bone morphogenetic protein-2 converts the differentiation pathway of C2C12 myoblasts into the osteoblast lineage. J. Cell Biol. 1994, 127, 1755–1766. [Google Scholar] [CrossRef] [Green Version]
- Wozney, J.M.; Rosen, V.; Celeste, A.J.; Mitsock, L.M.; Whitters, M.J.; Kriz, R.W.; Hewick, R.M.; Wang, E.A. Novel regulators of bone formation: Molecular clones and activities. Science 1988, 242, 1528–1534. [Google Scholar] [CrossRef]
- Miyazono, K.; Kamiya, Y.; Morikawa, M. Bone morphogenetic protein receptors and signal transduction. J. Biochem. 2010, 147, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Canalis, E.; Economides, A.N.; Gazzerro, E. Bone morphogenetic proteins, their antagonists, and the skeleton. Endocr. Rev. 2003, 24, 218–235. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Kim, Y.J.; Kim, H.J.; Park, H.D.; Kang, A.R.; Kyung, H.M.; Sung, J.H.; Wozney, J.M.; Ryoo, H.M. BMP-2-induced Runx2 expression is mediated by Dlx5, and TGF-beta 1 opposes the BMP-2-induced osteoblast differentiation by suppression of Dlx5 expression. J. Biol. Chem. 2003, 278, 34387–34394. [Google Scholar] [CrossRef] [Green Version]
- Gaur, T.; Lengner, C.J.; Hovhannisyan, H.; Bhat, R.A.; Bodine, P.V.; Komm, B.S.; Javed, A.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; et al. Canonical WNT signaling promotes osteogenesis by directly stimulating Runx2 gene expression. J. Biol. Chem. 2005, 280, 33132–33140. [Google Scholar] [CrossRef] [Green Version]
- Phimphilai, M.; Zhao, Z.; Boules, H.; Roca, H.; Franceschi, R.T. BMP signaling is required for RUNX2-dependent induction of the osteoblast phenotype. J. Bone Miner. Res. 2006, 21, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Rawadi, G.; Vayssiere, B.; Dunn, F.; Baron, R.; Roman-Roman, S. BMP-2 controls alkaline phosphatase expression and osteoblast mineralization by a Wnt autocrine loop. J. Bone Miner. Res. 2003, 18, 1842–1853. [Google Scholar] [CrossRef]
- Fukuda, T.; Kokabu, S.; Ohte, S.; Sasanuma, H.; Kanomata, K.; Yoneyama, K.; Kato, H.; Akita, M.; Oda, H.; Katagiri, T. Canonical Wnts and BMPs cooperatively induce osteoblastic differentiation through a GSK3beta-dependent and beta-catenin-independent mechanism. Differentiation 2010, 80, 46–52. [Google Scholar] [CrossRef]
- Zhang, R.; Oyajobi, B.O.; Harris, S.E.; Chen, D.; Tsao, C.; Deng, H.W.; Zhao, M. Wnt/beta-catenin signaling activates bone morphogenetic protein 2 expression in osteoblasts. Bone 2013, 52, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.F.; Li, G.; Chan, C.Y.; Meng, C.L.; Lin, M.C.; Chen, Y.C.; He, M.L.; Leung, P.C.; Kung, H.F. Flavonoids of Herba Epimedii regulate osteogenesis of human mesenchymal stem cells through BMP and Wnt/beta-catenin signaling pathway. Mol. Cell Endocrinol. 2010, 314, 70–74. [Google Scholar] [CrossRef]
- Day, T.F.; Guo, X.; Garrett-Beal, L.; Yang, Y. Wnt/beta-catenin signaling in mesenchymal progenitors controls osteoblast and chondrocyte differentiation during vertebrate skeletogenesis. Dev. Cell 2005, 8, 739–750. [Google Scholar] [CrossRef] [Green Version]
- Lo, Y.C.; Chang, Y.H.; Wei, B.L.; Huang, Y.L.; Chiou, W.F. Betulinic acid stimulates the differentiation and mineralization of osteoblastic MC3T3-E1 cells: Involvement of BMP/Runx2 and beta-catenin signals. J. Agric. Food Chem. 2010, 58, 6643–6649. [Google Scholar] [CrossRef]
- Bodine, P.V.; Komm, B.S. Wnt signaling and osteoblastogenesis. Rev. Endocr. Metab. Disord. 2006, 7, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Andersen, T.E.; Bollerslev, J.; Mandrup, S.; Abdallah, B.M.; Kassem, M. Patients with high bone mass phenotype exhibit enhanced osteoblast differentiation and inhibition of adipogenesis of human mesenchymal stem cells. J. Bone Miner. Res. 2007, 22, 1720–1731. [Google Scholar] [CrossRef]
- MacDonald, B.T.; He, X. Frizzled and LRP5/6 receptors for Wnt/beta-catenin signaling. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Reya, T.; Clevers, H. Wnt signalling in stem cells and cancer. Nature 2005, 434, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Moon, R.T.; Bowerman, B.; Boutros, M.; Perrimon, N. The promise and perils of Wnt signaling through beta-catenin. Science 2002, 296, 1644–1646. [Google Scholar] [CrossRef] [PubMed]
- Ghiasi, M.S.; Chen, J.; Vaziri, A.; Rodriguez, E.K.; Nazarian, A. Bone fracture healing in mechanobiological modeling: A review of principles and methods. Bone Rep. 2017, 6, 87–100. [Google Scholar] [CrossRef]
- Enoki, Y.; Sato, T.; Kokabu, S.; Hayashi, N.; Iwata, T.; Yamato, M.; Usui, M.; Matsumoto, M.; Tomoda, T.; Ariyoshi, W.; et al. Netrin-4 Promotes Differentiation and Migration of Osteoblasts. In Vivo 2017, 31, 793–799. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, H.; Zhang, H. Kaempferol promotes proliferation, migration and differentiation of MC3T3-E1 cells via up-regulation of microRNA-101. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1050–1056. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Kim, M.; Jho, E.H. Wnt/beta-catenin signalling: From plasma membrane to nucleus. Biochem. J. 2013, 450, 9–21. [Google Scholar] [CrossRef]
- Westhoff, M.A.; Serrels, B.; Fincham, V.J.; Frame, M.C.; Carragher, N.O. SRC-mediated phosphorylation of focal adhesion kinase couples actin and adhesion dynamics to survival signaling. Mol. Cell. Biol. 2004, 24, 8113–8133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.M.; Shih, Y.T.; Chen, Y.S.; Liu, C.L.; Fang, W.K.; Tsai, C.H.; Tsai, F.J.; Kuo, W.W.; Lai, T.Y.; Huang, C.Y. Schwann Cell Migration Induced by Earthworm Extract via Activation of PAs and MMP2/9 Mediated through ERK1/2 and p38. Evid. Based Complement. Altern. Med. 2011, 2011, 395458. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Liu, F.C.; Chou, P.Y.; Chien, Y.C.; Chang, W.S.; Huang, G.J.; Wu, C.H.; Sheu, M.J. Ethanol extracts of fruiting bodies of Antrodia cinnamomea suppress CL1-5 human lung adenocarcinoma cells migration by inhibiting matrix metalloproteinase-2/9 through ERK, JNK, p38, and PI3K/Akt signaling pathways. Evid. Based Complement. Altern. Med. 2012, 2012, 378415. [Google Scholar] [CrossRef] [Green Version]
- Liao, X.; Lu, S.; Zhuo, Y.; Winter, C.; Xu, W.; Wang, Y. Visualization of Src and FAK activity during the differentiation process from HMSCs to osteoblasts. PLoS ONE 2012, 7, e42709. [Google Scholar] [CrossRef]
- Soelaiman, I.N.; Das, S.; Shuid, A.N.; Mo, H.; Mohamed, N. Use of medicinal plants and natural products for treatment of osteoporosis and its complications. Evid. Based Complement. Altern. Med. 2013, 2013, 764701. [Google Scholar] [CrossRef]
- Whelan, A.M.; Jurgens, T.M.; Bowles, S.K. Natural health products in the prevention and treatment of osteoporosis: Systematic review of randomized controlled trials. Ann. Pharmacother. 2006, 40, 836–849. [Google Scholar] [CrossRef]
- Bordea, R.; Lucaciu, O.; Câmpian, R.S. Student’s knowledge and opinion regarding the need of implementation of Lasers in Dental Faculty curriculum. Hum. Vet. Med. 2016, 8, 157–160. [Google Scholar]
- Bordea, I.R.; Lucaciu, P.O.; Crişan, B.; Pelekanos, S.; Câmpian, R.S. The influence f chromophore presence in an experimental bleaching gel on laser assisted tooth whitening efficiency. Studia Ubb Chemia. 2016, 2, 215–223. [Google Scholar]
- Inchingolo, F.; Tatullo, M.; Pacifici, A.; Abenavoli, F.M.; Pacifici, L. Use of dermal-fat grafts in the post-oncological reconstructive surgery of atrophies in the zygomatic region: Clinical evaluations in the patients undergone to previous radiation therapy. Head Face Med. 2012, 8, 33. [Google Scholar] [CrossRef] [Green Version]
- Park, K.R.; Lee, H.; Cho, M.; Yun, H.M. A Phytochemical Constituent, (E)-Methyl-Cinnamate Isolated from Alpinia katsumadai Hayata Suppresses Cell Survival, Migration, and Differentiation in Preosteoblasts. Int. J. Mol. Sci. 2020, 21, 3700. [Google Scholar] [CrossRef]
- Park, K.R.; Yun, H.M. RANKL-induced osteoclastogenesis in bone marrow-derived macrophages is suppressed by cisapride. Toxicology 2019, 422, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Park, K.R.; Kim, E.C.; Hong, J.T.; Yun, H.M. Dysregulation of 5-hydroxytryptamine 6 receptor accelerates maturation of bone-resorbing osteoclasts and induces bone loss. Theranostics 2018, 8, 3087–3098. [Google Scholar] [CrossRef] [PubMed]
- Park, K.R.; Yun, H.M.; Hong, J.T. G721-0282 inhibits cell growth and induces apoptosis in human osteosarcoma through down-regulation of the STAT3 pathway. Int. J. Biol. Sci. 2020, 16, 330–341. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, K.-R.; Kwon, Y.-J.; Park, J.-E.; Yun, H.-M. 7-HYB, a Phenolic Compound Isolated from Myristica fragrans Houtt Increases Cell Migration, Osteoblast Differentiation, and Mineralization through BMP2 and β-catenin Signaling. Int. J. Mol. Sci. 2020, 21, 8059. https://doi.org/10.3390/ijms21218059
Park K-R, Kwon Y-J, Park J-E, Yun H-M. 7-HYB, a Phenolic Compound Isolated from Myristica fragrans Houtt Increases Cell Migration, Osteoblast Differentiation, and Mineralization through BMP2 and β-catenin Signaling. International Journal of Molecular Sciences. 2020; 21(21):8059. https://doi.org/10.3390/ijms21218059
Chicago/Turabian StylePark, Kyung-Ran, Yoon-Ju Kwon, Ji-Eun Park, and Hyung-Mun Yun. 2020. "7-HYB, a Phenolic Compound Isolated from Myristica fragrans Houtt Increases Cell Migration, Osteoblast Differentiation, and Mineralization through BMP2 and β-catenin Signaling" International Journal of Molecular Sciences 21, no. 21: 8059. https://doi.org/10.3390/ijms21218059