The Mechanism of Bisdemethoxycurcumin Enhances Conventional Antibiotics against Methicillin-Resistant Staphylococcus aureus


Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Susceptibility Testing and Synergic Effect
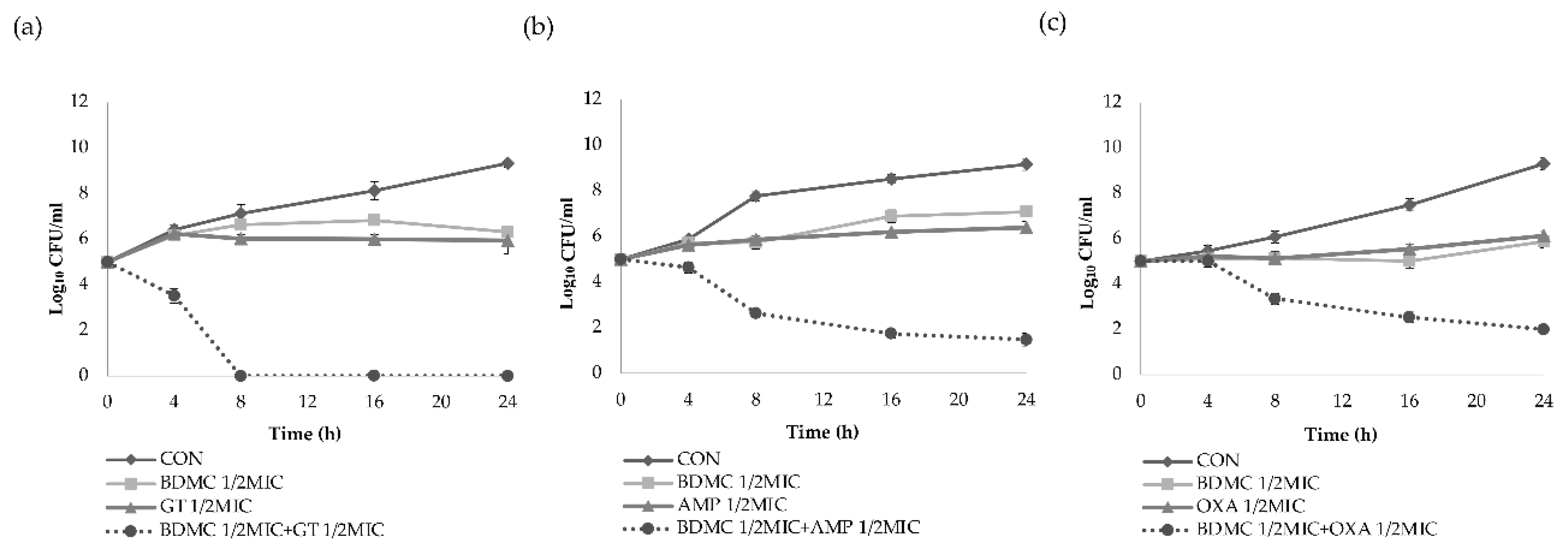
2.2. Time–Kill Curve Assay
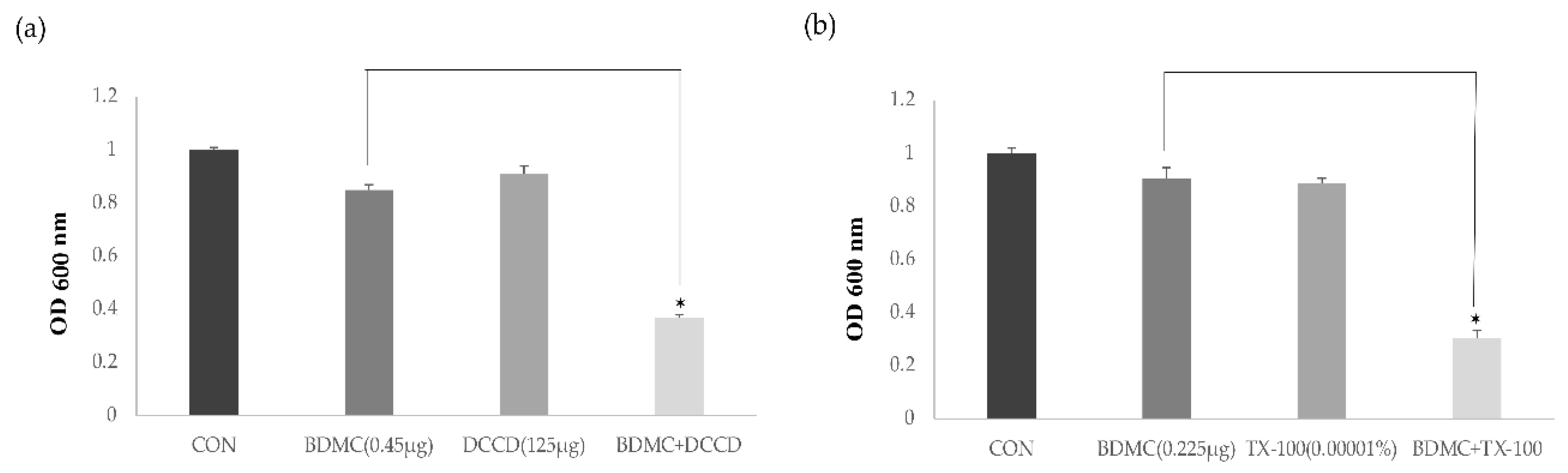
2.3. Antimicrobial Activity with the ATPase Inhibitor and the Membrane-Permeabilizing Agent
2.4. Expression of PBP2a in Methicillin-Resistant Staphylococcus aureus (MRSA)
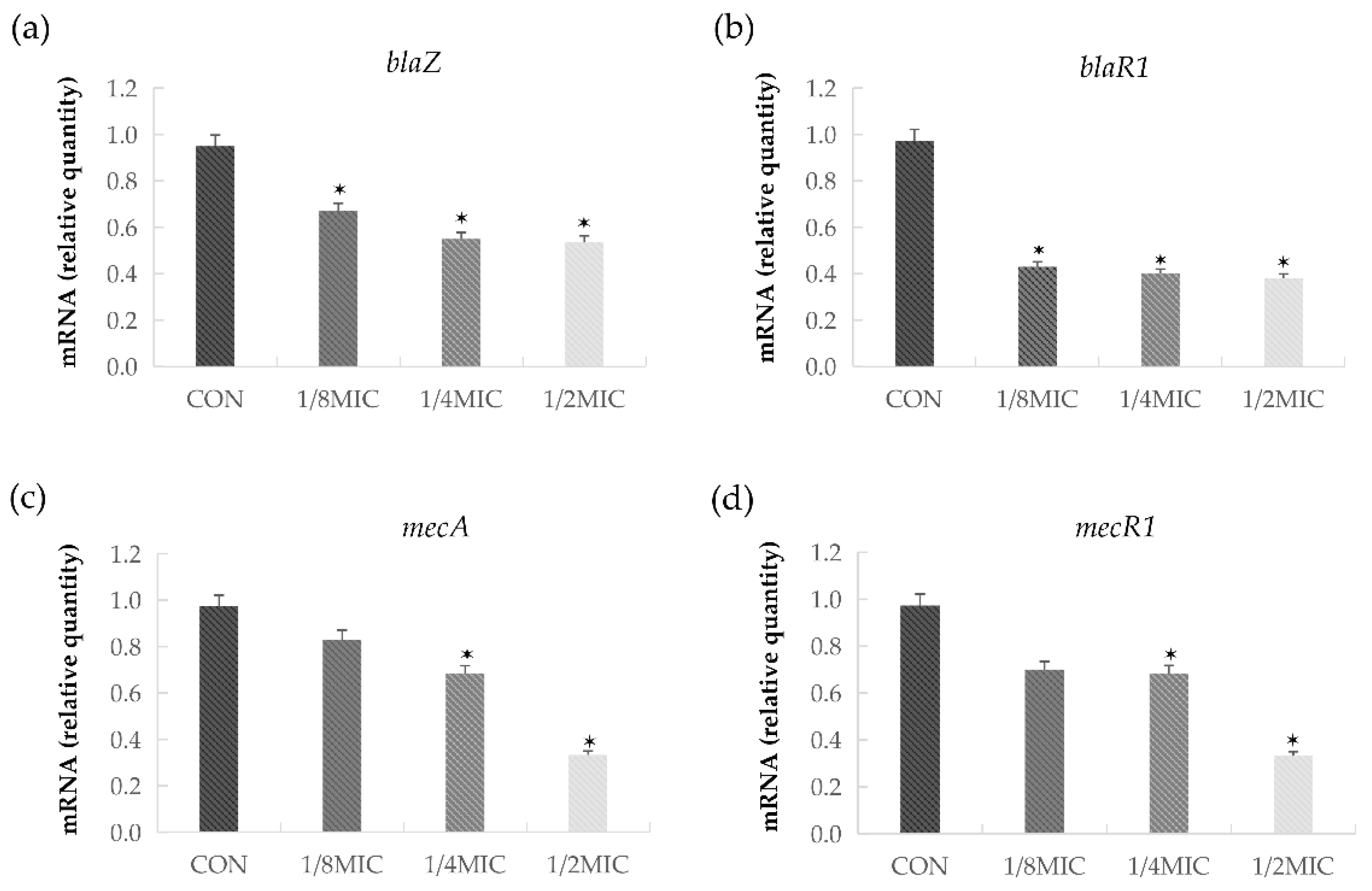
2.5. BDMC Represses the Transcription of MecA, BlaZ, BlaR1, and MecR1 in S. aureus
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Bacterial Strains and Growth Medium
4.3. Susceptibility Testing of BDMC with Antibiotics
4.4. Time–Kill Curve Assay
4.5. Antibacterial Activity of BDMC in the Presence of ATPase Inhibitor or the Membrane-Permeabilizing Agent
4.6. Western Blot Analysis
4.7. Reverse Transcription and qRT-PCR
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Zhen, X.; Lundborg, C.S.; Zhang, M.; Sun, X.; Li, Y.; Hu, X.; Gu, S.; Gu, Y.; Wei, J.; Dong, H. Clinical and economic impact of methicillin-resistant Staphylococcus aureus: A multicentre study in China. Sci. Rep. 2020, 10, 60825–60826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, C.L.; Dalecki, A.G.; Perez, M.D.; Schaaf, K.; Wolschendorf, F.; Kutsch, O. A copper-dependent compound restores ampicillin sensitivity in multidrug-resistant Staphylococcus aureus. Sci. Rep. 2020, 10, 8955. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, F.; Torres, V.J. The bicomponent pore-forming leucocidins of Staphylococcus aureus. Microbiol. Mol. Biol. Rev. 2014, 78, 199–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. 2020, 33, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- Alfouzan, W.; Udo, E.E.; Modhaffer, A.; Alosaimi, A. Molecular Characterization of Methicillin-Resistant Staphylococcus aureus in a Tertiary Care hospital in Kuwait. Sci. Rep. 2019, 9, 18527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddad Kashani, H.; Schmelcher, M.; Sabzalipoor, H.; Seyed Hosseini, E.; Moniri, R. Recombinant Endolysins as Potential Therapeutics against Antibiotic-Resistant Staphylococcus aureus: Current Status of Research and Novel Delivery Strategies. Clin. Microbiol. Rev. 2017, 31, e00071-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, M. Community-associated MRSA: What makes them special. Int. J. Med. Microbiol. 2013, 303, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Miceli, M.; Bontempo, P.; Nebbioso, A.; Altucci, L. Natural compounds in epigenetics: A current view. Food Chem. Toxicol. 2014, 73, 71–83. [Google Scholar] [CrossRef]
- Sinha, D.; Biswas, J.; Sung, B.; Aggarwal, B.B.; Bishayee, A. Chemopreventive and chemotherapeutic potential of curcumin in breast cancer. Curr. Drug Targets. 2012, 13, 1799–1819. [Google Scholar] [CrossRef]
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “Curecumin”: From kitchen to clinic. Biochem. Pharmacol. 2008, 75, 787–809. [Google Scholar] [CrossRef] [Green Version]
- Itaya, M.; Miyazawa, T.; Zingg, J.M.; Eitsuka, T.; Azzi, A.; Meydani, M.; Miyazawa, T.; Nakagawa, K. The differential cellular uptake of curcuminoids in vitro depends dominantly on albumin interaction. Phytomedicine 2019, 59, 152902. [Google Scholar] [CrossRef]
- Qiu, C.; Liu, K.; Zhang, S.; Gao, S.; Chen, W.; Li, D.; Huang, Y. Bisdemethoxycurcumin Inhibits Hepatocellular Carcinoma Proliferation Through Akt Inactivation via CYLD-Mediated Deubiquitination. Drug Des Devel Ther. 2020, 14, 993–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Huo, C.; Xiao, Y.; Xu, R.; Liu, Y.; Jia, X.; Wang, X. Bisdemethoxycurcumin Protection of Cardiomyocyte Mainly Depends on Nrf2/HO-1 Activation Mediated by the PI3K/AKT Pathway. Chem. Res. Toxicol. 2019, 32, 1871–1879. [Google Scholar] [CrossRef]
- Fu, M.; Fu, S.; Ni, S.; Wang, D.; Hong, T. Inhibitory effects of bisdemethoxycurcumin on mast cell-mediated allergic diseases. Int. Immunopharmacol. 2018, 65, 182–189. [Google Scholar] [CrossRef]
- Mun, S.H.; Joung, D.K.; Kim, Y.S.; Kang, O.H.; Kim, S.B.; Seo, Y.S.; Kim, Y.C.; Lee, D.S.; Shin, D.W.; Kweon, K.T.; et al. Synergistic antibacterial effect of curcumin against methicillin-resistant Staphylococcus aureus. Phytomedicine 2013, 20, 714–718. [Google Scholar] [CrossRef]
- Ramezani, M.; Hatamipour, M.; Sahebkar, A. Promising Anti-Tumor Properties of Bisdemethoxycurcumin: A Naturally Occurring Curcumin Analogue. J. Cell Physiol. 2018, 233, 880–887. [Google Scholar] [CrossRef]
- Mun, S.H.; Kang, O.H.; Kong, R.; Zhou, T.; Kim, S.A.; Shin, D.W.; Kwon, D.Y. Punicalagin suppresses methicillin resistance of Staphylococcus aureus to oxacillin. J. Pharmacol. Sci. 2017, 137, 317–323. [Google Scholar] [CrossRef]
- Choi, J.G.; Kang, O.H.; Brice, O.O.; Lee, Y.S.; Chae, H.S.; Oh, Y.C.; Sohn, D.H.; Park, H.; Choi, H.G.; Kim, S.G.; et al. Antibacterial activity of Ecklonia cava against methicillin-resistant Staphylococcus aureus and Salmonella spp. Foodborne Pathog. Dis. 2010, 7, 435–441. [Google Scholar] [CrossRef]
- Irvin, R.T.; MacAlister, T.J.; Costerton, J.W. Tris (hydroxymethyl) aminomethane buffer modification of Escherichia coli outer membrane permeability. J. Bacteriol. 1981, 145, 1397–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzales, P.R.; Pesesky, M.W.; Bouley, R.; Ballard, A.; Biddy, B.A.; Suckow, M.A.; Wolter, W.R.; Schroeder, V.A.; Burnham, C.A.D.; Mobashery, S.; et al. Synergistic, Collaterally Sensitive β-lactam Combinations Suppress Resistance in MRSA. Nat. Chem. Biol. 2015, 11, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Quijada, N.M.; Hernández, M.; Oniciuc, E.A.; Eiros, J.M.; Fernández-Natal, I.; Wagner, M.; Rodríguez-Lázaro, D. Oxacillin-susceptible mecA-positive Staphylococcus aureus associated with processed food in Europe. Food Microbiol. 2019, 82, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Dintcheva, N.T.; Infurna, G.; Baiamonte, M.; D’Anna, F. Natural Compounds as Sustainable Additives for Biopolymers. Polymers (Basel) 2020, 12, 732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, T.; Reker, D.; Schneider, P.; Schneider, G. Counting on natural products for drug design. Nat. Chem. 2016, 8, 531–541. [Google Scholar] [CrossRef]
- Joung, D.K.; Choi, S.H.; Kang, O.H.; Kim, S.B.; Mun, S.H.; Seo, Y.S.; Kang, D.H.; Gong, R.; Shin, D.W.; Kim, Y.C.; et al. Synergistic effects of oxyresveratrol in conjunction with antibiotics against methicillin-resistant Staphylococcus aureus. Mol. Med. Rep. 2015, 12, 663–667. [Google Scholar] [CrossRef]
- Mun, S.H.; Kang, O.H.; Joung, D.K.; Kim, S.B.; Choi, J.G.; Shin, D.W.; Kwon, D.Y. In vitro anti-MRSA activity of carvone with gentamicin. Exp. Ther. Med. 2019, 7, 891–896. [Google Scholar] [CrossRef]
- Zhou, T.; Li, Z.; Kang, O.H.; Mun, S.H.; Seo, Y.S.; Kong, R.; Shin, D.W.; Liu, X.Q.; Kwon, D.Y. Antimicrobial activity and synergism of ursolic acid 3-O-α-L-arabinopyranoside with oxacillin against methicillin-resistant Staphylococcus aureus. Int. J. Mol. Med. 2017, 40, 1285–1293. [Google Scholar] [CrossRef]
- Mun, S.H.; Kong, R.; Seo, Y.S.; Zhou, T.; Kang, O.H.; Shin, D.W.; Kwon, D.Y. Subinhibitory concentrations of punicalagin reduces expression of virulence-related exoproteins by Staphylococcus aureus. FEMS Microbiol. Lett. 2016, 363, e253. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MIC (μg/mL) | |||
|---|---|---|---|---|
| BDMC | GT | AMP | OXA | |
| ATCC 33591 | 7.8 | 3.9 | 62.5 | 250 |
| CCARM 3090 | 7.8 | 62.5 | 31.3 | 125 |
| CCARM 3091 | 7.8 | 250 | 62.5 | 1000 |
| CCARM 3095 | 15.6 | 125 | 31.3 | 250 |
| CCARM 3102 | 7.8 | 125 | 15.6 | 250 |
| DPS-1 | 7.8 | 125 | 62.5 | 250 |
| S. aureus Strains | Combination with GT | Combination with AMP | Combination with OXA | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Fold | FICI | Interpretation | Fold | FICI | Interpretation | Fold | FICI | Interpretation | |
| ATCC 33591 | 4 | 0.75 | partial synergy | 4 | 0.75 | partial synergy | 2 | 0.51 | partial synergy |
| CCARM 3090 | 8 | 0.2 | synergy | 4 | 0.75 | partial synergy | 4 | 0.75 | partial synergy |
| CCARM 3091 | 8 | 0.3 | synergy | 2 | 1 | additive effect | 4 | 0.75 | partial synergy |
| CCARM 3095 | 8 | 0.36 | synergy | 4 | 0.75 | partial synergy | 4 | 0.62 | partial synergy |
| CCARM 3102 | 4 | 0.5 | synergy | 8 | 0.62 | partial synergy | 4 | 0.62 | partial synergy |
| DPS-1 | 16 | 0.1 | synergy | 2 | 0.51 | partial synergy | 2 | 1 | additive effect |
| Primer | Sequence (5′-3′) |
|---|---|
| 16S RNA | F:ACTCCTACGGGAGGCAGCAG |
| R:ATTACCGCGGCTGCTGG | |
| mecA | F:CAATGCCAAAATCTCAGGTAAAGTG |
| R:AACCATCGTTACGGATTGCTTC | |
| mecR1 | F:GTGCTCGTCTCCACGTTAATTCCA |
| R:GACTAACCGAAGAAGTCGTGTCAG | |
| blaR1 | F:CACTATTCTCAGAATGACTTGGT |
| R:TGCATAATTCTCTTACTGTCATG | |
| blaZ | F:GCTTTAAAAGAACTTATTGAGGCTTC |
| R:CCACCGATYTCKTTTATAATTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Kim, M.-C.; Kang, O.-H.; Kwon, D.-Y. The Mechanism of Bisdemethoxycurcumin Enhances Conventional Antibiotics against Methicillin-Resistant Staphylococcus aureus. Int. J. Mol. Sci. 2020, 21, 7945. https://doi.org/10.3390/ijms21217945
Wang S, Kim M-C, Kang O-H, Kwon D-Y. The Mechanism of Bisdemethoxycurcumin Enhances Conventional Antibiotics against Methicillin-Resistant Staphylococcus aureus. International Journal of Molecular Sciences. 2020; 21(21):7945. https://doi.org/10.3390/ijms21217945
Chicago/Turabian StyleWang, Shu, Min-Chul Kim, Ok-Hwa Kang, and Dong-Yeul Kwon. 2020. "The Mechanism of Bisdemethoxycurcumin Enhances Conventional Antibiotics against Methicillin-Resistant Staphylococcus aureus" International Journal of Molecular Sciences 21, no. 21: 7945. https://doi.org/10.3390/ijms21217945