Copper(II) Binding by the Earliest Vertebrate Gonadotropin-Releasing Hormone, the Type II Isoform, Suggests an Ancient Role for the Metal

Abstract
:
1. Introduction
2. Results and Discussion
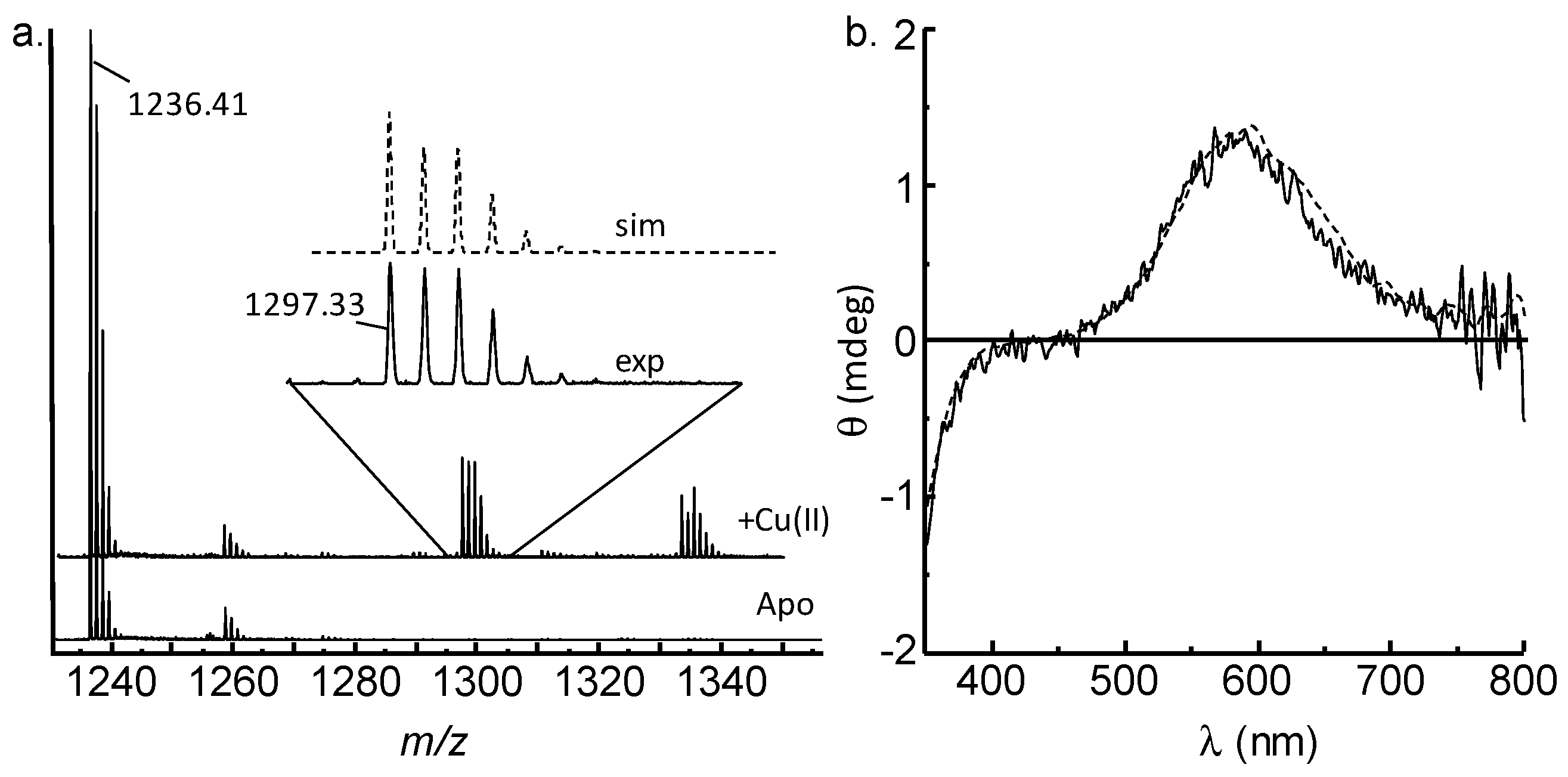
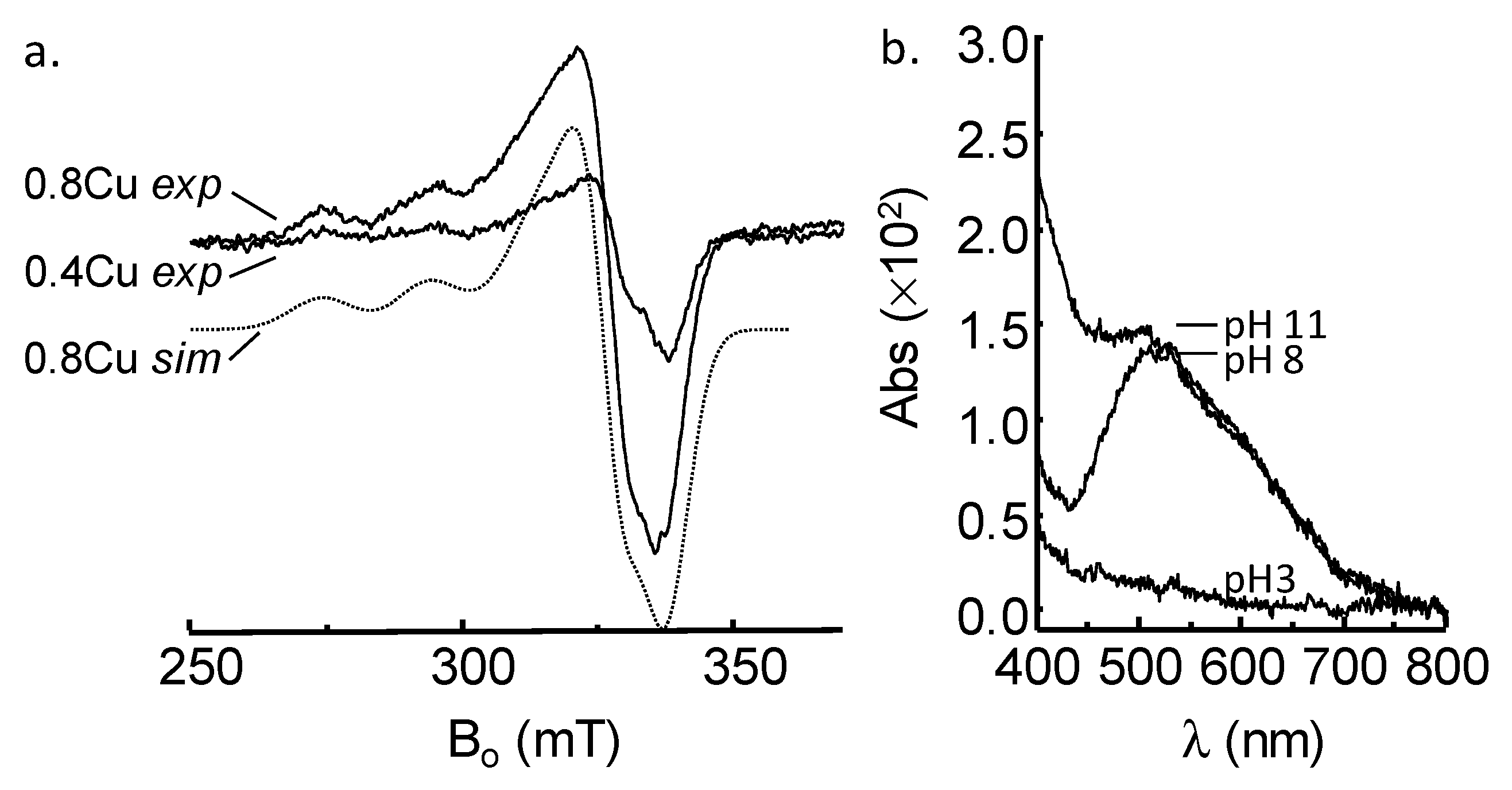
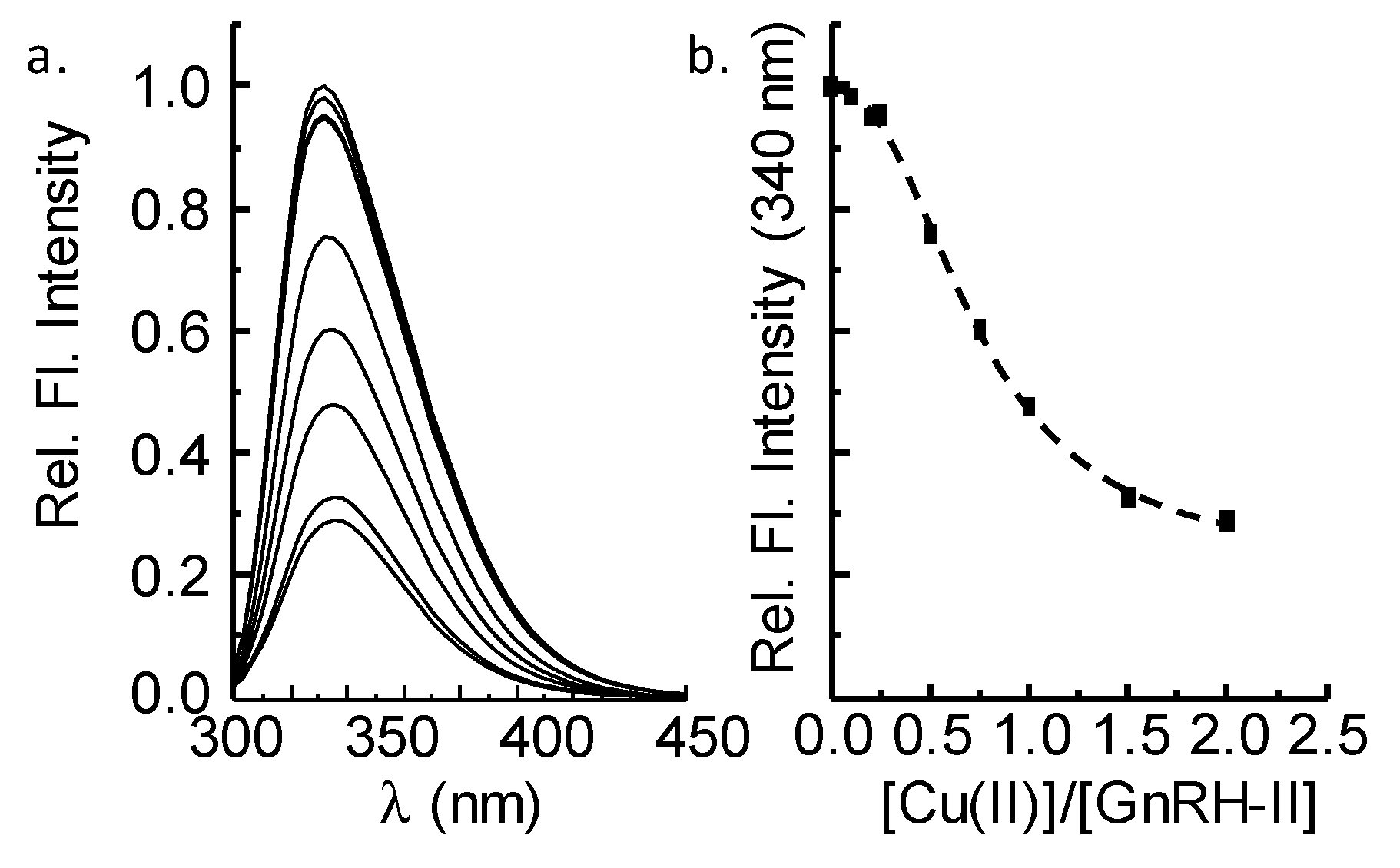
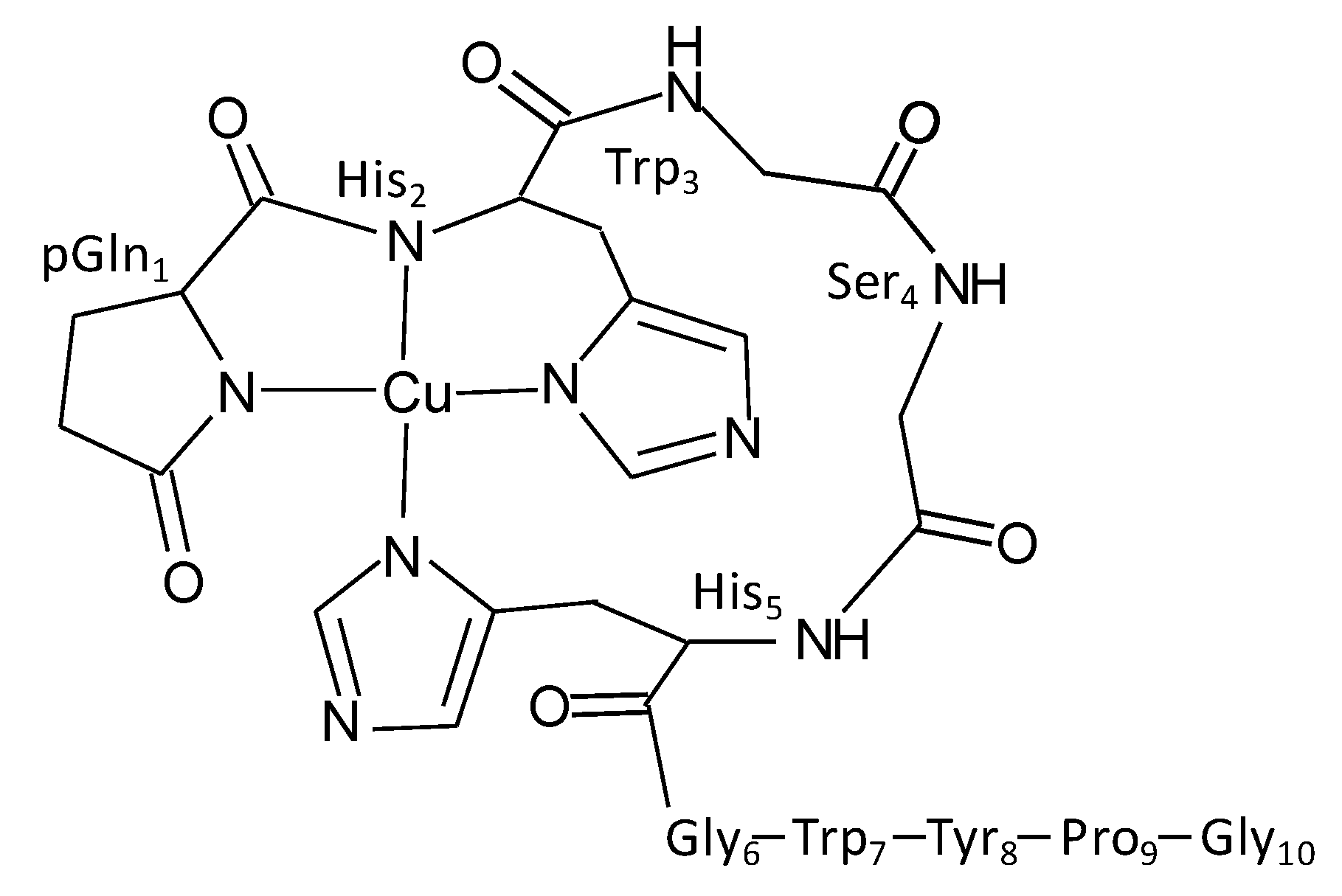
2.1. GnRH-II Binds Copper with a 1:1 Stoichiometry in a Four-Nitrogen Environment
2.2. Histidine Amino-Acids Are Highly Conserved in Vertebrate GnRH-II
3. Materials and Methods
3.1. Materials
3.2. Mass Spectroscopy (MS)
3.3. Electronic Spectroscopy
3.4. Fluorescence Spectroscopy
3.5. Electron Paramagnetic Resonance (EPR)
3.6. Circular Dichroism (CD) Spectroscopy
3.7. Phylogenetic Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sharpe, R.M.; Fraser, H.M. HCG stimulation of testicular LHRH-like activity. Nature 1980, 287, 642–643. [Google Scholar] [CrossRef]
- Moore, A.M.; Coolen, L.M.; Porter, D.T.; Goodman, R.L.; Lehman, M.N. KNDy Cells Revisited. Endocrinology 2018, 159, 3219–3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neill, J.D.; Musgrove, L.C.; Duck, L.W. Newly recognized GnRH receptors: Function and relative role. Trends Endocrinol. Metab. 2004, 15, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Urbanski, H.F. Differential roles of GnRH-I and GnRH-II neurons in the control of the primate reproductive axis. Front. Endocrinol. (Lausanne) 2012, 3, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desaulniers, A.T.; Cederberg, R.A.; Mills, G.A.; Ford, J.J.; Lents, C.A.; White, B.R. LH-Independent Testosterone Secretion Is Mediated by the Interaction Between GNRH2 and Its Receptor Within Porcine Testes. Biol. Reprod. 2015, 93, 45. [Google Scholar] [CrossRef]
- Zhao, J.; Dong, X.; Hu, X.; Long, Z.; Wang, L.; Liu, Q.; Sun, B.; Wang, Q.; Wu, Q.; Li, L. Zinc levels in seminal plasma and their correlation with male infertility: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 22386. [Google Scholar] [CrossRef]
- Duncan, F.E.; Que, E.L.; Zhang, N.; Feinberg, E.C.; O’Halloran, T.V.; Woodruff, T.K. The zinc spark is an inorganic signature of human egg activation. Sci. Rep. 2016, 6, 24737. [Google Scholar] [CrossRef] [Green Version]
- Fevold, H.L.; Hisaw, F.L.; Greep, R. Augmentation of the gonad stimulating action of pituitary extracts by inorganic substances, particularly copper salts. Am. J. Physiol. 1936, 117, 68–74. [Google Scholar] [CrossRef]
- Suzuki, M.; Hiroi, M.; Sugita, S. Ovulation in rabbits following intravenous and intracerebral administration of copper sulphate. Acta Endocrinol. (Copenh) 1965, 50, 512–516. [Google Scholar] [CrossRef]
- Hazum, E. Copper and thiol regulation of gonadotropin releasing hormone binding and luteinizing hormone release. Biochem. Biophys. Res. Commun. 1983, 112, 306–312. [Google Scholar] [CrossRef]
- Schvartz, I.; Hazum, E. Copper induces luteinizing hormone release and desensitization of pituitary gonadotropes. Biochem. Biophys. Res. Commun. 1986, 136, 417–425. [Google Scholar] [CrossRef]
- Barnea, A.; Colombani-Vidal, M.; Cho, G.; Hartter, D.E. Evidence for synergism between copper and prostaglandin E2 in stimulating the release of gonadotropin-releasing hormone from median eminence explants: Na+/Cl- requirements. Mol. Cell. Endocrinol. 1988, 56, 11–19. [Google Scholar] [CrossRef]
- Nakamura, K.; Kodaka, M.; El-Mehasseb, I.M.; Gajewska, A.; Okuno, H.; Ochwanowska, E.; Witek, B.; Kozlowski, H.; Kochman, K. Further structural analysis of GnRH complexes with metal ions. Neuro Endocrinol. Lett. 2005, 26, 247–252. [Google Scholar] [PubMed]
- Gajewska, A.; Zielinska-Gorska, M.; Wolinska-Witort, E.; Siawrys, G.; Baran, M.; Kotarba, G.; Biernacka, K. Intracellular mechanisms involved in copper-gonadotropin-releasing hormone (Cu-GnRH) complex-induced cAMP/PKA signaling in female rat anterior pituitary cells in vitro. Brain Res. Bull. 2016, 120, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Herman, A.; Kozlowski, H.; Czauderna, M.; Kochman, K.; Kulon, K.; Gajewska, A. Gonadoliberin (GnRH) and its copper complex (Cu-GnRH) enzymatic degradation in hypothalamic and pituitary tissue in vitro. J. Physiol. Pharmacol. 2012, 63, 69–75. [Google Scholar]
- Russino, D.; McDonald, E.; Hejazi, L.; Hanson, G.R.; Jones, C.E. The tachykinin peptide neurokinin B binds copper forming an unusual [CuII(NKB)2] complex and inhibits copper uptake into 1321N1 astrocytoma cells. ACS Chem. Neurosci. 2013, 4, 1371–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayawardena, B.M.; Jones, M.R.; Hong, Y.; Jones, C.E. Copper ions trigger disassembly of neurokinin B functional amyloid and inhibit de novo assembly. J. Struct. Biol. 2019, 208, 107394. [Google Scholar] [CrossRef] [PubMed]
- Peacey, L.; Elphick, M.R.; Jones, C.E. Roles of copper in neurokinin B and gonadotropin-releasing hormone structure and function and the endocrinology of reproduction. Gen. Comp. Endocrinol. 2020, 287, 113342. [Google Scholar] [CrossRef]
- Roy, D.; Dey, S.; Majumder, G.C.; Bhattacharyya, D. Copper: A biphasic regulator of caprine sperm forward progression. Syst. Biol. Reprod. Med. 2014, 60, 52–57. [Google Scholar] [CrossRef]
- Kuo, Y.M.; Gybina, A.A.; Pyatskowit, J.W.; Gitschier, J.; Prohaska, J.R. Copper transport protein (Ctr1) levels in mice are tissue specific and dependent on copper status. J. Nutr. 2006, 136, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Ogorek, M.; Lenartowicz, M.; Starzynski, R.; Jonczy, A.; Staron, R.; Doniec, A.; Krzeptowski, W.; Bednarz, A.; Pierzchala, O.; Lipinski, P.; et al. Atp7a and Atp7b regulate copper homeostasis in developing male germ cells in mice. Metallomics 2017, 9, 1288–1303. [Google Scholar] [CrossRef] [PubMed]
- Steiger, D.; Fetchko, M.; Vardanyan, A.; Atanesyan, L.; Steiner, K.; Turski, M.L.; Thiele, D.J.; Georgiev, O.; Schaffner, W. The Drosophila copper transporter Ctr1C functions in male fertility. J. Biol. Chem. 2010, 285, 17089–17097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowal, M.; Lenartowicz, M.; Pecio, A.; Golas, A.; Blaszkiewicz, T.; Styrna, J. Copper metabolism disorders affect testes structure and gamete quality in male mice. Syst. Biol. Reprod. Med. 2010, 56, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Lettieri, G.; D’Agostino, G.; Mele, E.; Cardito, C.; Esposito, R.; Cimmino, A.; Giarra, A.; Trifuoggi, M.; Raimondo, S.; Notari, T.; et al. Discovery of the Involvement in DNA Oxidative Damage of Human Sperm Nuclear Basic Proteins of Healthy Young Men Living in Polluted Areas. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Rasheed, O.F. Association between seminal plasma copper and magnesium levels with oxidative stress in iraqi infertile men. Oman Med. J. 2010, 25, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Zandawala, M.; Tian, S.; Elphick, M.R. The evolution and nomenclature of GnRH-type and corazonin-type neuropeptide signaling systems. Gen. Comp. Endocrinol. 2018, 264, 64–77. [Google Scholar] [CrossRef]
- Roch, G.J.; Busby, E.R.; Sherwood, N.M. GnRH receptors and peptides: Skating backward. Gen. Comp. Endocrinol. 2014, 209, 118–134. [Google Scholar] [CrossRef]
- Tran, K.K.; Jayawardena, B.M.; Elphick, M.R.; Jones, C.E. A gonadotropin-releasing hormone type neuropeptide with a high affinity binding site for copper(ii) and nickel(ii). Metallomics 2019, 11, 404–414. [Google Scholar] [CrossRef]
- Tsangaris, J.M.; Martin, R.B. Visible circular dichroism of copper(II) complexes of amino acids and peptides. J. Am. Chem. Soc. 1970, 92, 4255–4260. [Google Scholar] [CrossRef]
- Stanyon, H.F.; Cong, X.; Chen, Y.; Shahidullah, N.; Rossetti, G.; Dreyer, J.; Papamokos, G.; Carloni, P.; Viles, J.H. Developing predictive rules for coordination geometry from visible circular dichroism of copper(II) and nickel(II) ions in histidine and amide main-chain complexes. FEBS J. 2014, 281, 3945–3954. [Google Scholar] [CrossRef] [Green Version]
- La Mendola, D.; Farkas, D.; Bellia, F.; Magri, A.; Travaglia, A.; Hansson, O.; Rizzarelli, E. Probing the copper(II) binding features of angiogenin. Similarities and differences between a N-terminus peptide fragment and the recombinant human protein. Inorg. Chem. 2012, 51, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Gerega, K.; Kozlowski, H.; Masiukiewicz, E.; Pettit, L.D.; Pyburn, S.; Rzeszotarska, B. Metal complexes of luteinizing hormone-releasing hormone (LHRH). potentiometric and spectroscopic studies. J. Inorg. Biochem. 1988, 33, 11–18. [Google Scholar] [CrossRef]
- Gul, A.S.; Tran, K.K.; Jones, C.E. Neurokinin B and serum albumin limit copper binding to mammalian gonadotropin releasing hormone. Biochem. Biophys. Res. Commun. 2018, 497, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Peisach, J.; Blumberg, W.E. Structural implications derived from the analysis of electron paramagnetic resonance spectra of natural and artificial copper proteins. Arch. Biochem. Biophys. 1974, 165, 691–708. [Google Scholar] [CrossRef]
- Bennett, B.; Kowalski, J.M. EPR Methods for Biological Cu(II): L-Band CW and NARS. Methods Enzymol. 2015, 563, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Hyde, J.S.; Bennett, B.; Walter, E.D.; Millhauser, G.L.; Sidabras, J.W.; Antholine, W.E. EPR of Cu2+ prion protein constructs at 2 GHz using the g(perpendicular) region to characterize nitrogen ligation. Biophys. J. 2009, 96, 3354–3362. [Google Scholar] [CrossRef] [Green Version]
- Syme, C.D.; Nadal, R.C.; Rigby, S.E.; Viles, J.H. Copper binding to the amyloid-beta (Abeta) peptide associated with Alzheimer’s disease: Folding, coordination geometry, pH dependence, stoichiometry, and affinity of Abeta-(1-28): Insights from a range of complementary spectroscopic techniques. J. Biol. Chem. 2004, 279, 18169–18177. [Google Scholar] [CrossRef] [Green Version]
- Jancso, A.; Paksi, Z.; Jakab, N.; Gyurcsik, B.; Rockenbauer, A.; Gajda, T. Solution chemical properties and catecholase-like activity of the copper(II)-Ac-His-His-Gly-His-OH system, a relevant functional model for copper containing oxidases. Dalton Trans. 2005, 3187–3194. [Google Scholar] [CrossRef]
- Millar, R.P. GnRHs and GnRH receptors. Anim. Reprod. Sci. 2005, 88, 5–28. [Google Scholar] [CrossRef]
- Sherwood, N.M.; Lovejoy, D.A.; Coe, I.R. Origin of mammalian gonadotropin-releasing hormones. Endocr. Rev. 1993, 14, 241–254. [Google Scholar] [CrossRef]
- Kavanaugh, S.I.; Nozaki, M.; Sower, S.A. Origins of gonadotropin-releasing hormone (GnRH) in vertebrates: Identification of a novel GnRH in a basal vertebrate, the sea lamprey. Endocrinology 2008, 149, 3860–3869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, B.A.; Tello, J.A.; Erchegyi, J.; Warby, C.; Hong, D.J.; Akinsanya, K.O.; Mackie, G.O.; Vale, W.; Rivier, J.E.; Sherwood, N.M. Six novel gonadotropin-releasing hormones are encoded as triplets on each of two genes in the protochordate, Ciona intestinalis. Endocrinology 2003, 144, 1907–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, J.F.; Reska-Skinner, S.M.; Prakash, M.O.; Fischer, W.H.; Park, M.; Rivier, J.E.; Craig, A.G.; Mackie, G.O.; Sherwood, N.M. Two new forms of gonadotropin-releasing hormone in a protochordate and the evolutionary implications. Proc. Natl. Acad. Sci. USA 1996, 93, 10461–10464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terakado, K. Induction of gamete release by gonadotropin-releasing hormone in a protochordate, Ciona intestinalis. Gen. Comp. Endocrinol. 2001, 124, 277–284. [Google Scholar] [CrossRef]
- Stoll, S.; Schweiger, A. EasySpin, a comprehensive software package for spectral simulation and analysis in EPR. J. Magn. Reson. 2006, 178, 42–55. [Google Scholar] [CrossRef]
- Froncisz, W.; Hyde, J.S. Broadening of strains of lines in the G-parallel region of Cu2+ electron-paramagnetic resonance spectra. J. Chem. Phys. 1980, 73, 3123–3131. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| gi (i = x, y, z) | 2.067, 2.067, 2.222 |
| Ai (63Cu) (×10−4 cm−1) | 10.0, 10.0, 196.8 |
| Linewidth (×10−4 cm−1) | 4.67 |
| g-straini | 0.052, 0.058, 0.086 |
| A-straini (×10−4 cm−1) | 11.7, 11.7, 3.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peacey, L.; Peacey, C.; Gutzinger, A.; Jones, C.E. Copper(II) Binding by the Earliest Vertebrate Gonadotropin-Releasing Hormone, the Type II Isoform, Suggests an Ancient Role for the Metal. Int. J. Mol. Sci. 2020, 21, 7900. https://doi.org/10.3390/ijms21217900
Peacey L, Peacey C, Gutzinger A, Jones CE. Copper(II) Binding by the Earliest Vertebrate Gonadotropin-Releasing Hormone, the Type II Isoform, Suggests an Ancient Role for the Metal. International Journal of Molecular Sciences. 2020; 21(21):7900. https://doi.org/10.3390/ijms21217900
Chicago/Turabian StylePeacey, Lorraine, Charlotte Peacey, Adele Gutzinger, and Christopher E. Jones. 2020. "Copper(II) Binding by the Earliest Vertebrate Gonadotropin-Releasing Hormone, the Type II Isoform, Suggests an Ancient Role for the Metal" International Journal of Molecular Sciences 21, no. 21: 7900. https://doi.org/10.3390/ijms21217900