Nitric Oxide Signal, Nitrogen Metabolism, and Water Balance Affected by γ-Aminobutyric Acid (GABA) in Relation to Enhanced Tolerance to Water Stress in Creeping Bentgrass

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
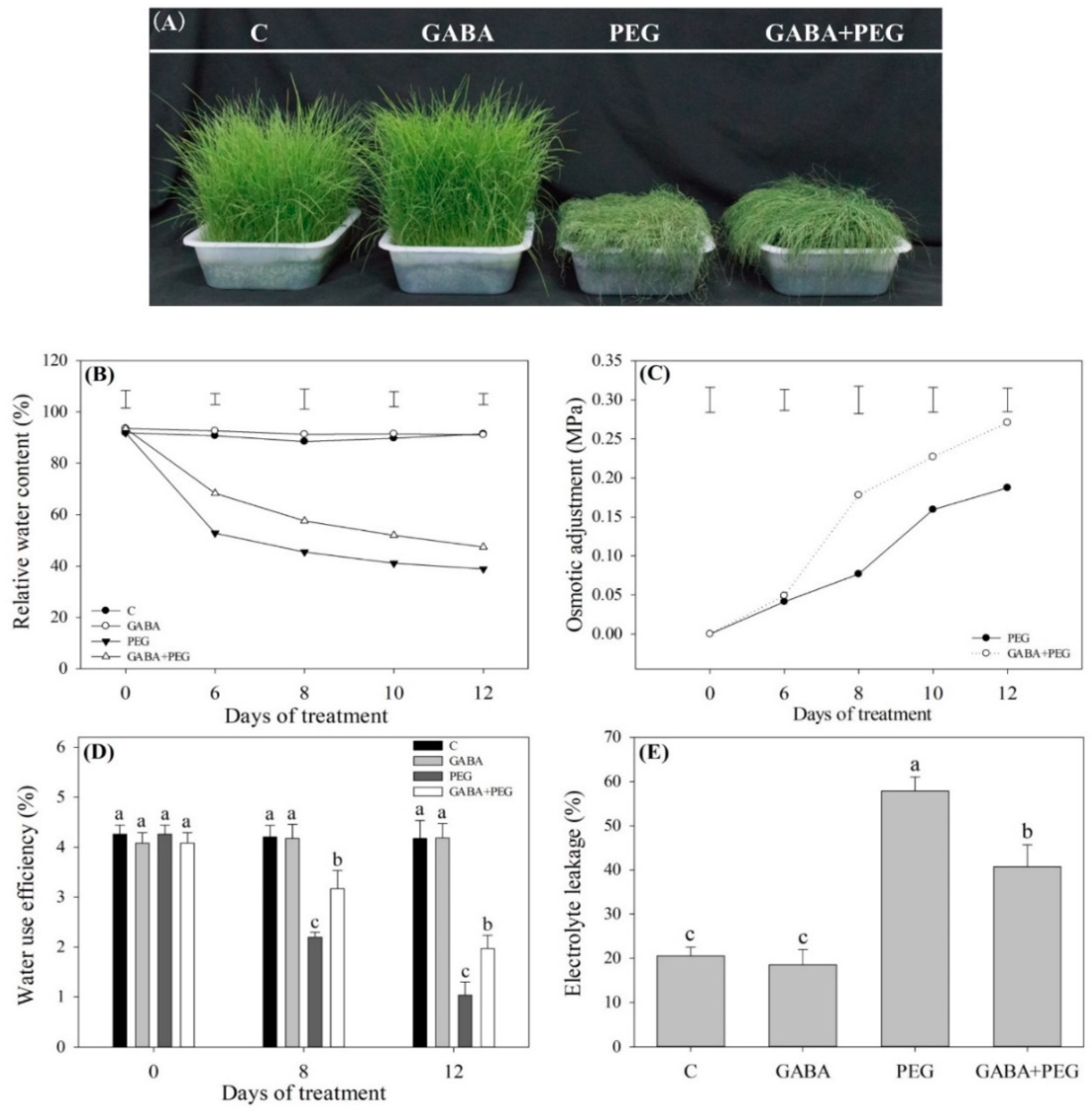
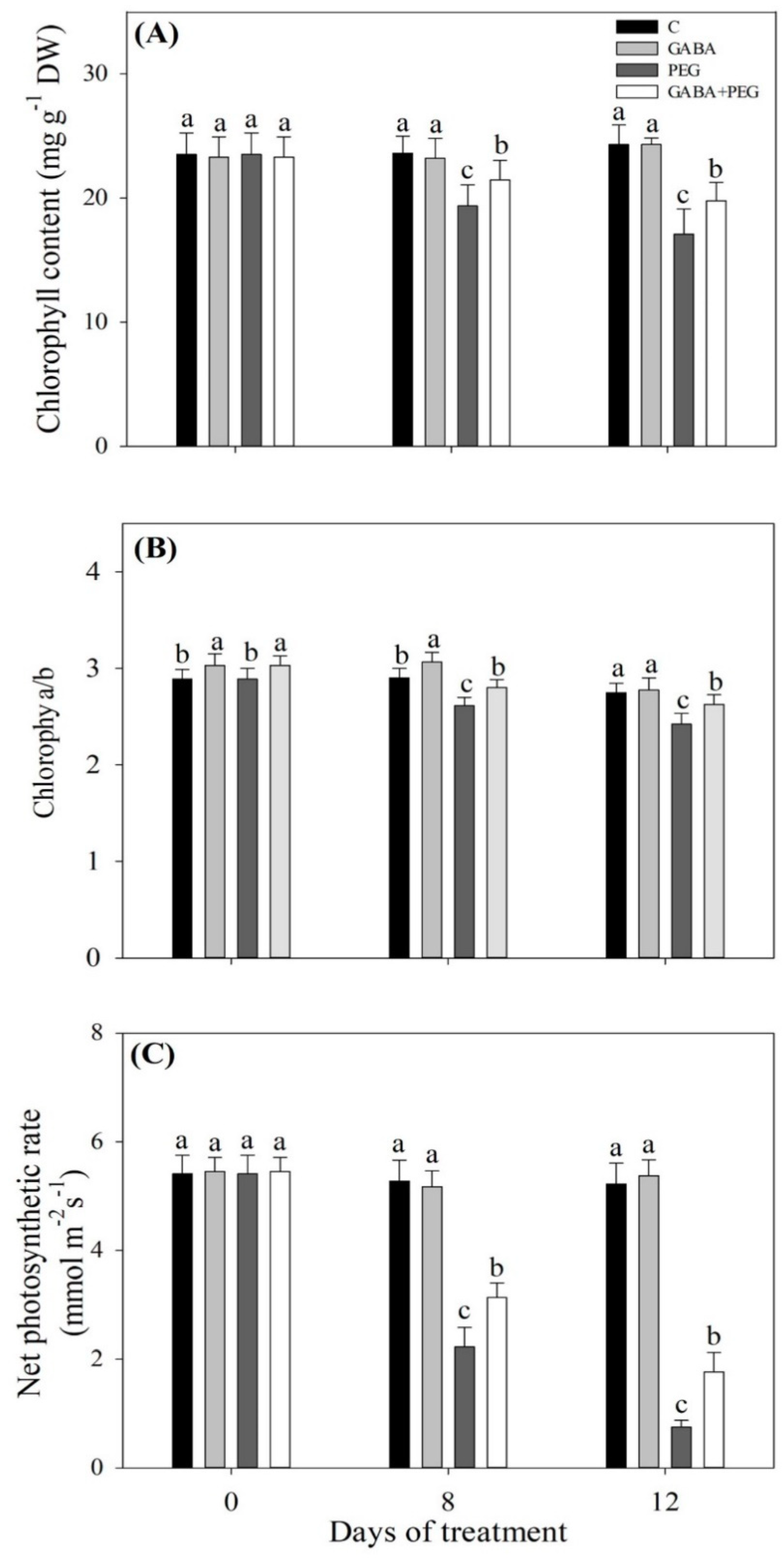
2.1. Cell Membrane Stability, Water Status, and Photosynthesis Affected by GABA
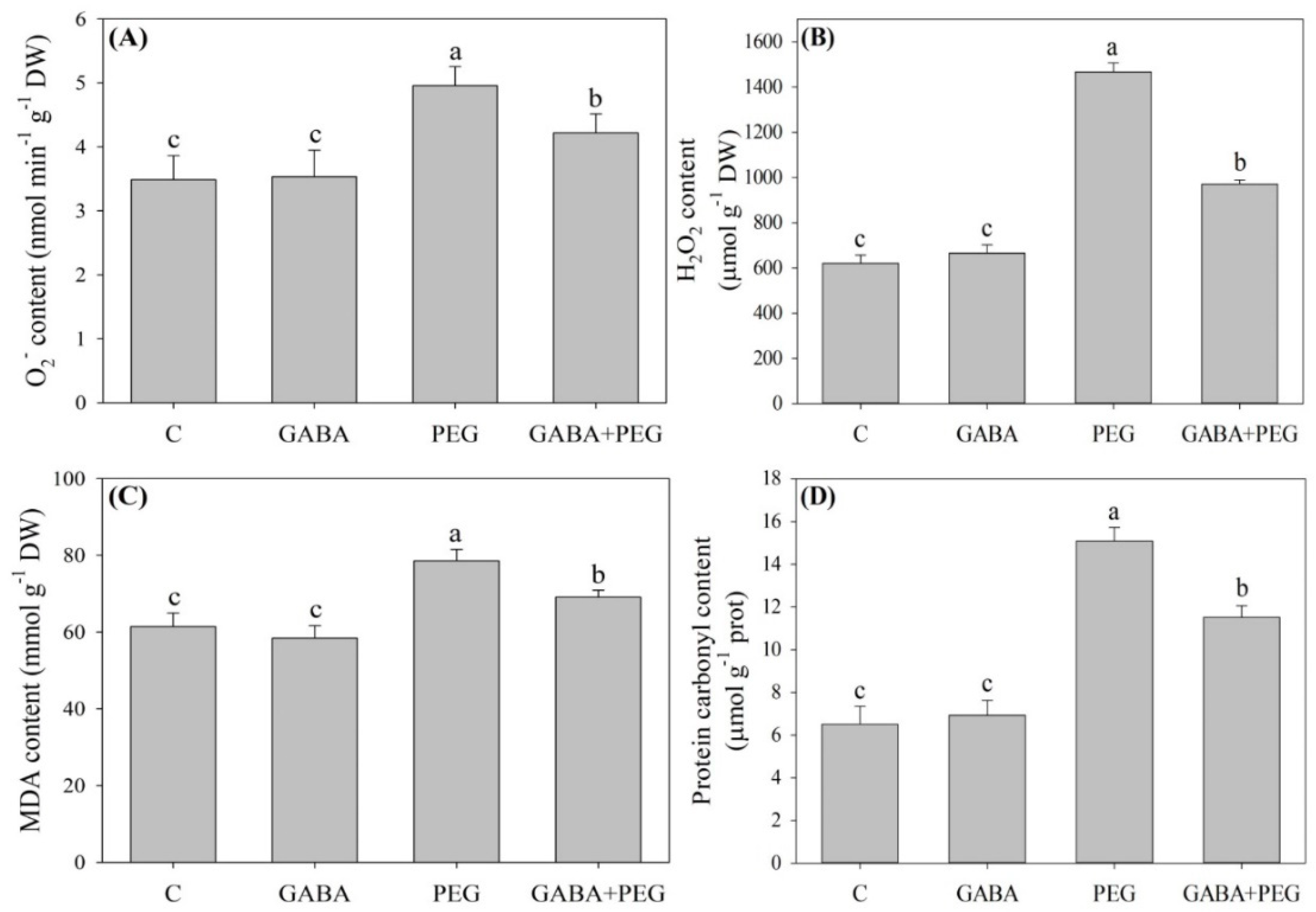
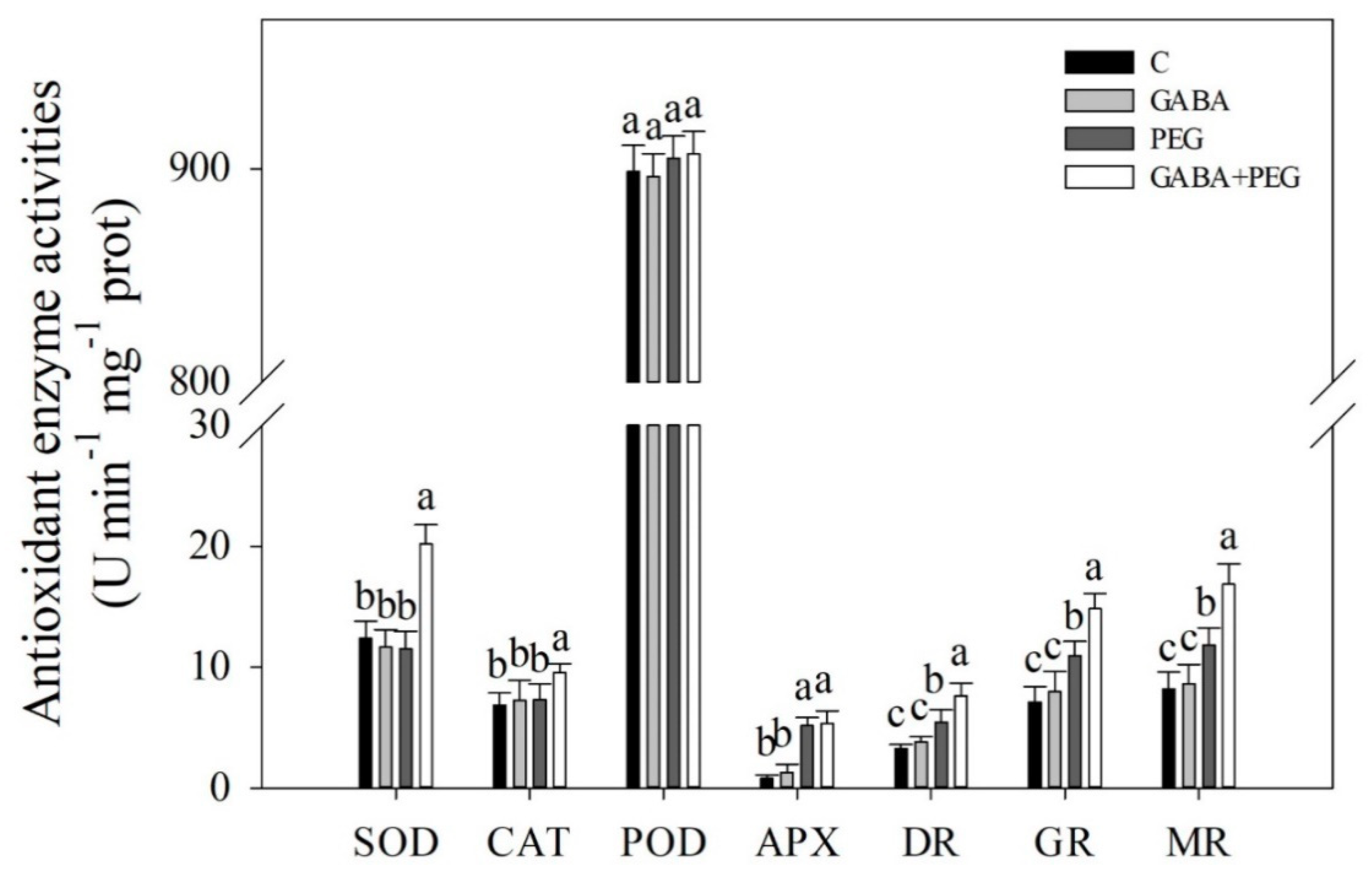
2.2. Oxidative Damage and Antioxidant Enzyme Activities Affected by GABA
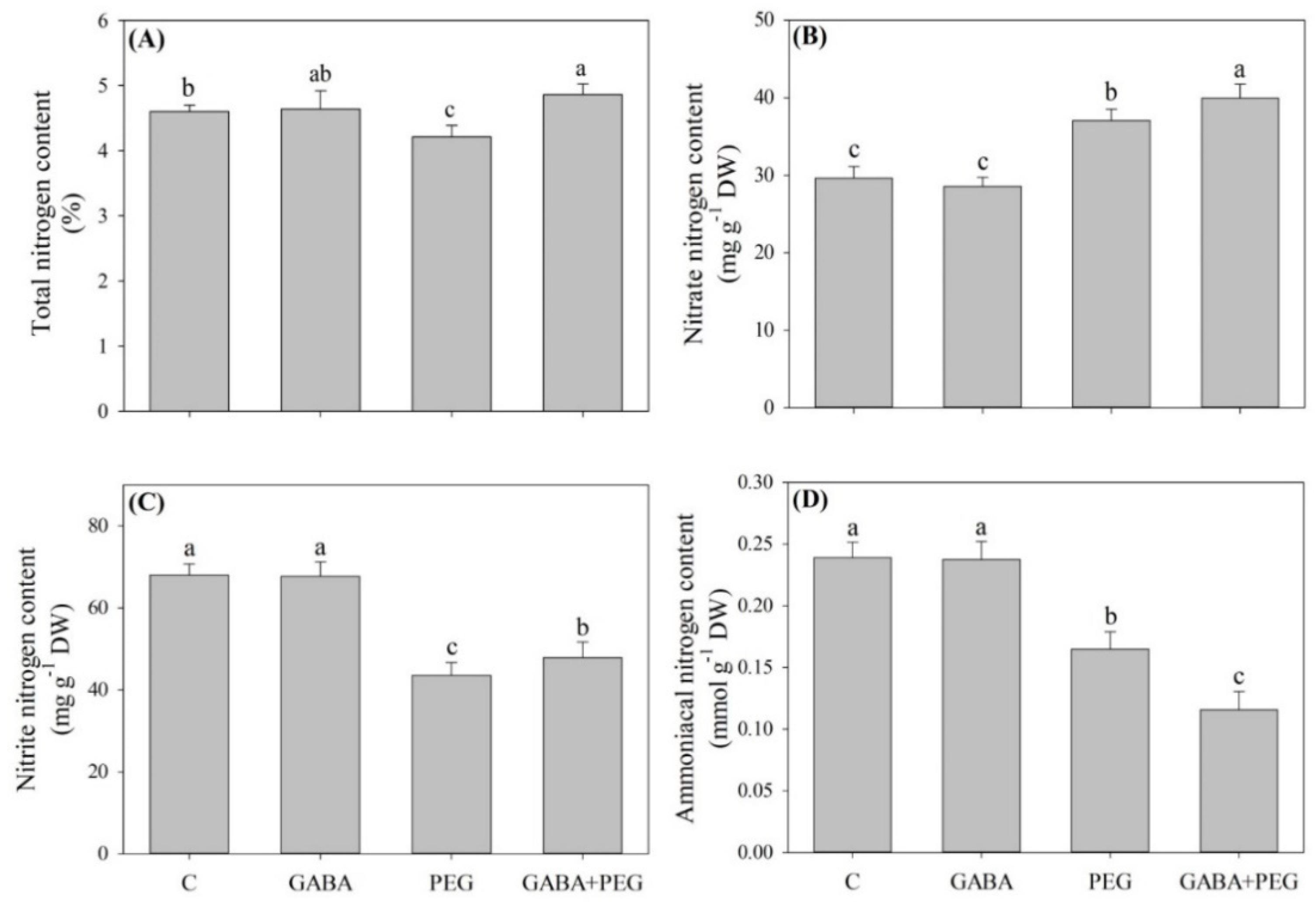
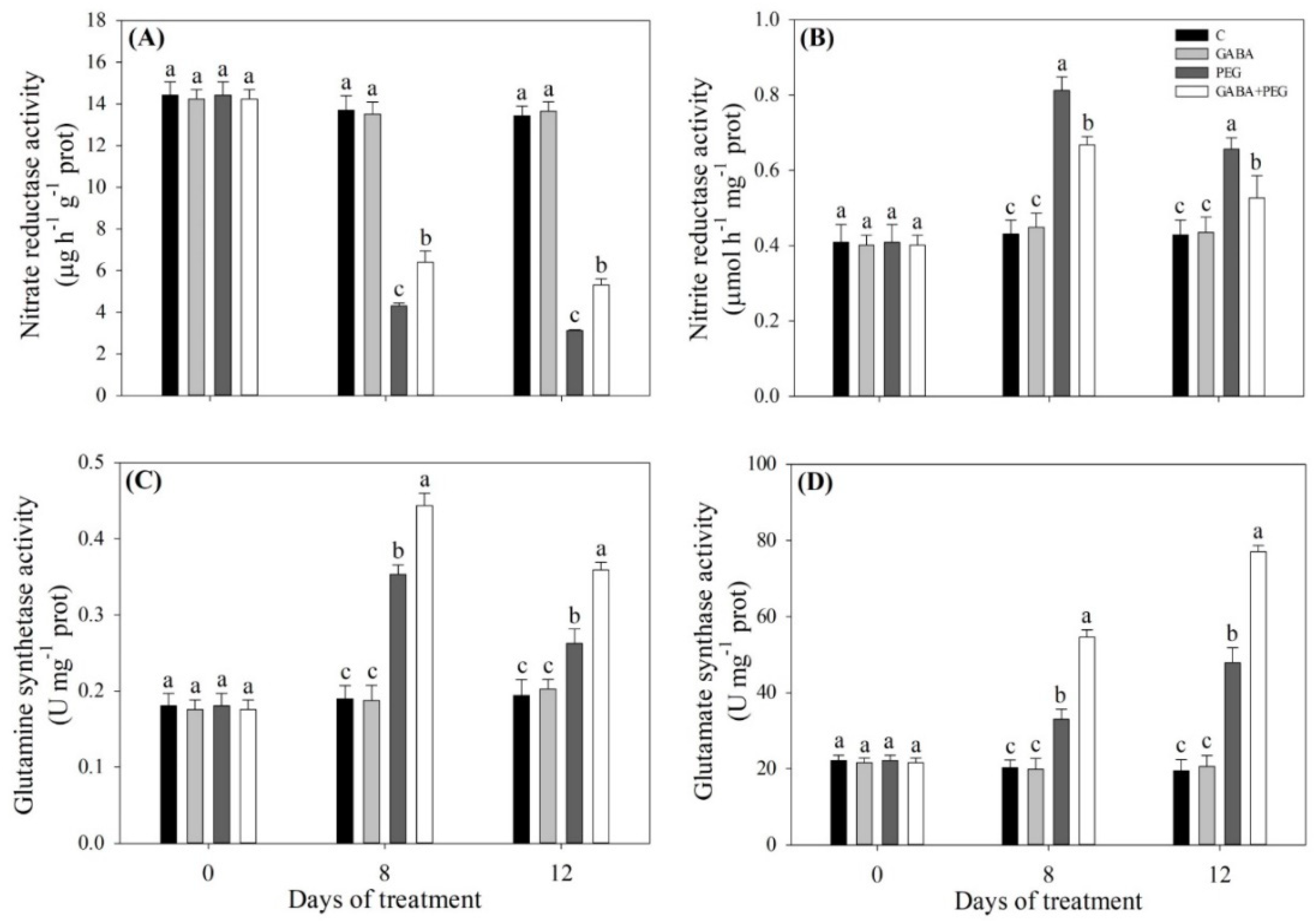
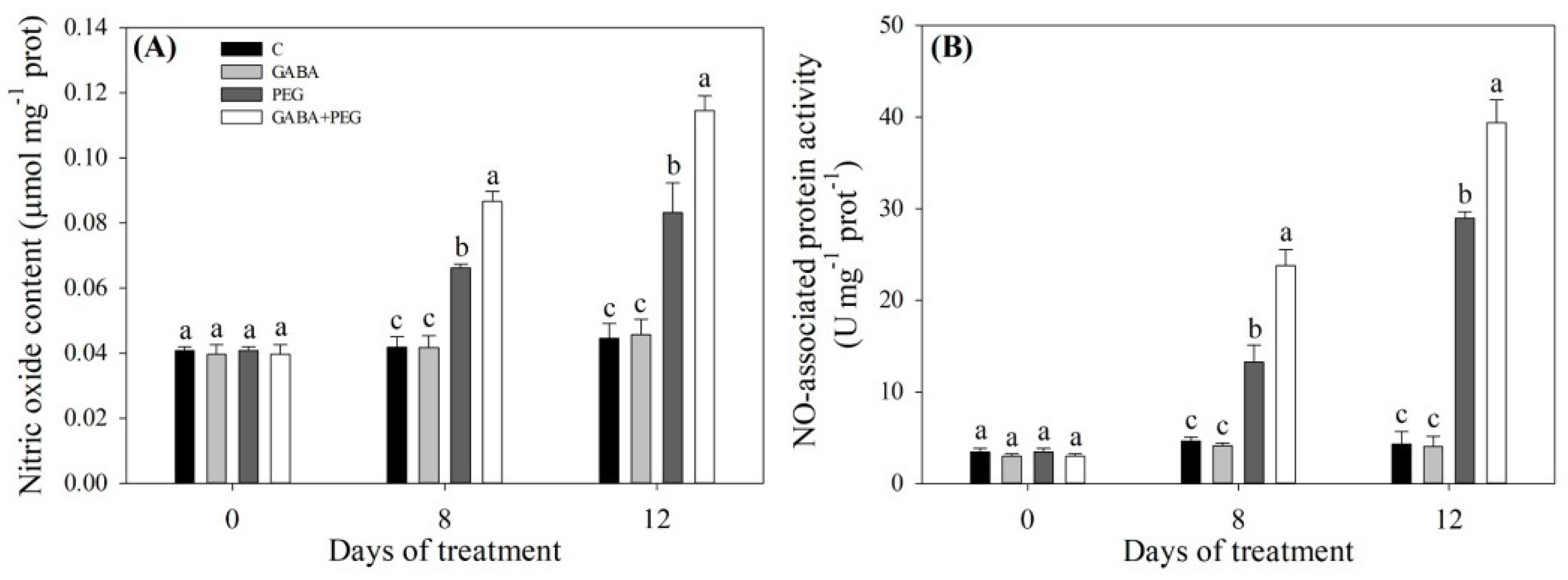
2.3. Nitrogen Metabolism and Nitric Oxide Affected by GABA
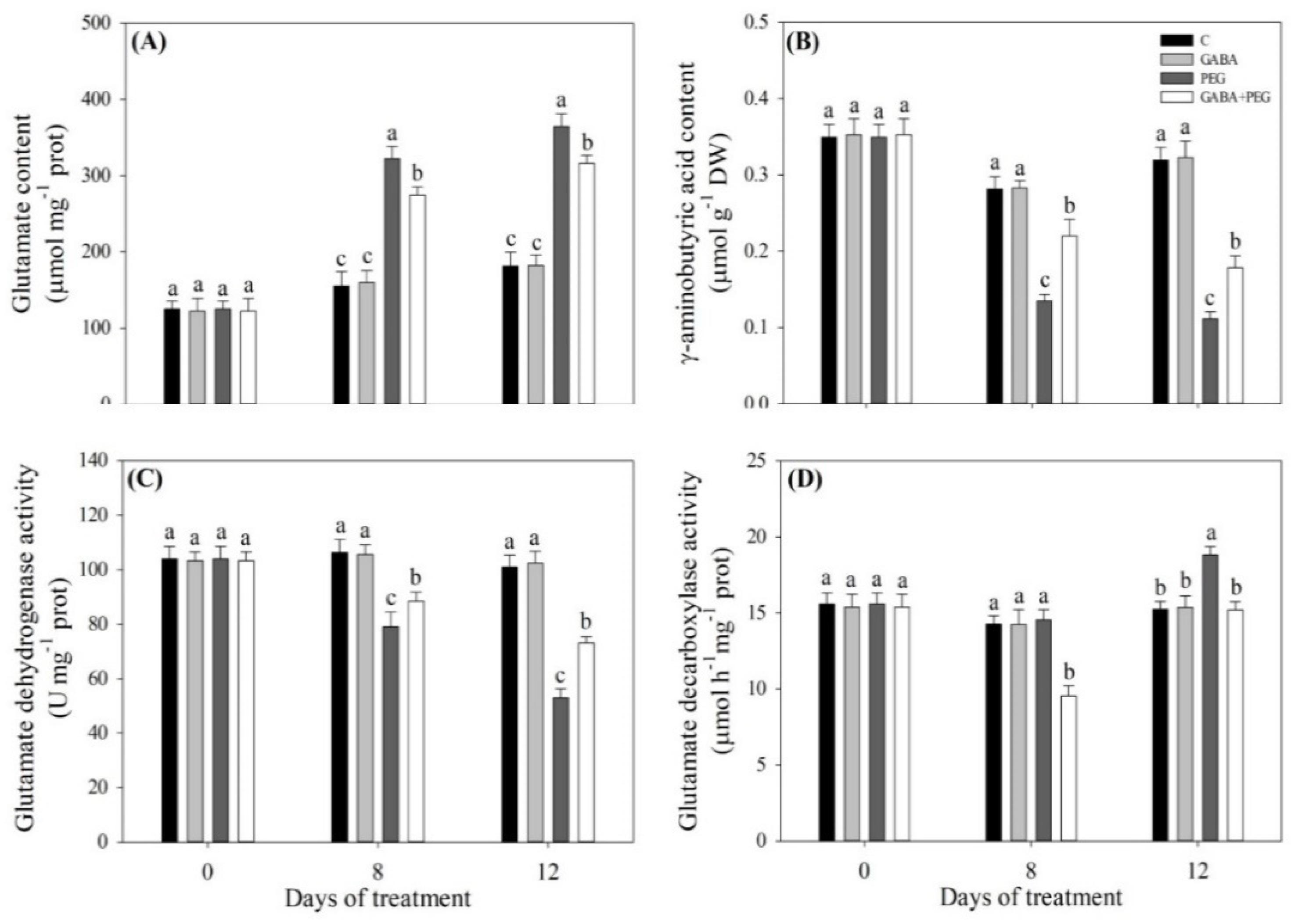
2.4. Endogenous Glutamic Acid and GABA Affected by GABA
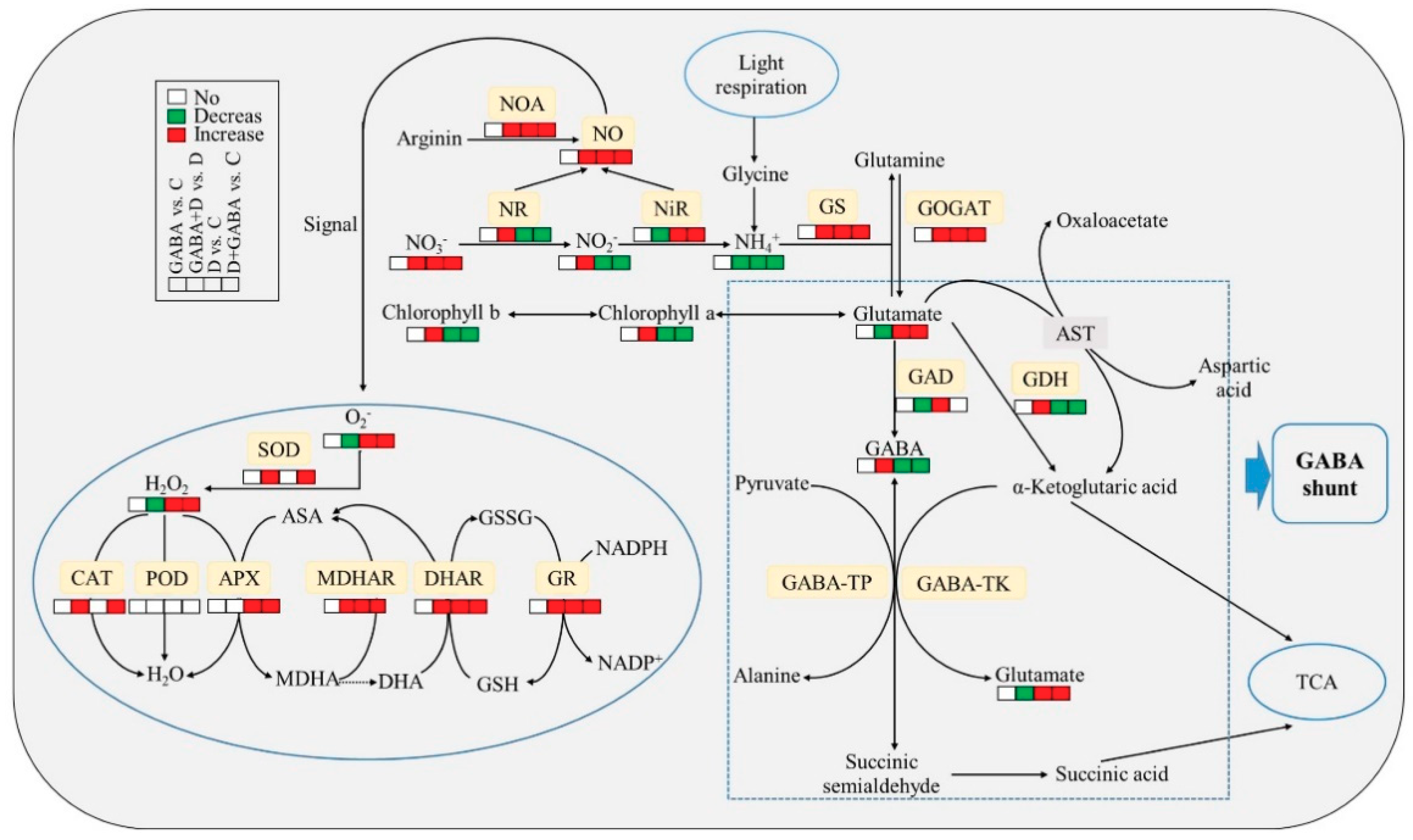
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Measurements of Cell Membrane Stability, Water Status and Photosynthesis
4.3. Determination of Oxidative Damage and Antioxidant Enzyme Activities
4.4. Determination of Nitrogen and γ-Aminobutyric Acid Metabolism
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| GABA | γ-aminobutyric acid |
| PEG-6000 | polyethylene glycol-6000 |
| SOD | superoxide dismutase |
| CAT | catalase |
| POD | peroxidase |
| APX | ascorbate peroxidase |
| DR | dehydroascorbate reductase |
| GR | glutathione reductase |
| MR | monodehydroascorbate reductase |
| RWC | relative water content |
| WUE | water use efficiency |
| Pn net | photosynthetic rate |
| EL | electrolyte leakage |
| OP | osmotic potential |
| OA | osmotic adjustment |
| Chl | total chlorophyll |
| TCA | tricarboxylic acid |
| NO | nitric oxide |
| NOA | NO-associated protein |
| NR | nitrate reductase |
| NiR | nitrite reductase |
| Glu | glutamate |
| GS | glutamine synthetase |
| GOGAT | glutamate synthase |
| GDH | glutamate dehydrogenase |
| GAD | glutamate decarboxylase |
| PGRs | plant growth regulators |
| ROS | reactive oxygen species |
| MDA | malondialdehyde |
| O2− | superoxide anion |
| H2O2 | hydrogen peroxide |
References
- Hu, L.X.; Wang, Z.L.; Huang, B.R. Diffusion limitations and metabolic factors associated with inhibition and recovery of photosynthesis from drought stress in a C perennial grass species. Physiol. Plant. 2010, 139, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, S.M.; Khan, M.Y.; Waqas, M.R. Arbuscular mycorrhizas and stress tolerance of drought stress in plants. In Arbuscular Mycorrhizas and Stress Tolerance of Plants; Wu, Q.S., Zuo, Y.N., Eds.; Springer: Gateway East, Singapore, 2017; pp. 25–41. [Google Scholar]
- Li, Z.; Zhou, H.; Peng, Y.; Zhang, X.; Ma, X.; Huang, L.; Yan, Y. Exogenously applied spermidine improves drought tolerance in creeping bentgrass associated with changes in antioxidant defense, endogenous polyamines and phytohormones. Plant Growth Regul. 2015, 76, 71–82. [Google Scholar] [CrossRef]
- Bian, X.J.; Merewitz, E.; Huang, B.R. Effects of trinexapac-ethyl on drought responses in creeping bentgrass associated with water use and osmotic adjustment. J. Amer. Soc. Hort. Sci. 2009, 134, 505–510. [Google Scholar] [CrossRef] [Green Version]
- Bouché, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Gilliham, M.; Tyerman, S.D. Linking metabolism to membrane signaling: The GABA–malate connection. Trends Plant Sci. 2016, 21, 136–138. [Google Scholar] [CrossRef] [Green Version]
- Serraj, R.; Shelp, B.J.; Sinclair, T.R. Accumulation of γ-aminobutyric acid in nodulated soybean in response to drought stress. Physiol. Plant. 1998, 102, 79–86. [Google Scholar] [CrossRef]
- Liu, T.; Liu, Z.; Li, Z.; Peng, Y.; Zhang, X.; Ma, X.; Huang, L.; Liu, W.; Nie, G.; He, L. Regulation of heat shock factor pathways by γ-aminobutyric acid (GABA) associated with thermotolerance of creeping bentgrass. Int. J. Mol. Sci. 2019, 20, 4713. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Zhao, L.; Yu, G. The dominant glutamic acid metabolic flux to produce γ-amino butyric acid over proline in Nicotiana tabacum leaves under water stress relates to its significant role in antioxidant activity. J. Integr. Plant Biol. 2011, 8, 608–618. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Tyerman, S.D.; Xu, B.; Bose, J.; Kaur, S.; Conn, V.; Domingos, P.; Ullah, S.; Wege, S.; Shabala, S.; et al. GABA signalling modulates plant growth by directly regulating the activity of plant-specific anion transporters. Nat. Commun. 2015, 6, 7879. [Google Scholar] [CrossRef] [Green Version]
- Allan, W.L.; Shelp, B.J. Fluctuations of γ-aminobutyrate, γ-hydroxybutyrate, and related amino acids in Arabidopsis leaves as a function of the light–dark cycle, leaf age, and N stress. Can. J. Bot. 2006, 84, 1339–1346. [Google Scholar] [CrossRef]
- Nayyar, H.; Kaur, R.; Kaur, S.; Singh, R. γ-Aminobutyric acid (GABA) imparts partial protection from heat stress injury to rice seedlings by improving leaf turgor and upregulating osmoprotectants and antioxidants. J. Plant Growth Regul. 2014, 33, 408–419. [Google Scholar] [CrossRef]
- Robertson, G.P.; Vitousek, P.M. Nitrogen in agriculture: Balancing the cost of an essential resource. Annu. Rev. Environ. Resour. 2009, 34, 97–125. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Huang, Y.; Chen, S.; Zou, J.; Zheng, X. Dependence of wheat and rice respiration on tissue nitrogen and the corresponding net carbon fixation efficiency under different rates of nitrogen application. Adv. Atmos. Sci. 2007, 24, 55–64. [Google Scholar] [CrossRef]
- Zhong, C.; Cao, X.; Hu, J.; Zhu, L.; Zhang, J.; Huang, J.; Jin, Q. Nitrogen metabolism in adaptation of photosynthesis to water stress in rice grown under different nitrogen levels. Front. Plant Sci. 2017, 8, 1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tawfik, A.A.; Kleinhenz, M.D.; Palta, J.P. Application of calcium and nitrogen for mitigating heat stress effects on potatoes. Amer. Potato. J. 1996, 73, 261–273. [Google Scholar] [CrossRef]
- Jespersen, D.; Huang, B. Proteins associated with heat-induced leaf senescence in creeping bentgrass as affected by foliar application of nitrogen, cytokinins, and an ethylene inhibitor. Proteomics. 2015, 4, 798–812. [Google Scholar] [CrossRef]
- Arasimowicz, M.; Floryszak-Wołyńska, J. Nitric oxide as a bioactive signalling molecule in plant stress responses. Plant Sci. 2007, 172, 876–887. [Google Scholar] [CrossRef]
- Rockel, P.; Kaiser, W.M. NO production in plants: Nitrate reductase versus nitric oxide synthase. In Progress in Botany; Esser, K., Beyschlag, W., Hellwig, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; Volume 63, pp. 246–257. [Google Scholar]
- Zhang, H.; Shen, W.B.; Xu, L.L. Effects of nitric oxide on the germination of wheat seeds and its reactive oxygen species metabolisms under osmotic stress. J. Bot. 2003, 45, 87–100. [Google Scholar]
- Uchida, A.; TJagendorf, A.; Hibino, T.; Takabe, T.; Takabe, T. Effects of hydrogen peroxide and nitric oxide on both salt and heat stress tolerance in rice. Plant. Sci. 2002, 163, 515–523. [Google Scholar] [CrossRef]
- Tian, X.; Lei, Y. Nitric oxide treatment alleviates drought stress in wheat seedlings. Biol. Plant. 2006, 50, 775–778. [Google Scholar] [CrossRef]
- Fu, J.J.; Chu, X.F.; Sun, Y.F.; Xu, Y.F.; Hu, T.M. Involvement of nitric oxide in 5-aminolevulinic acid-induced antioxidant defense in roots of Elymus nutans exposed to cold stress. Biol. Plant. 2016, 60, 585–594. [Google Scholar] [CrossRef]
- Han, Y.J.; Cho, K.C.; Hwang, O.J.; Choi, Y.S.; Shin, A.Y.; Hwang, I.; Kim, J. Overexpression of an Arabidopsis β-glucosidase gene enhances drought resistance with dwarf phenotype in creeping bentgrass. Plant Cell Rep. 2012, 31, 1677–1686. [Google Scholar] [CrossRef] [PubMed]
- Riseman, A.R.A.; Jensen, C.J.C.; Williams, M.W.M. Stomatal conductivity and osmotic adjustment during acclimation to multiple cycles of drought stress in potted miniature rose. J. Hort. Sci. Biotechnol. 2001, 76, 138–144. [Google Scholar] [CrossRef]
- Meng, X.; Wang, M.; Jiang, N.; Zhang, D.; Wang, L.; Liu, C. Regulation of both the reactive oxygen species level and antioxidant enzyme activity in drought stressed rice organs by benzimidazolate based SOD1 mimics. J. Agr. Food Chem. 2012, 60, 11211–11221. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.H.; Xue, J.Q.; Pu, J.; Zhao, B.; Bu, L.D. Influence of drought stress on plant growth and photosynthetic traits in maize seedlings. Acta Agron. Sinica. 2011, 37, 521–528. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.C.; Campbell, R.W.; Paulsen, G.M. Effects of drought stress and succinic acid-2,2-dimethylhydrazide treatment on water relations and photosynthesis in pea seedlings. Crop Sci. 1974, 14, 279–282. [Google Scholar] [CrossRef]
- Dwyer, L.M.; Stewart, D.W.; Tollenaar, M. Analysis of maize leaf photosynthesis under drought stress. Can. J. Plant Sci. 1992, 72, 477–481. [Google Scholar] [CrossRef]
- Li, Z.; Yu, J.; Peng, Y. Metabolic pathways regulated by abscisic acid, salicylic acid, and γ-aminobutyric acid in association with improved drought tolerance in creeping bentgrass (Agrostis stolonifera). Physiol Plant. 2017, 159, 42–58. [Google Scholar] [CrossRef]
- Helena, M.; Carvalho, M. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar]
- Hernández, I.; Cela, J.; Alegre, L.; Munné-Bosch, S. Antioxidant defenses against drought stress. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 823, pp. 231–258. [Google Scholar]
- Fedorova, M.; Bollineni, R.C.; Hoffmann, R. Protein carbonylation as a major hallmark of oxidative damage: Update of analytical strategies. Mass. Spectrom. Rev. 2014, 33, 79–97. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Peng, Y.; Huang, B. Alteration of transcripts of stress-protective genes and transcriptional factors by γ-Aminobutyric acid (GABA) associated with improved heat and drought tolerance in creeping bentgrass (Agrostis stolonifera). Int. J. Mol. Sci. 2018, 19, 1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolas, B.; Fait, A.; Zik, M. The root-specific glutamate decarboxylase (GAD1) is essential for sustaining GABA levels in Arabidopsis. Plant Mol. Biol. 2004, 55, 315–325. [Google Scholar]
- Miyashita, Y.; Good, A.G. Contribution of the GABA shunt to hypoxia-induced alanine accumulation in roots of Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Esteban, R.; Ariz, I.; Cruz, C.; Moran, J.F. Mechanisms of ammonium toxicity and the quest for tolerance. Plant Sci. 2016, 248, 92–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, S.; Zhao, D.; Lü, D.; Zhang, Y.; Liu, G.; Ma, H. Effects of water stress on carbon and nitrogen metabolism of hanfu apple leaves. J. Jilin. Agr. 2010, 32, 402–406. [Google Scholar]
- Zhen, A.; Hu, X.H.; Ren, W.Q.; Su, C.J.; Jin, X.Q.; Sun, X.P. Effect of exogenous γ-aminobutyric acid on NO3--N assimilation in muskmelon under Ca(NO3)2 stress. Indian. J. Plant Physiol. 1984, 27, 3987–3995. [Google Scholar]
- Bernard, S.M.; Habash, D.Z. The importance of cytosolic glutamine synthetase in nitrogen assimilation and recycling. New Phytol. 2009, 182, 608–620. [Google Scholar] [CrossRef]
- Chen, J.W.; Liu, Q.; Rong, X.; Xie, G.; Peng, J.; Zhu, H. Effects of different cultivations on activities of nitrogen metabolisms key enzymes of fodder rice. J. Hunan. Agr. 2005, 31, 238–241. [Google Scholar]
- Cao, Z.Z.; Zhang, Q.F.; Wei, K.S.; Yang, W.L.; Liu, G.K.; Cheng, F. Response of some key enzyme activities involved in nitrogen metabolism to high temperature at filling stage and its relation to storage protein accumulation in rice grain. Acta Agron. Sinica. 2013, 38, 99–106. [Google Scholar] [CrossRef]
- Li, Z.; Yu, J.; Peng, Y.; Huang, B. Metabolic pathways regulated by γ-aminobutyric acid (GABA) contributing to heat tolerance in creeping bentgrass (Agrostis stolonifera). Sci. Rep. 2016, 6, 30338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedroso, M.C.; Magalhaes, J.R.; Durzan, D. Nitric oxide induces cell death in Taxus cells. Plant Sci. 2000, 157, 178–180. [Google Scholar] [CrossRef]
- Yamasaki, H.; Sakihama, Y. Simultaneous production of nitric oxide and peroxynitrite by plant nitrate reductase: In vitro evidence for the NR-dependent formation of active nitrogen species. FEBS Lett. 2000, 468, 89–92. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, H.; Sakihama, Y.; Takahashi, S. An alternative pathway for nitric oxide production in plants: New features of an old enzyme. Trends Plant Sci. 1999, 4, 128–129. [Google Scholar] [CrossRef]
- Stöhr, C.; Strube, F.; Marx, G.; Ullrich, W.R.; Rockel, P. A plasma membrane-bound enzyme of tobacco roots catalyses the formation of nitric oxide from nitrite. Planta 2001, 212, 835–841. [Google Scholar] [CrossRef]
- Astier, J.; Gross, I.; Durner, J. Nitric oxide production in plants: An update. J. Exp. Bot. 2018, 69, 3401–3411. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.; Wang, X.; Li, Z.; Zhang, Y.; Peng, Y.; Li, Y.; He, X.; Zhang, X.; Ma, X.; Huang, L.; et al. NO is involved in spermidine-induced drought tolerance in white clover via activation of antioxidant enzymes and genes. Protoplasma. 2016, 253, 1243–1254. [Google Scholar] [CrossRef]
- Fan, H.; Li, T.; Guan, L.; Li, Z.; Guo, N.; Cai, Y.; Lin, Y. Effects of exogenous nitric oxide on antioxidation and DNA methylation of Dendrobium huoshanense grown under drought stress. Plant Cell Tissue Organ Cult. 2012, 109, 307–314. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water culture method for growing plants without soil. Calif. Agr. 1950, 347, 357–359. [Google Scholar]
- Li, Z.; Peng, Y.; Huang, B. Physiological effects of γ-aminobutyric acid application on improving heat and drought tolerance in creeping bentgrass. J. Amer. Soc. Hort. Sci. 2016, 141, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop Sci. 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Osmotic adjustment and growth of barley genotypes under drought stress. Crop Sci. 1989, 29, 230–233. [Google Scholar] [CrossRef]
- Feng, Y.L.; Cao, K.F. Photosynthetic characteristics, dark respiration, and leaf mass per unit area in seedlings of four tropical tree species grown under three irradiances. Photosynthetica. 2004, 42, 431–437. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, G.K. Superoxide dismutases: I. occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Cakmak, I.; Strbac, D.; Marschner, H. Activities of hydrogen peroxide scavenging enzymes in germinating wheat seeds. J. Exp. Bot. 1993, 44, 127–132. [Google Scholar] [CrossRef]
- Bremner, J.M. Determination of nitrogen in soil by the Kjeldahl method. J. Agr. Sci. 1960, 55, 11–33. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Li, L.; Xie, X.; Cao, Z. Effects of sulfur and salicylic acid on nitrate accumulation and nitrogen metabolism in leaves of Chinese chive. J. Plant Nutr. Fert. 2013, 19, 1264–1271. [Google Scholar]
- Rathore, D.P.S.; Tarafder, P.K. A rapid spectrophotometric method for the determination of nitrite in water. J. Indian. Chem. Soc. 1989, 66, 185–187. [Google Scholar]
- Sahrawat, K.L.; Prasad, R. A rapid method for determination of nitrate, nitrite, and ammoniacal nitrogen in soils. Plant Siol. 1975, 42, 305–308. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Srivastava, H.S. Increase in glutamate synthase activity (NADH-dependent) in excised root and leaf of maize seedlings in response to acidic amino acids and amides. Physiol. Plant. 1987, 182, 497–500. [Google Scholar] [CrossRef]
- Kanamori, T.; Konishi, S.; Takahashi, E. Inducible formation of glutamate dehydrogenase in rice plant roots by the addition of ammonia to the media. Physiol. Plant. 1972, 26, 1–6. [Google Scholar] [CrossRef]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, M.; Li, Z.; Luo, L.; Cheng, B.; Zhang, Y.; Zeng, W.; Peng, Y. Nitric Oxide Signal, Nitrogen Metabolism, and Water Balance Affected by γ-Aminobutyric Acid (GABA) in Relation to Enhanced Tolerance to Water Stress in Creeping Bentgrass. Int. J. Mol. Sci. 2020, 21, 7460. https://doi.org/10.3390/ijms21207460
Tang M, Li Z, Luo L, Cheng B, Zhang Y, Zeng W, Peng Y. Nitric Oxide Signal, Nitrogen Metabolism, and Water Balance Affected by γ-Aminobutyric Acid (GABA) in Relation to Enhanced Tolerance to Water Stress in Creeping Bentgrass. International Journal of Molecular Sciences. 2020; 21(20):7460. https://doi.org/10.3390/ijms21207460
Chicago/Turabian StyleTang, Mingyan, Zhou Li, Ling Luo, Bizhen Cheng, Youzhi Zhang, Weihang Zeng, and Yan Peng. 2020. "Nitric Oxide Signal, Nitrogen Metabolism, and Water Balance Affected by γ-Aminobutyric Acid (GABA) in Relation to Enhanced Tolerance to Water Stress in Creeping Bentgrass" International Journal of Molecular Sciences 21, no. 20: 7460. https://doi.org/10.3390/ijms21207460