Application of the MSAP Technique to Evaluate Epigenetic Changes in Plant Conservation

 , , , and
, , , and
Abstract
:1. Epigenetic Variation in Ex Situ Plant Conservation: The Role of DNA Methylation Changes
2. Methylation Sensitive Amplified Polymorphism (MSAP) Technique among Other Techniques to Detect DNA Methylation Changes
3. Seed Conservation
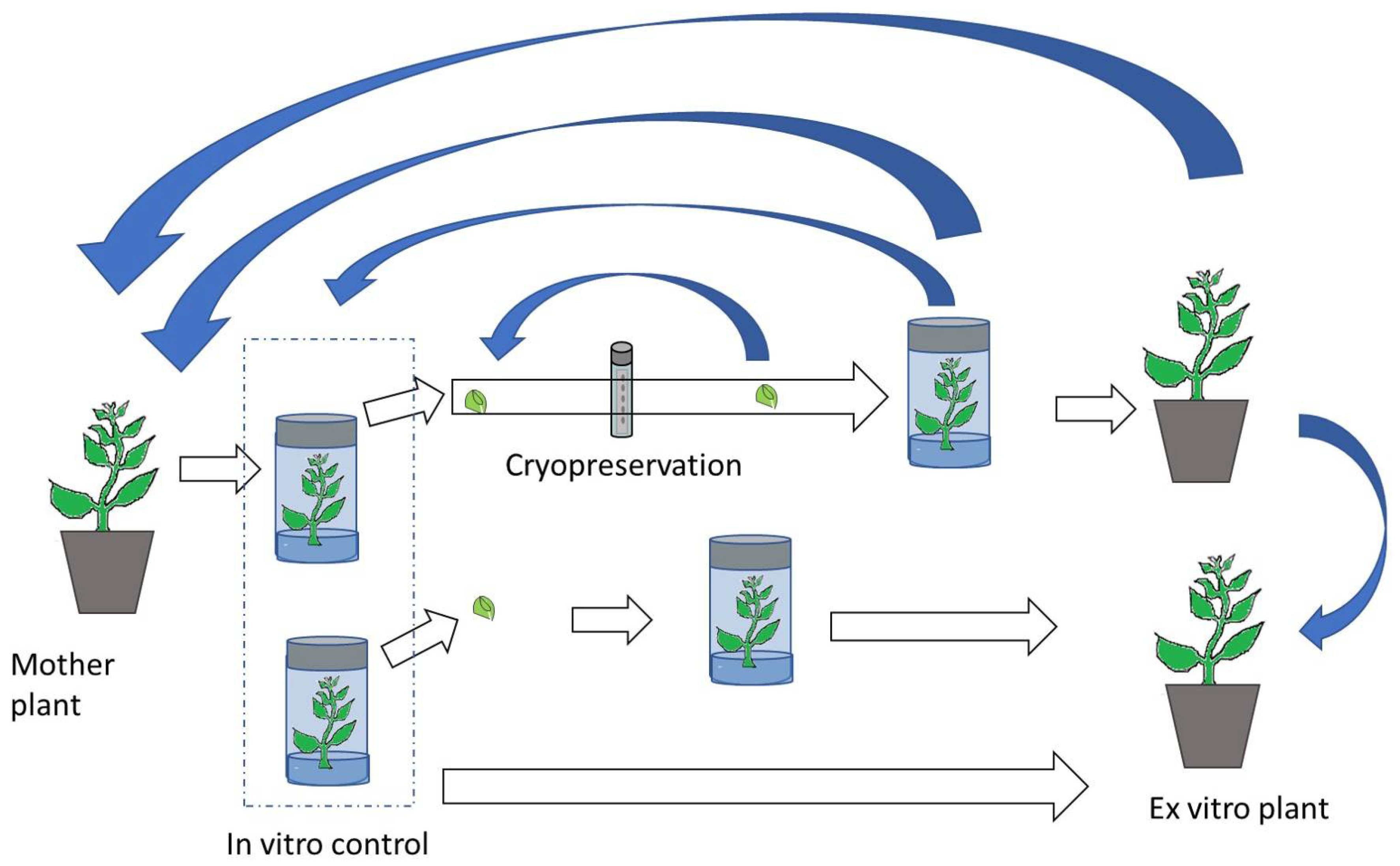
4. In Vitro Plant Conservation
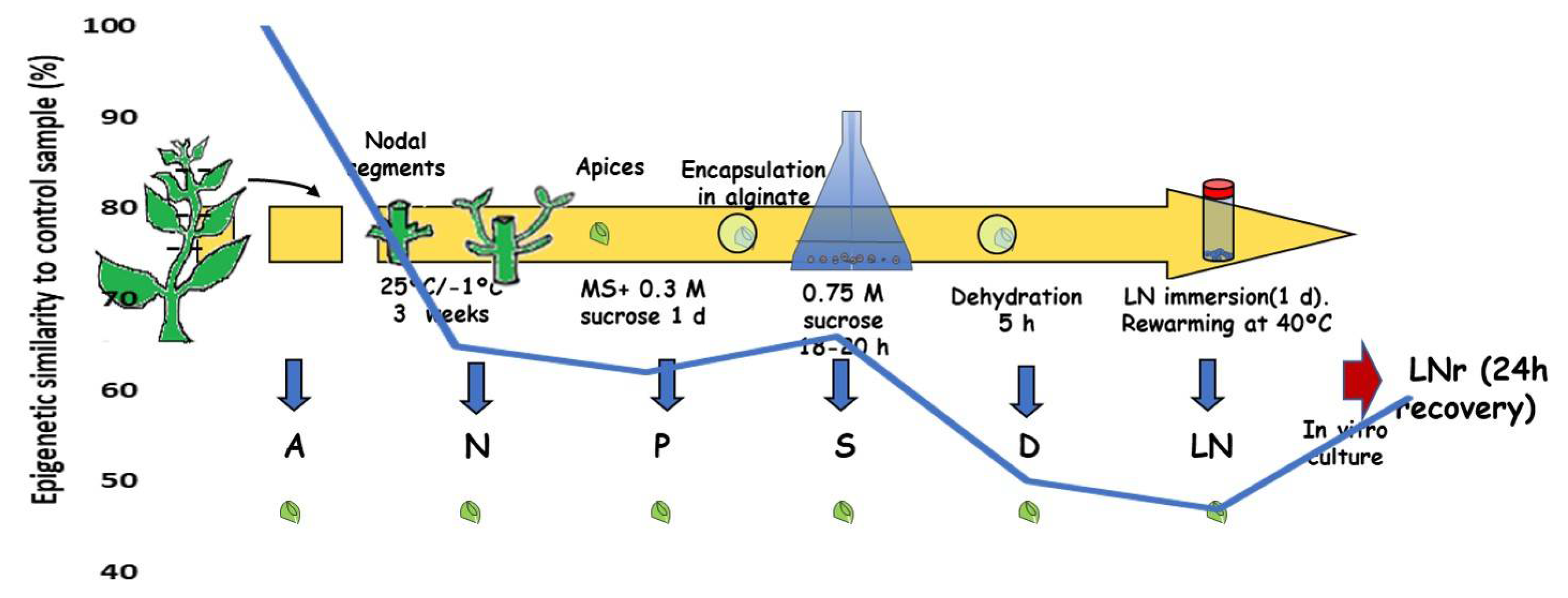
5. Cryopreservation

{kind=link}
{kind=link}
| Species | Studied Organ | Control | Conservation Technique | Genetic Stability | Detected DNA Methylation Variability | Ref. |
|---|---|---|---|---|---|---|
| Seed conservation | ||||||
| Zea mays | Caryopsis | Non-cryopreserved caryopsis | Caryopsis stored 12% wc. fwb. and storage in liquid nitrogen for 1 year | NA | Increase in the DNA methylation percentage from 65.2% to 72.6% | [65] |
| Secale cereale | Embryo and seedlings | Embryos and seedlings from non-stored caryopsis | Caryopsis stored 35 °C and 15% wc. fwb. stored for 13 or 29 days | √ RAPD in embryos, 5% changes in seedlings | 15–30% DNA methylation changes in seeds; 13–27% in seedlings | [58] |
| Mentha aquatica | Seeds and seedlings | Non-stored seeds, and seedlings from non-stored seeds | Seeds stored at 35 °C and 12% wc. for 28 days | √ RAPD in seeds; 13 % in seedlings | 8% DNA methylation changes in seeds, 16% in seedlings | [59] |
| In vitro slow growth | ||||||
| Malus pumila cv. Gala | In vitro shoots, from single bud | In vitro buds; state of development not stated | Half-strength medium, sucrose reduction, 2% mannitol, 4 °C, 12 h photoperiod, for 1 year | √ AFLP | 6 changed markers out of 389 (changes from DNA hemi-methylation to full methylation status) | [78] |
| Citrus paradise cv. Red Marsh | Embryogenic callus | Embryogenic callus | Half-strength medium, sucrose reduction, 10 °C, darkness, for 1 year | √ RAPD | 1 DNA demethylation marker out of 314 | [79] |
| Humulus lupulus | In vitro shoot cultures | Greenhouse plants | 4 °C, 12 h photoperiod, for 1 year | √ RAPD, AFLP | 35.7% loci changed, of which 4–11% DNA demethylation | [80,81] |
| Cryopreservation | ||||||
| Malus pumila cv. M26 | In vitro shoots from cryopreser-ved apices, from single bud | In vitro buds; state of development not stated | Encapsulation–Dehydration | √ AFLP | 5 DNA demethylation markers out of 380 | [106] |
| Fragaria vesca | In vitro shoots from cryopreser-ved apices, from single bud | In vitro shoots | Encapsulation–Dehydration | √ AFLP | 1 DNA demethylation markers out of 314 | [108] |
| Citrus | Callus after cryopreser-vation, single cell line | Callus | Vitrification PVS2 | √ RAPD | 1 DNA de novo methylation, 3 DNA demethylation markers, out of 358 | [109] |
| Humulus lupulus | In vitro shoot form cryopreser-ved apices | Greenhouse plants | Slow cooling | √ RAPD, AFLP | 36.73% loci polymorphic, of which aprox 47% DNA demethylation | [80] |
| Solanum tuberosum | In vitro plants from cryopreser-ved shoot tips | In vitro plants | DMSO-droplet method | NA | 3 DNA demethylation and 1 DNA de novo methylation markers out of 469 | [110] |
| Theobroma cacao | SE (cryo + in vitro SE) | Leave from ortet tree | Vitrification | NA | DNA methylation-related distances of 0.5 (similar to those of in vitro SE) | [98] |
| Menthax piperita | In vitro shoot apices after each step of the protocol | In vitro shoot apices | Encapsulation-dehydration | √ AFLP, RAPD | 53% DNA methylation changes were observed (being 59% de novo methylation), which was reduced to 40.8% after one day recovery | [112] |
| Actinidia chinensis var. deliciosa. | 8wk-old in vitro shoots derived from cryopreser-ved apices, and 3 mo-old ex vitro plants | Corresponding in vitro-derived samples | Droplet-vitrification | √ ISSR, AFLP | In vitro: 22 DNA de novo methylation and 30 DNA demethylation markers out of 718 Ex vitro: 6 DNA de novo methylation and one DNA demethylation marker out of 701 | [111] |
6. Statistical Methods for MSAP Analysis in Plant Conservation
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AFLP | Amplified Fragment Length Polymorphism |
| AMP | Amplified DNA methylation polymorphism |
| CMT | Chromomethylase |
| DNMT | DNA Methyltransferases |
| DRM | Domains Rearranged Methyltransferase |
| fwb | Fresh weight basis |
| HPLC | high-performance liquid chromatography |
| ISSR | Inter Simple Sequence Repeat |
| LN | Liquid Nitrogen |
| MAI | Methylation Analysis Inference |
| MET | Methyltransferase |
| metAFLP | methylation-sensitive Amplified Fragment Length Polymorphism |
| MSAP | Methylation Sensitive Amplified Polymorphism |
| RAPD | Random Amplified Polymorphic DNA |
| RdDM | RNA-directed DNA methylation |
| ROS | Reactive Oxygen Species |
| SSR | Simple Sequence Repeat |
| TLC | Thin-layer chromatography |
| UPGMA | Unweighted pair group method with arithmetic mean |
| wc. | water content |
References
- Corlett, R.T. A bigger toolbox: Biotechnology in biodiversity conservation. Trends Biotechnol. 2017, 35, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Coelho, N.; Gonçalves, S.; Romano, A. Endemic plant species conservation: Biotechnological approaches. Plants 2020, 9, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.-B. The vulnerability of plant genetic resources conserved ex situ. Crop Sci. 2017, 57, 2314–2328. [Google Scholar] [CrossRef] [Green Version]
- Buitink, J.; Leprince, O. Intracellular glasses and seed survival in the dry state. C. R. Biol. 2008, 331, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Zinsmeister, J.; Leprince, O.; Buitink, J. Molecular and environmental factors regulating seed longevity. Biochem. J. 2020, 477, 305–323. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-R.; Lambardi, M.; Engelmann, F.; Pathirana, R.; Panis, B.; Volk, G.M.; Wang, Q.-C. Advances in cryopreservation of in vitro-derived propagules: Technologies and explant sources. Plant Cell Tissue Organ Cult. 2020. [Google Scholar] [CrossRef]
- Gonzalez Benito, M.E.; Clavero-Ramírez, I.; López-Aranda, J.M. Review. The use of cryopreservation for germplasm conservation of vegetatively propagated crops. Span. J. Agric. Res. 2004, 2, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Flanagan, S.P.; Forester, B.R.; Latch, E.K.; Aitken, S.N.; Hoban, S. Guidelines for planning genomic assessment and monitoring of locally adaptive variation to inform species conservation. Evol. Appl. 2018, 11, 1035–1052. [Google Scholar] [CrossRef]
- Eizaguirre, C.; Baltazar-Soares, M. Evolutionary conservation-evaluating the adaptive potential of species. Evol. Appl. 2014, 7, 963–967. [Google Scholar] [CrossRef] [Green Version]
- Keller, T.E.; Lasky, J.R.; Yi, S.V. The multivariate association between genomewide DNA methylation and climate across the range of Arabidopsis thaliana. Mol. Ecol. 2016, 25, 1823–1837. [Google Scholar] [CrossRef]
- Jeremias, G.; Barbosa, J.; Marques, S.M.; Asselman, J.; Gonçalves, F.J.M.; Pereira, J.L. Synthesizing the role of epigenetics in the response and adaptation of species to climate change in freshwater ecosystems. Mol. Ecol. 2018, 27, 2790–2806. [Google Scholar] [CrossRef] [Green Version]
- Bender, J. DNA methylation and epigenetics. Annu. Rev. Plant Biol. 2004, 55, 41–68. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X. Comparative epigenomics: A powerful tool to understand the evolution of DNA methylation. New Phytol. 2016, 210, 76–80. [Google Scholar] [CrossRef]
- Pei, L.; Zhang, L.; Li, J.; Shen, C.; Qiu, P.; Tu, L.; Zhang, X.; Wang, M. Tracing the origin and evolution history of methylation-related genes in plants. BMC Plant Biol. 2019, 19, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, J.A.; Jacobsen, S.E. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat. Rev. Genet. 2010, 11, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, E.; Peacock, W.; Dennis, E. DNA methylation, a key regulator of plant development and other processes. Curr. Opin. Genet. Dev. 2000, 10, 217–223. [Google Scholar] [CrossRef]
- Martienssen, R.A.; Colot, V. DNA methylation and epigenetic inheritance in plants and filamentous fungi. Science 2001, 293, 1070–1074. [Google Scholar] [CrossRef] [Green Version]
- Matzke, M.A.; Mosher, R.A. RNA-directed DNA methylation: An epigenetic pathway of increasing complexity. Nat. Rev. Genet. 2014, 15, 394–408. [Google Scholar] [CrossRef]
- Rey, O.; Eizaguirre, C.; Angers, B.; Baltazar-Soares, M.; Sagonas, K.; Prunier, J.G.; Blanchet, S. Linking epigenetics and biological conservation: Towards a conservation epigenetics perspective. Funct. Ecol. 2020, 34, 414–427. [Google Scholar] [CrossRef] [Green Version]
- Vogt, G. Facilitation of environmental adaptation and evolution by epigenetic phenotype variation: Insights from clonal, invasive, polyploid, and domesticated animals. Environ. Epigenet. 2017, 3. [Google Scholar] [CrossRef]
- Feng, S.; Jacobsen, S.; Reik, W. Epigenetic reprogramming in plant and animal development. Science 2010, 330, 622–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quadrana, L.; Colot, V. Plant transgenerational epigenetics. Annu. Rev. Genet. 2016, 50, 467–491. [Google Scholar] [CrossRef] [PubMed]
- Dubrovina, A.S.; Kiselev, K.V. Age-associated alterations in the somatic mutation and DNA methylation levels in plants. Plant Biol. 2016, 18, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Mithani, A.; Belfield, E.J.; Mott, R.; Hurst, L.D.; Harberd, N.P. Environmentally responsive genome-wide accumulation of de novo Arabidopsis thaliana mutations and epimutations. Genome Res. 2014, 24, 1821–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhoeven, K.J.F.; Preite, V. Epigenetic variation in asexually reproducing organisms. Evolution 2013, 68, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Yong, W.-S.; Hsu, F.-M.; Chen, P.-Y. Profiling genome-wide DNA methylation. Epigenet. Chromatin 2016, 9, 26. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.M.; Bird, A. DNA methylation landscapes: Provocative insights from epigenomics. Nat. Rev. Genet. 2008, 9, 465–476. [Google Scholar] [CrossRef]
- Dahl, C.; Guldberg, P. DNA methylation analysis techniques. Biogerontology 2003, 4, 233–250. [Google Scholar] [CrossRef]
- Grunau, C.; Clark, S.J.; Rosenthal, A. Bisulfite genomic sequencing: Systematic investigation of critical experimental parameters. Nucleic Acids Res. 2001, 29, e65. [Google Scholar] [CrossRef]
- Reyna-López, G.E.; Simpson, J.; Ruiz-Herrera, J. Differences in DNA methylation patterns are detectable during the dimorphic transition of fungi by amplification of restriction polymorphisms. Mol. Gen. Genet. MGG 1997, 253, 703–710. [Google Scholar] [CrossRef]
- Xiong, L.Z.; Xu, C.G.; Maroof, M.S.; Zhang, Q. Patterns of cytosine methylation in an elite rice hybrid and its parental lines, detected by a methylation-sensitive amplification polymorphism technique. Mol. Gen. Genet. MGG 1999, 261, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Portis, E.; Acquadro, A.; Comino, C.; Lanteri, S. Analysis of DNA methylation during germination of pepper (Capsicum annuum L.) seeds using methylation-sensitive amplification polymorphism (MSAP). Plant Sci. 2004, 166, 169–178. [Google Scholar] [CrossRef]
- Meng, F.R.; Li, Y.C.; Yin, J.; Liu, H.; Chen, X.J.; Ni, Z.F.; Sun, Q.X. Analysis of DNA methylation during the germination of wheat seeds. Biol. Plant. 2012, 56, 269–275. [Google Scholar] [CrossRef]
- Rodriguez, M.P.; Cervigni, G.D.L.; Quarin, C.L.; Ortiz, J.P.A. Frequencies and variation in cytosine methylation patterns in diploid and tetraploid cytotypes of Paspalum notatum. Biol. Plant. 2012, 56, 276–282. [Google Scholar] [CrossRef]
- Takata, M.; Kishima, Y.; Sano, Y. DNA Methylation polymorphisms in rice and wild rice strains: Detection of epigenetic markers. Breed. Sci. 2005, 55, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.-S.; Pan, Y.-J.; Zhao, X.-Q.; Dwivedi, D.; Zhu, L.-H.; Ali, J.; Fu, B.-Y.; Li, Z.-K. Drought-induced site-specific DNA methylation and its association with drought tolerance in rice (Oryza sativa L.). J. Exp. Bot. 2011, 62, 1951–1960. [Google Scholar] [CrossRef]
- Aina, R.; Sgorbati, S.; Santagostino, A.; Labra, M.; Ghiani, A.; Citterio, S. Specific hypomethylation of DNA is induced by heavy metals in white clover and industrial hemp. Physiol. Plant. 2004, 121, 472–480. [Google Scholar] [CrossRef]
- Greco, M.; Chiappetta, A.; Bruno, L.; Bitonti, M.B. Effects of light deficiency on genome methylation in Posidonia oceanica. Mar. Ecol. Prog. Ser. 2013, 473, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Schulz, B.; Eckstein, R.L.; Durka, W. Scoring and analysis of methylation-sensitive amplification polymorphisms for epigenetic population studies. Mol. Ecol. Resour. 2013, 13, 642–653. [Google Scholar] [CrossRef] [Green Version]
- Fulnecek, J.; Kovarik, A. How to interpret methylation sensitive amplified polymorphism (MSAP) profiles? BMC Genet. 2014, 15, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Salmon, A.; Clotault, J.; Jenczewski, E.; Chable, V.; Manzanares-Dauleux, M.J. Brassica oleracea displays a high level of DNA methylation polymorphism. Plant Sci. 2008, 174, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Herrera, C.; Bazaga, P. Epigenetic differentiation and relationship to adaptive genetic divergence in discrete populations of the violet Viola cazorlensis. New Phytol. 2010, 187, 867–876. [Google Scholar] [CrossRef]
- Richards, C.L.; Schrey, A.W.; Pigliucci, M. Invasion of diverse habitats by few Japanese knotweed genotypes is correlated with epigenetic differentiation. Ecol. Lett. 2012, 15, 1016–1025. [Google Scholar] [CrossRef]
- REBASE. The Restriction Enzyme Database. Available online: http://rebase.neb.com/rebase/rebase.html (accessed on 7 August 2020).
- Schrey, A.W.; Alvarez, M.; Foust, C.M.; Kilvitis, H.J.; Lee, J.D.; Liebl, A.L.; Martin, L.B.; Richards, C.L.; Robertson, M. Ecological epigenetics: Beyond MS-AFLP. Integr. Comp. Biol. 2013, 53, 340–350. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Aranzana, M.J.; Kim, S.; Lister, C.; Shindo, C.; Tang, C.; Toomajian, C.; Zheng, H.; Dean, C.; Marjoram, P.; et al. An Arabidopsis example of association mapping in structured samples. PLoS Genet. 2007, 3, e4. [Google Scholar] [CrossRef] [Green Version]
- Bednarek, P.T.; Orłowska, R.; Koebner, R.M.D.; Zimny, J. Quantification of the tissue-culture induced variation in barley (Hordeum vulgare L.). BMC PlantBiol. 2007, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.-B.; Ahmed, Z.; Diederichsen, A. Towards a better monitoring of seed ageing under ex situ seed conservation. Conserv. Physiol. 2015, 3, cov026. [Google Scholar] [CrossRef] [Green Version]
- WIEWS. World Information and Early Warning System on Plant Genetic Resources for Food and Agriculture. Available online: http://www.fao.org/wiews/en/ (accessed on 15 June 2020).
- Kurek, K.; Plitta-Michalak, B.; Ratajczak, E. Reactive oxygen species as potential drivers of the seed aging process. Plants 2019, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- Roach, T.; Nagel, M.; Börner, A.; Eberle, C.; Kranner, I. Changes in tocochromanols and glutathione reveal differences in the mechanisms of seed ageing under seedbank conditions and controlled deterioration in barley. Environ. Exp. Bot. 2018, 156, 8–15. [Google Scholar] [CrossRef]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.Oxford University Press: New York, NY, USA, 2015; ISBN 978-0-19-180213-3. [Google Scholar]
- Fleming, M.B.; Hill, L.M.; Walters, C. The kinetics of ageing in dry-stored seeds: A comparison of viability loss and RNA degradation in unique legacy seed collections. Ann. Bot. 2019, 123, 1133–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streczynski, R.; Clark, H.; Whelehan, L.M.; Ang, S.-T.; Hardstaff, L.K.; Funnekotter, B.; Bunn, E.; Offord, C.A.; Sommerville, K.D.; Mancera, R.L. Current issues in plant cryopreservation and importance for ex situ conservation of threatened Australian native species. Aust. J. Bot. 2019, 67, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Z.; Shah, Z.H.; Rehman, H.M.; Shahzad, K.; Daur, I.; Elfeel, A.; Hassan, M.U.; Elsafori, A.K.; Yang, S.H.; Chung, G. Genomics: A hallmark to monitor molecular and biochemical processes leading toward a better perceptive of seed aging and ex-situ conservation. Curr. Issues Mol. Biol. 2017, 89–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirredda, M.; González-Benito, M.E.; Martín, C.; Mira, S. Genetic and epigenetic stability in rye seeds under different storage conditions: Ageing and oxygen effect. Plants 2020, 9, 393. [Google Scholar] [CrossRef] [Green Version]
- Mira, S.; Pirredda, M.; Martín-Sánchez, M.; Marchessi, J.E.; Martín, C. DNA methylation and integrity in aged seeds and regenerated plants. Seed Sci. Res. 2020, 1–9. [Google Scholar] [CrossRef]
- Barciszewska, M.Z.; Barciszewska, A.M.; Rattan, S.I.S. TLC-based detection of methylated cytosine: Application to aging epigenetics. Biogerontology 2007, 8, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Guzy-Wrobelska, J.; Filek, M.; Kaliciak, A.; Szarejko, I.; Macháčková, I.; Krekule, J.; Barciszewska, M. Vernalization and photoperiod-related changes in the DNA methylation state in winter and spring rapeseed. Acta Physiol. Plant 2013, 35, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Michalak, M.; Barciszewska, M.Z.; Barciszewski, J.; Plitta, B.P.; Chmielarz, P. Global changes in DNA methylation in seeds and seedlings of Pyrus communis after seed desiccation and storage. PLoS ONE 2013, 8, e70693. [Google Scholar] [CrossRef] [Green Version]
- Plitta, B.P.; Michalak, M.; Bujarska-Borkowska, B.; Barciszewska, M.Z.; Barciszewski, J.; Chmielarz, P. Effect of desiccation on the dynamics of genome-wide DNA methylation in orthodox seeds of Acer platanoides L. Plant Physiol. Biochem. 2014, 85, 71–77. [Google Scholar] [CrossRef]
- Plitta-Michalak, B.P.; Naskręt-Barciszewska, M.Z.; Kotlarski, S.; Tomaszewski, D.; Tylkowski, T.; Barciszewski, J.; Chmielarz, P.; Michalak, M. Changes in genomic 5-methylcytosine level mirror the response of orthodox (Acer platanoides L.) and recalcitrant (Acer pseudoplatanus L.) seeds to severe desiccation. Tree Physiol. 2018, 38, 617–629. [Google Scholar] [CrossRef] [Green Version]
- Pérez, J.; Araya-Valverde, E.; Garro-Monge, G.; Abdelnour-Esquivel, A. Analysis of stress indicators during cryopreservation of seeds of landrace maize (Zea mays). CryoLetters 2017, 38, 445–454. [Google Scholar]
- Lu, J.; Greene, S.; Reid, S.; Cruz, V.M.V.; Dierig, D.A.; Byrne, P. Phenotypic changes and DNA methylation status in cryopreserved seeds of rye (Secale cereale L.). Cryobiology 2018, 82, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Engelmann, F. Use of biotechnologies for the conservation of plant biodiversity. Vitro Cell. Dev. Biol. Plant 2011, 47, 5–16. [Google Scholar] [CrossRef]
- Ashmore, S.E. Status Report on The Development and Application of In Vitro Techniques for the Conservation and Use of Plant Genetic Resources; International Plant Genetics Research Institute: Rome, Italy, 1997; ISBN 978-92-9043-339-2. [Google Scholar]
- Reed, B.M.; Engelmann, F.; Dulloo, M.E.; Engels, J.M. Technical Guidelines for the Management of Field and In Vitro Germplasm Collections; International Plant Genetic Resources Institute: Rome, Italy, 2004; ISBN 978-92-9043-640-9. [Google Scholar]
- Da Silva, D.P.C.; Ozudogru, E.A.; Dos Reis, M.V.; Lambardi, M. In vitro conservation of ornamental plants. Ornam. Hortic. 2018, 24, 28–33. [Google Scholar] [CrossRef]
- Sarasan, V.; Cripps, R.; Ramsay, M.M.; Atherton, C.; McMichen, M.; Prendergast, G.; Rowntree, J.K. Conservation in vitro of threatened plants—Progress in the past decade. Vitro Cell. Dev. Biol. Plant 2006, 42, 206–214. [Google Scholar] [CrossRef]
- Bednarek, P.T.; Orłowska, R. Plant tissue culture environment as a switch-key of (epi)genetic changes. Plant Cell Tissue Organ Cult. 2020, 140, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Azizi, P.; Hanafi, M.M.; Sahebi, M.; Harikrishna, J.A.; Taheri, S.; Yassoralipour, A.; Nasehi, A. Epigenetic changes and their relationship to somaclonal variation: A need to monitor the micropropagation of plantation crops. Funct. Plant Biol. 2020, 47, 508–523. [Google Scholar] [CrossRef]
- Lázaro-Castellanos, J.O.; Mata-Rosas, M.; González, D.; Arias, S.; Reverchon, F. In vitro propagation of endangered Mammillaria genus (Cactaceae) species and genetic stability assessment using SSR markers. Vitro Cell. Dev. Biol. Plant 2018, 54, 518–529. [Google Scholar] [CrossRef]
- Hammond Hammond, S.D.; Viehmannova, I.; Zamecnik, J.; Panis, B.; Cepkova, P.H. Efficient slow-growth conservation and assessment of clonal fidelity of Ullucus tuberosus Caldas microshoots. Plant Cell Tissue Organ Cult. 2019, 138, 559–570. [Google Scholar] [CrossRef]
- Samarina, L.; Gvasaliya, M.; Koninskaya, N.; Rakhmangulov, R.; Efremov, A.; Kiselyova, N.; Ryndin, A.; Hanke, M.-V. A comparison of genetic stability in tea [Camellia sinensis (L.) Kuntze] plantlets derived from callus with plantlets from long-term in vitro propagation. Plant Cell Tissue Organ Cult. 2019, 138, 467–474. [Google Scholar] [CrossRef]
- Harding, K. The methylation status of DNA derived from potato plants recovered from slow growth. Plant Cell Tissue Organ Cult. 1994, 37, 31–38. [Google Scholar] [CrossRef]
- Hao, Y.-J.; Deng, X. Genetically stable regeneration of apple plants from slow growth. Plant Cell Tissue Organ Cult. 2003, 72, 253–260. [Google Scholar] [CrossRef]
- Hao, Y.-J.; Wen, X.-P.; Deng, X. Genetic and epigenetic evaluations of citrus calluses recovered from slow-growth culture. J. Plant Physiol. 2004, 161, 479–484. [Google Scholar] [CrossRef]
- Peredo, E.L.; Arroyo-García, R.; Reed, B.M.; Revilla, M.A. Genetic and epigenetic stability of cryopreserved and cold-stored hops (Humulus lupulus L.). Cryobiology 2008, 57, 234–241. [Google Scholar] [CrossRef]
- Peredo, E.L.; Arroyo-García, R.; Reed, B.M.; Revilla, M.A. Genetic stability of in vitro conserved germplasm of Humulus lupulus L. Agric. Food Sci. 2009, 18, 144–151. [Google Scholar] [CrossRef]
- Kundu, S.; Salma, U.; Ali, M.N.; Mandal, N. Conservation, ex vitro direct regeneration, and genetic uniformity assessment of alginate-encapsulated nodal cuttings of Sphagneticola calendulacea (L.) Pruski. Acta Physiol. Plant. 2018, 40, 53. [Google Scholar] [CrossRef]
- Arumugam, G.; Sinniah, U.R.; Swamy, M.K.; Lynch, P.T. Encapsulation of in vitro Plectranthus amboinicus (Lour.) Spreng. shoot apices for propagation and conservation. 3 Biotech 2019, 9, 298. [Google Scholar] [CrossRef]
- Kikowska, M.; Sliwinska, E.; Thiem, B. Micropropagation and production of somatic seeds for short-term storage of the endangered species Eryngium alpinum L. Plants 2020, 9, 498. [Google Scholar] [CrossRef] [Green Version]
- Tavazza, R.; Rey, N.A.; Papacchioli, V.; Pagnotta, M.A. A validated slow-growth in vitro conservation protocol for globe artichoke germplasm: A cost-effective tool to preserve from wild to elite genotypes. Sci. Hortic. 2015, 197, 135–143. [Google Scholar] [CrossRef]
- Koeda, S.; Matsumoto, S.; Matsumoto, Y.; Takisawa, R.; Nishikawa, K.; Kataoka, K. Medium-term in vitro conservation of virus-free parthenocarpic tomato plants. Vitro Cell. Dev. Biol. Plant 2018, 54, 392–398. [Google Scholar] [CrossRef]
- Kamińska, M.; Gołębiewski, M.; Tretyn, A.; Trejgell, A. Efficient long-term conservation of Taraxacum pieninicum synthetic seeds in slow growth conditions. Plant Cell Tissue Organ Cult. 2018, 132, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Kaeppler, S.M.; Kaeppler, H.F.; Rhee, Y. Epigenetic aspects of somaclonal variation in plants. Plant Mol. Biol. 2000, 43, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, M.D.; Yañez-Santos, A.M.; Paz, R.C.; Quiroga, M.P.; Marfil, C.F.; Conci, V.C.; García-Lampasona, S.C. Assessment of genetic and epigenetic changes in virus-free garlic (Allium sativum L.) plants obtained by meristem culture followed by in vitro propagation. Plant Cell Rep. 2016, 35, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Normah, M.N.; Sulong, N.; Reed, B.M. Cryopreservation of shoot tips of recalcitrant and tropical species: Advances and strategies. Cryobiology 2019, 87, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Höfer, M.; Hanke, M.-V. Cryopreservation of fruit germplasm. Vitro Cell. Dev. Biol. Plant 2017, 53, 372–381. [Google Scholar] [CrossRef]
- Niino, T.; Valle Arizaga, M. Cryopreservation for preservation of potato genetic resources. Breed. Sci. 2015, 65, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Popova, E.; Shukla, M.; Kim, H.H.; Saxena, P.K. Plant cryopreservation for biotechnology and breeding. In Advances in Plant Breeding Strategies: Breeding, Biotechnology and Molecular Tools; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: New York, NY, USA, 2015; pp. 63–93. ISBN 978-3-319-22521-0. [Google Scholar]
- Reed, B.M. Plant Cryopreservation: A Practical Guide; Reed, B.M., Ed.; Springer: New York, NY, USA, 2008; ISBN 978-0-387-72275-7. [Google Scholar]
- Kaczmarczyk, A.; Funnekotter, B.; Menon, A.; Phang, P.; Al-Hanbali, A.; Bunn, E.; Mancera, R.L. Current issues in plant cryopreservation. In Current Frontiers in Cryobiology; Katkov, I.I., Ed.; IntechOpen Ltd.: London, UK, 2012; pp. 417–438. ISBN 978-953-51-0191-8. [Google Scholar]
- Benson, E.E.; Bremner, D. Oxidative stress in the frozen plant: A free radical point of view. In Life in the Frozen State; Fuller, B.J., Lan, N., Benson, E.E., Eds.; CRC Press: Boca Raton, FL, USA, 2004; pp. 205–242. ISBN 978-0-429-21248-2. [Google Scholar]
- Uchendu, E.E.; Leonard, S.W.; Traber, M.G.; Reed, B.M. Vitamins C and E improve regrowth and reduce lipid peroxidation of blackberry shoot tips following cryopreservation. Plant Cell Rep. 2010, 29, 25–35. [Google Scholar] [CrossRef]
- Adu-Gyamfi, R.; Wetten, A.; Rodríguez López, C.M. Effect of cryopreservation and post-cryopreservation somatic embryogenesis on the epigenetic fidelity of cocoa (Theobroma cacao L.). PLoS ONE 2016, 11, e0158857. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Saha, D.; Niemann, H.; Gryshkov, O.; Glasmacher, B.; Hofmann, N. Effects of cryopreservation on the epigenetic profile of cells. Cryobiology 2017, 74, 1–7. [Google Scholar] [CrossRef]
- Funnekotter, B.; Mancera, R.L.; Bunn, E. Advances in understanding the fundamental aspects required for successful cryopreservation of Australian flora. Vitro Cell. Dev. Biol. Plant 2017, 53, 289–298. [Google Scholar] [CrossRef]
- Matsumoto, T. Cryopreservation of plant genetic resources: Conventional and new methods. Rev. Agric. Sci. 2017, 5, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Li, J.-W.; Ozudogru, E.A.; Li, J.; Wang, M.-R.; Bi, W.-L.; Lambardi, M.; Wang, Q.-C. Cryobiotechnology of forest trees: Recent advances and future prospects. Biodivers. Conserv. 2018, 27, 795–814. [Google Scholar] [CrossRef]
- Kaity, A.; Drew, R.A.; Ashmore, S.E. Genetic and epigenetic integrity assessment of acclimatised papaya plants regenerated directly from shoot-tips following short- and long-term cryopreservation. Plant Cell Tissue Organ Cult. 2013, 112, 75–86. [Google Scholar] [CrossRef]
- Mikuła, A.; Tomiczak, K.; Rybczyński, J.J. Cryopreservation enhances embryogenic capacity of Gentiana cruciata (L.) suspension culture and maintains (epi)genetic uniformity of regenerants. Plant Cell Rep. 2011, 30, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Johnston, J.W.; Benson, E.E.; Harding, K. Cryopreservation induces temporal DNA methylation epigenetic changes and differential transcriptional activity in Ribes germplasm. Plant Physiol. Biochem. 2009, 47, 123–131. [Google Scholar] [CrossRef]
- Hao, Y.-J.; Liu, Q.-L.; Deng, X.-X. Effect of cryopreservation on apple genetic resources at morphological, chromosomal, and molecular levels. Cryobiology 2001, 43, 46–53. [Google Scholar] [CrossRef]
- Osorio-Montalvo, P.; Sáenz-Carbonell, L.; De-la-Peña, C. 5-Azacytidine: A promoter of epigenetic changes in the quest to improve plant somatic embryogenesis. Int. J. Mol. Sci. 2018, 19, 3182. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.-J.; You, C.-X.; Deng, X.-X. Analysis of ploidy and the patterns of amplified fragment length polymorphism and methylation sensitive amplified polymorphism in strawberry plants recovered from cryopreservation. CryoLetters 2002, 23, 37–46. [Google Scholar]
- Hao, Y.-J.; You, C.-X.; Deng, X.-X. Effects of cryopreservation on developmental competency, cytological and molecular stability of citrus callus. CryoLetters 2002, 23, 27–35. [Google Scholar]
- Kaczmarczyk, A.; Houben, A.; Keller, E.R.J.; Mette, M.F. Influence of cryopreservation on the cytosine methylation state of potato genomic DNA. CryoLetters 2010, 31, 380–391. [Google Scholar] [PubMed]
- Zhang, X.-C.; Bao, W.-W.; Zhang, A.; Pathirana, R.; Wang, Q.-C.; Liu, Z.-D. Cryopreservation of shoot tips, evaluations of vegetative growth, and assessments of genetic and epigenetic changes in cryo-derived plants of Actinidia spp. Cryobiology 2020, 94, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, M.A.; Alvarez-Mari, A.; Rodríguez-Sanz, H.; Kremer, C.; González-Benito, M.E.; Martín, C. Genetic and epigenetic stability of recovered mint apices after several steps of a cryopreservation protocol by encapsulation-dehydration. A new approach for epigenetic analysis. Plant Physiol. Biochem. 2019, 143, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lang, Z.; Zhu, J.-K. Dynamics and function of DNA methylation in plants. Nat. Rev. Mol. Cell Biol. 2018, 19, 489–506. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Muñoz, R.; Moyano, E.; Khojasteh, A.; Bonfill, M.; Cusido, R.M.; Palazon, J. Genomic methylation in plant cell cultures: A barrier to the development of commercial long-term biofactories. Eng. Life Sci. 2019, 19, 872–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lele, L.; Ning, D.; Cuiping, P.; Xiao, G.; Weihua, G. Genetic and epigenetic variations associated with adaptation to heterogeneous habitat conditions in a deciduous shrub. Ecol. Evol. 2018, 8, 2594–2606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigo, J.M.; Zappacosta, D.C.; Selva, J.P.; Garbus, I.; Albertini, E.; Echenique, V. Apomixis frequency under stress conditions in weeping lovegrass (Eragrostis curvula). PLoS ONE 2017, 12, e0175852. [Google Scholar] [CrossRef]
- Liu, Z.-Z.; Chen, T.; Peng, F.-R.; Liang, Y.-W.; Tan, P.-P.; Mo, Z.-H.; Cao, F.; Shang, Y.-J.; Zhang, R.; Li, Y.-R. Variation in cytosine methylation among pecan cultivars at different developmental stages. J. Am. Soc. Hortic. Sci. 2018, 143, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Bonin, A.; Bellemain, E.; BronkenEidesen, P.; Pompanon, F.; Brochmann, C.; Taberlet, P. How to track and assess genotyping errors in population genetics studies: Tracking and assessing genotyping errors. Mol. Ecol. 2004, 13, 3261–3273. [Google Scholar] [CrossRef]
- Alsdurf, J.; Anderson, C.; Siemens, D.H. Epigenetics of drought-induced trans-generational plasticity: Consequences for range limit development. AoB Plants 2016, 8, plv146. [Google Scholar] [CrossRef] [Green Version]
- Alonso, C.; Pérez, R.; Bazaga, P.; Medrano, M.; Herrera, C.M. MSAP markers and global cytosine methylation in plants: A literature survey and comparative analysis for a wild-growing species. Mol. Ecol. Resour. 2016, 16, 80–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, L.; Li, M.; Fan, H.X.; Jiang, J.J.; Wang, Y.P.; Sokolov, V. Epigenetic variation in the callus of Brassica napus under different inducement conditions. Russ. J. Genet. 2016, 52, 802–809. [Google Scholar] [CrossRef]
- Goyali, J.C.; Igamberdiev, A.U.; Debnath, S.C. DNA methylation in lowbush blueberry (Vaccinium angustifolium Ait.) propagated by softwood cutting and tissue culture. Can. J. Plant Sci. 2018, 98, 1035–1044. [Google Scholar] [CrossRef]
- Jekayinoluwa, T.; Gueye, B.; Bhattacharjee, R.; Osibanjo, O.; Shah, T.; Abberton, M. Agromorphologic, genetic and methylation profiling of Dioscorea and Musa species multiplied under three micropropagation systems. PLoS ONE 2019, 14, e0216717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, A.; Igamberdiev, A.U.; Debnath, S.C. Detection of DNA methylation pattern in thidiazuron-induced blueberry callus using methylation-sensitive amplification polymorphism. Biol. Plant. 2017, 61, 511–519. [Google Scholar] [CrossRef]
- Bobadilla Landey, R.; Cenci, A.; Guyot, R.; Bertrand, B.; Georget, F.; Dechamp, E.; Herrera, J.-C.; Aribi, J.; Lashermes, P.; Etienne, H. Assessment of genetic and epigenetic changes during cell culture ageing and relations with somaclonal variation in Coffea arabica. Plant Cell Tissue Organ Cult. 2015, 122, 517–531. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Benito, M.E.; Ibáñez, M.Á.; Pirredda, M.; Mira, S.; Martín, C. Application of the MSAP Technique to Evaluate Epigenetic Changes in Plant Conservation. Int. J. Mol. Sci. 2020, 21, 7459. https://doi.org/10.3390/ijms21207459
González-Benito ME, Ibáñez MÁ, Pirredda M, Mira S, Martín C. Application of the MSAP Technique to Evaluate Epigenetic Changes in Plant Conservation. International Journal of Molecular Sciences. 2020; 21(20):7459. https://doi.org/10.3390/ijms21207459
Chicago/Turabian StyleGonzález-Benito, María Elena, Miguel Ángel Ibáñez, Michela Pirredda, Sara Mira, and Carmen Martín. 2020. "Application of the MSAP Technique to Evaluate Epigenetic Changes in Plant Conservation" International Journal of Molecular Sciences 21, no. 20: 7459. https://doi.org/10.3390/ijms21207459