MicroRNA-31 Regulates Expression of Wntless in Both Drosophila melanogaster and Human Oral Cancer Cells

 , , , and
, , , and
Abstract
:1. Introduction
2. Result
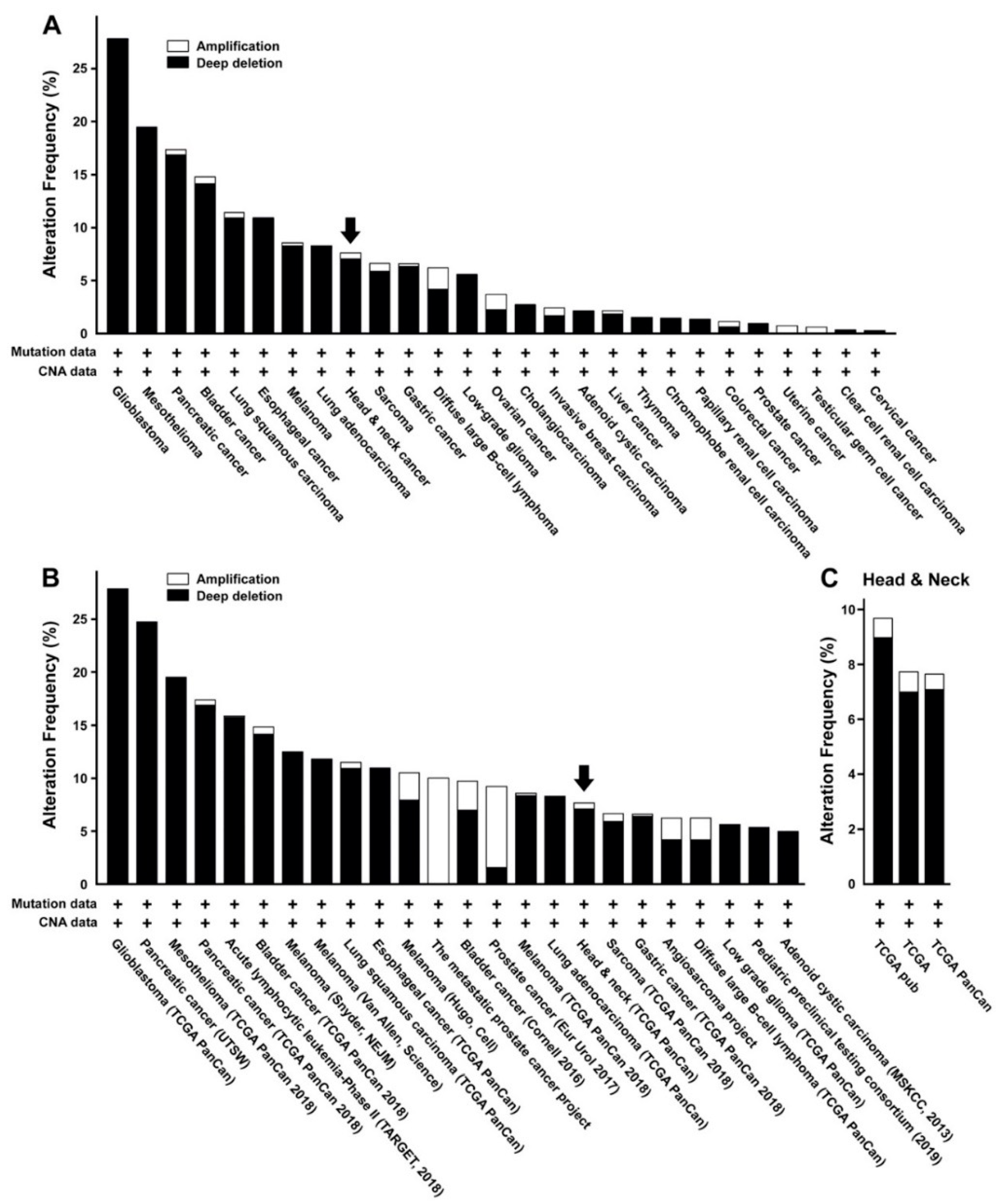
2.1. miR-31 is Differentially Regulated in Various Types of Malignant Tumors, Including Oral Cancer
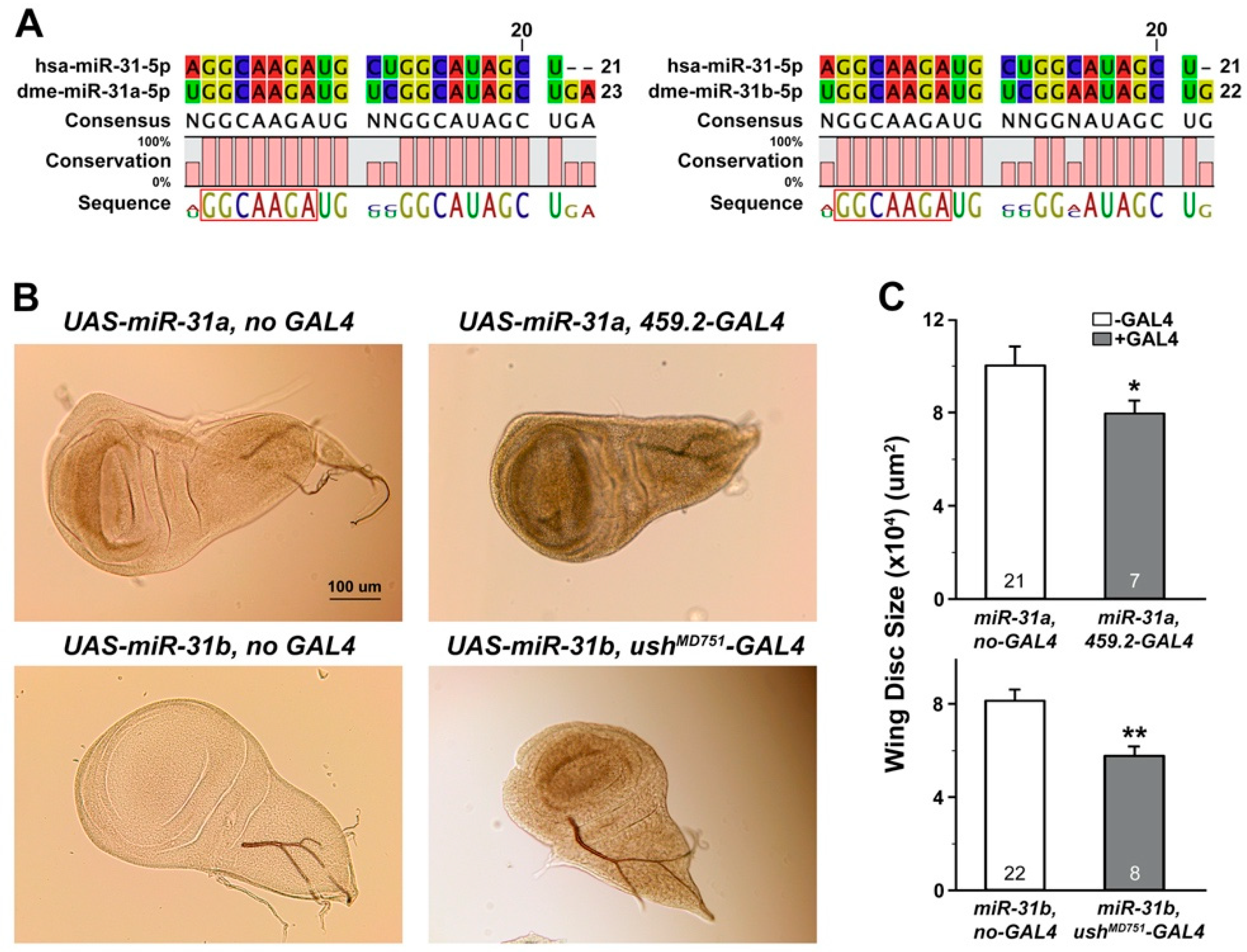
2.2. Overexpression of Drosophila miR-31a and miR-31b Induced Changes in the Size of Wing Discs and the Levels of Putative Target mRNAs
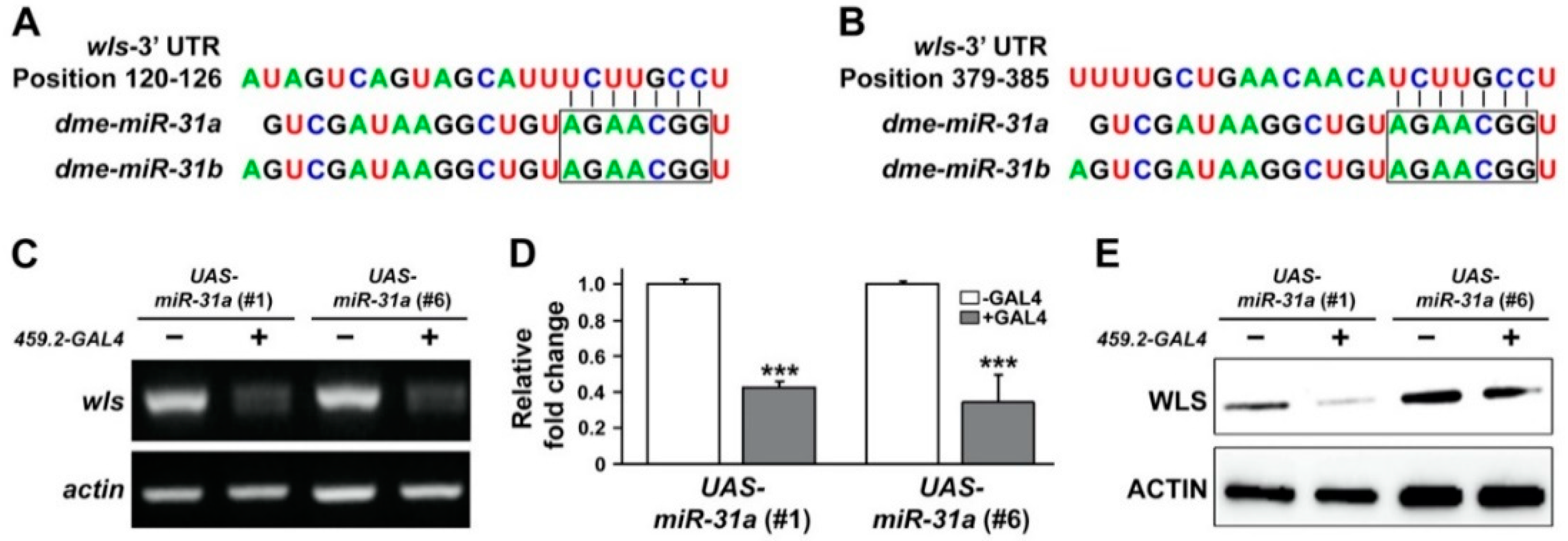
2.3. Overexpression of Drosophila miR-31a Reduced the Transcript Level of a Predicted Target, WLS
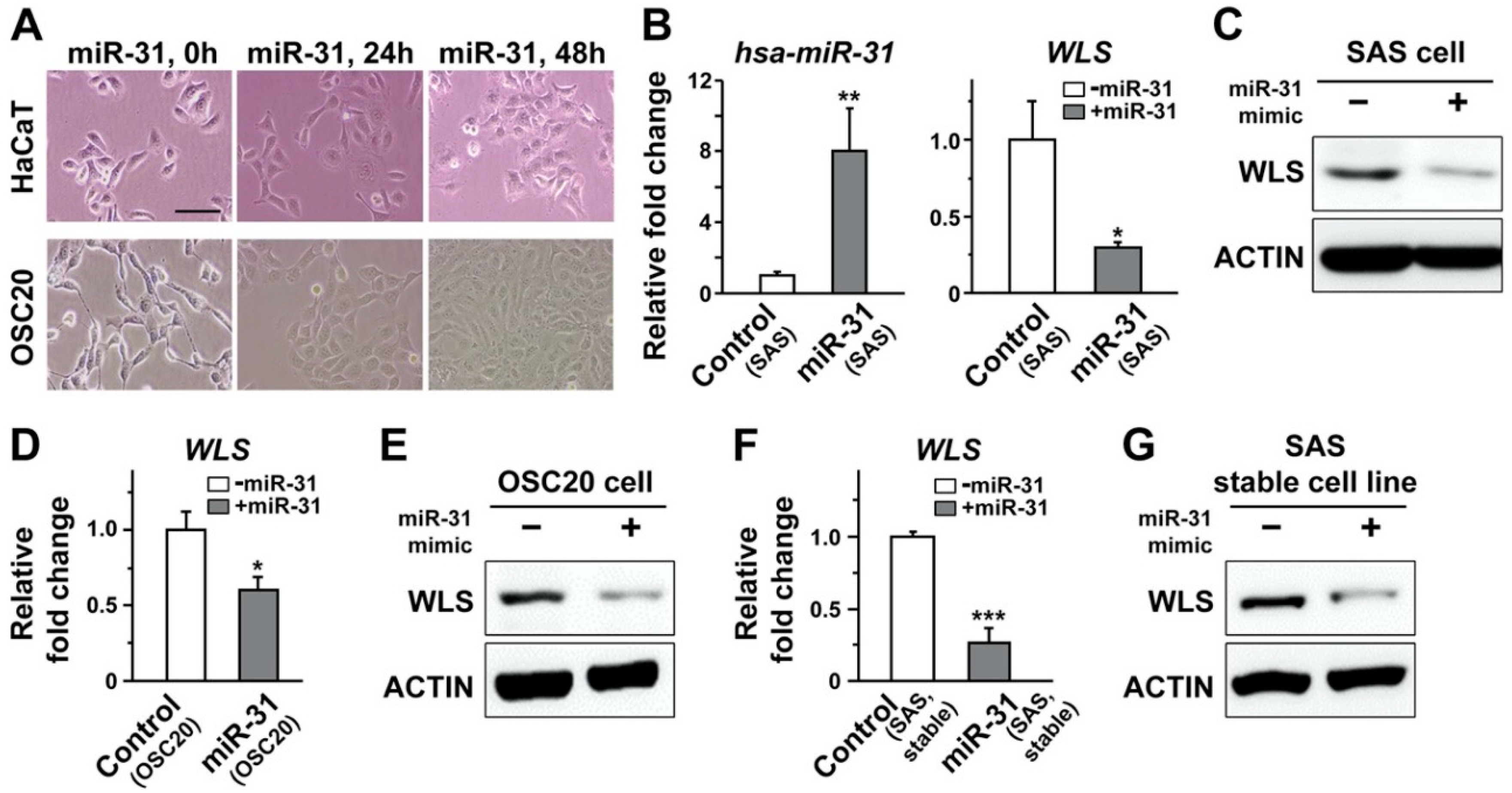
2.4. Overexpression of miR-31 in Human Oral Cancer Cells Induced Downregulation of Human WLS
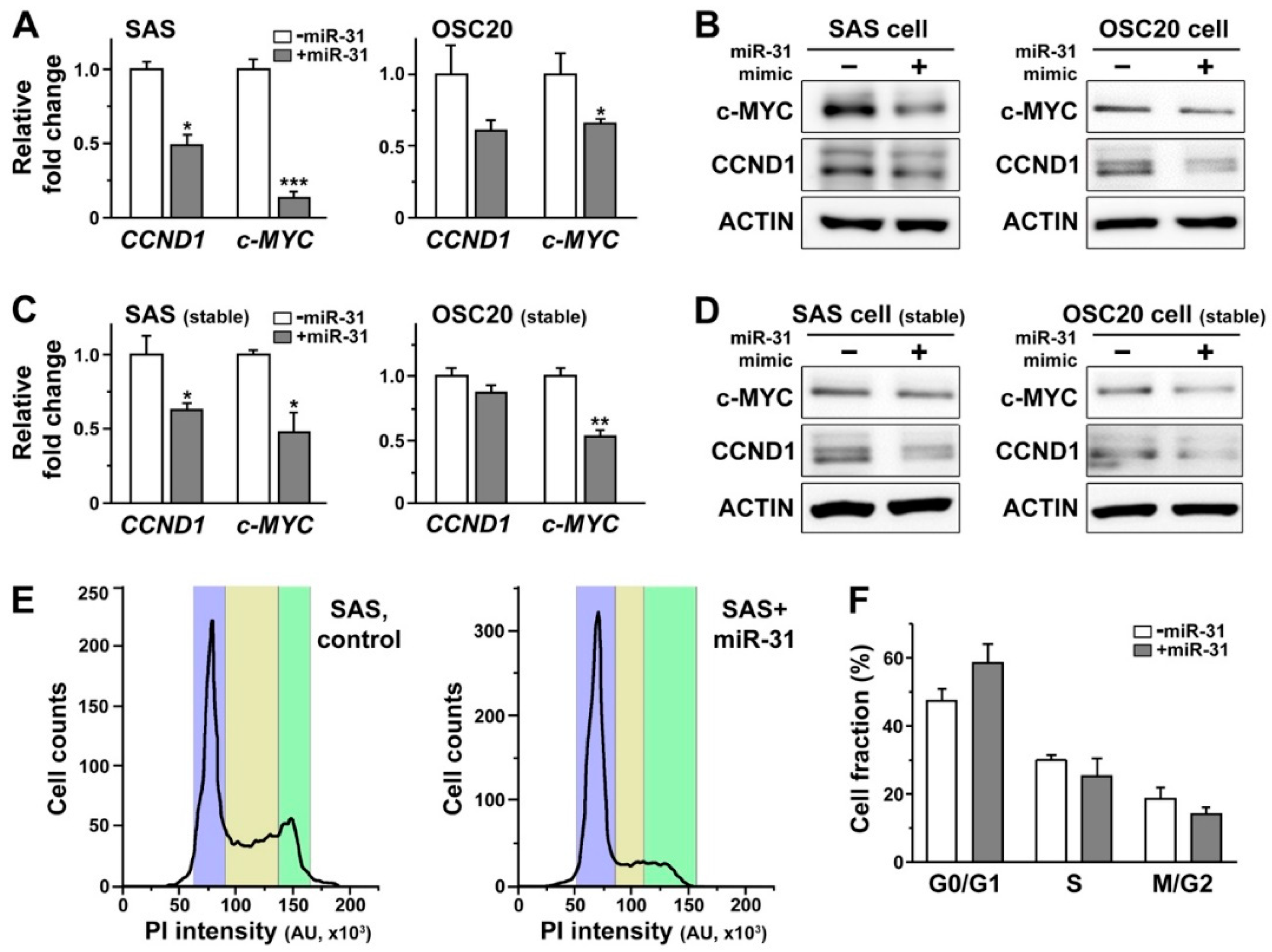
2.5. Overexpression of miR-31 in Human Oral Cancer Cells Induced Downregulation of Cyclin D1 and c-MYC
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fly Stocks
5.2. Image Analysis
5.3. Cell Culture
5.4. Transfections of miR-31 Mimic
5.5. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
5.6. Quantitative Measurement of miR-31 and Its Target Transcripts
5.7. Western Blot Analysis
5.8. Luciferase Assay
5.9. Flow Cytometry Analysis
5.10. Gene Expression and Survival Analysis of TCGA Collection
5.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| OSCC | oral squamous cell carcinoma |
| WLS | Wntless |
| miR-31 | microRNA-31 |
| CCND1 | cyclin D1 |
References
- Sharma, R.P. Wingless—A new mutant in D. melanogaster. Drosoph. Inf. Serv. 1973, 50, 134. [Google Scholar]
- Baker, N.E. Molecular cloning of sequences from wingless, a segment polarity gene in Drosophila: The spatial distribution of a transcript in embryos. EMBO J. 1987, 6, 1765–1773. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R.; Varmus, H.E. Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome. Cell 1982, 31, 99–109. [Google Scholar] [CrossRef]
- Rijsewijk, F.; Schuermann, M.; Wagenaar, E.; Parren, P.; Weigel, D.; Nusse, R. The Drosophila homolog of the mouse mammary oncogene int-1 is identical to the segment polarity gene wingless. Cell 1987, 50, 649–657. [Google Scholar] [CrossRef]
- Cadigan, K.M.; Nusse, R. Wnt signaling: A common theme in animal development. Genes Dev. 1997, 11, 3286–3305. [Google Scholar] [CrossRef] [Green Version]
- Willert, K.; Nusse, R. Wnt proteins. Cold Spring Harb. Perspect. Biol. 2012, 4, a007864. [Google Scholar] [CrossRef]
- Carreira-Barbosa, F.; Nunes, S.C. Wnt Signaling: Paths for Cancer Progression. Adv. Exp. Med. Biol. 2020, 1219, 189–202. [Google Scholar]
- Zhong, Z.; Yu, J.; Virshup, D.M.; Madan, B. Wnts and the hallmarks of cancer. Cancer Metastasis Rev. 2020, 39, 625–645. [Google Scholar] [CrossRef]
- Asem, M.S.; Buechler, S.; Wates, R.B.; Miller, D.L.; Stack, M.S. Wnt5a Signaling in Cancer. Cancers 2016, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, M.K.; Shao, C.; Wang, J.; Wei, Q.; Wang, X.; Collier, Z.; Tang, S.; Liu, H.; Zhang, F.; Huang, J.; et al. Wnt/beta-catenin signaling plays an ever-expanding role in stem cell self-renewal, tumorigenesis and cancer chemoresistance. Genes Dis. 2016, 3, 11–40. [Google Scholar] [CrossRef] [Green Version]
- Najdi, R.; Holcombe, R.F.; Waterman, M.L. Wnt signaling and colon carcinogenesis: Beyond APC. J. Carcinog 2011, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.D.; Mastriani, E.; Yan, Z.Q.; Yin, S.Y.; Zeng, Z.; Wang, H.; Li, Q.H.; Liu, H.Y.; Wang, X.Y.; Bao, H.X.; et al. SOX7 co-regulates Wnt/beta-catenin signaling with Axin-2: Both expressed at low levels in breast cancer. Sci. Rep. 2016, 6, 26136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguti, J.; DE MOURA, C.F.G.; Hossaka, T.A.; Franco, M.; Oshima, C.T.; Dedivitis, R.A.; Ribeiro, D.A. The role of canonical WNT signaling pathway in oral carcinogenesis: A comprehensive review. Anticancer Res. 2012, 32, 873–878. [Google Scholar] [PubMed]
- Shiah, S.G.; Shieh, Y.S.; Chang, J.Y. The Role of Wnt Signaling in Squamous Cell Carcinoma. J. Dent. Res. 2016, 95, 129–134. [Google Scholar] [CrossRef]
- Farooqi, A.A.; Shu, C.W.; Huang, H.W.; Wang, H.R.; Chang, Y.T.; Fayyaz, S.; Yuan, S.F.; Tang, J.Y.; Chang, H.W. TRAIL, Wnt, Sonic Hedgehog, TGFbeta, and miRNA Signalings Are Potential Targets for Oral Cancer Therapy. Int. J. Mol. Sci. 2017, 18, 1523. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.; Zheng, L.J.; Liu, J.Z.; Dobleman, T.; Hu, S.; Go, V.L.W.; Gao, G.; Xiao, G.G. MicroRNAs as effective surrogate biomarkers for early diagnosis of oral cancer. Clin. Oral Investig. 2018, 22, 571–581. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Scapoli, L.; Palmieri, A.; Lo Muzio, L.; Pezzetti, F.; Rubini, C.; Girardi, A.; Farinella, F.; Mazzotta, M.; Carinci, F. MicroRNA expression profiling of oral carcinoma identifies new markers of tumor progression. Int. J. Immunopathol. Pharm. 2010, 23, 1229–1234. [Google Scholar] [CrossRef]
- Gorenchtein, M.; Poh, C.F.; Saini, R.; Garnis, C. MicroRNAs in an oral cancer context—From basic biology to clinical utility. J. Dent. Res. 2012, 91, 440–446. [Google Scholar] [CrossRef]
- Perez-Sayans, M.; Pilar, G.D.; Barros-Angueira, F.; Suarez-Penaranda, J.M.; Fernandez, A.C.; Gandara-Rey, J.M.; Garcia-Garcia, A. Current trends in miRNAs and their relationship with oral squamous cell carcinoma. J. Oral Pathol. Med. 2012, 41, 433–443. [Google Scholar] [CrossRef]
- Kawakita, A.; Yanamoto, S.; Yamada, S.; Naruse, T.; Takahashi, H.; Kawasaki, G.; Umeda, M. MicroRNA-21 promotes oral cancer invasion via the Wnt/beta-catenin pathway by targeting DKK2. Pathol. Oncol. Res. 2014, 20, 253–261. [Google Scholar] [CrossRef]
- Min, A.; Zhu, C.; Peng, S.; Shuai, C.; Sun, L.; Han, Y.; Qian, Y.; Gao, S.; Su, T. Downregulation of Microrna-148a in Cancer-Associated Fibroblasts from Oral Cancer Promotes Cancer Cell Migration and Invasion by Targeting Wnt10b. J. Biochem. Mol. Toxicol. 2016, 30, 186–191. [Google Scholar] [CrossRef]
- Zhuang, Z.; Hu, F.; Hu, J.; Wang, C.; Hou, J.; Yu, Z.; Wang, T.T.; Liu, X.; Huang, H. MicroRNA-218 promotes cisplatin resistance in oral cancer via the PPP2R5A/Wnt signaling pathway. Oncol. Rep. 2017, 38, 2051–2061. [Google Scholar] [CrossRef]
- Chien, S.; Reiter, L.T.; Bier, E.; Gribskov, M. Homophila: Human disease gene cognates in Drosophila. Nucleic Acids Res. 2002, 30, 149–151. [Google Scholar] [CrossRef]
- Chang, C.J.; Hsu, C.C.; Chang, C.H.; Tsai, L.L.; Chang, Y.C.; Lu, S.W.; Yu, C.H.; Huang, H.S.; Wang, J.J.; Tsai, C.H.; et al. Let-7d functions as novel regulator of epithelial-mesenchymal transition and chemoresistant property in oral cancer. Oncol. Rep. 2011, 26, 1003–1010. [Google Scholar]
- Lu, Y.C.; Chen, Y.J.; Wang, H.M.; Tsai, C.Y.; Chen, W.H.; Huang, Y.C.; Fan, K.H.; Tsai, C.N.; Huang, S.F.; Kang, C.J.; et al. Oncogenic function and early detection potential of miRNA-10b in oral cancer as identified by microRNA profiling. Cancer Prev. Res. 2012, 5, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Tseng, H.H.; Tseng, Y.K.; You, J.J.; Kang, B.H.; Wang, T.H.; Yang, C.M.; Chen, H.C.; Liou, H.H.; Liu, P.F.; Ger, L.P.; et al. Next-generation Sequencing for microRNA Profiling: MicroRNA-21-3p Promotes Oral Cancer Metastasis. Anticancer Res. 2017, 37, 1059–1066. [Google Scholar]
- Hung, P.S.; Tu, H.F.; Kao, S.Y.; Yang, C.C.; Liu, C.J.; Huang, T.Y.; Chang, K.W.; Lin, S.C. miR-31 is upregulated in oral premalignant epithelium and contributes to the immortalization of normal oral keratinocytes. Carcinogenesis 2014, 35, 1162–1171. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.J.; Lin, S.C.; Yang, C.C.; Cheng, H.W.; Chang, K.W. Exploiting salivary miR-31 as a clinical biomarker of oral squamous cell carcinoma. Head Neck 2012, 34, 219–224. [Google Scholar] [CrossRef]
- Lu, W.C.; Kao, S.Y.; Yang, C.C.; Tu, H.F.; Wu, C.H.; Chang, K.W.; Lin, S.C. EGF up-regulates miR-31 through the C/EBPbeta signal cascade in oral carcinoma. PLoS ONE 2014, 9, e108049. [Google Scholar]
- Siow, M.Y.; Ng, L.P.; Vincent-Chong, V.K.; Jamaludin, M.; Abraham, M.T.; Abdul Rahman, Z.A.; Kallarakkal, T.G.; Yang, Y.H.; Cheong, S.C.; Zain, R.B. Dysregulation of miR-31 and miR-375 expression is associated with clinical outcomes in oral carcinoma. Oral Dis. 2014, 20, 345–351. [Google Scholar] [CrossRef]
- Li, T.; Li, L.; Li, D.; Wang, S.; Sun, J. MiR-34a inhibits oral cancer progression partially by repression of interleukin-6-receptor. Int. J. Clin. Exp. Pathol. 2015, 8, 1364–1373. [Google Scholar]
- Chen, W.; Yi, J.K.; Shimane, T.; Mehrazarin, S.; Lin, Y.L.; Shin, K.H.; Kim, R.H.; Park, N.H.; Kang, M.K. Grainyhead-like 2 regulates epithelial plasticity and stemness in oral cancer cells. Carcinogenesis 2016, 37, 500–510. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Zhang, L. MicroRNA-143 suppresses oral squamous cell carcinoma cell growth, invasion and glucose metabolism through targeting hexokinase 2. Biosci. Rep. 2017, 37, BSR20160404. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.C.; Chang, J.T.; Liao, C.T.; Kang, C.J.; Huang, S.F.; Chen, I.H.; Huang, C.C.; Huang, Y.C.; Chen, W.H.; Tsai, C.Y.; et al. OncomiR-196 promotes an invasive phenotype in oral cancer through the NME4-JNK-TIMP1-MMP signaling pathway. Mol. Cancer 2014, 13, 218. [Google Scholar] [CrossRef] [Green Version]
- Li, T.K.; Yin, K.; Chen, Z.; Bao, Y.; Zhang, S.X. MiR-214 regulates oral cancer KB cell apoptosis through targeting RASSF5. Genet Mol. Res. 2017, 16. [Google Scholar] [CrossRef]
- Peng, S.C.; Liao, C.T.; Peng, C.H.; Cheng, A.J.; Chen, S.J.; Huang, C.G.; Hsieh, W.P.; Yen, T.C. MicroRNAs MiR-218, MiR-125b, and Let-7g predict prognosis in patients with oral cavity squamous cell carcinoma. PLoS ONE 2014, 9, e102403. [Google Scholar] [CrossRef]
- Uesugi, A.; Kozaki, K.; Tsuruta, T.; Furuta, M.; Morita, K.; Imoto, I.; Omura, K.; Inazawa, J. The tumor suppressive microRNA miR-218 targets the mTOR component Rictor and inhibits AKT phosphorylation in oral cancer. Cancer Res. 2011, 71, 5765–5778. [Google Scholar] [CrossRef] [Green Version]
- Tachibana, H.; Sho, R.; Takeda, Y.; Zhang, X.; Yoshida, Y.; Narimatsu, H.; Otani, K.; Ishikawa, S.; Fukao, A.; Asao, H.; et al. Circulating miR-223 in Oral Cancer: Its Potential as a Novel Diagnostic Biomarker and Therapeutic Target. PLoS ONE 2016, 11, e0159693. [Google Scholar] [CrossRef]
- Xu, P.; Li, Y.; Zhang, H.; Li, M.; Zhu, H. MicroRNA-340 Mediates Metabolic Shift in Oral Squamous Cell Carcinoma by Targeting Glucose Transporter-1. J. Oral. Maxillofac. Surg. 2016, 74, 844–850. [Google Scholar] [CrossRef]
- Jung, H.M.; Patel, R.S.; Phillips, B.L.; Wang, H.; Cohen, D.M.; Reinhold, W.C.; Chang, L.J.; Yang, L.J.; Chan, E.K. Tumor suppressor miR-375 regulates MYC expression via repression of CIP2A coding sequence through multiple miRNA-mRNA interactions. Mol. Biol. Cell 2013, 24, 1638–1648. [Google Scholar] [CrossRef]
- Wiklund, E.D.; Gao, S.; Hulf, T.; Sibbritt, T.; Nair, S.; Costea, D.E.; Villadsen, S.B.; Bakholdt, V.; Bramsen, J.B.; Sorensen, J.A.; et al. MicroRNA alterations and associated aberrant DNA methylation patterns across multiple sample types in oral squamous cell carcinoma. PLoS ONE 2011, 6, e27840. [Google Scholar] [CrossRef]
- Liborio-Kimura, T.N.; Jung, H.M.; Chan, E.K. miR-494 represses HOXA10 expression and inhibits cell proliferation in oral cancer. Oral Oncol. 2015, 51, 151–157. [Google Scholar] [CrossRef]
- Ries, J.; Baran, C.; Wehrhan, F.; Weber, M.; Motel, C.; Kesting, M.; Nkenke, E. The altered expression levels of miR-186, miR-494 and miR-3651 in OSCC tissue vary from those of the whole blood of OSCC patients. Cancer Biomark 2019, 24, 19–30. [Google Scholar] [CrossRef]
- Bhatnagar, N.; Li, X.; Padi, S.K.; Zhang, Q.; Tang, M.S.; Guo, B. Downregulation of miR-205 and miR-31 confers resistance to chemotherapy-induced apoptosis in prostate cancer cells. Cell Death Dis. 2010, 1, e105. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Padi, S.K.; Tindall, D.J.; Guo, B. Polycomb protein EZH2 suppresses apoptosis by silencing the proapoptotic miR-31. Cell Death Dis. 2014, 5, e1486. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Sun, M.; Zhou, S.; Guo, B. Class I HDAC inhibitor mocetinostat induces apoptosis by activation of miR-31 expression and suppression of E2F6. Cell Death Discov. 2016, 2, 16036. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Huang, C.; Diao, X.; Fan, M.; Wang, P.; Xiao, Y.; Zhong, X.; Wu, R. Screening biomarkers of prostate cancer by integrating microRNA and mRNA microarrays. Genet Test Mol. Biomark. 2013, 17, 807–813. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, K.S.; Bae, H.J.; Eun, J.W.; Shen, Q.; Park, S.J.; Shin, W.C.; Yang, H.D.; Park, M.; Park, W.S.; et al. MicroRNA-31 functions as a tumor suppressor by regulating cell cycle and epithelial-mesenchymal transition regulatory proteins in liver cancer. Oncotarget 2015, 6, 8089–8102. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Han, C.; Zhang, Z.; Wang, L.; Xu, J. Increased expression of microRNA-31-5p inhibits cell proliferation, migration, and invasion via regulating Sp1 transcription factor in HepG2 hepatocellular carcinoma cell line. Biochem. Biophys. Res. Commun. 2017, 490, 371–377. [Google Scholar] [CrossRef]
- Hu, C.; Huang, F.; Deng, G.; Nie, W.; Huang, W.; Zeng, X. miR-31 promotes oncogenesis in intrahepatic cholangiocarcinoma cells via the direct suppression of RASA1. Exp. Med. 2013, 6, 1265–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karakatsanis, A.; Papaconstantinou, I.; Gazouli, M.; Lyberopoulou, A.; Polymeneas, G.; Voros, D. Expression of microRNAs, miR-21, miR-31, miR-122, miR-145, miR-146a, miR-200c, miR-221, miR-222, and miR-223 in patients with hepatocellular carcinoma or intrahepatic cholangiocarcinoma and its prognostic significance. Mol. Carcinog 2013, 52, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ma, J.; Zheng, J.; Wu, J.; Qu, C.; Sun, F.; Xu, S. MiR-31 Functions as a Tumor Suppressor in Lung Adenocarcinoma Mainly by Targeting HuR. Clin. Lab. 2016, 62, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.; Sun, B.; Jiang, Y.; Zheng, J.; Yang, N.; Ji, C.; Liang, Z.; Shi, J.; Zhang, R.; Liu, Y.; et al. MicroRNA-31 inhibits lung adenocarcinoma stem-like cells via down-regulation of MET-PI3K-Akt signaling pathway. Anticancer Agents Med. Chem. 2016, 16, 501–518. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, M.D.; Boyd, K.L.; Moyo, T.; Mitra, R.; Duszynski, R.; Arrate, M.P.; Chen, X.; Zhao, Z.; Blackwell, T.S.; Andl, T.; et al. MicroRNA-31 initiates lung tumorigenesis and promotes mutant KRAS-driven lung cancer. J. Clin. Investig. 2016, 126, 349–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, W.; Ye, Z.; Cui, R.; Perry, J.; Dedousi-Huebner, V.; Huebner, A.; Wang, Y.; Li, B.; Volinia, S.; Nakanishi, H.; et al. MicroRNA-31 predicts the presence of lymph node metastases and survival in patients with lung adenocarcinoma. Clin. Cancer Res. 2013, 19, 5423–5433. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Shi, J.; Wu, Y.; Xu, W.; Wang, Q.; Zhang, J.; Jiang, M.; Gu, G. Use of Luminex xMAP bead-based suspension array for detecting microRNA in NSCLC tissues and its clinical application. Tumori 2012, 98, 792–799. [Google Scholar] [CrossRef]
- Yu, M.; Liang, H.; Fu, Z.; Wang, X.; Liao, Z.; Zhou, Y.; Liu, Y.; Wang, Y.; Hong, Y.; Zhou, X.; et al. BAP1 suppresses lung cancer progression and is inhibited by miR-31. Oncotarget 2016, 7, 13742–13753. [Google Scholar] [CrossRef] [Green Version]
- Augoff, K.; McCue, B.; Plow, E.F.; Sossey-Alaoui, K. miR-31 and its host gene lncRNA LOC554202 are regulated by promoter hypermethylation in triple-negative breast cancer. Mol. Cancer 2012, 11. [Google Scholar] [CrossRef] [Green Version]
- Ge, F.; Wang, C.; Wang, W.; Liu, W.; Wu, B. MicroRNA-31 inhibits tumor invasion and metastasis by targeting RhoA in human gastric cancer. Oncol. Rep. 2017, 38, 1133–1139. [Google Scholar] [CrossRef] [Green Version]
- Korourian, A.; Roudi, R.; Shariftabrizi, A.; Madjd, Z. MicroRNA-31 inhibits RhoA-mediated tumor invasion and chemotherapy resistance in MKN-45 gastric adenocarcinoma cells. Exp. Biol. Med. 2017, 242, 1842–1847. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, X.; Liu, Y.; Ni, Z.; Lin, Y.; Duan, Z.; Shi, Y.; Wang, G.; Li, F. Downregulated miR-31 level associates with poor prognosis of gastric cancer and its restoration suppresses tumor cell malignant phenotypes by inhibiting E2F2. Oncotarget 2016, 7, 36577–36589. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.B.; Song, L.; Wen, H.J.; Bai, X.X.; Li, Z.J.; Ma, L.J. Upregulation of microRNA-31 targeting integrin alpha5 suppresses tumor cell invasion and metastasis by indirectly regulating PI3K/AKT pathway in human gastric cancer SGC7901 cells. Tumour Biol. 2016, 37, 8317–8325. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, J.; Li, D.; Xiao, B.; Miao, Y.; Jiang, Z.; Zhuo, H. Down-regulation of miR-31 expression in gastric cancer tissues and its clinical significance. Med. Oncol. 2010, 27, 685–689. [Google Scholar] [CrossRef]
- Geekiyanage, H.; Galanis, E. MiR-31 and miR-128 regulates poliovirus receptor-related 4 mediated measles virus infectivity in tumors. Mol. Oncol. 2016, 10, 1387–1403. [Google Scholar] [CrossRef] [Green Version]
- Hua, D.; Ding, D.; Han, X.; Zhang, W.; Zhao, N.; Foltz, G.; Lan, Q.; Huang, Q.; Lin, B. Human miR-31 targets radixin and inhibits migration and invasion of glioma cells. Oncol. Rep. 2012, 27, 700–706. [Google Scholar]
- Visani, M.; de Biase, D.; Marucci, G.; Cerasoli, S.; Nigrisoli, E.; Bacchi Reggiani, M.L.; Albani, F.; Baruzzi, A.; Pession, A. PERNO study group: Expression of 19 microRNAs in glioblastoma and comparison with other brain neoplasia of grades I-III. Mol. Oncol. 2014, 8, 417–430. [Google Scholar] [CrossRef]
- Asangani, I.A.; Harms, P.W.; Dodson, L.; Pandhi, M.; Kunju, L.P.; Maher, C.A.; Fullen, D.R.; Johnson, T.M.; Giordano, T.J.; Palanisamy, N.; et al. Genetic and epigenetic loss of microRNA-31 leads to feed-forward expression of EZH2 in melanoma. Oncotarget 2012, 3, 1011–1025. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, E.; Hershkovitz, L.; Itzhaki, O.; Hajdu, S.; Nemlich, Y.; Ortenberg, R.; Gefen, N.; Edry, L.; Modai, S.; Keisari, Y.; et al. Regulation of cancer aggressive features in melanoma cells by microRNAs. PLoS ONE 2011, 6, e18936. [Google Scholar] [CrossRef]
- Rokah, O.H.; Granot, G.; Ovcharenko, A.; Modai, S.; Pasmanik-Chor, M.; Toren, A.; Shomron, N.; Shpilberg, O. Downregulation of miR-31, miR-155, and miR-564 in chronic myeloid leukemia cells. PLoS ONE 2012, 7, e35501. [Google Scholar]
- Luo, X.; Burwinkel, B.; Tao, S.; Brenner, H. MicroRNA signatures: Novel biomarker for colorectal cancer? Cancer Epidemiol. Biomark. Prev. 2011, 20, 1272–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.S.; Wu, X.D.; Zhang, S.Q.; Li, C.F.; Yang, L.; Li, D.D.; Zhang, B.G.; Zhang, Y.; Jin, J.P.; Zhang, B. The tumor suppressor gene RhoBTB1 is a novel target of miR-31 in human colon cancer. Int. J. Oncol. 2013, 42, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.M.; Qian, J.C.; Deng, Z.L.; Cai, Z.; Tang, T.; Wang, P.; Zhang, K.H.; Cai, J.P. Expression of miR-21, miR-31, miR-96 and miR-135b is correlated with the clinical parameters of colorectal cancer. Oncol. Lett. 2012, 4, 339–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.H.; Yu, J.; Chen, N.; Wang, X.Y.; Liu, X.Y.; Wang, S.; Ding, Y.Q. Elevated microRNA-31 expression regulates colorectal cancer progression by repressing its target gene SATB2. PLoS ONE 2013, 8, e85353. [Google Scholar] [CrossRef] [Green Version]
- Earle, J.S.; Luthra, R.; Romans, A.; Abraham, R.; Ensor, J.; Yao, H.; Hamilton, S.R. Association of microRNA expression with microsatellite instability status in colorectal adenocarcinoma. J. Mol. Diagn. 2010, 12, 433–440. [Google Scholar] [CrossRef]
- Tateishi, Y.; Okudela, K.; Mitsui, H.; Umeda, S.; Suzuki, T.; Kojima, Y.; Watanabe, K.; Kawano, N.; Endo, I.; Ohashi, K. The potential role of microRNA-31 expression in early colorectal cancer. Pathol. Int. 2015, 65, 513–518. [Google Scholar] [CrossRef]
- Wang, N.; Li, Y.; Zhou, J. miR-31 Functions as an Oncomir Which Promotes Epithelial-Mesenchymal Transition via Regulating BAP1 in Cervical Cancer. BioMed Res. Int. 2017, 2017, 6361420. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Zhou, Y.; Zheng, L.; Li, H. MiR-31 is an independent prognostic factor and functions as an oncomir in cervical cancer via targeting ARID1A. Gynecol. Oncol. 2014, 134, 129–137. [Google Scholar] [CrossRef]
- Zheng, W.; Liu, Z.; Zhang, W.; Hu, X. miR-31 functions as an oncogene in cervical cancer. Arch. Gynecol. Obs. 2015, 292, 1083–1089. [Google Scholar] [CrossRef]
- Kao, S.Y.; Tsai, M.M.; Wu, C.H.; Chen, J.J.; Tseng, S.H.; Lin, S.C.; Chang, K.W. Co-targeting of multiple microRNAs on factor-Inhibiting hypoxia-Inducible factor gene for the pathogenesis of head and neck carcinomas. Head Neck 2016, 38, 522–528. [Google Scholar] [CrossRef]
- Liu, S.G.; Qin, X.G.; Zhao, B.S.; Qi, B.; Yao, W.J.; Wang, T.Y.; Li, H.C.; Wu, X.N. Differential expression of miRNAs in esophageal cancer tissue. Oncol. Lett. 2013, 5, 1639–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suresh, R.; Sethi, S.; Ali, S.; Giorgadze, T.; Sarkar, F.H. Differential Expression of MicroRNAs in Papillary Thyroid Carcinoma and Their Role in Racial Disparity. J. Cancer Sci. 2015, 7, 145–154. [Google Scholar]
- Yabushita, S.; Fukamachi, K.; Tanaka, H.; Sumida, K.; Deguchi, Y.; Sukata, T.; Kawamura, S.; Uwagawa, S.; Suzui, M.; Tsuda, H. Circulating microRNAs in serum of human K-ras oncogene transgenic rats with pancreatic ductal adenocarcinomas. Pancreas 2012, 41, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal 2013, 6, l1. [Google Scholar] [CrossRef] [Green Version]
- Soga, D.; Yoshiba, S.; Shiogama, S.; Miyazaki, H.; Kondo, S.; Shintani, S. microRNA expression profiles in oral squamous cell carcinoma. Oncol. Rep. 2013, 30, 579–583. [Google Scholar] [CrossRef] [Green Version]
- Herranz, H.; Milan, M. Signalling molecules, growth regulators and cell cycle control in Drosophila. Cell Cycle 2008, 7, 3335–3337. [Google Scholar] [CrossRef]
- Hartl, T.A.; Scott, M.P. Wing tips: The wing disc as a platform for studying Hedgehog signaling. Methods 2014, 68, 199–206. [Google Scholar] [CrossRef]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 1993, 118, 401–415. [Google Scholar]
- Banziger, C.; Soldini, D.; Schutt, C.; Zipperlen, P.; Hausmann, G.; Basler, K. Wntless, a conserved membrane protein dedicated to the secretion of Wnt proteins from signaling cells. Cell 2006, 125, 509–522. [Google Scholar] [CrossRef] [Green Version]
- Bartscherer, K.; Pelte, N.; Ingelfinger, D.; Boutros, M. Secretion of Wnt ligands requires Evi, a conserved transmembrane protein. Cell 2006, 125, 523–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malka, Y.; Steiman-Shimony, A.; Rosenthal, E.; Argaman, L.; Cohen-Daniel, L.; Arbib, E.; Margalit, H.; Kaplan, T.; Berger, M. Post-transcriptional 3′-UTR cleavage of mRNA transcripts generates thousands of stable uncapped autonomous RNA fragments. Nat. Commun. 2017, 8, 2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Wei, D.; Wang, W.; Shen, B.; Xu, S.; Cao, Y. TRAF4 enhances oral squamous cell carcinoma cell growth, invasion and migration by Wnt-beta-catenin signaling pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 11837–11846. [Google Scholar] [PubMed]
- Liao, D.J.; Thakur, A.; Wu, J.; Biliran, H.; Sarkar, F.H. Perspectives on c-Myc, Cyclin D1, and their interaction in cancer formation, progression, and response to chemotherapy. Crit. Rev. Oncog. 2007, 13, 93–158. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- De Angelis, R.; Sant, M.; Coleman, M.P.; Francisci, S.; Baili, P.; Pierannunzio, D.; Trama, A.; Visser, O.; Brenner, H.; Ardanaz, E.; et al. Cancer survival in Europe 1999-2007 by country and age: Results of EUROCARE--5-a population-based study. Lancet Oncol. 2014, 15, 23–34. [Google Scholar] [CrossRef]
- Hussein, A.A.; Helder, M.N.; de Visscher, J.G.; Leemans, C.R.; Braakhuis, B.J.; de Vet, H.C.W.; Forouzanfar, T. Global incidence of oral and oropharynx cancer in patients younger than 45 years versus older patients: A systematic review. Eur. J. Cancer 2017, 82, 115–127. [Google Scholar] [CrossRef]
- Lekka, E.; Hall, J. Noncoding RNAs in disease. FEBS Lett. 2018, 592, 2884–2900. [Google Scholar] [CrossRef]
- Lujambio, A.; Lowe, S.W. The microcosmos of cancer. Nature 2012, 482, 347–355. [Google Scholar] [CrossRef]
- Pinzon, N.; Li, B.; Martinez, L.; Sergeeva, A.; Presumey, J.; Apparailly, F.; Seitz, H. microRNA target prediction programs predict many false positives. Genome Res. 2017, 27, 234–245. [Google Scholar] [CrossRef] [Green Version]
- Carthew, R.W.; Agbu, P.; Giri, R. MicroRNA function in Drosophila melanogaster. Semin Cell Dev. Biol. 2017, 65, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loganantharaj, R.; Randall, T.A. The Limitations of Existing Approaches in Improving MicroRNA Target Prediction Accuracy. Methods Mol. Biol. 2017, 1617, 133–158. [Google Scholar] [PubMed]
- Xiao, W.; Bao, Z.X.; Zhang, C.Y.; Zhang, X.Y.; Shi, L.J.; Zhou, Z.T.; Jiang, W.W. Upregulation of miR-31* is negatively associated with recurrent/newly formed oral leukoplakia. PLoS ONE 2012, 7, e38648. [Google Scholar] [CrossRef] [PubMed]
- Shiah, S.G.; Hsiao, J.R.; Chang, W.M.; Chen, Y.W.; Jin, Y.T.; Wong, T.Y.; Huang, J.S.; Tsai, S.T.; Hsu, Y.M.; Chou, S.T.; et al. Downregulated miR329 and miR410 promote the proliferation and invasion of oral squamous cell carcinoma by targeting Wnt-7b. Cancer Res. 2014, 74, 7560–7572. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA ID | Expression Profile of miRNAs |
|---|---|
| hsa-let-7d | Downregulated [25] |
| hsa-miR-10b | Upregulated [26] |
| hsa-miR-21 | Upregulated [21,27] |
| hsa-miR-31 | Upregulated [28,29,30,31] |
| hsa-miR-34a | Downregulated [32] |
| hsa-miR-141/200/429 | Downregulated [33] |
| hsa-miR-143 | Downregulated [34] |
| hsa-miR-196b | Upregulated [35] |
| hsa-miR-214 | Upregulated [36] |
| hsa-miR-218 | Downregulated [37,38] |
| hsa-miR-223 | Upregulated [39] |
| hsa-miR-340 | Downregulated [40] |
| hsa-miR-375 | Downregulated [41,42] |
| hsa-miR-494 | Downregulated [43,44] |
| Family/Cluster Cancer | Expression Profile of miR-31 |
|---|---|
| Prostate cancer | Downregulated [45,46,47] |
| Upregulated [48] | |
| Liver cancer | Downregulated [49,50] |
| Upregulated [51,52] | |
| Lung cancer | Downregulated [53,54] |
| Upregulated [55,56,57,58] | |
| Breast cancer | Downregulated [59] |
| Gastric cancer | Downregulated [60,61,62,63,64] |
| Glioblastoma | Downregulated [65,66,67] |
| Melanoma | Downregulated [68,69] |
| Leukemia | Downregulated [70] |
| Colon/Colorectal cancer | Upregulated [71,72,73,74,75,76] |
| Cervical cancer | Upregulated [77,78,79] |
| Head and neck cancer | Upregulated [80] |
| Esophageal cancer | Upregulated [81] |
| Thyroid cancer | Upregulated [82] |
| Pancreatic cancer | Upregulated [83] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, J.E.; Lee, J.Y.; Kim, I.R.; Park, S.M.; Kang, J.W.; Kim, Y.H.; Park, H.R.; Lee, J.H. MicroRNA-31 Regulates Expression of Wntless in Both Drosophila melanogaster and Human Oral Cancer Cells. Int. J. Mol. Sci. 2020, 21, 7232. https://doi.org/10.3390/ijms21197232
Jung JE, Lee JY, Kim IR, Park SM, Kang JW, Kim YH, Park HR, Lee JH. MicroRNA-31 Regulates Expression of Wntless in Both Drosophila melanogaster and Human Oral Cancer Cells. International Journal of Molecular Sciences. 2020; 21(19):7232. https://doi.org/10.3390/ijms21197232
Chicago/Turabian StyleJung, Ji Eun, Joo Young Lee, In Ryoung Kim, Sang Mee Park, Ji Wan Kang, Yun Hak Kim, Hae Ryoun Park, and Ji Hye Lee. 2020. "MicroRNA-31 Regulates Expression of Wntless in Both Drosophila melanogaster and Human Oral Cancer Cells" International Journal of Molecular Sciences 21, no. 19: 7232. https://doi.org/10.3390/ijms21197232