Parallelized Manipulation of Adherent Living Cells by Magnetic Nanoparticles-Mediated Forces

 , , , , , ,
, , , , , ,  , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
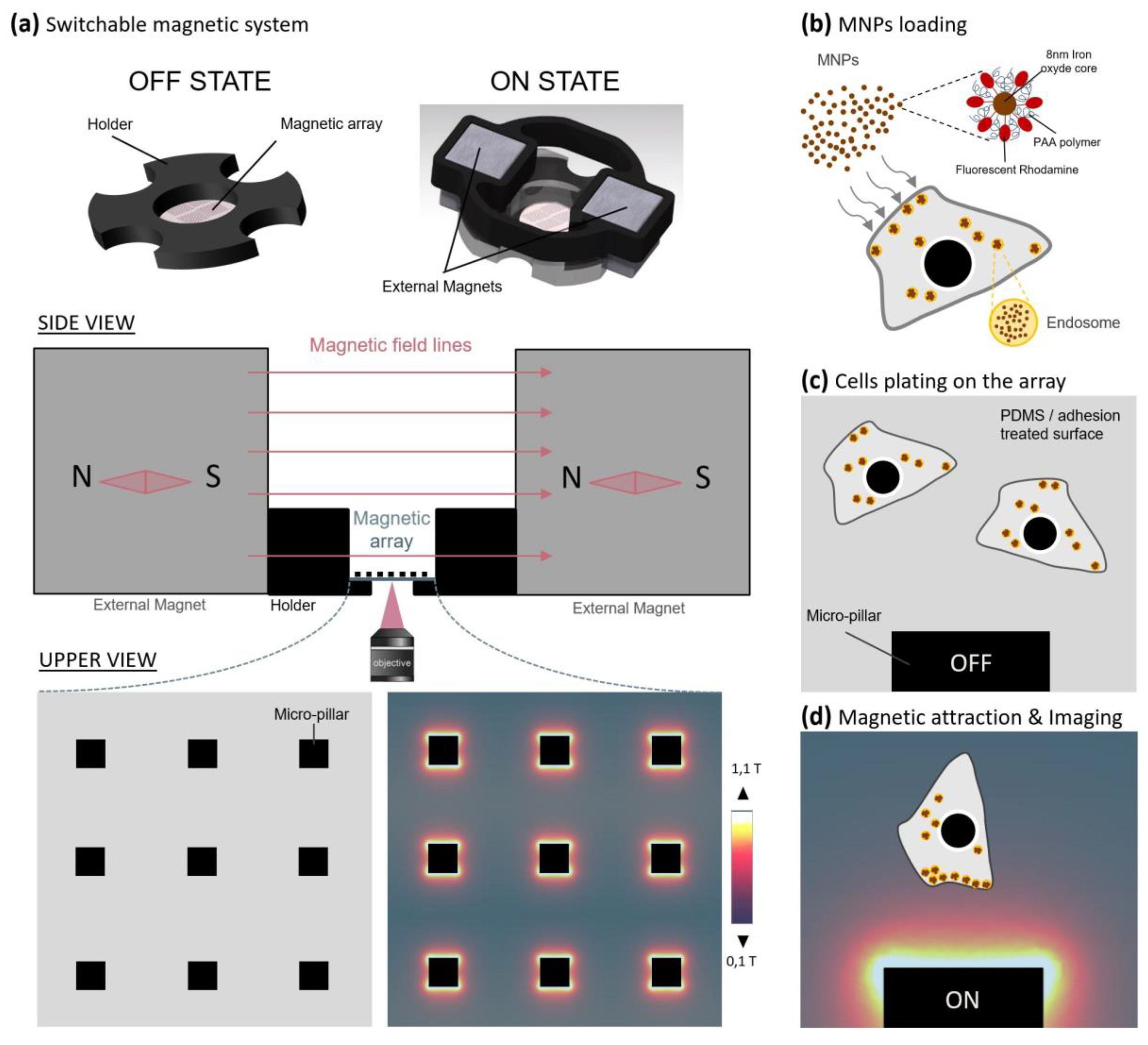
2.1. A Parallelized Magnetic Tool to Actuate Migration and Neurite Outgrowth through Forces
2.2. MNPs Are Stable in Lysosomal Compartment up to Several Days
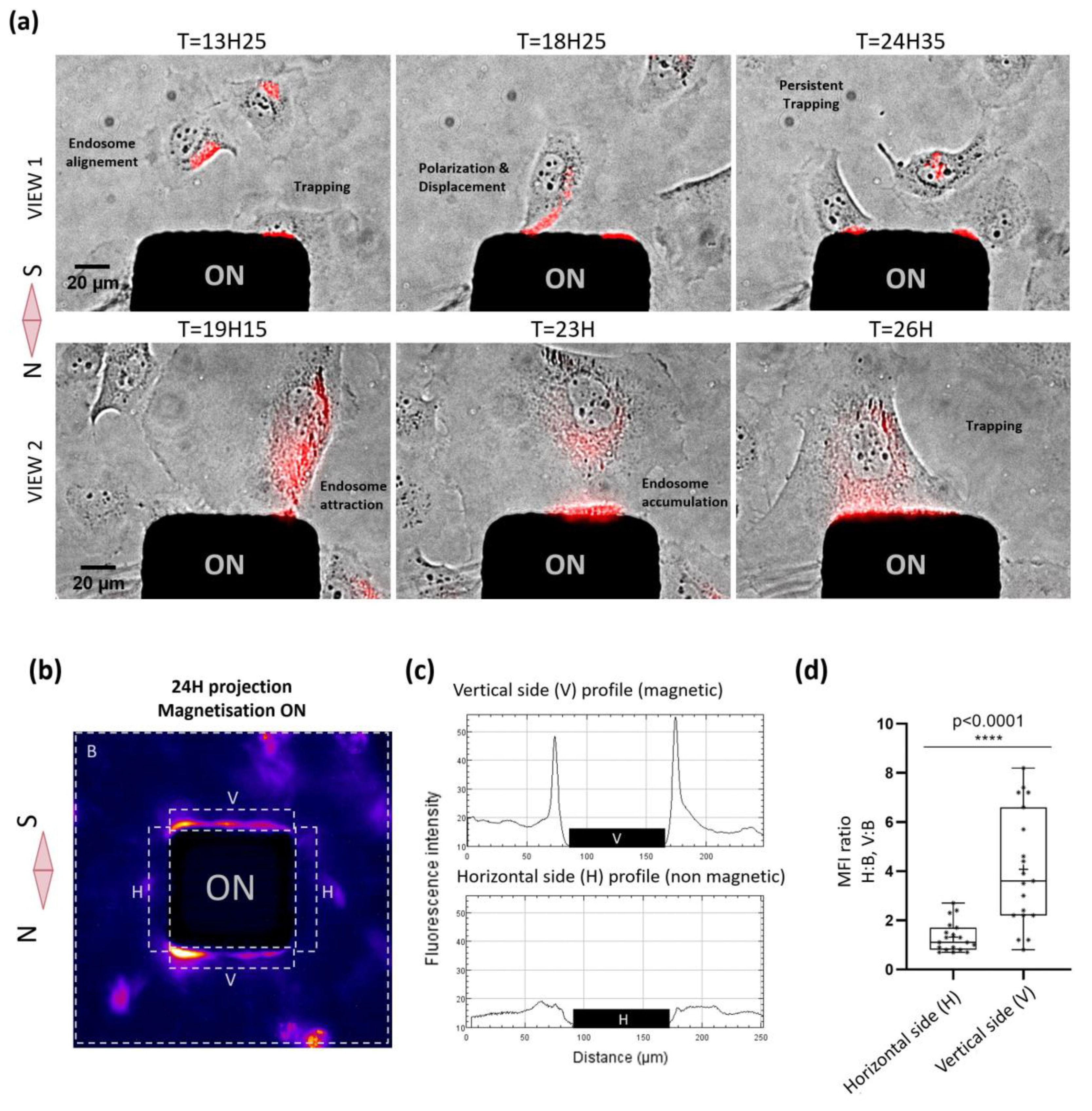
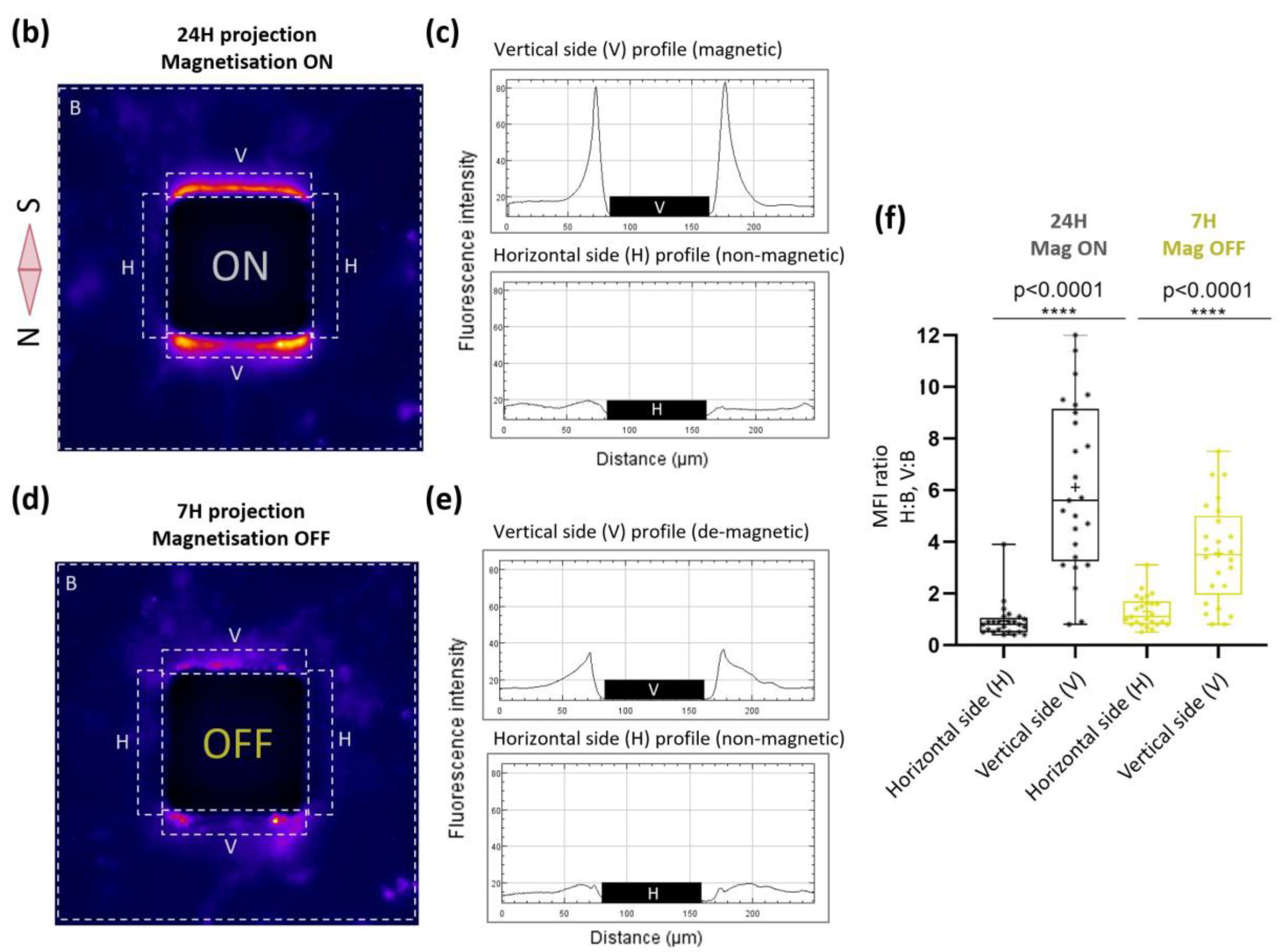
2.3. Nanoparticle-Mediated Forces Direct Migration of Adherent Cells and Can Trap Them
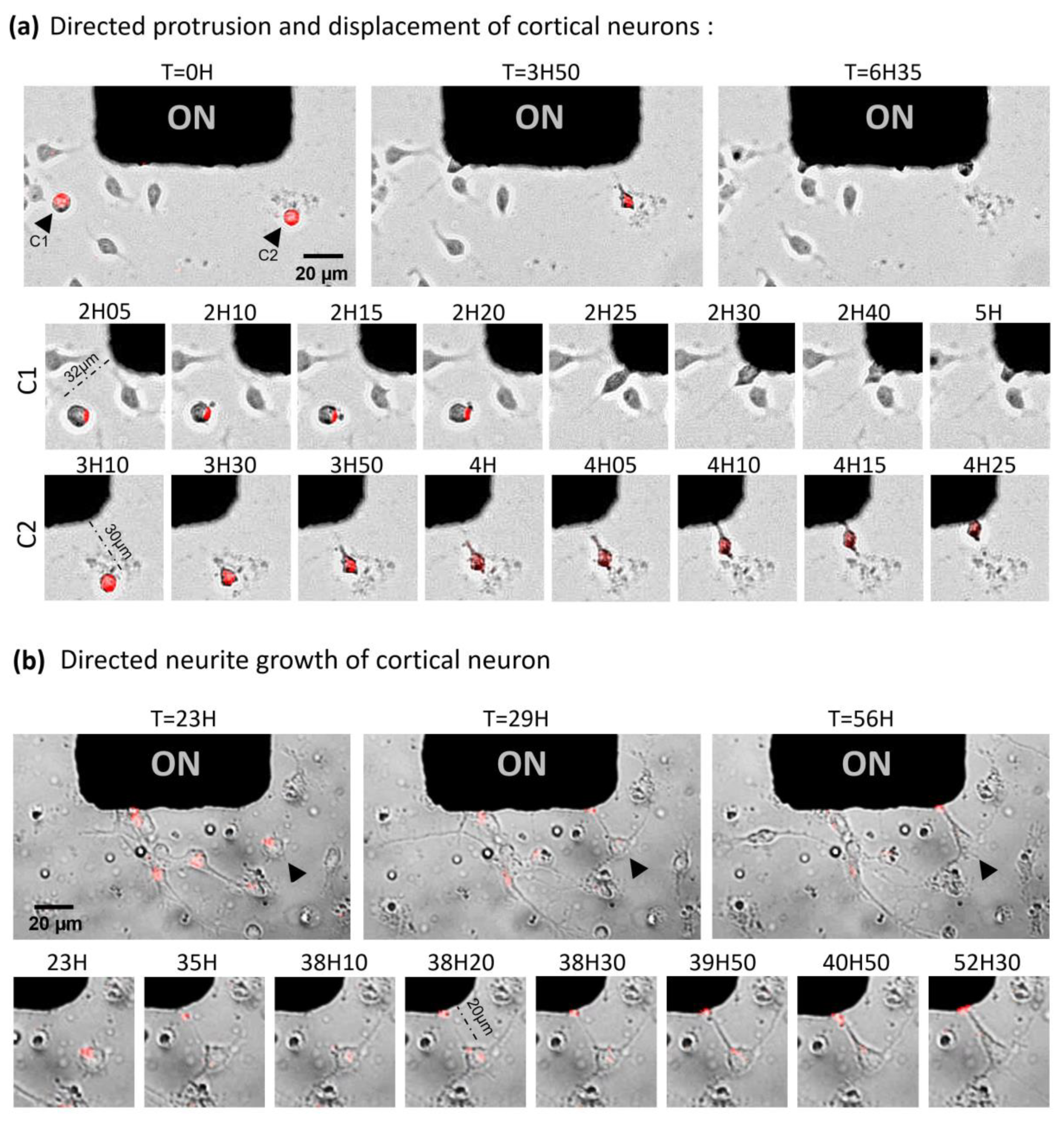
2.4. Toward the Parallelized Magnetic Manipulation of Neurite Outgrowth
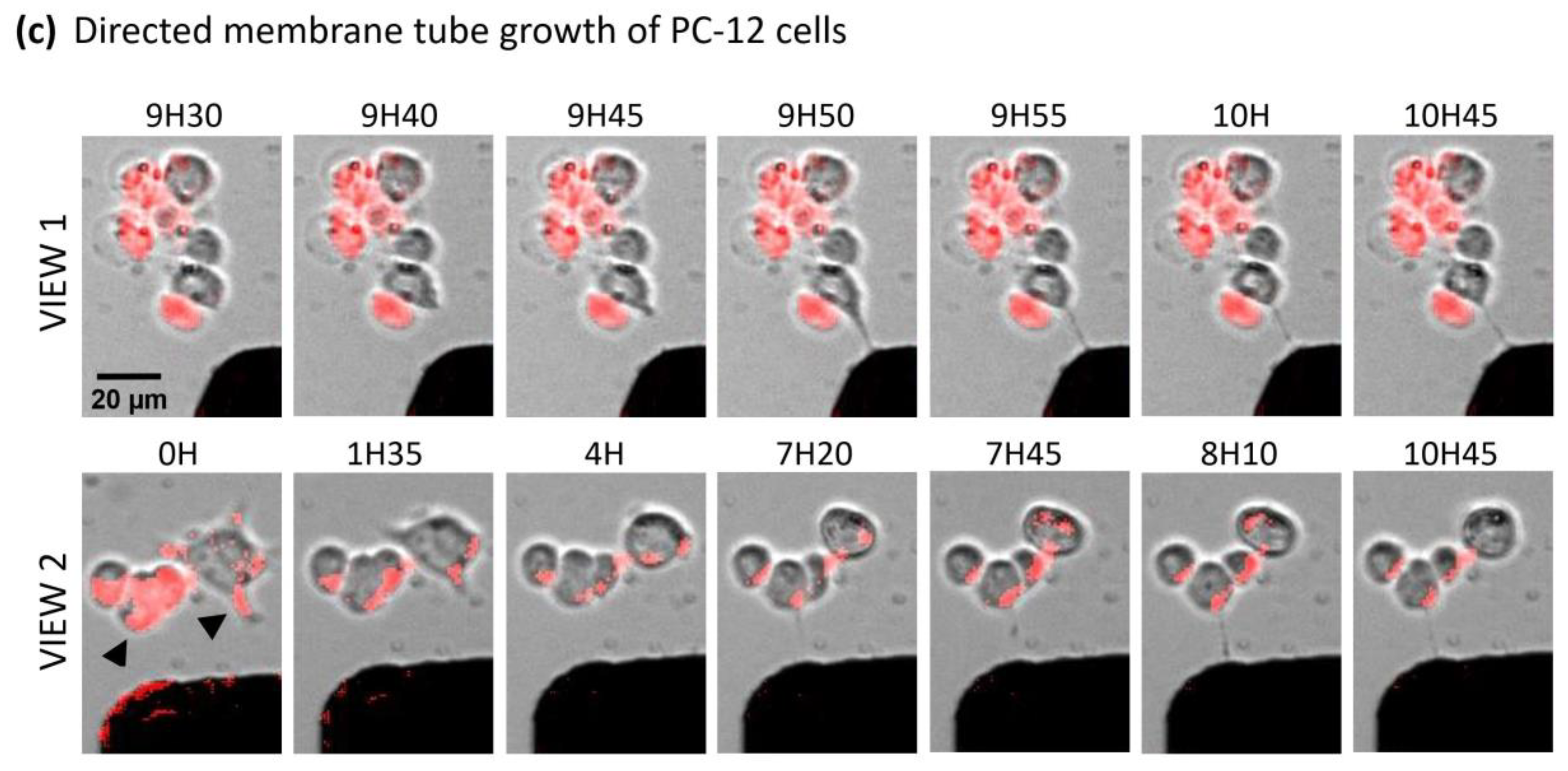
2.5. Magnetic Endosomes Can Induce and Direct Neurite-Like Tubes from Pc12 Cells but Cannot Guide Preexisting Neurites
3. Discussion
4. Materials and Methods
4.1. Synthesis of Fe2O3-PAA2k-Rhodamine Magnetic Nanoparticles (MNPs)
4.2. Characterization of MNPs
4.3. Cell Culture
4.4. Cell Transfection for Endosome Labeling
4.5. MNP Loading by Passive Endocytosis
4.6. Micro-Magnetic Array Fabrication and Characterization
4.7. Magnetic Control Experiments on Micro-Magnetic Array (MMA)
4.8. Image Analysis and Statistic Evaluation
4.9. Force Calculation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schöneborn, H.; Raudzus, F.; Coppey, M.; Neumann, S.; Heumann, R. Perspectives of RAS and RHEB GTPase Signaling Pathways in Regenerating Brain Neurons. Int. J. Mol. Sci. 2018, 19, 4052. [Google Scholar] [CrossRef] [Green Version]
- Karunarathne, A.; Giri, L.; Patel, A.K.; Venkatesh, K.V.; Gautam, N. Optical control demonstrates switch-like PIP3 dynamics underlying the initiation of immune cell migration. Proc. Natl. Acad. Sci. USA 2013, 110, E1575–E1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, P.R.; Gautam, N. Subcellular optogenetic inhibition of G proteins generates signaling gradients and cell migration. Mol. Boil. Cell 2014, 25, 2305–2314. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, P.R.; Kalyanaraman, V.; Gautam, N. Subcellular optogenetic activation of Cdc42 controls local and distal signaling to drive immune cell migration. Mol. Boil. Cell 2016, 27, 1442–1450. [Google Scholar] [CrossRef] [PubMed]
- Valon, L.; Etoc, F.; Remorino, A.; Di Pietro, F.; Morin, X.; Dahan, M.; Coppey, M. Predictive Spatiotemporal Manipulation of Signaling Perturbations Using Optogenetics. Biophys. J. 2015, 109, 1785–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunarathne, A.; Giri, L.; Kalyanaraman, V.; Gautam, N. Optically triggering spatiotemporally confined GPCR activity in a cell and programming neurite initiation and extension. Proc. Natl. Acad. Sci. USA 2013, 110, E1565–E1574. [Google Scholar] [CrossRef] [Green Version]
- Endo, M.; Hattori, M.; Toriyabe, H.; Ohno, H.; Kamiguchi, H.; Iino, Y.; Ozawa, T. Optogenetic activation of axon guidance receptors controls direction of neurite outgrowth. Sci. Rep. 2016, 6, 23976. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Weitemier, A.; Zeng, X.; He, L.; Wang, X.; Tao, Y.; Huang, A.J.Y.; Hashimotodani, Y.; Kano, M.; Iwasaki, H.; et al. Near-infrared deep brain stimulation via upconversion nanoparticle–mediated optogenetics. Science 2018, 359, 679–684. [Google Scholar] [CrossRef] [Green Version]
- Monzel, C.; Vicario, C.; Piehler, J.; Coppey, M.; Dahan, M. Magnetic control of cellular processes using biofunctional nanoparticles. Chem. Sci. 2017, 8, 7330–7338. [Google Scholar] [CrossRef] [Green Version]
- Liße, D.; Monzel, C.; Vicario, C.; Manzi, J.; Maurin, I.; Coppey, M.; Piehler, J.; Dahan, M. Engineered Ferritin for Magnetogenetic Manipulation of Proteins and Organelles Inside Living Cells. Adv. Mater. 2017, 29, 1700189. [Google Scholar] [CrossRef]
- Etoc, F.; Vicario, C.; Lisse, D.; Siaugue, J.-M.; Piehler, J.; Coppey, M.; Dahan, M. Magnetogenetic Control of Protein Gradients Inside Living Cells with High Spatial and Temporal Resolution. Nano Lett. 2015, 15, 3487–3494. [Google Scholar] [CrossRef]
- Jin, Y.; Lee, J.-U.; Chung, E.; Yang, K.; Kim, J.; Kim, J.-W.; Lee, J.S.; Cho, A.-N.; Oh, T.; Lee, J.-H.; et al. Magnetic Control of Axon Navigation in Reprogrammed Neurons. Nano Lett. 2019, 19, 6517–6523. [Google Scholar] [CrossRef] [PubMed]
- Schöneborn, H.; Raudzus, F.; Secret, E.; Otten, N.; Michel, A.; Fresnais, J.; Menager, C.; Siaugue, J.-M.; Zaehres, H.; Dietzel, I.; et al. Novel Tools towards Magnetic Guidance of Neurite Growth: (I) Guidance of Magnetic Nanoparticles into Neurite Extensions of Induced Human Neurons and In Vitro Functionalization with RAS Regulating Proteins. J. Funct. Biomater. 2019, 10, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gahl, T.J.; Kunze, A. Force-Mediating Magnetic Nanoparticles to Engineer Neuronal Cell Function. Front. Mol. Neurosci. 2018, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Svitkina, T.M. The Actin Cytoskeleton and Actin-Based Motility. Cold Spring Harb. Perspect. Biol. 2018, 10, a018267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamoureux, P.; Buxbaum, R.E.; Heidemann, S.R. Direct evidence that growth cones pull. Nature 1989, 340, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Suter, D.M.; Miller, K.E. The emerging role of forces in axonal elongation. Prog. Neurobiol. 2011, 94, 91–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, C.-M.; Wang, H.-B.; Dembo, M.; Wang, Y.L. Cell Movement Is Guided by the Rigidity of the Substrate. Biophys. J. 2000, 79, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Isenberg, B.C.; DiMilla, P.A.; Walker, M.; Kim, S.; Wong, J.Y. Vascular Smooth Muscle Cell Durotaxis Depends on Substrate Stiffness Gradient Strength. Biophys. J. 2009, 97, 1313–1322. [Google Scholar] [CrossRef] [Green Version]
- Lamoureux, P.; Ruthel, G.; Buxbaum, R.E.; Heidemann, S.R. Mechanical tension can specify axonal fate in hippocampal neurons. J. Cell Biol. 2002, 159, 499–508. [Google Scholar] [CrossRef]
- Schnell, E.; Klinkhammer, K.; Balzer, S.; Brook, G.; Klee, D.; Dalton, P.; Mey, J. Guidance of glial cell migration and axonal growth on electrospun nanofibers of poly-ε-caprolactone and a collagen/poly-ε-caprolactone blend. Biomaterials 2007, 28, 3012–3025. [Google Scholar] [CrossRef] [PubMed]
- Berns, E.J.; Sur, S.; Pan, L.; Goldberger, J.E.; Suresh, S.; Zhang, S.; Kessler, J.A.; Stupp, S.I. Aligned neurite outgrowth and directed cell migration in self-assembled monodomain gels. Biomaterials 2014, 35, 185–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liverani, C.; Mercatali, L.; Cristofolini, L.; Giordano, E.; Minardi, S.; Della Porta, G.; De Vita, A.; Miserocchi, G.; Spadazzi, C.; Tasciotti, E.; et al. Investigating the Mechanobiology of Cancer Cell-ECM Interaction Through Collagen-Based 3D Scaffolds. Cell. Mol. Bioeng. 2017, 10, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.C.; Cámara-Torres, M.; Rahimi, K.; Köhler, J.; Möller, M.; De Laporte, L. Nerve Cells Decide to Orient inside an Injectable Hydrogel with Minimal Structural Guidance. Nano Lett. 2017, 17, 3782–3791. [Google Scholar] [CrossRef] [PubMed]
- Antman-Passig, M.; Shefi, O. Remote Magnetic Orientation of 3D Collagen Hydrogels for Directed Neuronal Regeneration. Nano Lett. 2016, 16, 2567–2573. [Google Scholar] [CrossRef]
- Tseng, P.; Judy, J.W.; Di Carlo, D. Magnetic nanoparticle–mediated massively parallel mechanical modulation of single-cell behavior. Nat. Methods 2012, 9, 1113–1119. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, M.; Clemons, T.D.; Ho, D.; Gutierrez, L.; Lázaro, F.J.; House, M.J.; Pierre, T.G.S.; Fear, M.W.; Wood, F.M.; Iyer, K.S. Manipulating directional cell motility using intracellular superparamagnetic nanoparticles. Nanoscale 2015, 7, 4884–4889. [Google Scholar] [CrossRef] [Green Version]
- White, E.E.; Pai, A.; Weng, Y.; Suresh, A.K.; Van Haute, D.; Pailevanian, T.; Alizadeh, D.; Hajimiri, A.; Badie, B.; Berlin, J.M. Functionalized iron oxide nanoparticles for controlling the movement of immune cells. Nanoscale 2015, 7, 7780–7789. [Google Scholar] [CrossRef] [Green Version]
- Alon, N.; Havdala, T.; Skaat, H.; Baranes, K.; Levy, I.; Margel, S.; Shefi, O.; Marcus, M.; Sharoni, A. Magnetic micro-device for manipulating PC12 cell migration and organization. Lab Chip 2015, 15, 2030–2036. [Google Scholar] [CrossRef]
- Bing, X.; Huang, L.; Zhu, L.; Liu, Z.; Ma, T.; Zhu, S.; Huang, J.; Luo, Z.; Xia, B. Manipulation of Schwann cell migration across the astrocyte boundary by polysialyltransferase-loaded superparamagnetic nanoparticles under magnetic field. Int. J. Nanomed. 2016, 11, 6727–6741. [Google Scholar] [CrossRef] [Green Version]
- Fass, J.N.; Odde, D.J. Tensile Force-Dependent Neurite Elicitation via Anti-β1 Integrin Antibody-Coated Magnetic Beads. Biophys. J. 2003, 85, 623–636. [Google Scholar] [CrossRef] [Green Version]
- Kunze, A.; Tseng, P.; Godzich, C.; Murray, C.; Caputo, A.; Schweizer, F.E.; Di Carlo, D. Engineering Cortical Neuron Polarity with Nanomagnets on a Chip. ACS Nano 2015, 9, 3664–3676. [Google Scholar] [CrossRef] [PubMed]
- Riggio, C.; Calatayud, M.P.; Giannaccini, M.; Sanz, B.; Torres, T.E.; Fernández-Pacheco, R.; Ripoli, A.; Ibarra, R.; Dente, L.; Cuschieri, A.; et al. The orientation of the neuronal growth process can be directed via magnetic nanoparticles under an applied magnetic field. Nanomed. Nanotechnol. Biol. Med. 2014, 10, 1549–1558. [Google Scholar] [CrossRef] [PubMed]
- Raffa, V.; Falcone, F.; De Vincentiis, S.; Falconieri, A.; Calatayud, M.P.; Goya, G.; Cuschieri, A.; De Vincentis, S. Piconewton Mechanical Forces Promote Neurite Growth. Biophys. J. 2018, 115, 2026–2033. [Google Scholar] [CrossRef] [Green Version]
- Marcus, M.; Karni, M.; Baranes, K.; Levy, I.; Alon, N.; Margel, S.; Shefi, O. Iron oxide nanoparticles for neuronal cell applications: Uptake study and magnetic manipulations. J. Nanobiotechnol. 2016, 14, 37. [Google Scholar] [CrossRef] [Green Version]
- Toraille, L.; Aïzel, K.; Balloul, E.; Vicario, C.; Monzel, C.; Coppey, M.; Secret, E.; Siaugue, J.-M.; Sampaio, J.; Rohart, S.; et al. Optical Magnetometry of Single Biocompatible Micromagnets for Quantitative Magnetogenetic and Magnetomechanical Assays. Nano Lett. 2018, 18, 7635–7641. [Google Scholar] [CrossRef]
- Sangnier, A.P.; Van De Walle, A.B.; Curcio, A.; Le Borgne, R.; Motte, L.; Lalatonne, Y.; Wilhelm, C. Impact of magnetic nanoparticle surface coating on their long-term intracellular biodegradation in stem cells. Nanoscale 2019, 11, 16488–16498. [Google Scholar] [CrossRef]
- Safi, M.; Courtois, J.; Seigneuret, M.; Conjeaud, H.; Berret, J.-F. The effects of aggregation and protein corona on the cellular internalization of iron oxide nanoparticles. Biomaterials 2011, 32, 9353–9363. [Google Scholar] [CrossRef] [Green Version]
- Gazeau, F.; Wilhelm, C. Magnetic labeling, imaging and manipulation of endothelial progenitor cells using iron oxide nanoparticles. Futur. Med. Chem. 2010, 2, 397–408. [Google Scholar] [CrossRef]
- Maiuri, P.; Terriac, E.; Paul-Gilloteaux, P.; Vignaud, T.; McNally, K.; Onuffer, J.; Thorn, K.S.; Nguyen, P.A.; Georgoulia, N.; Soong, D.; et al. The first World Cell Race. Curr. Biol. 2012, 22, R673–R675. [Google Scholar] [CrossRef] [Green Version]
- De Anda, F.C.; Pollarolo, G.; Da Silva, J.S.; Camoletto, P.G.; Feiguin, F.; Dotti, C.G. Centrosome localization determines neuronal polarity. Nature 2005, 436, 704–708. [Google Scholar] [CrossRef] [PubMed]
- Lamoureux, P.; Heidemann, S.R.; Martzke, N.R.; Miller, K.E. Growth and elongation within and along the axon. Dev. Neurobiol. 2009, 70, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Lamoureux, P.L.; O’Toole, M.R.; Heidemann, S.R.; Miller, K.E. Slowing of axonal regeneration is correlated with increased axonal viscosity during aging. BMC Neurosci. 2010, 11, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falk, J.; Konopacki, F.A.; Zivraj, K.H.; Holt, C.E. Rab5 and Rab4 regulate axon elongation in the Xenopus visual system. J. Neurosci. 2014, 34, 373–391. [Google Scholar] [CrossRef] [Green Version]
- Pfister, B.J.; Bonislawski, D.P.; Smith, D.H.; Cohen, A.S. Stretch-grown axons retain the ability to transmit active electrical signals. FEBS Lett. 2006, 580, 3525–3531. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, J.L. How does an axon grow? Genes Dev. 2003, 17, 941–958. [Google Scholar] [CrossRef] [Green Version]
- Pfister, B.J.; Iwata, A.; Meaney, D.F.; Smith, U.H. Extreme Stretch Growth of Integrated Axons. J. Neurosci. 2004, 24, 7978–7983. [Google Scholar] [CrossRef] [Green Version]
- Steketee, M.B.; Moysidis, S.N.; Jin, X.-L.; Weinstein, J.E.; Pita-Thomas, W.; Raju, H.B.; Iqbal, S.; Goldberg, J.L. Nanoparticle-mediated signaling endosome localization regulates growth cone motility and neurite growth. Proc. Natl. Acad. Sci. USA 2011, 108, 19042–19047. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bongaerts, M.; Aizel, K.; Secret, E.; Jan, A.; Nahar, T.; Raudzus, F.; Neumann, S.; Telling, N.; Heumann, R.; Siaugue, J.-M.; et al. Parallelized Manipulation of Adherent Living Cells by Magnetic Nanoparticles-Mediated Forces. Int. J. Mol. Sci. 2020, 21, 6560. https://doi.org/10.3390/ijms21186560
Bongaerts M, Aizel K, Secret E, Jan A, Nahar T, Raudzus F, Neumann S, Telling N, Heumann R, Siaugue J-M, et al. Parallelized Manipulation of Adherent Living Cells by Magnetic Nanoparticles-Mediated Forces. International Journal of Molecular Sciences. 2020; 21(18):6560. https://doi.org/10.3390/ijms21186560
Chicago/Turabian StyleBongaerts, Maud, Koceila Aizel, Emilie Secret, Audric Jan, Tasmin Nahar, Fabian Raudzus, Sebastian Neumann, Neil Telling, Rolf Heumann, Jean-Michel Siaugue, and et al. 2020. "Parallelized Manipulation of Adherent Living Cells by Magnetic Nanoparticles-Mediated Forces" International Journal of Molecular Sciences 21, no. 18: 6560. https://doi.org/10.3390/ijms21186560