Anterograde Neuronal Circuit Tracers Derived from Herpes Simplex Virus 1: Development, Application, and Perspectives



Abstract
:1. Introduction
2. Neuronal Circuit and Traditional Tracers
3. Viral Tracers
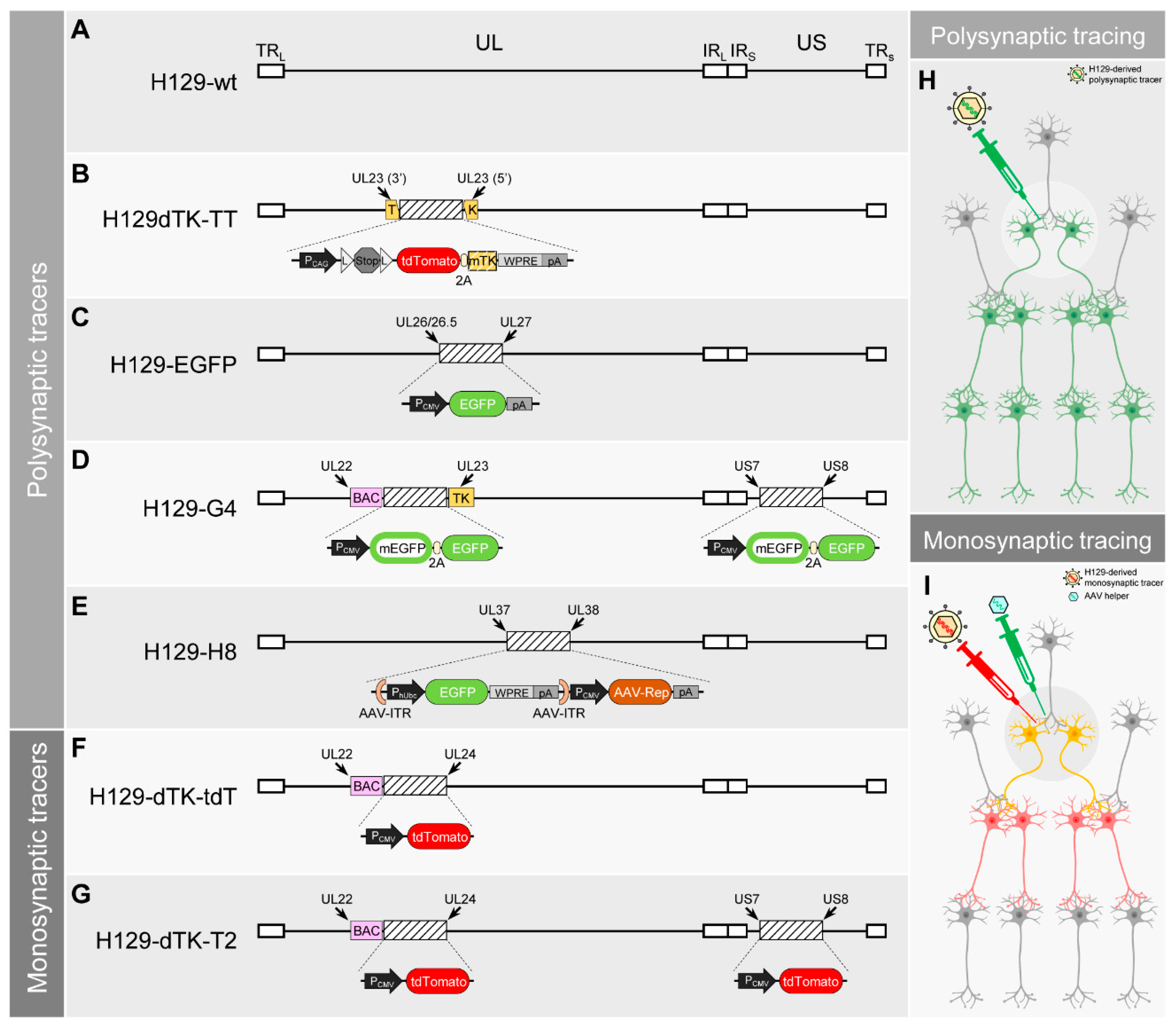
4. H129-Derived Anterograde Tracers
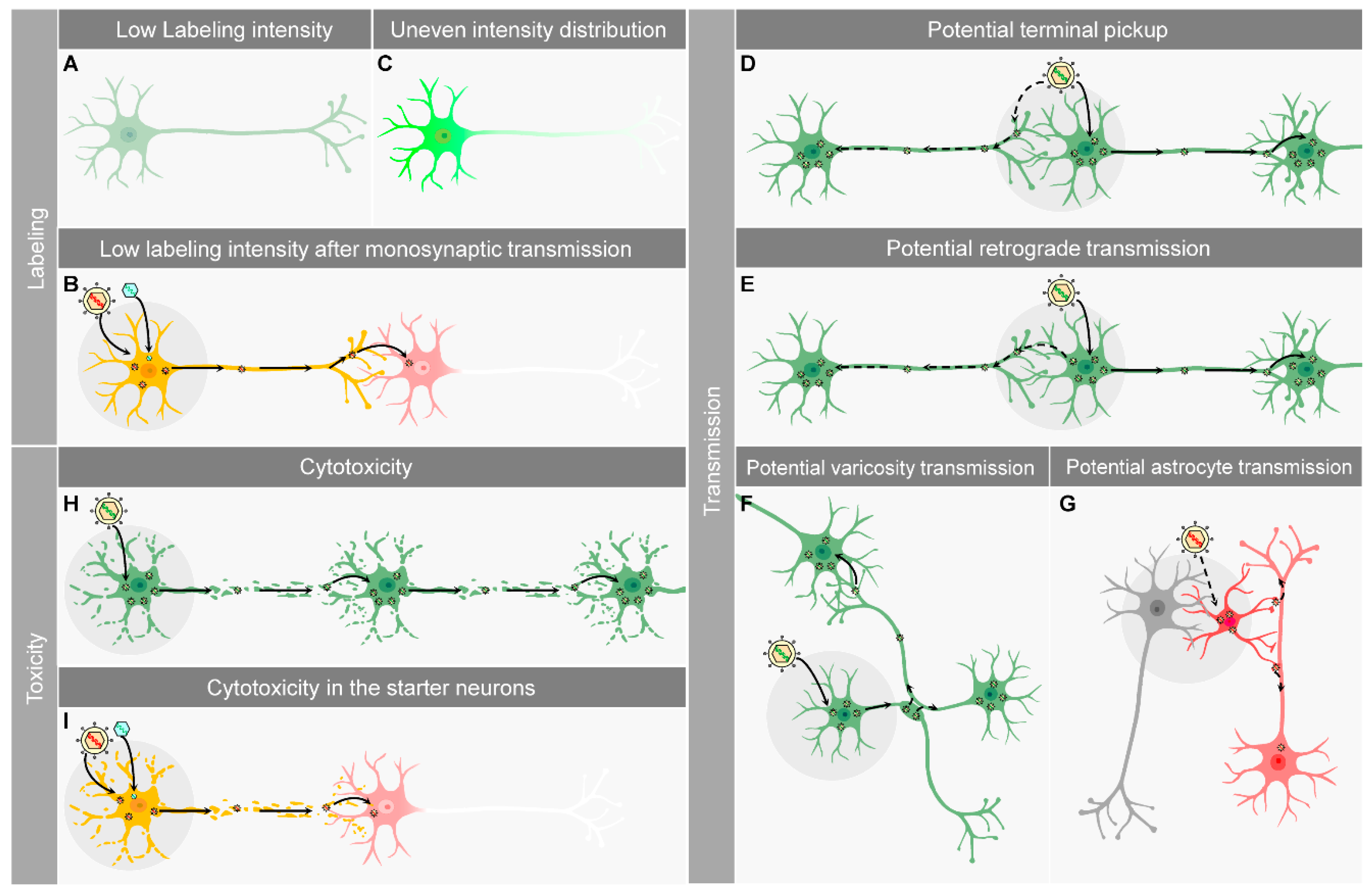
5. Limitations of Current H129-Derived Anterograde Tracers
5.1. Labeling Intensity and Distribution
5.2. Potential Retrograde Labeling and Transmission
5.3. Transneuronal Transmission vs. Transsynaptic Transmission
5.4. Toxicity to the Infected Neurons
5.5. Difficulties in H129 Genetic Manipulation
6. Strategies for Optimizing and Developing Future H129-Derived Tracers
6.1. Increasing Labeling Intensity
6.2. Reducing Non-Specific Tracing
6.3. Attenuating the Toxicity
6.4. Developing High Throughput Modification and Screening Methods
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HSV-1 | Herpes simplex virus 1 |
| H129 | Herpes simplex virus 1 (HSV-1) strain H129 |
| IE | immediate early gene |
| E | early gene |
| L | late gene |
| kb | kilobase |
| CNS | central nervous system |
| AAV | adeno-associated virus |
| CAV | canine adenoviral vector |
| RABV | rabies virus |
| PRV | pseudorabies virus |
| VSV | vesicular stomatitis virus |
| AAV1 | adeno-associated virus serotype 1 |
| AAV9 | adeno-associated virus serotype 9 |
| LMCV | lymphocytic choriomeningitis virus |
| TK | thymidine kinases |
| CMV | cytomegalovirus |
| tdT | tdTomato |
| GFP | green fluorescent protein |
| EGFP | enhanced GFP |
| mGFP | membrane-bound EGFP |
| fMOST | fluorescence Micro-Optical Sectioning Tomography |
| ITR | Inverted terminal repeat |
| RVdG | G-protein deleted monosynaptic rabies virus tracer |
| BAC | bacterial artificial chromosome |
| E.coli | Escherichia coli |
| Cre | Cre recombinase |
| UTR | untranslated region |
| saRNA | self-amplifying RNA |
| SINV | Sindbis virus |
| EnvA | subgroup A envelope glycoprotein of avian sarcoma leukosis virus |
| TVA | cellular receptor for EnvA |
| scFv | single-chain variable fragment |
| KOS | Herpes simplex virus 1 (HSV-1) strain KOS |
| AdV | adenovirus |
| ORF | open reading frame |
| siR | self-inactivating rabies virus |
| CRISPR | clustered regularly interspaced short palindromic repeats |
| Cas9 | CRISPR-associated protein 9 |
| sgRNA | single guide RNA |
| TRL | terminal repeat long |
| TRs | terminal repeat short |
| IRL | internal repeat long |
| IRs | internal repeat short |
| UL | unique long |
| UL | unique short |
| PCMV | CMV promoter |
| WPRE | woodchuck hepatitis virus posttranscriptional regulatory element |
| pA | poly(A) |
| AAV-Rep | Rep gene of AAV |
| Flp | flippase recombinase |
References
- Denes, C.E.; Everett, R.D.; Diefenbach, R.J. Tour de Herpes: Cycling Through the Life and Biology of HSV-1. Methods Mol. Biol. (Clifton, N.J.) 2020, 2060. [Google Scholar] [CrossRef]
- Gottlieb, S.L.; Giersing, B.K.; Hickling, J.; Jones, R.; Deal, C.; Kaslow, D.C. Meeting report: Initial World Health Organization consultation on herpes simplex virus (HSV) vaccine preferred product characteristics, March 2017. Vaccine 2019, 37, 7408–7418. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; He, N.; Su, M.; Feng, J.; Chen, L.; Gao, M. Herpes simplex virus infections among rural residents in eastern China. BMC Infect Dis. 2011, 11, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, L.; Qiu, Y.; Cheng, S.; Jiang, X.; Ma, Y.-P.; Fang, W.; Wang, W.; Cui, J.; Ruan, Q.; Zhao, F.; et al. Serologic and viral genome prevalence of HSV, EBV, and HCMV among healthy adults in Wuhan, China. J. Med. Virol. 2018, 90, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Lobo, A.M.; Agelidis, A.M.; Shukla, D. Pathogenesis of herpes simplex keratitis: The host cell response and ocular surface sequelae to infection and inflammation. Ocul. Surf. 2018. [Google Scholar] [CrossRef] [PubMed]
- Tyler, K.L. Acute Viral Encephalitis. N. Engl. J. Med. 2018, 379, 557–566. [Google Scholar] [CrossRef]
- Michaux, C.; Morlat, P.; Bonnet, F. Cytomegalovirus and other herpes virus infections in systemic diseases. Presse. Med. 2010, 39, 34–41. [Google Scholar] [CrossRef]
- Caruso Brown, A.E.; Cohen, M.N.; Tong, S.; Braverman, R.S.; Rooney, J.F.; Giller, R.; Levin, M.J. Pharmacokinetics and safety of intravenous cidofovir for life-threatening viral infections in pediatric hematopoietic stem cell transplant recipients. Antimicrob. Agents Chemother. 2015, 59, 3718–3725. [Google Scholar] [CrossRef] [Green Version]
- Cairns, D.M.; Rouleau, N.; Parker, R.N.; Walsh, K.G.; Gehrke, L.; Kaplan, D.L. A 3D human brain-like tissue model of herpes-induced Alzheimer’s disease. Sci. Adv. 2020, 6, eaay8828. [Google Scholar] [CrossRef]
- Linard, M.; Letenneur, L.; Garrigue, I.; Doize, A.; Dartigues, J.-F.; Helmer, C. Interaction between APOE4 and herpes simplex virus type 1 in Alzheimer’s disease. Alzheimers Dement. 2020, 16, 200–208. [Google Scholar] [CrossRef]
- Kumar, S.P.; Chandy, M.L.; Shanavas, M.; Khan, S.; Suresh, K.V. Pathogenesis and life cycle of herpes simplex virus infection-stages of primary, latency and recurrence. J. Oral Maxillofac. Surg. Med. Pathol. 2016, 28, 350–353. [Google Scholar] [CrossRef]
- Yan, C.; Luo, Z.; Li, W.; Li, X.; Dallmann, R.; Kurihara, H.; Li, Y.-F.; He, R.-R. Disturbed Yin-Yang balance: Stress increases the susceptibility to primary and recurrent infections of herpes simplex virus type 1. Acta. Pharm. Sin. B 2020, 10, 383–398. [Google Scholar] [CrossRef] [PubMed]
- Artusi, S.; Miyagawa, Y.; Goins, W.F.; Cohen, J.B.; Glorioso, J.C. Herpes Simplex Virus Vectors for Gene Transfer to the Central Nervous System. Diseases 2018, 6, 74. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; He, H.; Wang, H. Oncolytic herpes simplex virus and immunotherapy. BMC Immunol. 2018, 19, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Totsch, S.K.; Schlappi, C.; Kang, K.-D.; Ishizuka, A.S.; Lynn, G.M.; Fox, B.; Beierle, E.A.; Whitley, R.J.; Markert, J.M.; Gillespie, G.Y.; et al. Oncolytic herpes simplex virus immunotherapy for brain tumors: Current pitfalls and emerging strategies to overcome therapeutic resistance. Oncogene 2019, 38, 6159–6171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.-R.; Zhao, H.; Abdul-Muneer, P.M.; Cao, H.; Li, X.; Geller, A.I. Neurons can be labeled with unique hues by helper virus-free HSV-1 vectors expressing Brainbow. J. Neurosci. methods 2015, 240, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Zemanick, M.C.; Strick, P.L.; Dix, R.D. Direction of transneuronal transport of herpes simplex virus 1 in the primate motor system is strain-dependent. Proc. Natl. Acad. Sci. USA 1991, 88, 8048–8051. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.B.; Jiang, H.F.; Gang, Y.D.; Song, Y.G.; Shen, Z.Z.; Yang, H.; Dong, X.; Tian, Y.L.; Ni, R.J.; Liu, Y.; et al. Anterograde monosynaptic transneuronal tracers derived from herpes simplex virus 1 strain H129. Mol. Neurodegener. 2017, 12, 38. [Google Scholar] [CrossRef]
- Schmued, L.C.; Fallon, J.H. Fluoro-Gold: A new fluorescent retrograde axonal tracer with numerous unique properties. Brain Res. 1986, 377, 147–154. [Google Scholar] [CrossRef]
- Kuypers, H.G.; Bentivoglio, M.; van der Kooy, D.; Catsman-Berrevoets, C.E. Retrograde transport of bisbenzimide and propidium iodide through axons to their parent cell bodies. Neurosci. Lett. 1979, 12, 1–7. [Google Scholar] [CrossRef]
- Schwab, M.E.; Javoy-Agid, F.; Agid, Y. Labeled wheat germ agglutinin (WGA) as a new, highly sensitive retrograde tracer in the rat brain hippocampal system. Brain Res. 1978, 152, 145–150. [Google Scholar] [CrossRef]
- Schwab, M.E.; Suda, K.; Thoenen, H. Selective retrograde transsynaptic transfer of a protein, tetanus toxin, subsequent to its retrograde axonal transport. J. Cell Biol. 1979, 82, 798–810. [Google Scholar] [CrossRef] [Green Version]
- Cowan, W.M.; Gottlieb, D.I.; Hendrickson, A.E.; Price, J.L.; Woolsey, T.A. The autoradiographic demonstration of axonal connections in the central nervous system. Brain Res. 1972, 37, 21–51. [Google Scholar] [CrossRef]
- Wouterlood, F.G.; Bloem, B.; Mansvelder, H.D.; Luchicchi, A.; Deisseroth, K. A fourth generation of neuroanatomical tracing techniques: Exploiting the offspring of genetic engineering. J. Neurosci. Methods 2014, 235, 331–348. [Google Scholar] [CrossRef] [PubMed]
- Taymans, J.M.; Vandenberghe, L.H.; Haute, C.V.; Thiry, I.; Deroose, C.M.; Mortelmans, L.; Wilson, J.M.; Debyser, Z.; Baekelandt, V. Comparative analysis of adeno-associated viral vector serotypes 1, 2, 5, 7, and 8 in mouse brain. Hum. Gene. Ther. 2007, 18, 195–206. [Google Scholar] [CrossRef]
- Tervo, D.G.; Hwang, B.Y.; Viswanathan, S.; Gaj, T.; Lavzin, M.; Ritola, K.D.; Lindo, S.; Michael, S.; Kuleshova, E.; Ojala, D.; et al. A Designer AAV Variant Permits Efficient Retrograde Access to Projection Neurons. Neuron 2016, 92, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.Y.; Jang, M.J.; Yoo, B.B.; Greenbaum, A.; Ravi, N.; Wu, W.L.; Sanchez-Guardado, L.; Lois, C.; Mazmanian, S.K.; Deverman, B.E.; et al. Engineered AAVs for efficient noninvasive gene delivery to the central and peripheral nervous systems. Nat. Neurosci. 2017, 20, 1172–1179. [Google Scholar] [CrossRef]
- Peltekian, E.; Garcia, L.; Danos, O. Neurotropism and retrograde axonal transport of a canine adenoviral vector: A tool for targeting key structures undergoing neurodegenerative processes. Mol. Ther. 2002, 5, 25–32. [Google Scholar] [CrossRef]
- Soudais, C.; Laplace-Builhe, C.; Kissa, K.; Kremer, E.J. Preferential transduction of neurons by canine adenovirus vectors and their efficient retrograde transport in vivo. FASEB J. 2001, 15, 2283–2285. [Google Scholar] [CrossRef]
- Callaway, E.M.; Luo, L. Monosynaptic Circuit Tracing with Glycoprotein-Deleted Rabies Viruses. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 8979–8985. [Google Scholar] [CrossRef] [Green Version]
- Ugolini, G. Rabies virus as a transneuronal tracer of neuronal connections. Adv. Virus Res. 2011, 79, 165–202. [Google Scholar] [CrossRef] [PubMed]
- Wickersham, I.R.; Finke, S.; Conzelmann, K.K.; Callaway, E.M. Retrograde neuronal tracing with a deletion-mutant rabies virus. Nat. Methods 2007, 4, 47–49. [Google Scholar] [CrossRef] [PubMed]
- Ugolini, G. Specificity of rabies virus as a transneuronal tracer of motor networks: Transfer from hypoglossal motoneurons to connected second-order and higher order central nervous system cell groups. J. Comp. Neurol. 1995, 356, 457–480. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.N.; Banfield, B.W.; Smeraski, C.A.; Wilcox, C.L.; Dudek, F.E.; Enquist, L.W.; Pickard, G.E. Pseudorabies virus expressing enhanced green fluorescent protein: A tool for in vitro electrophysiological analysis of transsynaptically labeled neurons in identified central nervous system circuits. Proc. Natl. Acad. Sci. USA 2000, 97, 9264–9269. [Google Scholar] [CrossRef] [Green Version]
- Card, J.P.; Kobiler, O.; McCambridge, J.; Ebdlahad, S.; Shan, Z.; Raizada, M.K.; Sved, A.F.; Enquist, L.W. Microdissection of neural networks by conditional reporter expression from a Brainbow herpesvirus. Proc. Natl. Acad. Sci. USA 2011, 108, 3377–3382. [Google Scholar] [CrossRef] [Green Version]
- Ekstrand, M.I.; Enquist, L.W.; Pomeranz, L.E. The alpha-herpesviruses: Molecular pathfinders in nervous system circuits. Trends Mol. Med. 2008, 14, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Szpara, M.L.; Tafuri, Y.R.; Parsons, L.; Shreve, J.T.; Engel, E.A.; Enquist, L.W. Genome Sequence of the Anterograde-Spread-Defective Herpes Simplex Virus 1 Strain MacIntyre. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Tang, Y.; Yan, K.; Yu, J.; Zou, Y.; Xu, W.; Xiao, K.; Zhang, Z.; Li, W.; Wu, B.; et al. Differences in neurotropism and neurotoxicity among retrograde viral tracers. Mol. Neurodegener. 2019, 14, 8. [Google Scholar] [CrossRef]
- Luo, L.; Callaway, E.M.; Svoboda, K. Genetic dissection of neural circuits. Neuron 2008, 57, 634–660. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Callaway, E.M.; Svoboda, K. Genetic Dissection of Neural Circuits: A Decade of Progress. Neuron 2018, 98, 865. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Holmes, T.C.; Luo, M.-H.; Beier, K.T.; Horwitz, G.D.; Zhao, F.; Zeng, W.; Hui, M.; Semler, B.L.; Sandri-Goldin, R.M. Viral Vectors for Neural Circuit Mapping and Recent Advances in Trans-synaptic Anterograde Tracers. Neuron 2020. [Google Scholar] [CrossRef] [PubMed]
- Zingg, B.; Chou, X.L.; Zhang, Z.G.; Mesik, L.; Liang, F.; Tao, H.W.; Zhang, L.I. AAV-Mediated Anterograde Transsynaptic Tagging: Mapping Corticocollicular Input-Defined Neural Pathways for Defense Behaviors. Neuron 2017, 93, 33–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beier, K.T.; Saunders, A.; Oldenburg, I.A.; Miyamichi, K.; Akhtar, N.; Luo, L.; Whelan, S.P.; Sabatini, B.; Cepko, C.L. Anterograde or retrograde transsynaptic labeling of CNS neurons with vesicular stomatitis virus vectors. Proc. Natl. Acad. Sci. USA 2011, 108, 15414–15419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, K.; Zhong, X.; Ying, M.; Li, L.; Tao, S.; Zhu, X.; He, X.; Xu, F. A mutant vesicular stomatitis virus with reduced cytotoxicity and enhanced anterograde trans-synaptic efficiency. Mol. brain 2020, 13, 45. [Google Scholar] [CrossRef] [PubMed]
- Song, C.K.; Schwartz, G.J.; Bartness, T.J. Anterograde transneuronal viral tract tracing reveals central sensory circuits from white adipose tissue. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R501–R511. [Google Scholar] [CrossRef] [Green Version]
- Rinaman, L.; Schwartz, G. Anterograde transneuronal viral tracing of central viscerosensory pathways in rats. J. Neurosci. 2004, 24, 2782–2786. [Google Scholar] [CrossRef]
- Sun, N.; Cassell, M.D.; Perlman, S. Anterograde, transneuronal transport of herpes simplex virus type 1 strain H129 in the murine visual system. J. Virol. 1996, 70, 5405–5413. [Google Scholar] [CrossRef] [Green Version]
- Lo, L.; Anderson, D.J. A Cre-dependent, anterograde transsynaptic viral tracer for mapping output pathways of genetically marked neurons. Neuron 2011, 72, 938–950. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Xiong, F.; Song, Y.G.; Jiang, H.F.; Qin, H.B.; Zhou, J.; Lu, S.; Grieco, S.F.; Xu, X.; Zeng, W.B.; et al. HSV-1 H129-Derived Anterograde Neural Circuit Tracers: Improvements, Production and Applications. Neurosci. Bull. 2020, in press. [Google Scholar]
- Beier, K.T. Hitchhiking on the neuronal highway: Mechanisms of transsynaptic specificity. J. Chem. Neuroanat. 2019, 99, 9–17. [Google Scholar] [CrossRef]
- Dix, R.D.; McKendall, R.R.; Baringer, J.R. Comparative neurovirulence of herpes simplex virus type 1 strains after peripheral or intracerebral inoculation of BALB/c mice. Infect. Immun. 1983, 40, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnett, E.M.; Evans, G.D.; Sun, N.; Perlman, S.; Cassell, M.D. Anterograde tracing of trigeminal afferent pathways from the murine tooth pulp to cortex using herpes simplex virus type 1. J. Neurosci. 1995, 15, 2972–2984. [Google Scholar] [CrossRef] [PubMed]
- Archin, N.M.; Atherton, S.S. Rapid spread of a neurovirulent strain of HSV-1 through the CNS of BALB/c mice following anterior chamber inoculation. J. Neurovirol. 2002, 8, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Dum, R.P.; Levinthal, D.J.; Strick, P.L. The spinothalamic system targets motor and sensory areas in the cerebral cortex of monkeys. J. Neurosci. 2009, 29, 14223–14235. [Google Scholar] [CrossRef] [Green Version]
- Padilla, S.L.; Qiu, J.; Soden, M.E.; Sanz, E.; Nestor, C.C.; Barker, F.D.; Quintana, A.; Zweifel, L.S.; Rønnekleiv, O.K.; Kelly, M.J.; et al. Agouti-related peptide neural circuits mediate adaptive behaviors in the starved state. Nat. Neurosci. 2016, 19, 734–741. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Tellez, L.A.; Perkins, M.H.; Perez, I.O.; Qu, T.; Ferreira, J.; Ferreira, T.L.; Quinn, D.; Liu, Z.-W.; Gao, X.-B.; et al. A Neural Circuit for Gut-Induced Reward. Cell 2018, 175. [Google Scholar] [CrossRef] [Green Version]
- Lo, L.; Yao, S.; Kim, D.-W.; Cetin, A.; Harris, J.; Zeng, H.; Anderson, D.J.; Weissbourd, B. Connectional architecture of a mouse hypothalamic circuit node controlling social behavior. Proc. Natl. Acad. Sci. USA 2019, 116, 7503–7512. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Jin, S.; Lin, X.; Chen, L.; Qiao, X.; Jiang, L.; Zhou, P.; Johnston, K.G.; Golshani, P.; Nie, Q.; et al. CA1-projecting subiculum neurons facilitate object-place learning. Nat. Neurosci. 2019, 22, 1857–1870. [Google Scholar] [CrossRef]
- McGovern, A.E.; Davis-Poynter, N.; Rakoczy, J.; Phipps, S.; Simmons, D.G.; Mazzone, S.B. Anterograde neuronal circuit tracing using a genetically modified herpes simplex virus expressing EGFP. J. Neurosci. Methods 2012, 209, 158–167. [Google Scholar] [CrossRef]
- Deng, K.; Yang, L.; Xie, J.; Tang, H.; Wu, G.S.; Luo, H.R. Whole-brain mapping of projection from mouse lateral septal nucleus. Biol. Open 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Gong, H.; Zhang, B.; Wang, Q.; Yan, C.; Wu, J.; Liu, Q.; Zeng, S.; Luo, Q. Micro-optical sectioning tomography to obtain a high-resolution atlas of the mouse brain. Science 2010, 330, 1404–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, K.; Ahrens, S.; Zhang, X.; Schiff, H.; Ramakrishnan, C.; Fenno, L.; Deisseroth, K.; Zhao, F.; Luo, M.H.; Gong, L.; et al. The central amygdala controls learning in the lateral amygdala. Nat. Neurosci. 2017, 20, 1680–1685. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Zhou, W.; Jin, Y.; Tang, H.; Cao, P.; Mao, Y.; Xie, W.; Zhang, X.; Zhao, F.; Luo, M.H.; et al. A Central Amygdala Input to the Parafascicular Nucleus Controls Comorbid Pain in Depression. Cell Rep. 2019, 29, 3847–3858.e3845. [Google Scholar] [CrossRef] [Green Version]
- Pi, G.; Gao, D.; Wu, D.; Wang, Y.; Lei, H.; Zeng, W.; Gao, Y.; Yu, H.; Xiong, R.; Jiang, T.; et al. Posterior basolateral amygdala to ventral hippocampal CA1 drives approach behaviour to exert an anxiolytic effect. Nat. Commun. 2020, 11, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Yang, C.; Cui, Y.; Su, F.; Wang, Y.; Wang, Y.; Yuan, P.; Shang, S.; Li, H.; Zhao, J.; et al. An Excitatory Neural Assembly Encodes Short-Term Memory in the Prefrontal Cortex. Cell Rep. 2018, 22, 1734–1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Y.; Wang, Y.; Jiang, H.F.; Liu, J.H.; Jia, J.; Wang, K.; Zhao, F.; Luo, M.H.; Luo, M.M.; Wang, X.M. Impaired glutamatergic projection from the motor cortex to the subthalamic nucleus in 6-hydroxydopamine-lesioned hemi-parkinsonian rats. Exp. Neurol. 2017, 300, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Song, Z.; Tian, Y.; Tian, W.; Zhu, C.; Ji, G.; Luo, Y.; Chen, S.; Wang, L.; Mao, Y.; et al. Basolateral amygdala input to the medial prefrontal cortex controls obsessive-compulsive disorder-like checking behavior. Proc. Natl. Acad. Sci. USA 2019, 116, 3799–3804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Chen, J.; Xu, X.; Sun, W.J.; Chen, X.; Zhao, F.; Luo, M.H.; Liu, C.; Guo, Y.; Xie, W.; et al. Direct auditory cortical input to the lateral periaqueductal gray controls sound-driven defensive behavior. PLoS Biol. 2019, 17, e3000417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Fu, Y.; Cao, Z.; Shao, D.; Song, J.; Sheng, H.; Yang, L.; Cui, D.; Chen, M.; Zhao, F.; et al. A Conditioning-Strengthened Circuit From CA1 of Dorsal Hippocampus to Basolateral Amygdala Participates in Morphine-Withdrawal Memory Retrieval. Front. Neurosci. 2020, 14. [Google Scholar] [CrossRef]
- Su, P.; Ying, M.; Han, Z.; Xia, J.; Jin, S.; Li, Y.; Wang, H.; Xu, F. High-brightness anterograde transneuronal HSV1 H129 tracer modified using a Trojan horse-like strategy. Mol. Brain 2020, 13, 5. [Google Scholar] [CrossRef] [Green Version]
- Wickersham, I.R.; Lyon, D.C.; Barnard, R.J.; Mori, T.; Finke, S.; Conzelmann, K.K.; Young, J.A.; Callaway, E.M. Monosynaptic restriction of transsynaptic tracing from single, genetically targeted neurons. Neuron 2007, 53, 639–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Tucciarone, J.; Jiang, S.; Yin, F.; Wang, B.-S.; Wang, D.; Jia, Y.; Jia, X.; Li, Y.; Yang, T.; et al. Genetic Single Neuron Anatomy Reveals Fine Granularity of Cortical Axo-Axonic Cells. Cell Rep. 2019, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diefenbach, R.J.; Miranda-Saksena, M.; Douglas, M.W.; Cunningham, A.L. Transport and egress of herpes simplex virus in neurons. Rev. Med. Virol. 2008, 18, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Wojaczynski, G.J.; Engel, E.A.; Steren, K.E.; Enquist, L.W.; Patrick Card, J. The neuroinvasive profiles of H129 (herpes simplex virus type 1) recombinants with putative anterograde-only transneuronal spread properties. Brain Struct. Funct. 2015, 220, 1395–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, P.; Wang, H.; Xia, J.; Zhong, X.; Hu, L.; Li, Y.; Li, Y.; Ying, M.; Xu, F. Evaluation of retrograde labeling profiles of HSV1 H129 anterograde tracer. J. Chem. Neuroanat. 2019, 100, 101662. [Google Scholar] [CrossRef]
- Kakooza-Mwesige, A.; Tshala-Katumbay, D.; Juliano, S.L. Viral infections of the central nervous system in Africa. Brain Res. Bull. 2019, 145, 2–17. [Google Scholar] [CrossRef]
- Sattentau, Q. Avoiding the void: Cell-to-cell spread of human viruses. Nat. Rev. Microbiol. 2008, 6, 815–826. [Google Scholar] [CrossRef]
- Lee, E.C.; Yu, D.; Martinez de Velasco, J.; Tessarollo, L.; Swing, D.A.; Court, D.L.; Jenkins, N.A.; Copeland, N.G. A highly efficient Escherichia coli-based chromosome engineering system adapted for recombinogenic targeting and subcloning of BAC DNA. Genomics 2001, 73, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Ellis, H.M.; Lee, E.C.; Jenkins, N.A.; Copeland, N.G.; Court, D.L. An efficient recombination system for chromosome engineering in Escherichia coli. Proc. Natl. Acad. Sci. USA 2000, 97, 5978–5983. [Google Scholar] [CrossRef] [Green Version]
- Wiens, M.D.; Shen, Y.; Li, X.; Salem, M.A.; Smisdom, N.; Zhang, W.; Brown, A.; Campbell, R.E. A Tandem Green-Red Heterodimeric Fluorescent Protein with High FRET Efficiency. Chembiochem 2016, 17, 2361–2367. [Google Scholar] [CrossRef]
- Miyagawa, Y.; Marino, P.; Verlengia, G.; Uchida, H.; Goins, W.F.; Yokota, S.; Geller, D.A.; Yoshida, O.; Mester, J.; Cohen, J.B.; et al. Herpes simplex viral-vector design for efficient transduction of nonneuronal cells without cytotoxicity. Proc. Natl. Acad. Sci. USA 2015, 112, E1632–E1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verlengia, G.; Miyagawa, Y.; Ingusci, S.; Cohen, J.B.; Simonato, M.; Glorioso, J.C. Engineered HSV vector achieves safe long-term transgene expression in the central nervous system. Sci. Rep. 2017, 7, 1507. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, Y.; Verlengia, G.; Reinhart, B.; Han, F.; Uchida, H.; Zucchini, S.; Goins, W.F.; Simonato, M.; Cohen, J.B.; Glorioso, J.C. Deletion of the Virion Host Shut-off Gene Enhances Neuronal-Selective Transgene Expression from an HSV Vector Lacking Functional IE Genes. Mol. Ther. Methods Clin. Dev. 2017, 6, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannemann, H. Viral replicons as valuable tools for drug discovery. Drug Discov Today 2020, 25, 1026–1033. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, T.; Banerjee, S.; Kim, C.; Leubner, M.; Lamar, C.; Gupta, P.; Lee, B.; Neve, R.; Lisman, J. Memory Erasure Experiments Indicate a Critical Role of CaMKII in Memory Storage. Neuron 2017, 96. [Google Scholar] [CrossRef] [Green Version]
- Ferenczy, M.W.; DeLuca, N.A. Reversal of heterochromatic silencing of quiescent herpes simplex virus type 1 by ICP0. J. Virol. 2011, 85, 3424–3435. [Google Scholar] [CrossRef] [Green Version]
- Kameda, H.; Furuta, T.; Matsuda, W.; Ohira, K.; Nakamura, K.; Hioki, H.; Kaneko, T. Targeting green fluorescent protein to dendritic membrane in central neurons. Neurosci. Res. 2008, 61, 79–91. [Google Scholar] [CrossRef]
- Weed, D.J.; Nicola, A.V. Herpes simplex virus Membrane Fusion. Adv. Anat. Embryol. Cell Biol. 2017, 223, 29–47. [Google Scholar] [CrossRef] [Green Version]
- Sathiyamoorthy, K.; Chen, J.; Longnecker, R.; Jardetzky, T.S. The COMPLEXity in herpesvirus entry. Curr. Opin. Virol. 2017, 24, 97–104. [Google Scholar] [CrossRef]
- Eisenberg, R.J.; Atanasiu, D.; Cairns, T.M.; Gallagher, J.R.; Krummenacher, C.; Cohen, G.H. Herpes virus fusion and entry: A story with many characters. Viruses 2012, 4, 800–832. [Google Scholar] [CrossRef]
- Goins, W.F.; Hall, B.; Cohen, J.B.; Glorioso, J.C. Retargeting of herpes simplex virus (HSV) vectors. Curr. Opin. Virol. 2016, 21, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campadelli-Fiume, G.; Petrovic, B.; Leoni, V.; Gianni, T.; Avitabile, E.; Casiraghi, C.; Gatta, V. Retargeting Strategies for Oncolytic Herpes Simplex Viruses. Viruses 2016, 8, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, R.S.; Georgieva, E.R.; Borbat, P.P.; Freed, J.H.; Heldwein, E.E. Structural basis for membrane anchoring and fusion regulation of the herpes simplex virus fusogen gB. Nat. Struct. Mol. Biol. 2018, 25, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Leoni, V.; Gatta, V.; Casiraghi, C.; Nicosia, A.; Petrovic, B.; Campadelli-Fiume, G. A Strategy for Cultivation of Retargeted Oncolytic Herpes Simplex Viruses in Non-cancer Cells. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjulson, L.; Cassataro, D.; DasGupta, S.; Miesenbock, G. Cell-Specific Targeting of Genetically Encoded Tools for Neuroscience. Annu. Rev. Genet. 2016, 50, 571–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, A.T.; Saied, A.; Charles, A.; Subramanian, R.; Chouljenko, V.N.; Kousoulas, K.G. A herpes simplex virus 1 (McKrae) mutant lacking the glycoprotein K gene is unable to infect via neuronal axons and egress from neuronal cell bodies. MBIO 2012, 3, e00144-12. [Google Scholar] [CrossRef] [Green Version]
- Crump, C. Virus Assembly and Egress of HSV. Adv. Exp. Med. Biol. 2018, 1045, 23–44. [Google Scholar] [CrossRef]
- Koyuncu, O.O.; Hogue, I.B.; Enquist, L.W. Virus infections in the nervous system. Cell Host Microbe. 2013, 13, 379–393. [Google Scholar] [CrossRef] [Green Version]
- Miranda-Saksena, M.; Denes, C.E.; Diefenbach, R.J.; Cunningham, A.L. Infection and Transport of Herpes Simplex Virus Type 1 in Neurons: Role of the Cytoskeleton. Viruses 2018, 10, 92. [Google Scholar] [CrossRef] [Green Version]
- Denes, C.E.; Miranda-Saksena, M.; Cunningham, A.L.; Diefenbach, R.J. Cytoskeletons in the Closet-Subversion in Alphaherpesvirus Infections. Viruses 2018, 10, 79. [Google Scholar] [CrossRef] [Green Version]
- Dodding, M.P.; Way, M. Coupling viruses to dynein and kinesin-1. EMBO J. 2011, 30, 3527–3539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuRaine, G.; Wisner, T.W.; Howard, P.; Johnson, D.C. Kinesin-1 Proteins KIF5A, -5B, and -5C Promote Anterograde Transport of Herpes Simplex Virus Enveloped Virions in Axons. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diefenbach, R.J.; Davis, A.; Miranda-Saksena, M.; Fernandez, M.A.; Kelly, B.J.; Jones, C.A.; LaVail, J.H.; Xue, J.; Lai, J.; Cunningham, A.L. The Basic Domain of Herpes Simplex Virus 1 pUS9 Recruits Kinesin-1 To Facilitate Egress from Neurons. J. Virol. 2016, 90, 2102–2111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuRaine, G.; Wisner, T.W.; Howard, P.; Williams, M.; Johnson, D.C. Herpes Simplex Virus gE/gI and US9 Promote both Envelopment and Sorting of Virus Particles in the Cytoplasm of Neurons, Two Processes That Precede Anterograde Transport in Axons. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaVail, J.H.; Tauscher, A.N.; Sucher, A.; Harrabi, O.; Brandimarti, R. Viral regulation of the long distance axonal transport of herpes simplex virus nucleocapsid. Neuroscience 2007, 146, 974–985. [Google Scholar] [CrossRef] [Green Version]
- LaVail, J.H.; Topp, K.S.; Giblin, P.A.; Garner, J.A. Factors that contribute to the transneuronal spread of herpes simplex virus. J. Neurosci. Res. 1997, 49, 485–496. [Google Scholar] [CrossRef]
- Dong, X.; Zhou, J.; Qin, H.-B.; Xin, B.; Huang, Z.-L.; Li, Y.-Y.; Xu, X.-M.; Zhao, F.; Zhao, C.-J.; Liu, J.-J.; et al. Anterograde Viral Tracer Herpes Simplex Virus 1 Strain H129 Transports Primarily as Capsids in Cortical Neuron Axons. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Pontejo, S.M.; Murphy, P.M. Chemokines encoded by herpesviruses. J. Leukoc. Biol. 2017, 102, 1199–1217. [Google Scholar] [CrossRef]
- Boutell, C.; Everett, R.D. Regulation of alphaherpesvirus infections by the ICP0 family of proteins. J. Gen. Virol. 2013, 94, 465–481. [Google Scholar] [CrossRef]
- Johnson, P.A.; Miyanohara, A.; Levine, F.; Cahill, T.; Friedmann, T. Cytotoxicity of a replication-defective mutant of herpes simplex virus type 1. J. Virol. 1992, 66, 2952–2965. [Google Scholar] [CrossRef] [Green Version]
- Oroskar, A.A.; Read, G.S. Control of mRNA stability by the virion host shutoff function of herpes simplex virus. J. Virol. 1989, 63, 1897–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manivanh, R.; Mehrbach, J.; Charron, A.J.; Grassetti, A.; Cerón, S.; Taylor, S.A.; Cabrera, J.R.; Gerber, S.; Leib, D.A. Herpes Simplex Virus 1 ICP34.5 Alters Mitochondrial Dynamics in Neurons. J. Virol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, S.; Miyagawa, Y.; Sato, Y.; Yamamoto, M.; Adachi, K.; Kinoh, H.; Goins, W.F.; Cohen, J.B.; Glorioso, J.C.; Taniai, N.; et al. Protocol Optimization for the Production of the Non-Cytotoxic JΔNI5 HSV Vector Deficient in Expression of Immediately Early Genes. Mol. Ther. Methods Clin. Dev. 2020, 17, 612–621. [Google Scholar] [CrossRef] [PubMed]
- LaVail, J.H.; Tauscher, A.N.; Hicks, J.W.; Harrabi, O.; Melroe, G.T.; Knipe, D.M. Genetic and molecular in vivo analysis of herpes simplex virus assembly in murine visual system neurons. J. Virol. 2005, 79, 11142–11150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Zhang, B.; Wu, Y.; Guo, X. Amino Acid Mutation in Position 349 of Glycoprotein Affect the Pathogenicity of Rabies Virus. Front. Microbiol. 2020, 11, 481. [Google Scholar] [CrossRef]
- Luo, J.; Zhang, B.; Lyu, Z.; Wu, Y.; Zhang, Y.; Guo, X. Single amino acid change at position 255 in rabies virus glycoprotein decreases viral pathogenicity. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020. [Google Scholar] [CrossRef]
- Whisnant, A.W.; Jürges, C.S.; Hennig, T.; Wyler, E.; Prusty, B.; Rutkowski, A.J.; L’Hernault, A.; Djakovic, L.; Göbel, M.; Döring, K.; et al. Integrative functional genomics decodes herpes simplex virus 1. Nat. Commun. 2020, 11, 2038. [Google Scholar] [CrossRef]
- Ciabatti, E.; Gonzalez-Rueda, A.; Mariotti, L.; Morgese, F.; Tripodi, M. Life-Long Genetic and Functional Access to Neural Circuits Using Self-Inactivating Rabies Virus. Cell 2017, 170, 382–392 e314. [Google Scholar] [CrossRef] [Green Version]
- Asuri, P.; Bartel, M.A.; Vazin, T.; Jang, J.H.; Wong, T.B.; Schaffer, D.V. Directed evolution of adeno-associated virus for enhanced gene delivery and gene targeting in human pluripotent stem cells. Mol. Ther. 2012, 20, 329–338. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| H129-wt | H129dTK-TT | H129-EGFP | H129-G4 | H129-H8 | H129-dTK-tdT | H129-dTK-T2 | |
|---|---|---|---|---|---|---|---|
| Polysynaptic Tracing a | ✓ | ✓ | ✓ | ✓ | ✓ | ✗ | ✗ |
| Monosynaptic Tracing a | ✗ | ✗ | ✗ | ✗ | ✗ | ✓ | ✓ |
| Labeling Brightness b | - | +/++ | ++ | +++ | ++/+++ | + | + |
| Starter Neuron Specificity a | ✗ | Cre+ neuron | ✗ | ✗ | ✗ | Naïve/Cre+/Flp+/… (controlled by helper AAV expressing TK) | Naïve/Cre+/Flp+/… (controlled by helper AAV expressing TK) |
| Advantages | Works in primates | Polysynaptic tracing from Cre+ neurons | Increased labeling intensity | With the brightest labeling so far | Enhanced brightness | Monosynaptic tracer, suitable for starter neuron specific or nonspecific tracing | Monosynaptic tracer, suitable for starter neuron specific or nonspecific tracing, increased labeling intensity |
| Limitations | No fluorescence, requires immunostaining, potential retrograde labeling, high toxicity | Low labeling intensity, can’t trace from naïve neurons, potential retrograde labeling, high toxicity | No starter cell specificity, potential retrograde labeling, high toxicity | No starter cell specificity, potential retrograde labeling, high toxicity | No starter cell specificity | Low labeling intensity, requires immunostaining to visualize post-synaptic neurons, potential retrograde labeling, relatively high toxicity in the starter neuron | Relatively low labeling intensity, potential retrograde labeling, relatively low toxicity in the postsynaptic neurons but still high in the starter neurons |
| Original Articles | [17,51] | [48] | [59] | [18,49] | [70] | [18,49] | [49] |
| Application Articles c | [45,46,47,52,53,54] | [55,56,57,58] * | [60] | [61,62,63,64,65,66,67,68] | / | [64] | / |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Yang, H.; Xiong, F.; Xu, X.; Zeng, W.-B.; Zhao, F.; Luo, M.-H. Anterograde Neuronal Circuit Tracers Derived from Herpes Simplex Virus 1: Development, Application, and Perspectives. Int. J. Mol. Sci. 2020, 21, 5937. https://doi.org/10.3390/ijms21165937
Li D, Yang H, Xiong F, Xu X, Zeng W-B, Zhao F, Luo M-H. Anterograde Neuronal Circuit Tracers Derived from Herpes Simplex Virus 1: Development, Application, and Perspectives. International Journal of Molecular Sciences. 2020; 21(16):5937. https://doi.org/10.3390/ijms21165937
Chicago/Turabian StyleLi, Dong, Hong Yang, Feng Xiong, Xiangmin Xu, Wen-Bo Zeng, Fei Zhao, and Min-Hua Luo. 2020. "Anterograde Neuronal Circuit Tracers Derived from Herpes Simplex Virus 1: Development, Application, and Perspectives" International Journal of Molecular Sciences 21, no. 16: 5937. https://doi.org/10.3390/ijms21165937