Early Transcriptomic Changes upon Thalidomide Exposure Influence the Later Neuronal Development in Human Embryonic Stem Cell-Derived Spheres

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
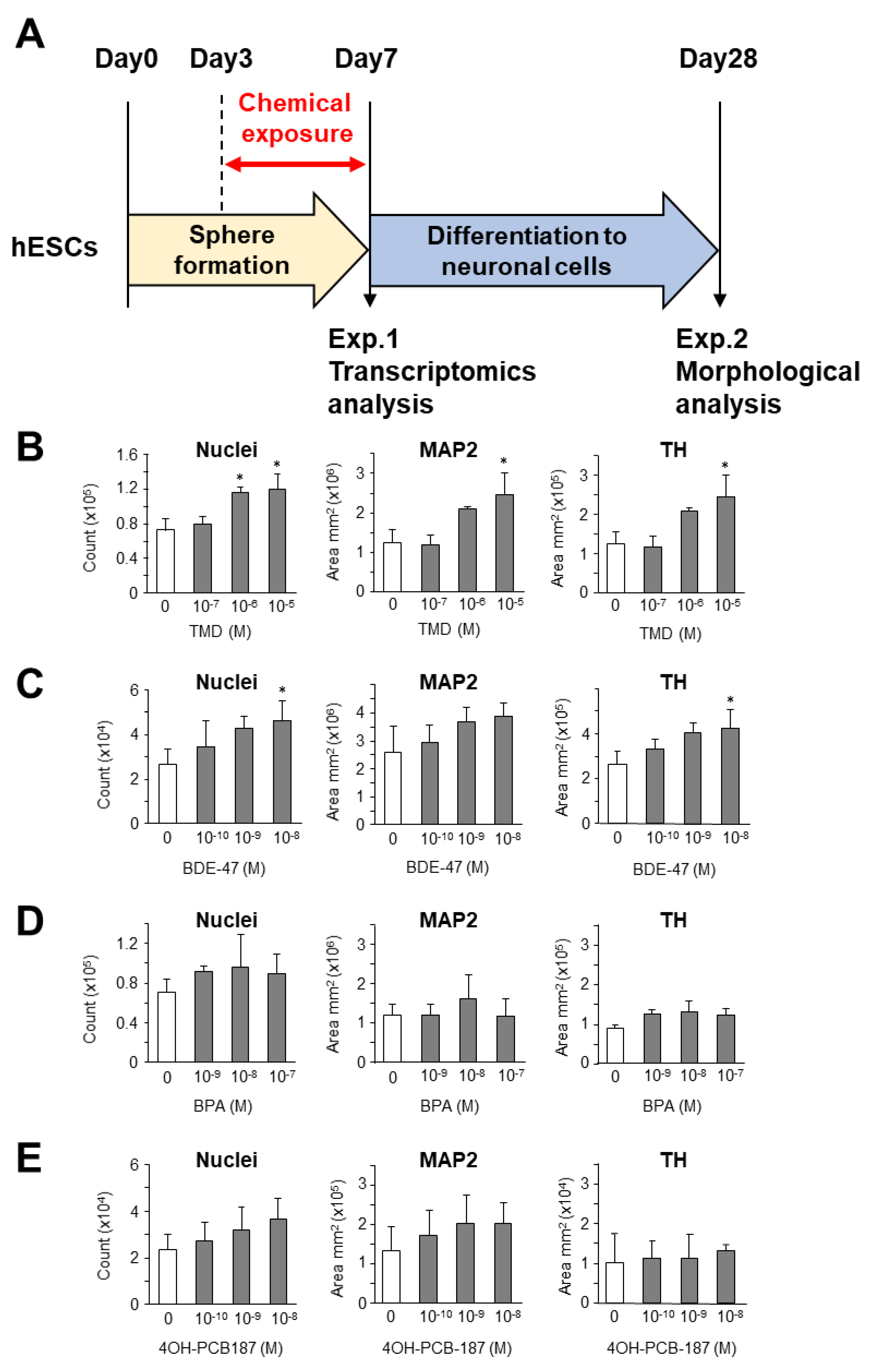
2.1. Morphological Analysis of the Effect of Chemical Exposure at the Early Stage of Development on the Neuronal Differentiation from hESC-Derived Spheres
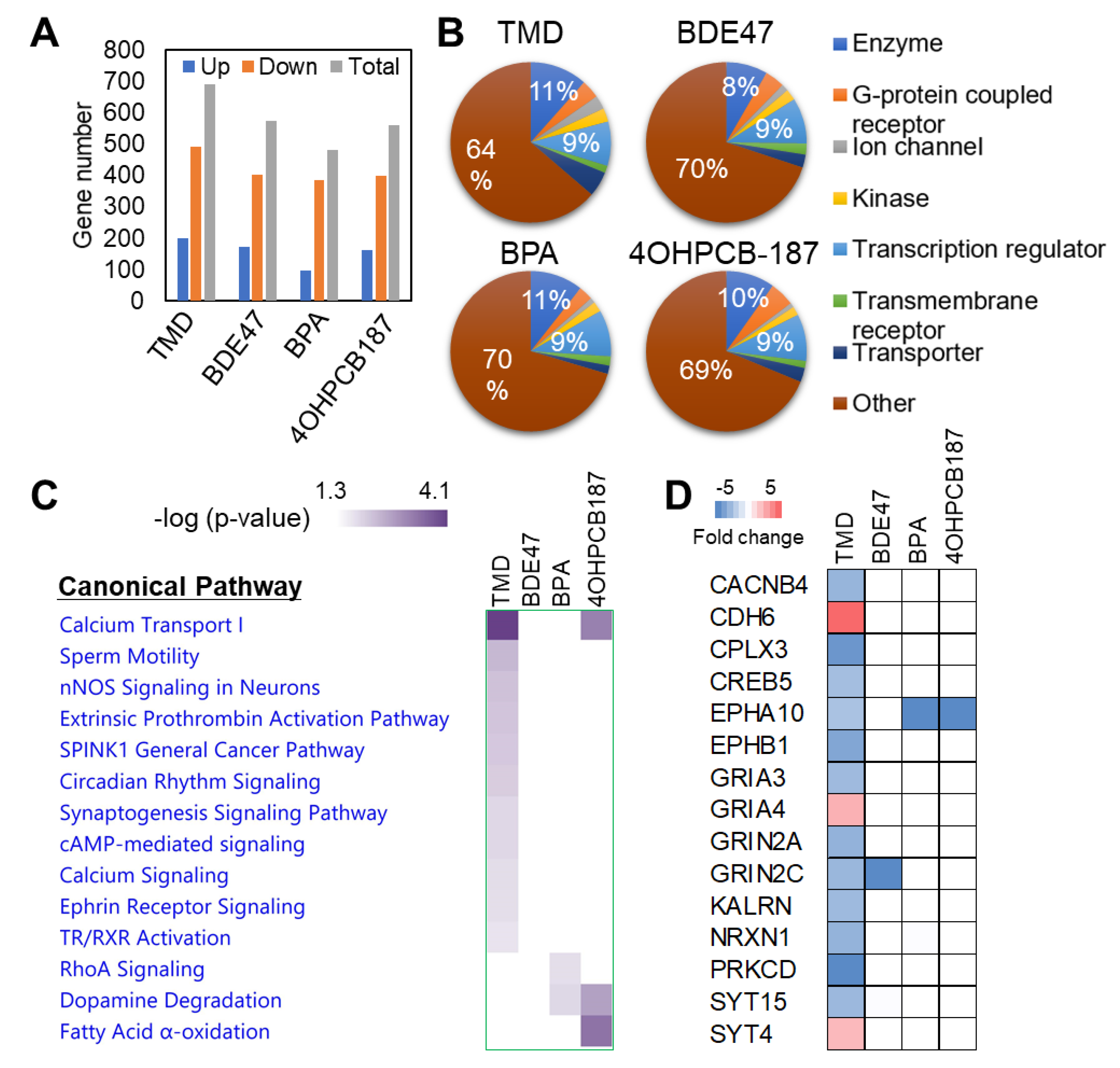
2.2. Transcriptional Analysis of the Effect of Chemical Exposure at the Early Stage of Neuronal Development Derived from hESCs
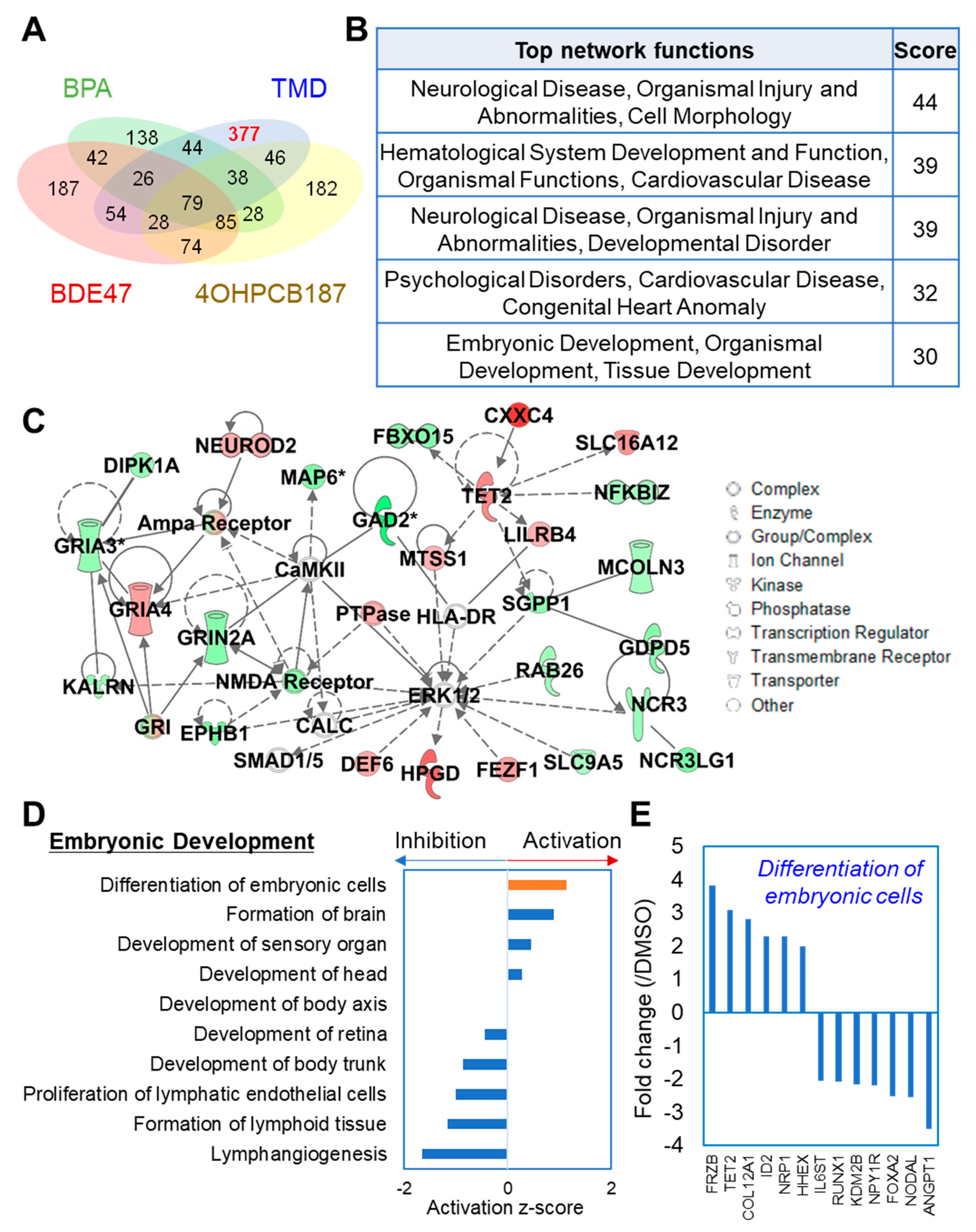
2.3. TMD-Specific Effect on Gene Expression at the Early Stage of Neuronal Development Derived from hESCs
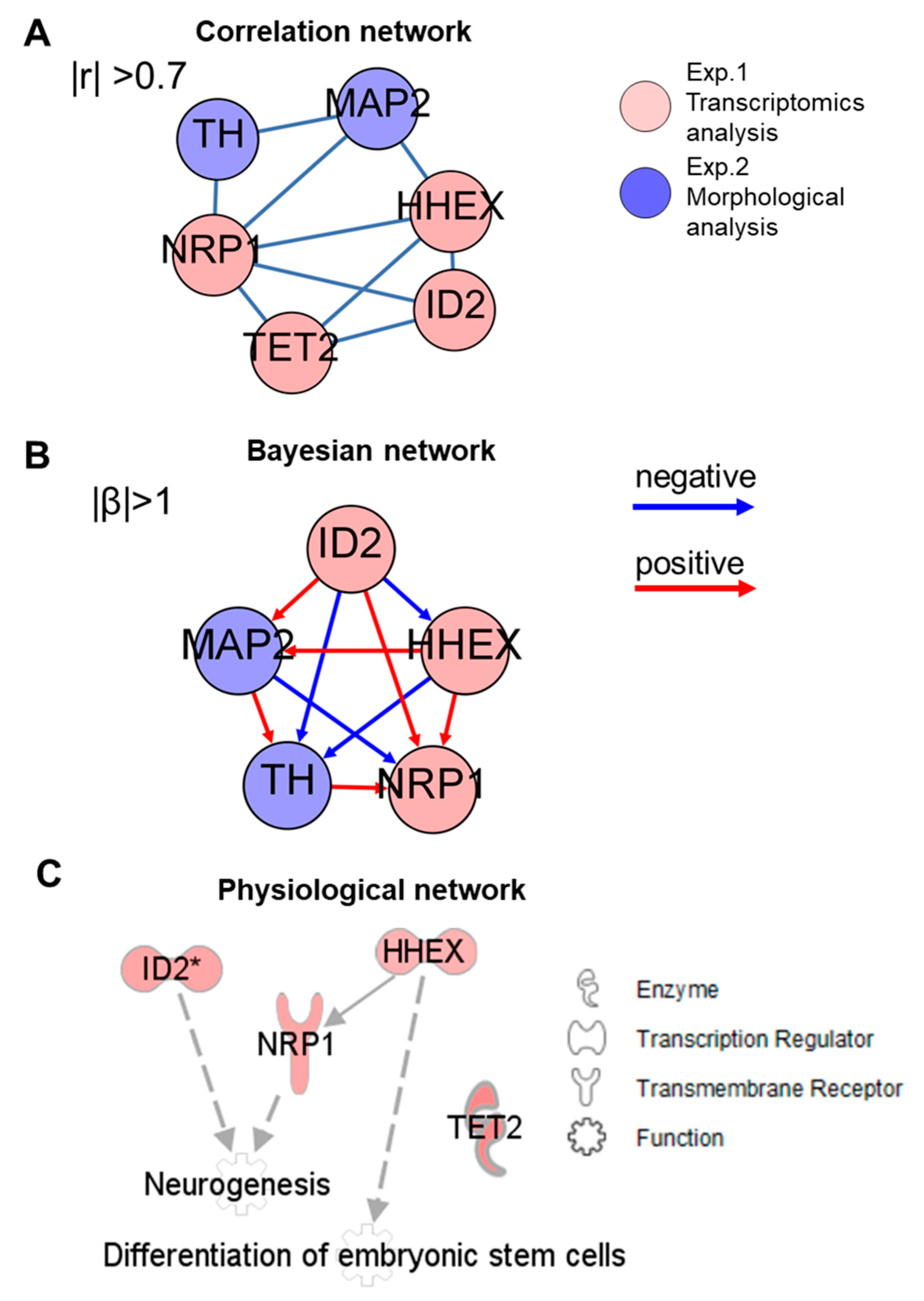
2.4. Integrated Network Analysis of Transcriptional and Morphological Changes during Neuronal Differentiation from hESC-Derived Spheres
3. Materials and Methods
3.1. Ethics Statement
3.2. Chemical Exposures, Culture and Neuronal Differentiation of Human ESCs
3.3. Immunocytochemistry and Image Analysis
3.4. Microarray Gene Expression Profiling
3.5. Knowledge-Based Pathway Analysis and Network Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BDE-47 | 2,2′,4,4′-tetrabromodiphenyl ether |
| BPA | bisphenol A |
| DMSO | dimethyl sulfoxide |
| hESC | human embryonic stem cell |
| 4OH-PCB187 | 4-hydroxy-2,2′,3,4′,5,5′,6-heptachlorobiphenyl |
| TMD | thalidomide |
References
- Patel, V.; Saxena, S.; Lund, C.; Thornicroft, S.G.; Baingana, F.; Bolton, P.; Chisholm, D.; Collins, P.Y.; Cooper, J.L.; Eaton, J.; et al. The Lancet Commission on global mental health and sustainable development. Lancet 2018, 392, 1553–1598. [Google Scholar] [CrossRef] [Green Version]
- Zablotsky, B.; Black, L.I.; Maenner, M.J.; Schieve, L.A.; Blumberg, S.J. Estimated Prevalence of Autism and Other Developmental Disabilities Following Questionnaire Changes in the 2014 National Health Interview Survey. Natl. Health Stat. Rep. 2015, 13, 1–20. [Google Scholar]
- Pessah, I.N.; Seegal, R.F.; Lein, P.J.; LaSalle, J.M.; Yee, B.K.; Van De Water, J.; Berman, R.F. Immunologic and neurodevelopmental susceptibilities of autism. Neurotoxicology 2008, 29, 532–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Council on Environmental Health, Pesticide exposure in children. Pediatrics 2012, 130, e1757–e1763. [CrossRef] [PubMed] [Green Version]
- Myhre, O.; Låg, M.; Villanger, G.D.; Oftedal, B.; Øvrevik, J.; Holme, J.A.; Aase, H.; Paulsen, R.E.; Bal-Price, A.; Dirven, H. Early life exposure to air pollution particulate matter (PM) as risk factor for attention deficit/hyperactivity disorder (ADHD): Need for novel strategies for mechanisms and causalities. Toxicol. Appl. Pharmacol. 2018, 354, 196–214. [Google Scholar] [CrossRef]
- O’Rahilly, R.; Müller, F. Significant features in the early prenatal development of the human brain. Ann. Anat. Anat. Anz. 2008, 190, 105–118. [Google Scholar] [CrossRef]
- Kim, J.Y.; Son, M.J.; Son, C.Y.; Radua, J.; Eisenhut, M.; Gressier, F.; Koyanagi, A.; Carvalho, A.F.; Stubbs, B.; Solmi, M.; et al. Environmental risk factors and biomarkers for autism spectrum disorder: An umbrella review of the evidence. Lancet Psychiatry 2019, 6, 590–600. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, M.P.; Hou, Z.; Propson, N.E.; Zhang, J.; Engstrom, C.J.; Costa, V.S.; Jiang, P.; Nguyen, B.K.; Bolin, J.M.; Daly, W.; et al. Human pluripotent stem cell-derived neural constructs for predicting, neural toxicity. Proc. Natl. Acad. Sci. USA 2015, 112, 12516–12521. [Google Scholar] [CrossRef] [Green Version]
- Nagano, R.; Akanuma, H.; Qin, X.-Y.; Imanishi, S.; Toyoshiba, H.; Yoshinaga, J.; Ohsako, S.; Sone, H. Multi-Parametric Profiling Network Based on Gene Expression and Phenotype Data: A Novel Approach to Developmental Neurotoxicity Testing. Int. J. Mol. Sci. 2012, 13, 187–207. [Google Scholar] [CrossRef]
- Pamies, D.; Block, K.; Lau, P.; Gribaldo, L.; Pardo, C.A.; Barreras, P.; Smirnova, L.; Wiersma, D.; Zhao, L.; Harris, G.; et al. Rotenone exerts developmental neurotoxicity in a human brain spheroid model. Toxicol. Appl. Pharmacol. 2018, 354, 101–114. [Google Scholar] [CrossRef]
- He, X.; Imanishi, S.; Sone, H.; Nagano, R.; Qin, X.-Y.; Yoshinaga, J.; Akanuma, H.; Yamane, J.; Fujibuchi, W.; Ohsako, S.; et al. Effects of methylmercury exposure on neuronal differentiation of mouse and human embryonic stem cells. Toxicol. Lett. 2012, 212, 1–10. [Google Scholar] [CrossRef]
- Zeng, Y.; Kurokawa, Y.; Win-Shwe, T.-T.; Nansai, H.; Zhang, Z.; Sone, H. Effects of polyamidoamine dendrimers on a 3-D neurosphere system using human neural progenitor cells. Toxicol. Sci. 2016, 152, 128–144. [Google Scholar] [CrossRef] [Green Version]
- Katsura, M.; Cyou-Nakamine, H.; Zen, Q.; Zen, Y.; Nansai, H.; Amagasa, S.; Kanki, Y.; Inoue, T.; Kaneki, K.; Taguchi, A.; et al. Effects of Chronic Low-Dose Radiation on Human Neural Progenitor Cells. Sci. Rep. 2016, 6, 20027. [Google Scholar] [CrossRef]
- Qin, X.-Y.; Akanuma, H.; Wei, F.; Nagano, R.; Zeng, Q.; Imanishi, S.; Ohsako, S.; Yoshinaga, J.; Yonemoto, J.; Tanokura, M.; et al. Effect of low-dose thalidomide on dopaminergic neuronal differentiation of human neural progenitor cells: A combined study of metabolomics and morphological analysis. NeuroToxicology 2012, 33, 1375–1380. [Google Scholar] [CrossRef]
- Miyazaki, K.; Narita, N.; Narita, M. Maternal administration of thalidomide or valproic acid causes abnormal serotonergic neurons in the offspring: Implication for pathogenesis of autism. Int. J. Dev. Neurosci. 2005, 23, 287–297. [Google Scholar] [CrossRef]
- Rostamian, A.; Gharedaghi, A.; Norouzi-Javidan, A.; Dehpour, A.R. Involvement of the nitric oxide pathway in the anti-depressant-like effects of thalidomide in mice. Physiol. Behav. 2019, 208, 112572. [Google Scholar] [CrossRef]
- Stoeckel, M.C.; Seitz, R.J.; Buetefisch, C.M. Congenitally altered motor experience alters somatotopic organization of human primary motor cortex. Proc. Natl. Acad. Sci. USA 2009, 106, 2395–2400. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.T.; Strömland, K.K. What can we learn from the thalidomide experience. Curr. Opin. Ophthalmol. 2011, 22, 356–364. [Google Scholar] [CrossRef] [Green Version]
- Hideshima, T.; Chauhan, D.; Shima, Y.; Raje, N.; Davies, F.E.; Tai, Y.T.; Treon, S.P.; Lin, B.; Schlossman, R.L.; Richardson, P.; et al. Thalidomide and its analogs overcome drug resistance of human multiple myeloma cells to conventional therapy. Blood. 2000, 96, 2943–2950. [Google Scholar] [CrossRef]
- Donovan, K.A.; An, J.; Nowak, R.P.; Yuan, J.C.; Fink, E.C.; Berry, B.C.; Ebert, B.L.; Fischer, E.S. Thalidomide promotes degradation of SALL4, a transcription factor implicated in Duane Radial Ray syndrome. eLife 2018, 7, e38430. [Google Scholar] [CrossRef]
- Chen, H.; Seifikar, H.; Larocque, N.; Kim, Y.; Khatib, I.; Fernandez, C.J.; Abello, N.; Robinson, J.F. Using a Multi-Stage hESC Model to Characterize BDE-47 Toxicity during Neurogenesis. Toxicol. Sci. 2019, 171, 221–234. [Google Scholar] [CrossRef]
- Gaylord, A.; Osborne, G.; Ghassabian, A.; Malits, J.; Attina, T.; Trasande, L. Trends in neurodevelopmental disability burden due to early life chemical exposure in the USA from 2001 to 2016: A population-based disease burden and cost analysis. Mol. Cell. Endocrinol. 2020, 502, 110666. [Google Scholar] [CrossRef] [PubMed]
- Kundakovic, M.; Gudsnuk, K.; Herbstman, J.B.; Tang, D.; Perera, F.P.; Champagne, F.A. DNA methylation of BDNF as a biomarker of early-life adversity. Proc. Natl. Acad. Sci. USA 2015, 112, 6807–6813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura-Kuroda, J.; Nagata, I.; Kuroda, Y. Disrupting effects of hydroxy-polychlorinated biphenyl (PCB) congeners on neuronal development of cerebellar Purkinje cells: A possible causal factor for developmental brain disorders? Chemosphere 2007, 67, S412–S420. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, P.; Landrigan, P. Neurobehavioural effects of developmental toxicity. Lancet Neurol. 2014, 13, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Haraguchi, K.; Ito, Y.; Takagi, M.; Fujii, Y.; Harada, K.H.; Koizumi, A. Levels, profiles and dietary sources of hydroxylated PCBs and hydroxylated and methoxylated PBDEs in Japanese women serum samples. Environ. Int. 2016, 97, 155–162. [Google Scholar] [CrossRef]
- Poon, E.; Powers, B.E.; McAlonan, R.M.; Ferguson, D.C.; Schantz, S.L. Effects of Developmental Exposure to Polychlorinated Biphenyls and/or Polybrominated Diphenyl Ethers on Cochlear Function. Toxicol. Sci. 2011, 124, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Meganathan, K.; Jagtap, S.; Wagh, V.; Winkler, J.; Gaspar, J.A.; Hildebrand, D.; Trusch, M.; Lehmann, K.; Hescheler, J.; Schlüter, H.; et al. Identification of thalidomide-specific transcriptomics and proteomics signatures during differentiation of human embryonic stem cells. PLoS ONE 2012, 7, e44228. [Google Scholar] [CrossRef]
- Schreiber, T.; Gassmann, K.; Götz, C.; Hübenthal, U.; Moors, M.; Krause, G.; Merk, H.F.; Nguyen, N.; Scanlan, T.S.; Abel, J.; et al. Polybrominated diphenyl ethers induce developmental neurotoxicity in a human in vitro model: Evidence for endocrine disruption. Environ. Health Perspect. 2010, 118, 572–578. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.E.; Lim, M.S.; Park, J.H.; Park, C.H.; Koh, H.C. PTEN promotes dopaminergic neuronal differentiation through regulation of erk-dependent inhibition of s6k signaling in human neural stem cells. Stem Cells Transl. Med. 2016, 5, 1319–1329. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.-Y.; Hara, M.; Arner, E.; Kawaguchi, Y.; Inoue, I.; Tatsukawa, H.; Furutani, Y.; Nagatsuma, K.; Matsuura, T.; Wei, F.; et al. Transcriptome Analysis Uncovers a Growth-Promoting Activity of Orosomucoid-1 on Hepatocytes. EBioMedicine 2017, 24, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libbrecht, M.W.; Noble, W.S. Machine Learning Applications in Genetics and Genomics. Nat. Rev. Genet. 2015, 16, 321–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, S.; Liang, S.; Zhou, H.; Yin, N.; Faiola, F. Typical halogenated flame retardants affect human neural stem cell gene expression during proliferation and differentiation via glycogen synthase kinase 3 beta and T3 signaling. Ecotoxicol. Environ. Saf. 2019, 183, 109498. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Yin, N.; Liang, S.; Yang, R.; Liu, S.; Lu, Y.; Jiang, L.; Zhou, Q.; Jiang, G.; Faiola, F. Bisphenol A and several derivatives exert neural toxicity in human neuron-like cells by decreasing neurite length. Food Chem. Toxicol. 2020, 135, 111015. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Li, Y.; Wu, K.; Hao, S.; Cai, Q.; Zhou, Z.; Yang, H. Effects of environmental chemicals on the proliferation and differentiation of neural stem cells. Environ. Toxicol. 2019, 34, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Caspersen, I.H.; Aase, H.; Biele, G.; Brantsæter, A.L.; Haugen, M.; Kvalem, H.E.; Skogan, A.H.; Zeiner, P.; Alexander, J.; Meltzer, H.M.; et al. The influence of maternal dietary exposure to dioxins and PCBs during pregnancy on ADHD symptoms and cognitive functions in Norwegian preschool children. Environ. Int. 2016, 94, 649–660. [Google Scholar] [CrossRef]
- Suemori, H.; Yasuchika, K.; Hasegawa, K.; Fujioka, T.; Tsuneyoshi, N.; Nakatsuji, N. Efficient establishment of human embryonic stem cell lines and long-term maintenance with stable karyotype by enzymatic bulk passage. Biochem. Biophys. Res. Commun. 2006, 345, 926–932. [Google Scholar] [CrossRef]
- Shimizukawa, R.; Sakata, A.; Hirose, M.; Takahashi, A.; Iseki, H.; Liu, Y.; Kunita, S.; Sugiyama, F.; Yagami, K.-I. Establishment of a new embryonic stem cell line derived from C57BL/6 mouse expressing EGFP ubiquitously. Genesis 2005, 42, 47–52. [Google Scholar] [CrossRef]
- Yamane, J.; Aburatani, S.; Imanishi, S.; Akanuma, H.; Nagano, R.; Kato, T.; Sone, H.; Ohsako, S.; Fujibuchi, W. Prediction of developmental chemical toxicity based on gene networks of human embryonic stem cells. Nucleic Acids Res. 2016, 44, 5515–5528. [Google Scholar] [CrossRef]
- Darras, V.M. Endocrine disrupting polyhalogenated organic pollutants interfere with thyroid hormone signalling in the developing brain. Cerebellum 2008, 7, 26–37. [Google Scholar] [CrossRef]
- Mustieles, V.; Pérez-Lobato, R.; Olea, N.; Fernández, M.F. Bisphenol A: Human exposure and neurobehavior. NeuroToxicology 2015, 49, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Mercogliano, R.; Santonicola, S. Investigation on bisphenol A levels in human milk and dairy supply chain: A review. Food Chem. Toxicol. 2018, 114, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Berghuis, S.A.; Soechitram, S.D.; Hitzert, M.M.; Sauer, P.J.; Bos, A.F. Prenatal exposure to polychlorinated biphenyls and their hydroxylated metabolites is associated with motor development of three-month-old infants. NeuroToxicology 2013, 38, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Jaspers, V.L.B.; Dietz, R.; Sonne, C.; Letcher, R.J.; Eens, M.; Neels, H.M.; Born, E.W.; Covaci, A. A screening of persistent organohalogenated contaminants in hair of East Greenland polar bears. Sci. Total Environ. 2010, 408, 5613–5618. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-S.; Bergman, Å.; Linderholm, L.; Athanasiadou, M.; Kočan, A.; Petřík, J.; Drobna, B.; Trnovec, T.; Charles, M.J.; Hertz-Picciotto, I. Placental transfer of polychlorinated biphenyls, their hydroxylated metabolites and pentachlorophenol in pregnant women from eastern Slovakia. Chemosphere 2008, 70, 1676–1684. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Kurokawa, Y.; Win-Shwe, T.-T.; Zeng, Q.; Hirano, S.; Zhang, Z.; Sone, H.; Sone, H. Effects of PAMAM dendrimers with various surface functional groups and multiple generations on cytotoxicity and neuronal differentiation using human neural progenitor cells. J. Toxicol. Sci. 2016, 41, 351–370. [Google Scholar] [CrossRef] [Green Version]
- Sone, H.; Ito, T. Comparative Analysis between Effects of Thalidomide and Environmental Chemicals on Neuronal Differentiations in Human Sphere. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE151239 (accessed on 28 May 2020).




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kikegawa, M.; Qin, X.-Y.; Ito, T.; Nishikawa, H.; Nansai, H.; Sone, H. Early Transcriptomic Changes upon Thalidomide Exposure Influence the Later Neuronal Development in Human Embryonic Stem Cell-Derived Spheres. Int. J. Mol. Sci. 2020, 21, 5564. https://doi.org/10.3390/ijms21155564
Kikegawa M, Qin X-Y, Ito T, Nishikawa H, Nansai H, Sone H. Early Transcriptomic Changes upon Thalidomide Exposure Influence the Later Neuronal Development in Human Embryonic Stem Cell-Derived Spheres. International Journal of Molecular Sciences. 2020; 21(15):5564. https://doi.org/10.3390/ijms21155564
Chicago/Turabian StyleKikegawa, Mami, Xian-Yang Qin, Tomohiro Ito, Hiromi Nishikawa, Hiroko Nansai, and Hideko Sone. 2020. "Early Transcriptomic Changes upon Thalidomide Exposure Influence the Later Neuronal Development in Human Embryonic Stem Cell-Derived Spheres" International Journal of Molecular Sciences 21, no. 15: 5564. https://doi.org/10.3390/ijms21155564