Comparative Analysis of Age-Related Changes in Lacrimal Glands and Meibomian Glands of a C57BL/6 Male Mouse Model


Abstract
:1. Introduction
2. Results
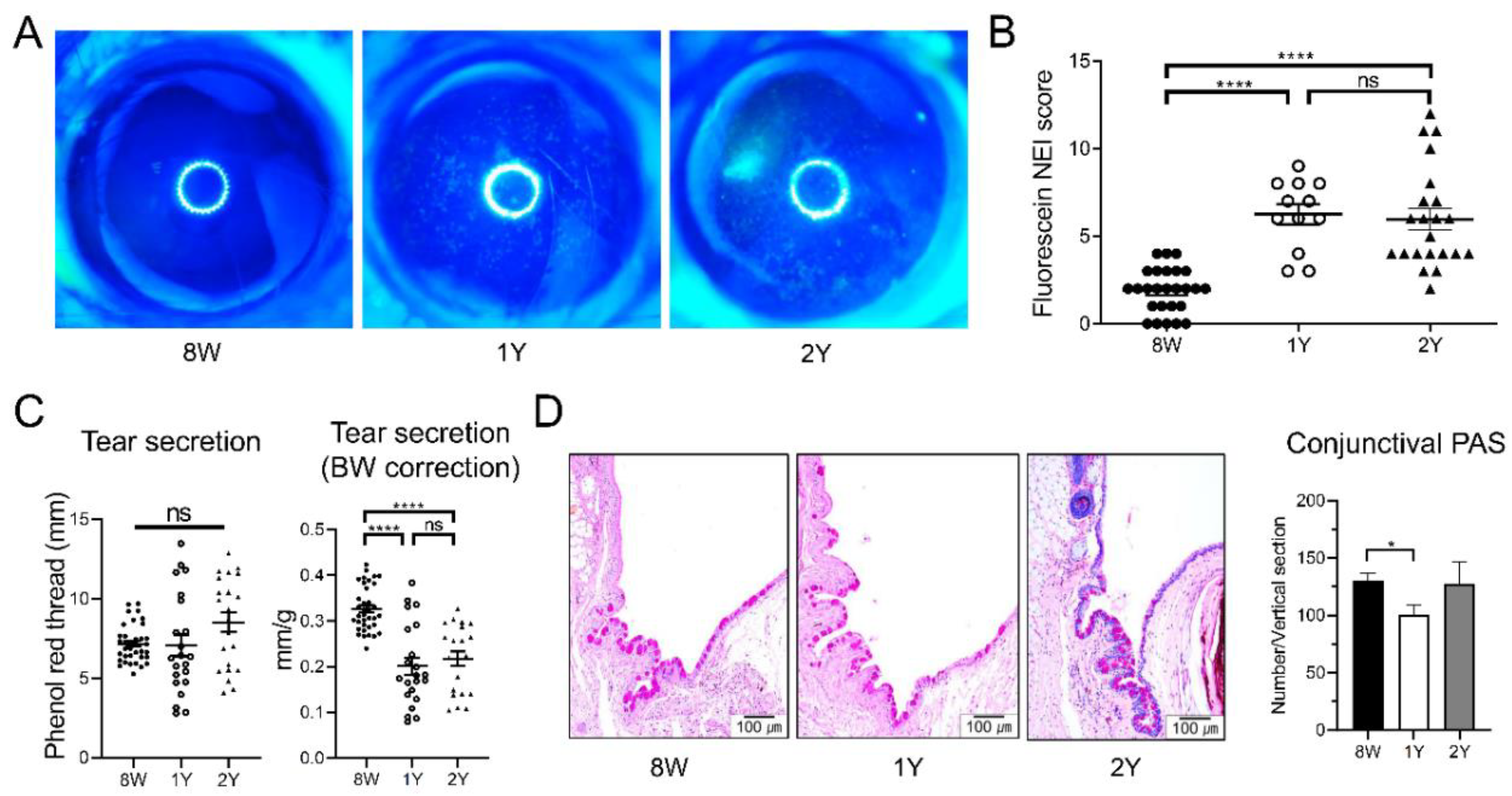
2.1. DED Was Established in Aged C57BL/6 Male Mice
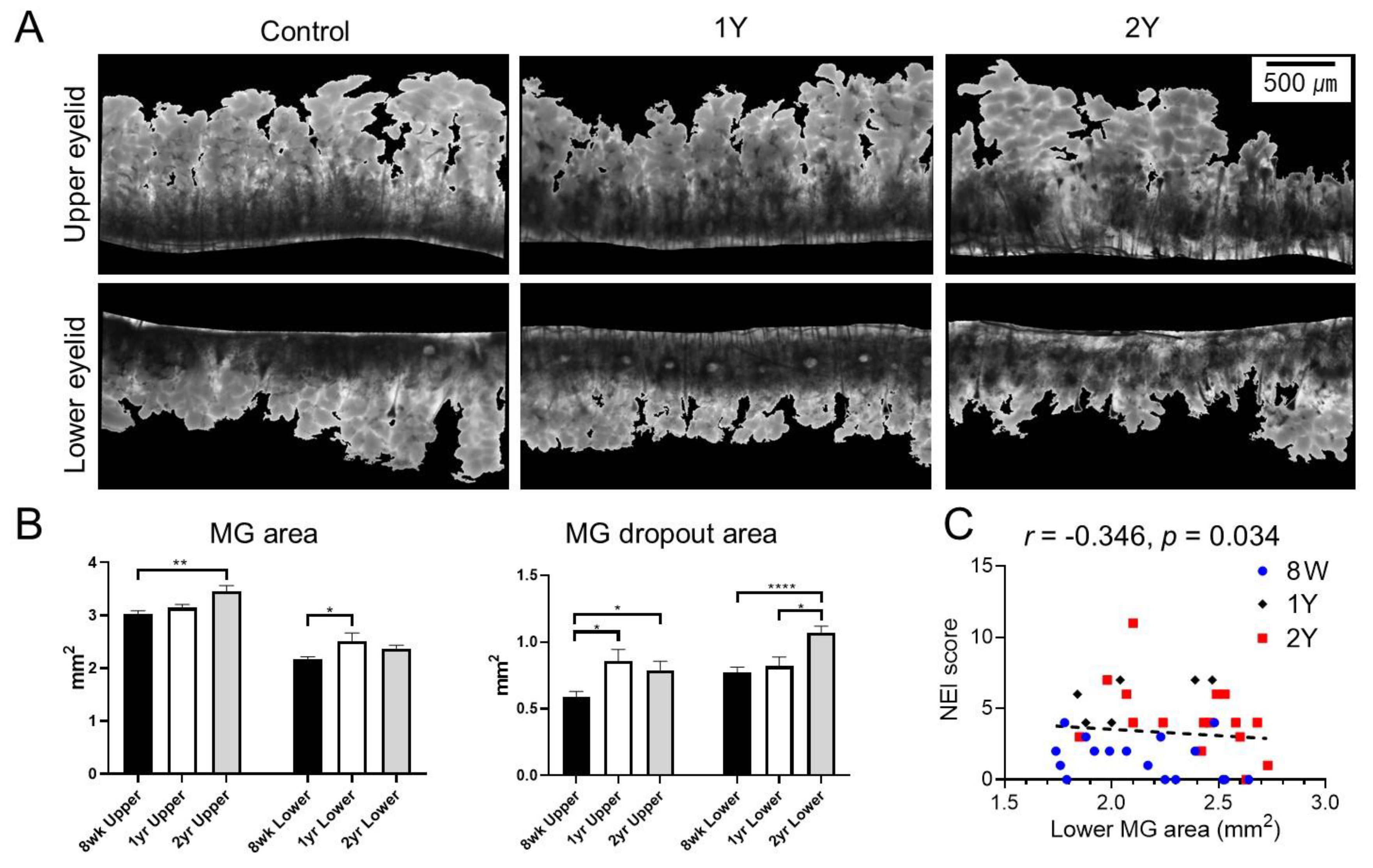
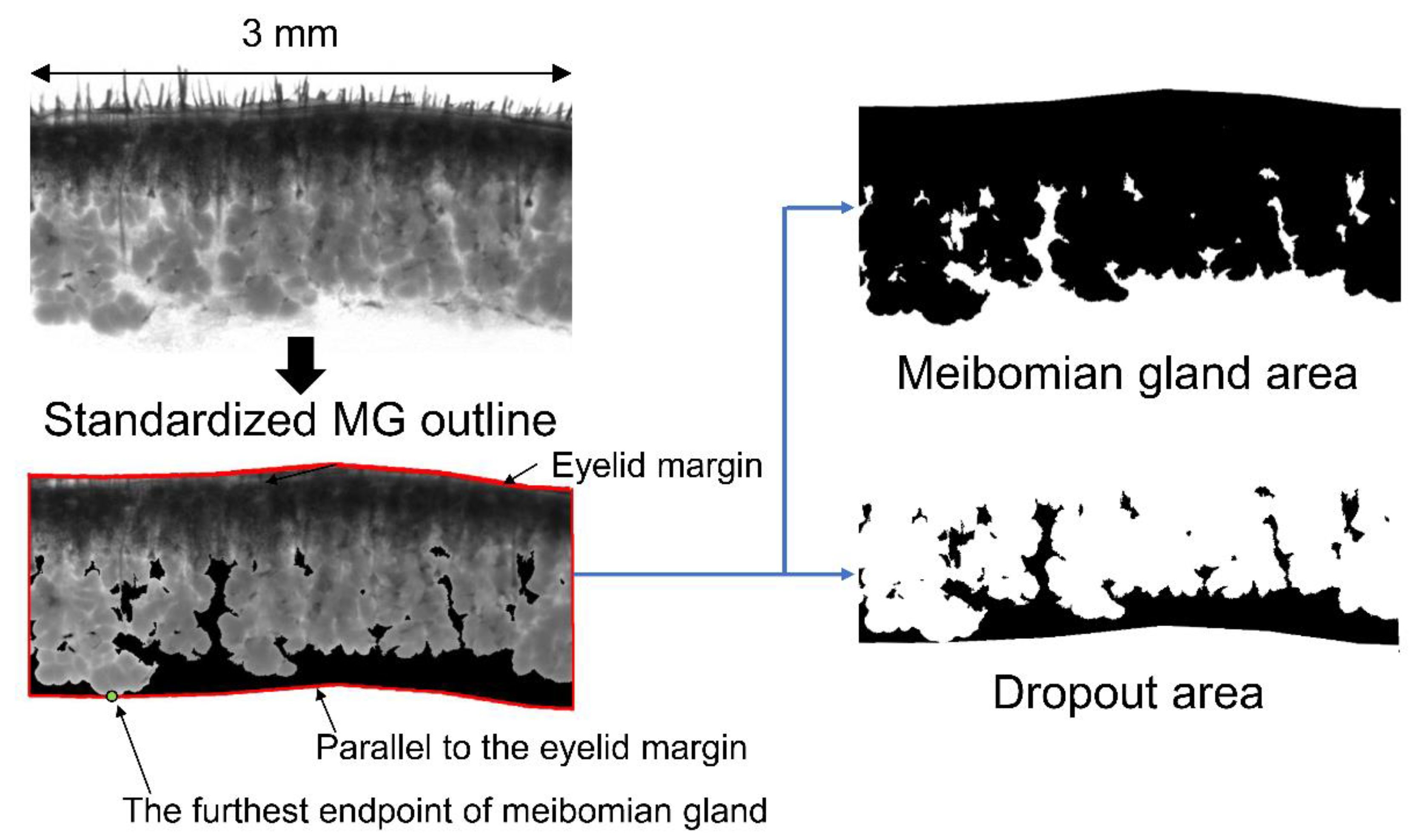
2.2. MG Dropout Area Increased with Aging, and Age-Adjusted Lower MG Area Was Negatively Correlated with the Severity of Corneal Epithelial Erosion
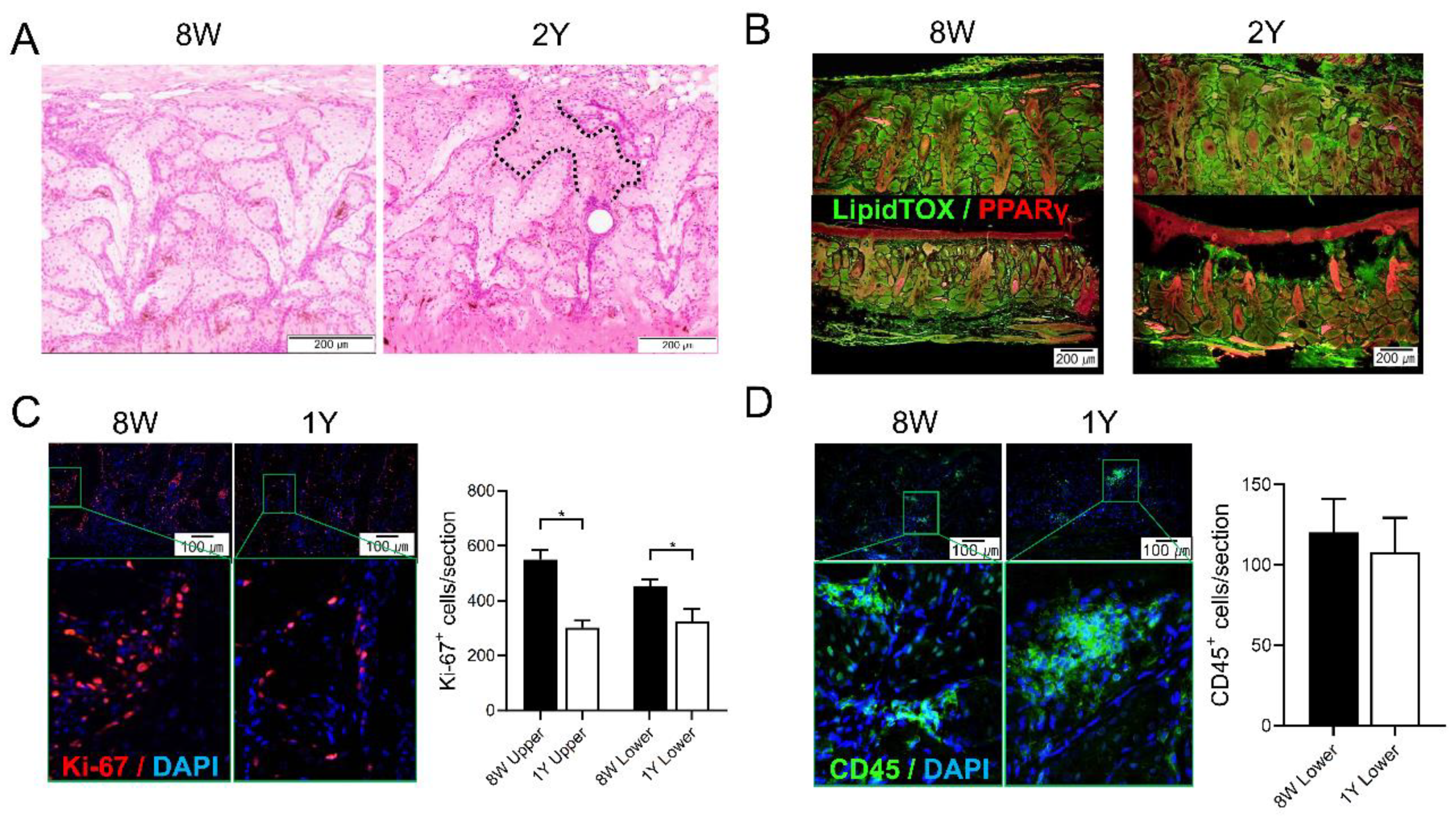
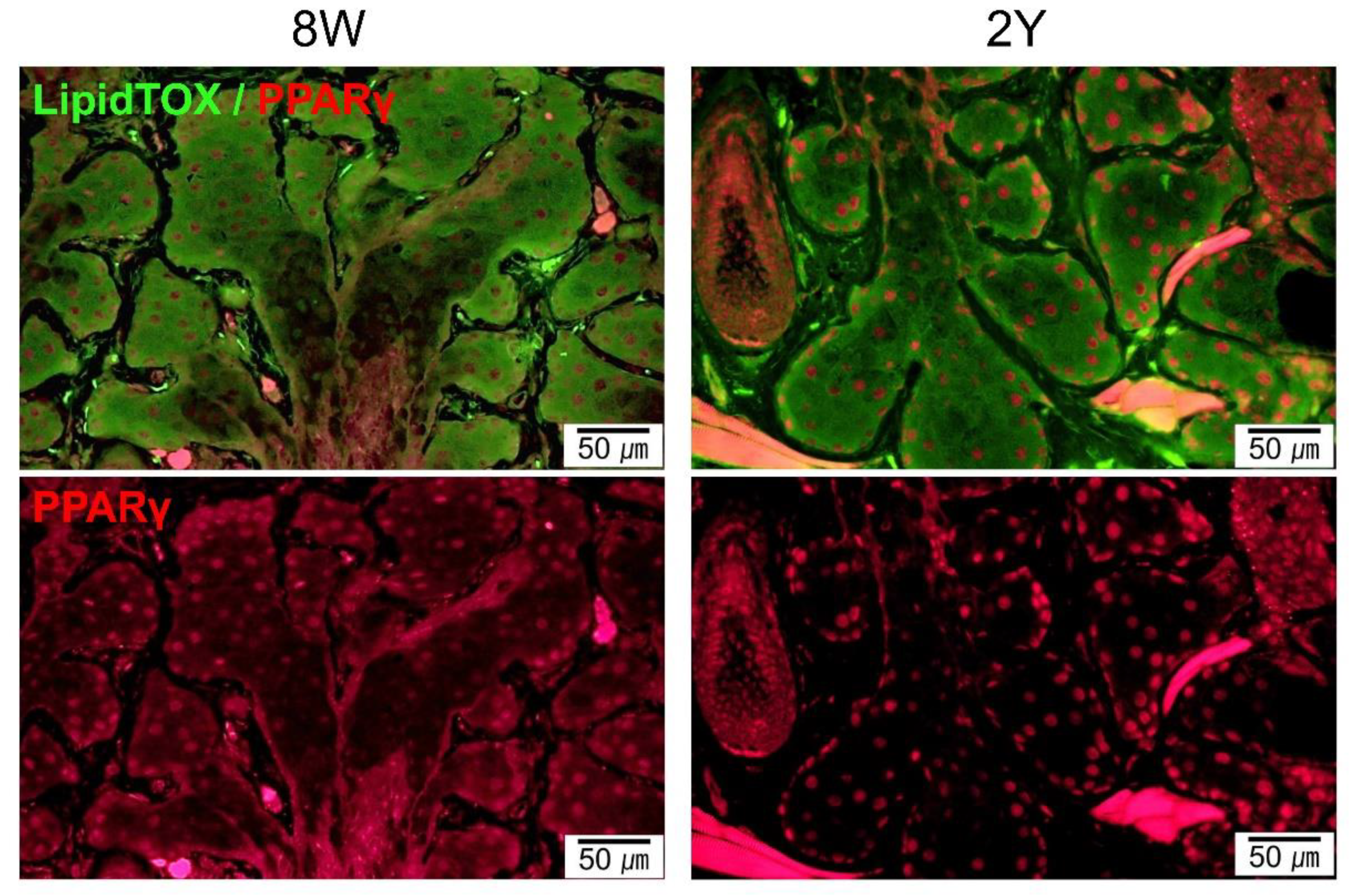
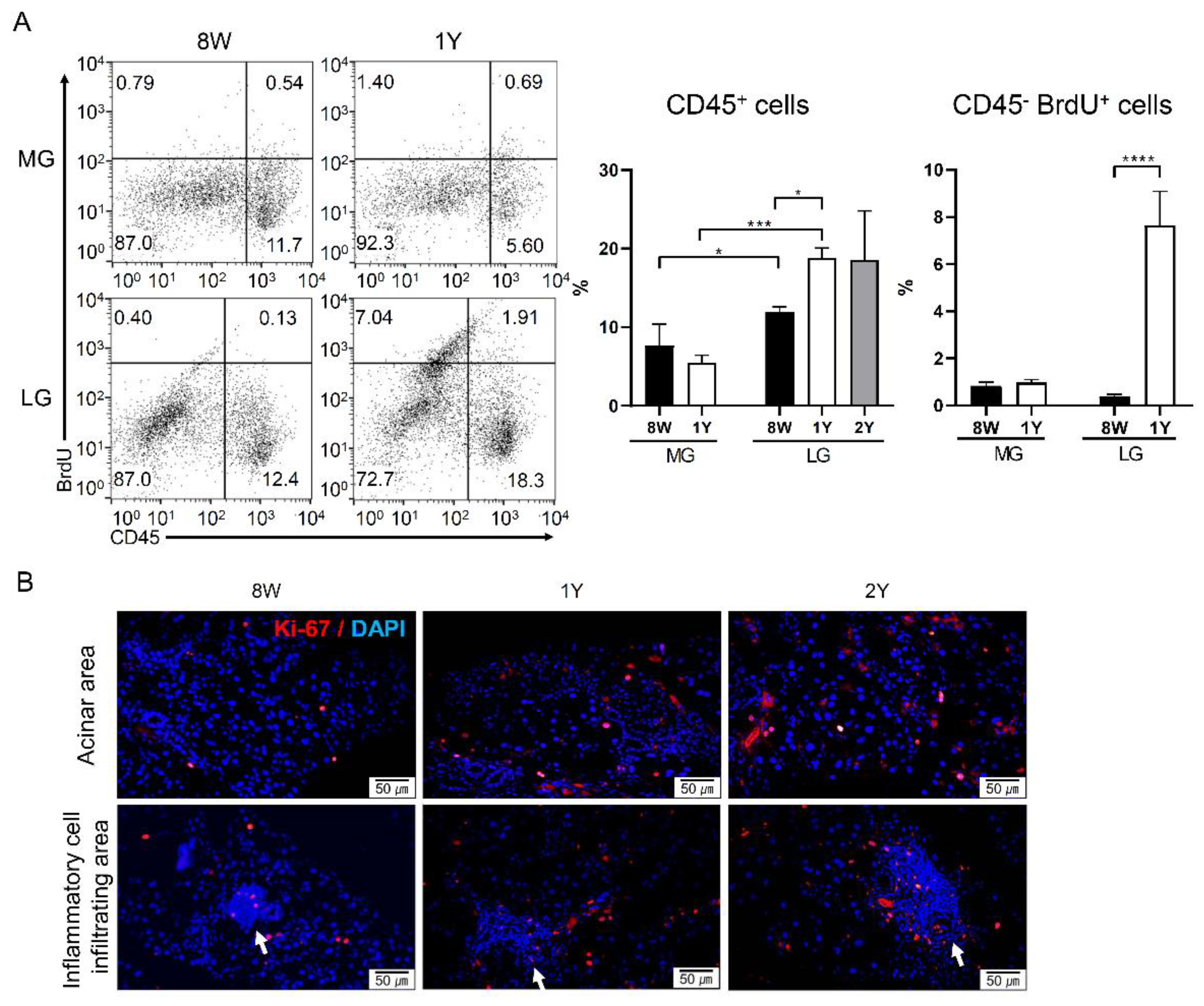
2.3. Aging-Dependent Inflammation Was Not Evident, but Decreased Potential of Differentiation Was Suspected in the MGs
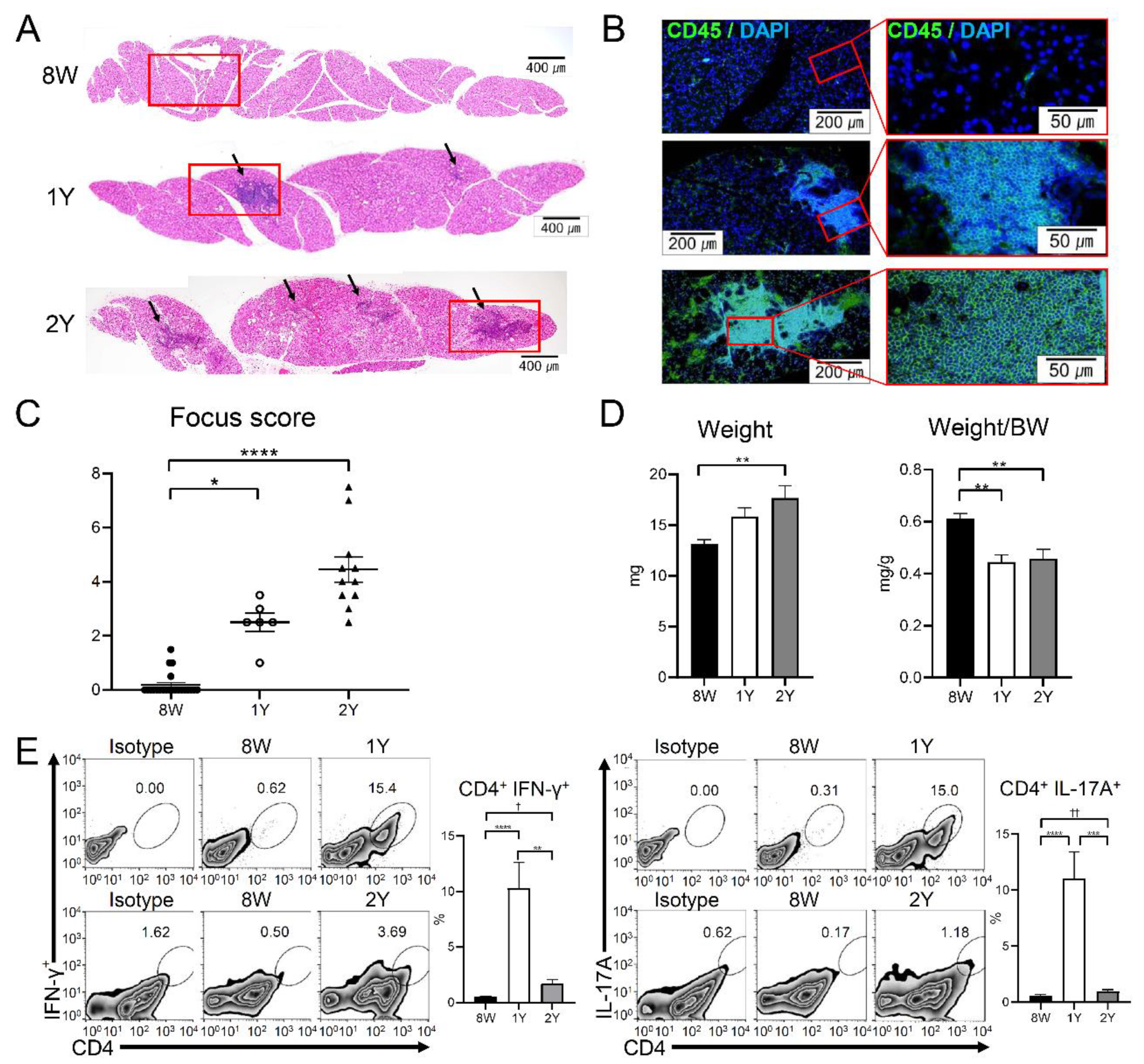
2.4. Aging-Dependent Inflammation Was Evident in the LGs
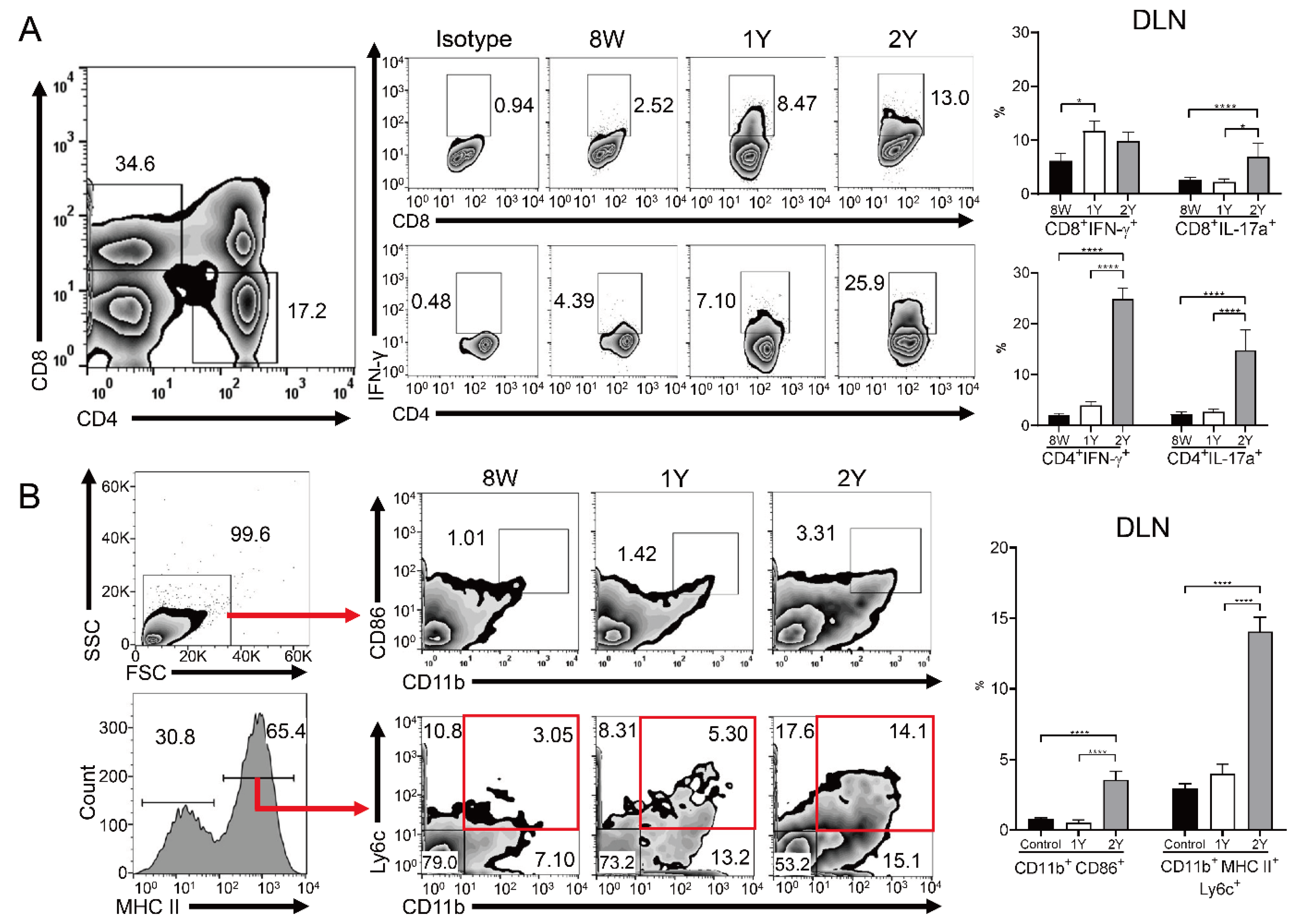
2.5. Effector T Cells and Activated Antigen-Presenting Cells Were Increased in the Drainage Lymph Nodes of Aged Mice
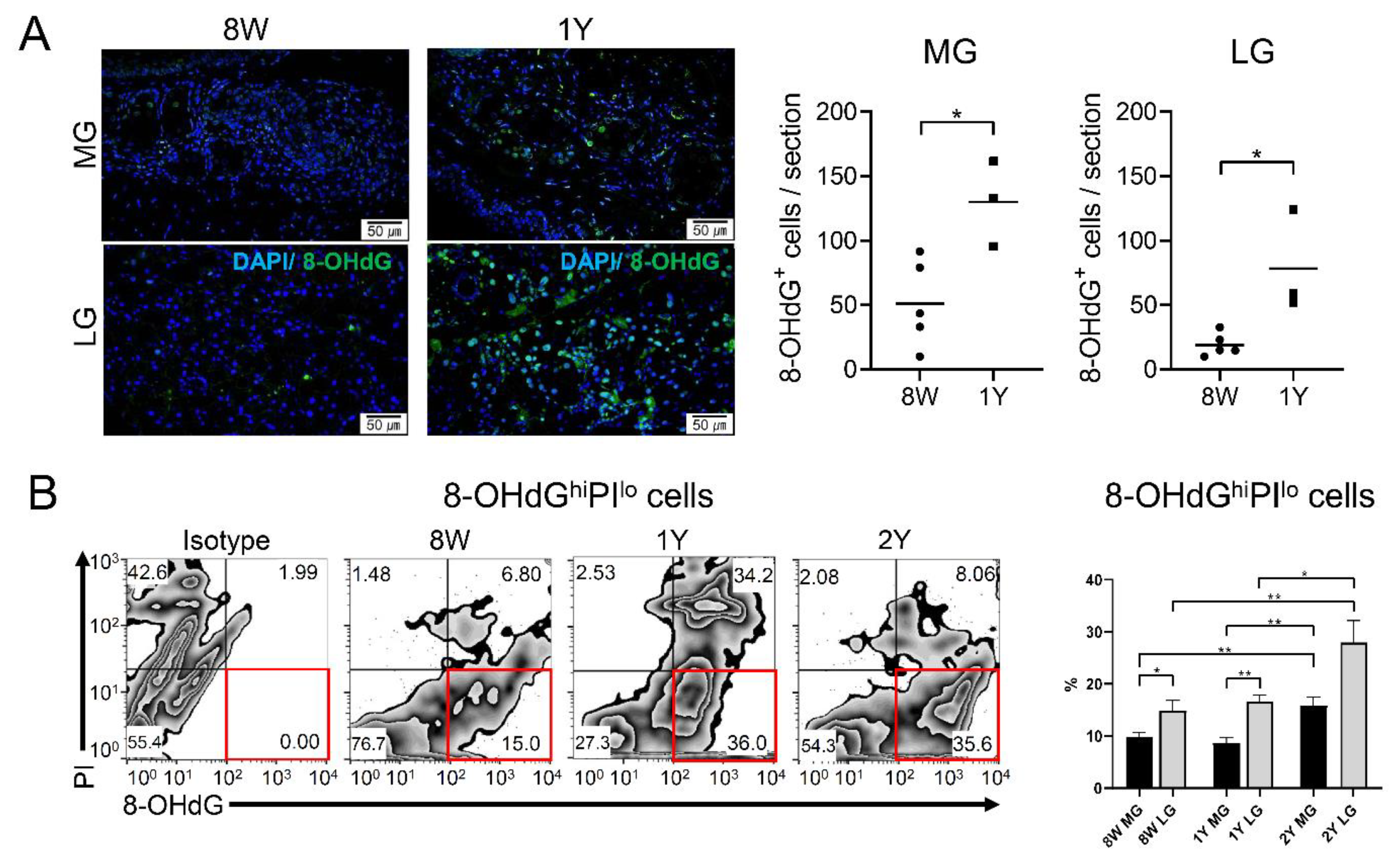
2.6. Oxidative Stress Increased in Both Glands of Aged Mice
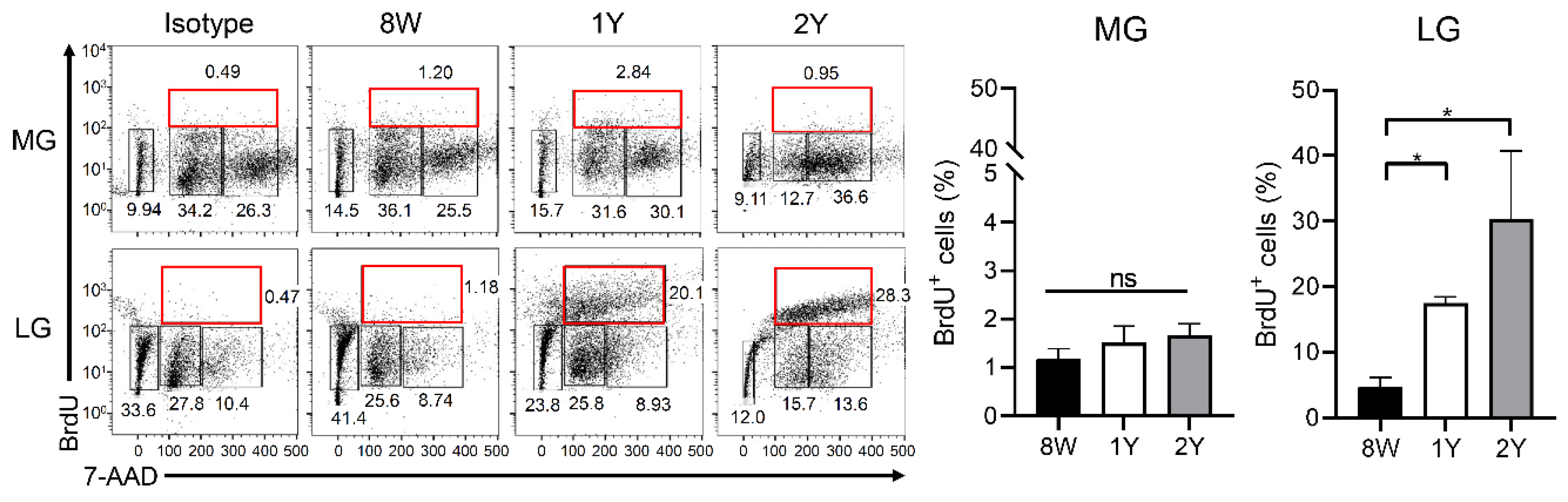
2.7. Senescence of Stem Cells Was Not Evident in Both Glands
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Clinical Evaluation
4.3. Histopathology
4.4. Transillumination Meibography of Mice
4.5. 5-Bromo-2′-Deoxyuridine (BrdU) Injection and Cell Cycle Analysis
4.6. Flow Cytometry for Immune Cells and Oxidatively Stressed Cells
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 1Y | One-year-old |
| 2Y | Two-year-old |
| 7-AAD | 7-aminoactinomycin D |
| 8W | Eight-week-old |
| BrdU | Bromo-2′-deoxyuridine |
| BW | Body weight |
| CCD | Charge-Coupled Device |
| DED | Dry eye disease |
| DLN | Drainage lymph node |
| H&E | Hematoxylin-eosin |
| IFN | Interferon |
| LG | Lacrimal gland |
| MHC | Major histocompatibility complex |
| MG | Meibomian gland |
| NEI | National Eye Institute |
| PAS | Periodic acid-Schiff |
| PI | Propidium iodide |
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Number of Samples | |||
|---|---|---|---|---|
| 8W | 1Y | 2Y | ||
| Dry eye assessment | Corneal staining (eyes) | 26 | 12 | 22 |
| Tear secretion (eyes) | 36 | 22 | 22 | |
| Conjunctival goblet cell count (eyes) | 12 | 6 | 3 | |
| Gross examination | MG area analyzed by meibography | 27 | 6 | 16 |
| LG weight | 8 | 6 | 6 | |
| MG analysis | Histology | |||
| H&E staining (coronal section) | 8 | 6 | 0 | |
| H&E staining, and LipidTOX/PPARγ IHC (coronal section) | 10 | 0 | 8 | |
| Ki-67 and CD45 IHC (coronal section) | 5 | 3 | 0 | |
| 8-OHdG IHC (sagittal section) | 5 | 3 | 0 | |
| Flow cytometry * | ||||
| 8-OHdGhiPIlo and BrdU+ | 4 | 0 | 3 | |
| 8-OHdGhiPIlo, BrdU+, and CD45+ | 5 | 5 | 0 | |
| LG analysis | Histology | |||
| H&E staining and CD45 IHC | 9 | 0 | 3 | |
| H&E staining, CD45 IHC, and Ki-67 IHC | 9 | 3 | 8 | |
| H&E staining, CD45 IHC, and 8-OHdG IHC | 4 | 3 | 0 | |
| 8-OHdG IHC | 1 | 0 | 0 | |
| Flow cytometry | ||||
| CD4+ IFNγ+ and CD4+ IL-17A+ | 9 | 6 | 0 | |
| 8-OHdGhiPIlo, BrdU+, and CD45+ | 4 | 5 | 0 | |
| CD4+ IFN-γ+, CD4+ IL-17A+, 8-OHdGhiPIlo, BrdU+, and CD45+ | 5 | 0 | 3 | |
| Secondary lymphoid organs | Flow cytometry of DLN † (mice) | 15 | 8 | 3 |
| Experiment | Included Groups | |
|---|---|---|
| Dry eye assessment | Corneal staining | 8W, 1Y, and 2Y |
| Tear secretion | 8W, 1Y, and 2Y | |
| Conjunctival goblet cell count | 8W, 1Y, and 2Y | |
| Gross examination | MG area analyzed by meibography | 8W, 1Y, and 2Y |
| LG weight | 8W, 1Y, and 2Y | |
| MG analysis | Histology | |
| H&E staining | 8W, 1Y, and 2Y | |
| LipidTOX/PPARγ IHC (coronal section) * | 8W and 2Y | |
| Ki-67 and CD45 IHC * | 8W and 1Y | |
| 8-OHdG IHC * | 8W and 1Y | |
| Flow cytometry | ||
| 8-OHdGhiPIlo | 8W, 1Y, and 2Y | |
| BrdU+/7-AAD | 8W, 1Y, and 2Y | |
| LG analysis | Histology | |
| H&E staining and CD45 IHC | 8W, 1Y, and 2Y | |
| Ki-67 IHC | 8W, 1Y, and 2Y | |
| 8-OHdG IHC * | 8W and 1Y | |
| Flow cytometry | ||
| CD4+ IFNγ+ and CD4+ IL-17A+ | 8W, 1Y, and 2Y | |
| 8-OHdGhiPIlo | 8W, 1Y, and 2Y | |
| BrdU+/7-AAD | 8W, 1Y, and 2Y | |
| Secondary lymphoid organs | Flow cytometry of DLN (mice) | 8W, 1Y, and 2Y |
References
- van Deursen, J.M. The role of senescent cells in ageing. Nature 2014, 509, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Muller, L.; Di Benedetto, S.; Pawelec, G. The Immune System and Its Dysregulation with Aging. Subcell. Biochem. 2019, 91, 21–43. [Google Scholar] [PubMed]
- Schaumberg, D.A.; Sullivan, D.A.; Buring, J.E.; Dana, M.R. Prevalence of dry eye syndrome among US women. Am. J. Ophthalmol. 2003, 136, 318–326. [Google Scholar] [CrossRef]
- Bian, F.; Xiao, Y.; Barbosa, F.L.; de Souza, R.G.; Hernandez, H.; Yu, Z.; Pflugfelder, S.C.; de Paiva, C.S. Age-associated antigen-presenting cell alterations promote dry-eye inducing Th1 cells. Mucosal Immunol. 2019, 12, 897–908. [Google Scholar] [CrossRef] [PubMed]
- de Souza, R.G.; de Paiva, C.S.; Alves, M.R. Age-related Autoimmune Changes in Lacrimal Glands. Immune Netw. 2019, 19, e3. [Google Scholar] [CrossRef] [PubMed]
- de Paiva, C.S. Effects of Aging in Dry Eye. Int. Ophthalmol. Clin. 2017, 57, 47–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nien, C.J.; Massei, S.; Lin, G.; Nabavi, C.; Tao, J.; Brown, D.J.; Paugh, J.R.; Jester, J.V. Effects of age and dysfunction on human meibomian glands. Arch. Ophthalmol. 2011, 129, 462–469. [Google Scholar] [CrossRef] [Green Version]
- Tsubota, K.; Kawashima, M.; Inaba, T.; Dogru, M.; Matsumoto, Y.; Ishida, R.; Kaido, M.; Kojima, T.; Uchino, M.; Uchino, Y.; et al. The antiaging approach for the treatment of dry eye. Cornea 2012, 31 (Suppl. 1), S3–S8. [Google Scholar] [CrossRef]
- Nasu, M.; Matsubara, O.; Yamamoto, H. Post-mortem prevalence of lymphocytic infiltration of the lacrymal gland: A comparative study in autoimmune and non-autoimmune diseases. J. Pathol. 1984, 143, 11–15. [Google Scholar] [CrossRef]
- Obata, H.; Yamamoto, S.; Horiuchi, H.; Machinami, R. Histopathologic study of human lacrimal gland. Statistical analysis with special reference to aging. Ophthalmology 1995, 102, 678–686. [Google Scholar] [CrossRef]
- Rocha, E.M.; Alves, M.; Rios, J.D.; Dartt, D.A. The aging lacrimal gland: Changes in structure and function. Ocul. Surf. 2008, 6, 162–174. [Google Scholar] [CrossRef] [Green Version]
- McClellan, A.J.; Volpe, E.A.; Zhang, X.; Darlington, G.J.; Li, D.Q.; Pflugfelder, S.C.; de Paiva, C.S. Ocular surface disease and dacryoadenitis in aging C57BL/6 mice. Am. J. Pathol. 2014, 184, 631–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Geest, K.S.; Abdulahad, W.H.; Tete, S.M.; Lorencetti, P.G.; Horst, G.; Bos, N.A.; Kroesen, B.J.; Brouwer, E.; Boots, A.M. Aging disturbs the balance between effector and regulatory CD4+ T cells. Exp. Gerontol. 2014, 60, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, V.; Rink, L.; Uciechowski, P. The Th17/Treg balance is disturbed during aging. Exp. Gerontol. 2013, 48, 1379–1386. [Google Scholar] [CrossRef]
- Paschides, C.A.; Petroutsos, G.; Psilas, K. Correlation of conjunctival impression cytology results with lacrimal function and age. Acta Ophthalmol. 1991, 69, 422–425. [Google Scholar] [CrossRef]
- Wei, A.; Hong, J.; Sun, X.; Xu, J. Evaluation of age-related changes in human palpebral conjunctiva and meibomian glands by in vivo confocal microscopy. Cornea 2011, 30, 1007–1012. [Google Scholar] [CrossRef]
- Dogru, M.; Kojima, T.; Simsek, C.; Tsubota, K.; Tsubota, K.; Kawashima, M.; Inaba, T.; Dogru, M.; Matsumoto, Y.; Ishida, R.; et al. Potential Role of Oxidative Stress in Ocular Surface Inflammation and Dry Eye Disease. The antiaging approach for the treatment of dry eye. Invest. Ophthalmol. Vis. Sci. 2018, 59, DES163–DES168. [Google Scholar] [CrossRef] [Green Version]
- Kojima, T.; Wakamatsu, T.H.; Dogru, M.; Ogawa, Y.; Igarashi, A.; Ibrahim, O.M.; Inaba, T.; Shimizu, T.; Noda, S.; Obata, H.; et al. Age-related dysfunction of the lacrimal gland and oxidative stress: Evidence from the Cu,Zn-superoxide dismutase-1 (Sod1) knockout mice. Am. J. Pathol. 2012, 180, 1879–1896. [Google Scholar] [CrossRef]
- Kawashima, M.; Tsubota, K. Effect of calorie restriction on change in lacrimal gland with age. Cornea 2011, 30 (Suppl. 1), S29–33. [Google Scholar] [CrossRef]
- You, S.; Tariq, A.; Kublin, C.L.; Zoukhri, D. Detection of BrdU-label retaining cells in the lacrimal gland: Implications for tissue repair. Cell Tissue Res. 2011, 346, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Eom, Y.; Choi, K.E.; Kang, S.Y.; Lee, H.K.; Kim, H.M.; Song, J.S. Comparison of meibomian gland loss and expressed meibum grade between the upper and lower eyelids in patients with obstructive meibomian gland dysfunction. Cornea 2014, 33, 448–452. [Google Scholar] [CrossRef]
- Pult, H. Relationships Between Meibomian Gland Loss and Age, Sex, and Dry Eye. Eye Contact Lens 2018, 44 (Suppl. 2), S318–S324. [Google Scholar] [CrossRef] [PubMed]
- Pult, H.; Riede-Pult, B.H.; Nichols, J.J. Relation between upper and lower lids’ meibomian gland morphology, tear film, and dry eye. Optom. Vis. Sci. 2012, 89, E310–315. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Menzies, K.; Sorbara, L.; Jones, L. Infrared imaging of meibomian gland structure using a novel keratograph. Optom. Vis. Sci. 2012, 89, 788–794. [Google Scholar] [CrossRef]
- Arita, R.; Minoura, I.; Morishige, N.; Shirakawa, R.; Fukuoka, S.; Asai, K.; Goto, T.; Imanaka, T.; Nakamura, M. Development of Definitive and Reliable Grading Scales for Meibomian Gland Dysfunction. Am. J. Ophthalmol. 2016, 169, 125–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, T.N.; Lin, M.C. Risk factors for severe Meibomian gland atrophy in a young adult population: A cross-sectional study. PLoS ONE 2017, 12, e0185603. [Google Scholar] [CrossRef] [Green Version]
- Ban, Y.; Shimazaki-Den, S.; Tsubota, K.; Shimazaki, J. Morphological evaluation of meibomian glands using noncontact infrared meibography. Ocul. Surf. 2013, 11, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Nien, C.J.; Paugh, J.R.; Massei, S.; Wahlert, A.J.; Kao, W.W.; Jester, J.V. Age-related changes in the meibomian gland. Exp. Eye Res. 2009, 89, 1021–1027. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Spiegelman, B.M. PPARgamma: A nuclear regulator of metabolism, differentiation, and cell growth. J. Biol. Chem. 2001, 276, 37731–37734. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPAR gamma is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef]
- Hwang, H.S.; Parfitt, G.J.; Brown, D.J.; Jester, J.V. Meibocyte differentiation and renewal: Insights into novel mechanisms of meibomian gland dysfunction (MGD). Exp. Eye Res. 2017, 163, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Jester, J.V.; Potma, E.; Brown, D.J. PPARgamma Regulates Mouse Meibocyte Differentiation and Lipid Synthesis. Ocul. Surf. 2016, 14, 484–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nocturne, G.; Mariette, X. B cells in the pathogenesis of primary Sjogren syndrome. Nat. Rev. Rheumatol. 2018, 14, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Shim, W.; Kim, C.E.; Choi, S.Y.; Lee, H.; Yang, J. Therapeutic Efficacy of Nanocomplex of Poly(Ethylene Glycol) and Catechin for Dry Eye Disease in a Mouse Model. Invest. Ophthalmol. Vis. Sci. 2017, 58, 1682–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, D.A.; Block, L.; Pena, J.D. Influence of androgens and pituitary hormones on the structural profile and secretory activity of the lacrimal gland. Acta Ophthalmol. 1996, 74, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Sato, E.H.; Sullivan, D.A. Comparative influence of steroid hormones and immunosuppressive agents on autoimmune expression in lacrimal glands of a female mouse model of Sjogren’s syndrome. Invest. Ophthalmol. Vis. Sci. 1994, 35, 2632–2642. [Google Scholar] [PubMed]
- Modulo, C.M.; Machado Filho, E.B.; Malki, L.T.; Dias, A.C.; de Souza, J.C.; Oliveira, H.C.; Jorge, I.C.; Santos Gomes, I.B.; Meyrelles, S.S.; Rocha, E.M. The role of dyslipidemia on ocular surface, lacrimal and meibomian gland structure and function. Curr. Eye Res. 2012, 37, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Kojima, T.; Nagata, T.; Kudo, H.; Muller-Lierheim, W.G.K.; van Setten, G.B.; Dogru, M.; Tsubota, K. The Effects of High Molecular Weight Hyaluronic Acid Eye Drop Application in Environmental Dry Eye Stress Model Mice. Int. J. Mol. Sci. 2020, 21, 3516. [Google Scholar] [CrossRef]
- Kim, J.; Choi, S.H.; Kim, Y.J.; Jeong, H.J.; Ryu, J.S.; Lee, H.J.; Kim, T.W.; Im, S.H.; Oh, J.Y.; Kim, M.K. Clinical Effect of IRT-5 Probiotics on Immune Modulation of Autoimmunity or Alloimmunity in the Eye. Nutrients 2017, 9, 1166. [Google Scholar] [CrossRef]
- De Roo, A.K.; Wouters, J.; Govaere, O.; Foets, B.; van den Oord, J.J. Identification of Circulating Fibrocytes and Dendritic Derivatives in Corneal Endothelium of Patients With Fuchs’ Dystrophy. Invest. Ophthalmol. Vis. Sci. 2017, 58, 670–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.H.; Kim, D.H.; Shin, E.J.; Lee, H.J.; Wee, W.R.; Jeon, S.; Kim, M.K. Comparative Analysis of Substrate-Free Cultured Oral Mucosal Epithelial Cell Sheets from Cells of Subjects with and without Stevens-Johnson Syndrome for Use in Ocular Surface Reconstruction. PLoS ONE 2016, 11, e0147548. [Google Scholar] [CrossRef] [PubMed]
- Subastri, A.; Suyavaran, A.; Preedia Babu, E.; Nithyananthan, S.; Barathidasan, R.; Thirunavukkarasu, C. Troxerutin with copper generates oxidative stress in cancer cells: Its possible chemotherapeutic mechanism against hepatocellular carcinoma. J. Cell Physiol. 2018, 233, 1775–1790. [Google Scholar] [CrossRef]
- Reyes, N.J.; Yu, C.; Mathew, R.; Kunnen, C.M.; Kalnitsky, J.; Redfern, R.L.; Leonardi, A.; Perez, V.L.; MacLeod, A.S.; Gupta, P.K.; et al. Neutrophils cause obstruction of eyelid sebaceous glands in inflammatory eye disease in mice. Sci. Transl. Med. 2018, 10, eaas9164. [Google Scholar] [CrossRef] [Green Version]
- Biosciences, B. Cell Cycle Analysis Using the BD BrdU FITC Assay on the BD FACSVerse™ System. Growth 2011, 2, G1. [Google Scholar]
- Lima, K.G.; Krause, G.C.; Schuster, A.D.; Catarina, A.V.; Basso, B.S.; De Mesquita, F.C.; Pedrazza, L.; Marczak, E.S.; Martha, B.A.; Nunes, F.B.; et al. Gallic acid reduces cell growth by induction of apoptosis and reduction of IL-8 in HepG2 cells. Biomed. Pharmacother. 2016, 84, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Paral, P.; Bajecny, M.; Savvulidi, F.; Necas, E. Cell Cycle Analysis Using In Vivo Staining of DNA-Synthesizing Cells. Methods Mol. Biol. 2019. [Google Scholar] [CrossRef]
- Braumuller, H.; Wieder, T.; Brenner, E.; Assmann, S.; Hahn, M.; Alkhaled, M.; Schilbach, K.; Essmann, F.; Kneilling, M.; Griessinger, C.; et al. T-helper-1-cell cytokines drive cancer into senescence. Nature 2013, 494, 361–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.K.; Kim, K.W.; Ryu, J.S.; Jeong, H.J.; Lee, S.M.; Kim, M.K. Bilateral Effect of the Unilateral Corneal Nerve Cut on Both Ocular Surface and Lacrimal Gland. Invest. Ophthalmol. Vis. Sci. 2019, 60, 430–441. [Google Scholar] [CrossRef]
- Cambi, M.; Tamburrino, L.; Marchiani, S.; Olivito, B.; Azzari, C.; Forti, G.; Baldi, E.; Muratori, M. Development of a specific method to evaluate 8-hydroxy, 2-deoxyguanosine in sperm nuclei: Relationship with semen quality in a cohort of 94 subjects. Reproduction 2013, 145, 227–235. [Google Scholar] [CrossRef] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, C.H.; Ryu, J.S.; Hwang, H.S.; Kim, M.K. Comparative Analysis of Age-Related Changes in Lacrimal Glands and Meibomian Glands of a C57BL/6 Male Mouse Model. Int. J. Mol. Sci. 2020, 21, 4169. https://doi.org/10.3390/ijms21114169
Yoon CH, Ryu JS, Hwang HS, Kim MK. Comparative Analysis of Age-Related Changes in Lacrimal Glands and Meibomian Glands of a C57BL/6 Male Mouse Model. International Journal of Molecular Sciences. 2020; 21(11):4169. https://doi.org/10.3390/ijms21114169
Chicago/Turabian StyleYoon, Chang Ho, Jin Suk Ryu, Ho Sik Hwang, and Mee Kum Kim. 2020. "Comparative Analysis of Age-Related Changes in Lacrimal Glands and Meibomian Glands of a C57BL/6 Male Mouse Model" International Journal of Molecular Sciences 21, no. 11: 4169. https://doi.org/10.3390/ijms21114169