Identification of circRNA-Associated-ceRNA Networks Involved in Milk Fat Metabolism under Heat Stress


Abstract
:1. Introduction
2. Results
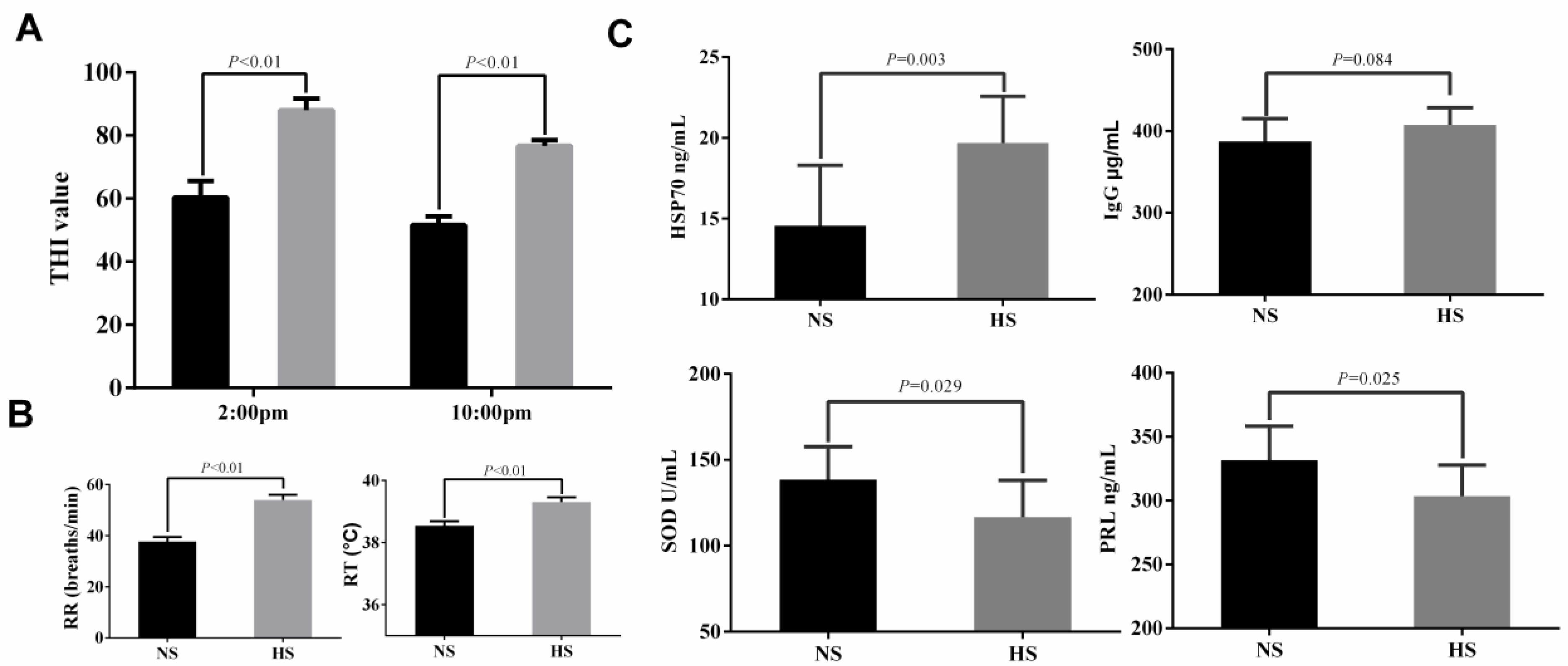
2.1. The Performances of Dairy Cows under Heat Stress
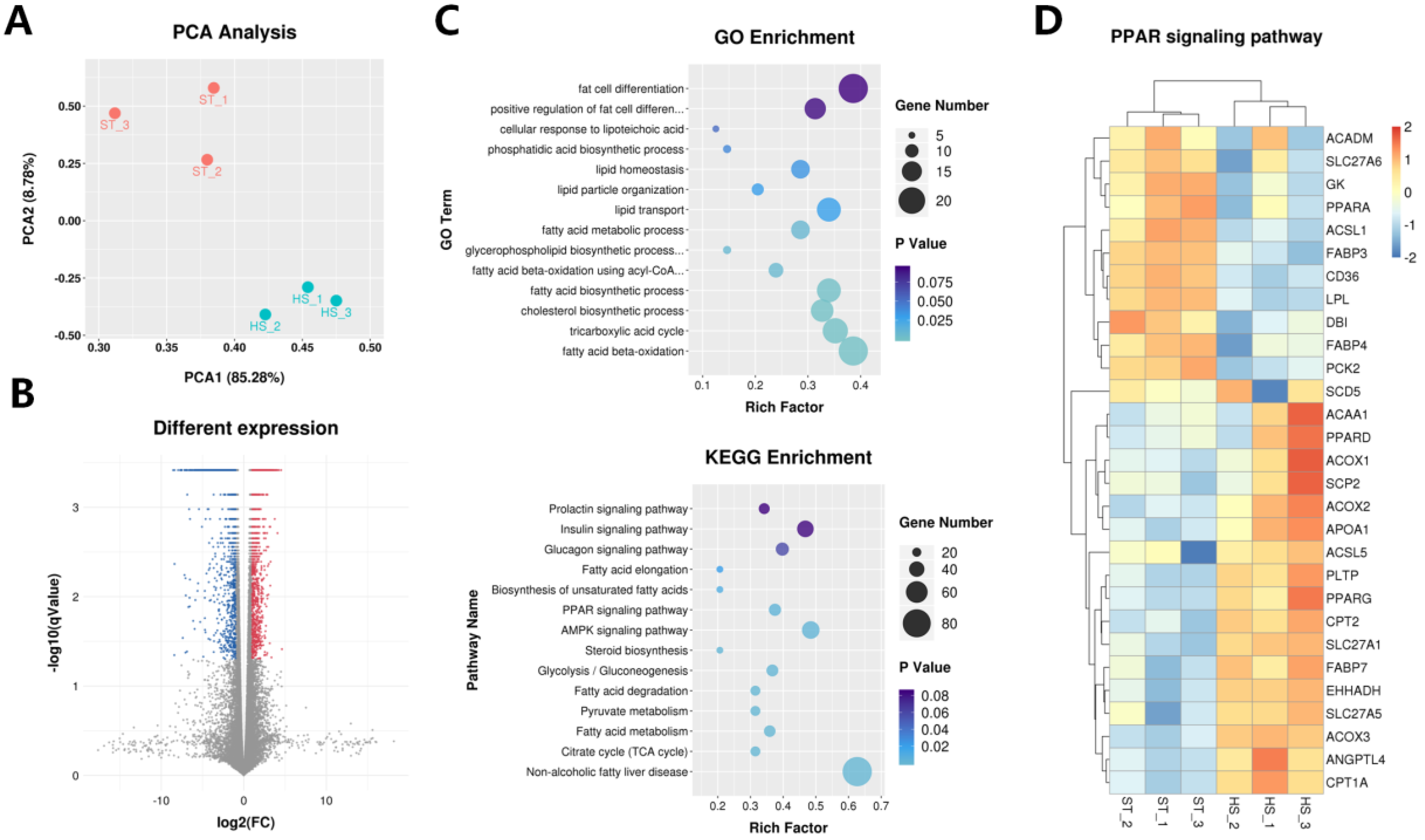
2.2. RNA Sequencing and Transcript Analysis
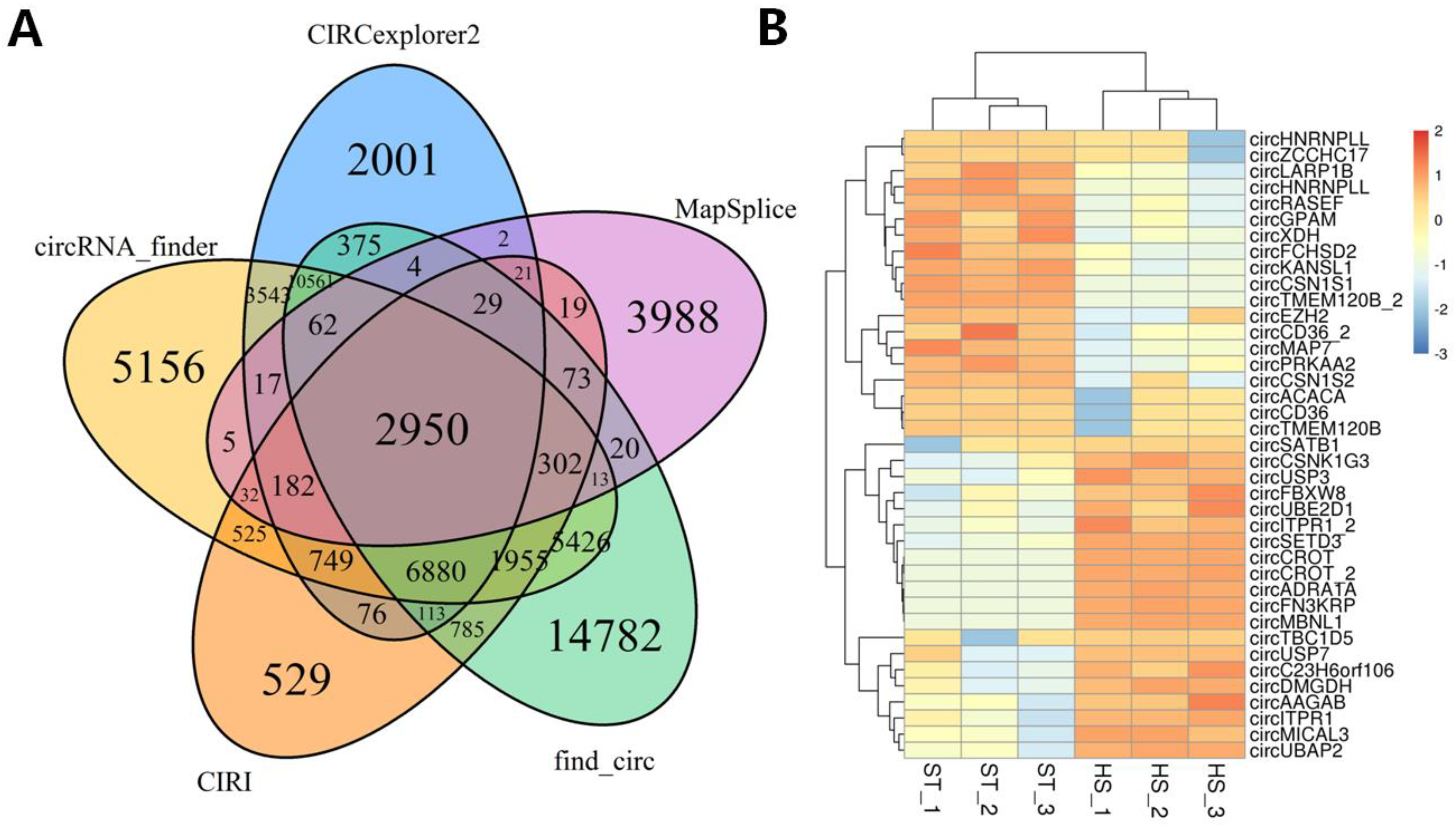
2.3. Identification of circRNAs
2.4. Functional Interactions between circRNAs and mRNAs
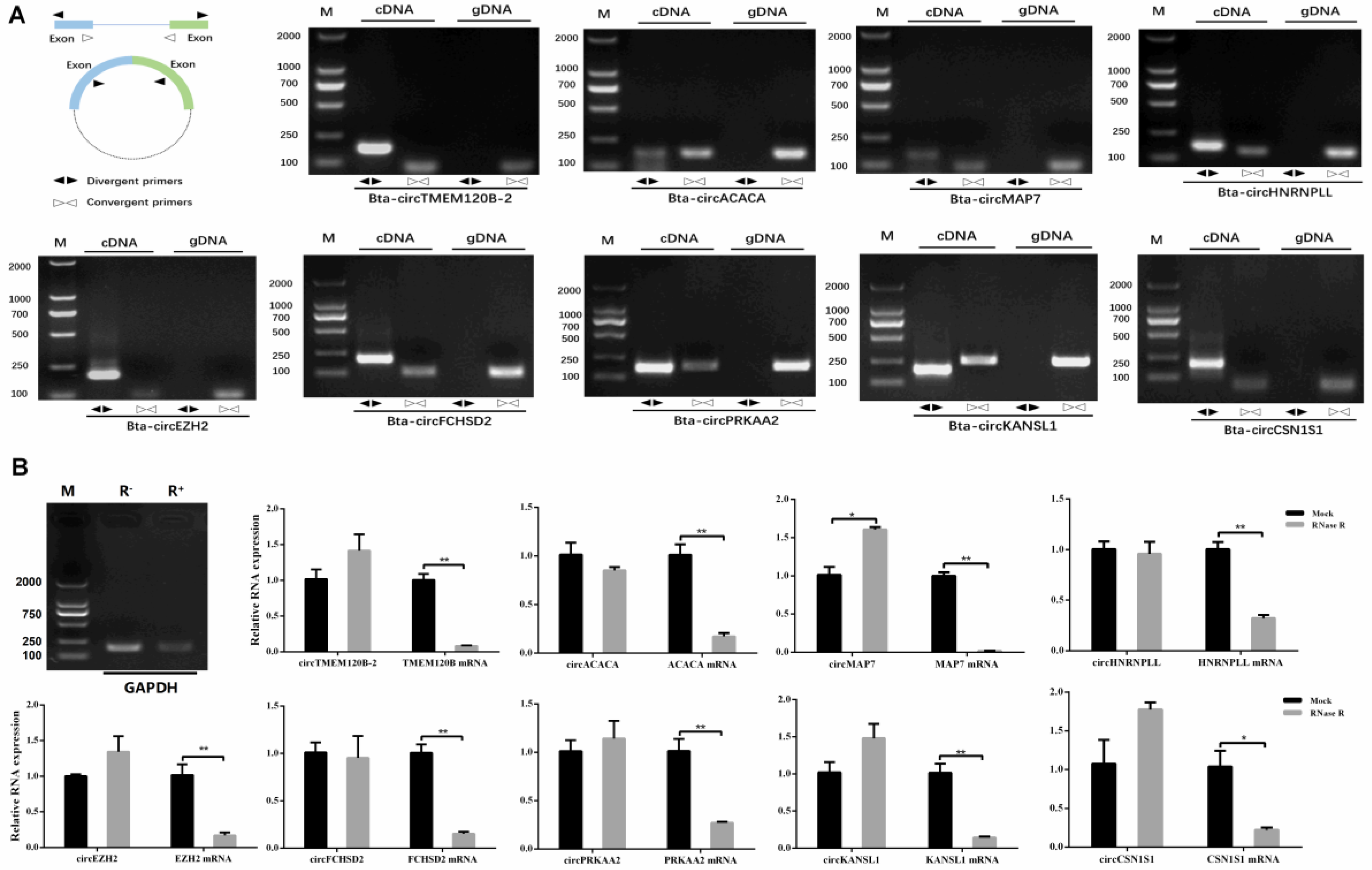
2.5. Characterization of circRNA
3. Discussion
4. Materials and methods
4.1. Experimental Design and Sample Collection
4.2. RNA Preparation and Sequencing
4.3. Bioinformatic Analysis
4.4. Validation of circRNAs
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| HSP70 | Heat shock protein 70 |
| SOD | Superoxide dismutase |
| PRL | Prolactin |
| GO | Gene ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| ncRNA | Non-coding RNAs |
| THI | Temperature-humidity index |
| FPKM | Fragments Per Kilobase of transcript per Million |
| CoA | Coenzyme A |
| PPAR | Peroxisome proliferative activated receptor |
| PPARG | Peroxisome proliferative activated receptor, gamma |
| SCD | Stearoyl-CoA desaturase |
References
- Luber, G.; McGeehin, M. Climate change and extreme heat events. Am. J. Prev. Med. 2008, 35, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Nardone, A.; Ronchi, B.; Lacetera, N.; Ranieri, M.S.; Bernabucci, U. Effects of climate changes on animal production and sustainability of livestock systems. Livest. Sci. 2010, 130, 57–69. [Google Scholar] [CrossRef]
- Kadzere, T.C.; Murphy, M.R.; Silanikove, N.; Maltz, E. Heat stress in lactating dairy cows: A review. Livest. Prod. Sci. 2002, 77, 59–91. [Google Scholar] [CrossRef]
- Carabaño, J.M.; Logar, B.; Bormann, J.; Minet, J.; Vanrobays, M.-L.; Díaz, C.; Tychon, B.; Gengler, N.; Hammami, H. Modeling heat stress under different environmental conditions. J. Dairy Sci. 2016, 99, 3798–3814. [Google Scholar] [CrossRef] [Green Version]
- Borad, G.S.; Kumar, A.; Singh, A.K. Effect of processing on nutritive values of milk protein. Crit. Rev. Food Sci. Nutr. 2017, 57, 3690–3702. [Google Scholar] [CrossRef]
- Hill, L.D.; Wall, E. Dairy cattle in a temperate climate: The effects of weather on milk yield and composition depend on management. Animal 2015, 9, 138–149. [Google Scholar] [CrossRef] [Green Version]
- Bernabucci, U.; Basirico, L.; Morera, P.; Dipasquale, D.; Vitali, A.; Cappelli, F.P.; Calamari, L. Effect of summer season on milk protein fractions in Holstein cows. J. Dairy Sci. 2015, 98, 1815–1827. [Google Scholar] [CrossRef] [Green Version]
- Rhoads, L.M.; Rhoads, R.P.; Vanbaale, M.J.; Collier, R.J.; Sanders, S.R.; Weber, W.J.; Crooker, B.A.; Baumgard, L.H. Effects of heat stress and plane of nutrition on lactating Holstein cows: I. Production, metabolism, and aspects of circulating somatotropin. J. Dairy Sci. 2009, 92, 852–864. [Google Scholar] [CrossRef] [Green Version]
- Pierre St, N.R.; Cobanov, B.; Schnitkey, G. Economic losses from heat stress by US livestock industries. J. Dairy Sci. 2003, 86, E52–E77. [Google Scholar] [CrossRef] [Green Version]
- Fuquay, J.W. Heat stress as it affects animal production. J. Anim. Sci. 1981, 52, 164–174. [Google Scholar] [CrossRef]
- West, W.J.; Mullinix, B.G.; Bernard, J.K. Effects of hot, humid weather on milk temperature, dry matter intake, and milk yield of lactating dairy cows. J. Dairy Sci. 2003, 86, 232–242. [Google Scholar] [CrossRef] [Green Version]
- Wheelock, B.J.; Rhoads, R.P.; Vanbaale, M.J.; Sanders, S.R.; Baumgard, L.H. Effects of heat stress on energetic metabolism in lactating Holstein cows. J. Dairy Sci. 2010, 93, 644–655. [Google Scholar] [CrossRef]
- Bernabucci, U.; Biffani, S.; Buggiotti, L.; Vitali, A.; Lacetera, N.; Nardone, A. The effects of heat stress in Italian Holstein dairy cattle. J. Dairy. Sci. 2014, 97, 471–486. [Google Scholar] [CrossRef] [PubMed]
- Leroy, L.J.; Rizos, D.; Sturmey, R.; Bossaert, P.; Gutierrez-Adan, A.; van Hoeck, V.; Valckx, S.; Bols, P.E. Intrafollicular conditions as a major link between maternal metabolism and oocyte quality: A focus on dairy cow fertility. Reprod. Fertil. Dev. 2012, 24, 1. [Google Scholar] [CrossRef] [PubMed]
- Cowley, C.F.; Barber, D.G.; Houlihan, A.V.; Poppi, D.P. Immediate and residual effects of heat stress and restricted intake on milk protein and casein composition and energy metabolism. J. Dairy Sci. 2015, 98, 2356–2368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Tian, S.Z.H.; Zhou, X.; Zhang, Y.; Li, S.; Yang, H.; Zheng, N.; Wang, J. Metabolic responses and “omics” technologies for elucidating the effects of heat stress in dairy cows. Int. J. Biometeorol. 2017, 61, 1149–1158. [Google Scholar]
- Vujanac, I.; Kirovski, D.; Šamanc, H.; Prodanovic, R.; Lakic, N.; Adamovic, M.; Valcic, O. Milk production in high-yielding dairy cows under different environment temperatures. Large Anim. Rev. 2012, 18, 31–36. [Google Scholar]
- Zielniok, K.; Sobolewska, A.; Gajewska, M. Mechanisms of autophagy induction by sex steroids in bovine mammary epithelial cells. J. Mol. Endocrinol. 2017, 59, 29–48. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, H.; Wang, Y.; Zhao, Y.; Fang, X.; Chen, C.; Chen, H. Expression patterns of circular RNAs from primary kinase transcripts in the mammary glands of lactating rats. J. Mammary Cancer 2015, 18, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, D.; Wang, Y.; Li, M.; Fang, X.; Chen, H.; Zhang, C. The landscape of circular RNAs and mRNAs in bovine milk exosomes. J. Food Compos. Anal. 2019, 76, 33–38. [Google Scholar] [CrossRef]
- Li, X.; Yang, L.; Chen, L. The Biogenesis, Functions, and Challenges of Circular RNAs. Mol. Cell 2018, 71, 428–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patop, L.I.; Kadener, S. circRNAs in Cancer. Curr. Opin. Genet. Dev. 2018, 48, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Sun, X.; Liu, Y.; Li, H.; Deng, G.; Lin, H.; Wang, S. Heat stress alters genome-wide profiles of circular RNAs in Arabidopsis. Plant Mol. Biol. 2018, 96, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Palmquist, D.L. Milk Fat: Origin of Fatty Acids and Influence of Nutritional Factors Thereon. In Advanced Dairy Chemistry Volume 2: Lipids; Springer: Boston, MA, USA, 2006; pp. 43–92. [Google Scholar]
- Bionaz, M.; Loor, J.J. ACSL1, AGPAT6, FABP3, LPIN1, and SLC27A6 Are the Most Abundant Isoforms in Bovine Mammary Tissue and Their Expression is affected by stage of lactation. J. Nutr. 2008, 138, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Min, L.; Cheng, J.B.; Shi, B.L.; Yang, H.J.; Zheng, N.; Wang, J.Q. Effects of heat stress on serum insulin, adipokines, AMP-activated protein kinase, and heat shock signal molecules in dairy cows. J. Zhejiang Univ. Sci. B 2015, 16, 541–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellagi, R.; Martin, B.; Chassaing, C.; Najar, T.; Pomiès, D. Evaluation of heat stress on tarentaise and holstein cow performance in the mediterranean climate. Int. J. Biometeorol. 2017, 61, 1371–1379. [Google Scholar] [CrossRef]
- Preez, J.H.D. Parameters for the determination and evaluation of heat stress in dairy cattle in South Africa. Onderstepoort J. Vet. Res. 2001, 67, 263–271. [Google Scholar]
- Dikmen, S.; Hansen, P.J. Is the temperature-humidity index the best indicator of heat stress in lactating dairy cows in a subtropical environment. J. Dairy Sci. 2009, 92, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Park, J.C.; Seo, Y.S. Heat Shock Proteins: A Review of the Molecular Chaperones for Plant Immunity. Plant Pathol. J. 2015, 31, 323–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimoto, I.R.; Sarge, K.D.; Abravaya, K. Transcriptional regulation of heat shock genes. A paradigm for inducible genomic responses. J. Biol. Chem. 1992, 267, 21987. [Google Scholar]
- Sabirzhanov, B.; Stoica, B.A.; Hanscom, M.; Piao, C.S.; Faden, A.I. Over-expression of HSP70 attenuates caspase-dependent and caspase-independent pathways and inhibits neuronal apoptosis. J. Neurochem. 2012, 123, 542–554. [Google Scholar] [CrossRef] [PubMed]
- Dangi, S.S.; Gupta, M.; Dangi, S.K.; Chouhan, V.S.; Maurya, V.P.; Kumar, P.; Singh, G.; Sarkar, M. Expression of HSPs: An adaptive mechanism during long-term heat stress in goats (Capra hircus). Int. J. Biometeorol. 2015, 59, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tang, S.; Yin, B.; Sun, J.R.; Song, E.B.; Bao, E.D. Co-enzyme Q10 and acetyl salicylic acid enhance Hsp70 expression in primary chicken myocardial cells to protect the cells during heat stress. Mol. Cell. Biochem. 2017, 435, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Sarup, P.; Dahlgaard, J.; Norup, A.M.; Rgensen, K.T.J.; Hebsgaard, M.B.; Loeschcke, V. Down Regulation of Hsp70 Expression Level Prolongs the Duration of Heat-Induced Male Sterility in Drosophila buzzatii. Funct. Ecolog. 2004, 18, 365–370. [Google Scholar] [CrossRef]
- Sun, L.L.; Gao, S.T.; Wang, K.; Xu, J.C. Effects of source on bioavailability of selenium, antioxidant status, and performance in lactating dairy cows during oxidative stress-inducing conditions. J. Dairy Sci. 2018, 102, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Yasui, T.; Mcart, J.A.A.; Ryan, C.M.; Gilbert, R.O.; Nydam, D.V.; Valdez, F.; Griswold, K.E.; Overton, T.R. Effects of chromium propionate supplementation during the periparturient period and early lactation on metabolism, performance, and cytological endometritis in dairy cows. J. Dairy Sci. 2014, 97, 6400–6410. [Google Scholar] [CrossRef]
- Song, J.H.; Xue, Q.L.; Bing, S.Z.; Jun, G.W. Response of the Photosynthesis and Antioxidant Systems to High-Temperature Stress in Euonymus japonicus Seedlings. For. Sci. 2010, 56, 172–180. [Google Scholar]
- Zhang, J.F.; Weng, X.G.; Wang, J.F.; Zhou, D.; Zhang, W.; Zhai, C.C.; Hou, Y.X.; Zhu, Y.H. Effects of temperature-humidity index and chromium supplementation on antioxidant capacity, heat shock protein 72, and cytokine responses of lactating cows. J. Anim. Sci. 2014, 92, 3026–3034. [Google Scholar] [CrossRef]
- Caroprese, M.; Marzano, A.; Entrican, G.; Wattegedera, S.; Albenzio, M.; Sevi, A. Immune response of cows fed polyunsaturated fatty acids under high ambient temperatures. J. Dairy Sci. 2009, 92, 2796–2803. [Google Scholar] [CrossRef] [Green Version]
- Tao, S.; Monteiro, A.P.A.; Thompson, I.M.; Hayen, M.J.; Dahl, G.E. Effect of late-gestation maternal heat stress on growth and immune function of dairy calves. J. Dairy Sci. 2012, 95, 7128–7136. [Google Scholar] [CrossRef] [Green Version]
- Tao, S.; Bubolz, J.W.; Amaral, B.C.D.; Thompson, I.M.; Hayen, M.J.; Johnson, S.E.; Dahl, G.E. Effect of heat stress during the dry period on mammary gland development. J. Dairy Sci. 2011, 94, 5976–5986. [Google Scholar] [CrossRef] [PubMed]
- Lacasse, P.; Lollivier, V.; Dessauge, F.; Bruckmaier, R.M.; Ollier, S.; Boutinaud, M. New developments on the galactopoietic role of prolactin in dairy ruminants. Domest. Anim. Endocrinol. 2012, 43, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Ravagnolo, O.; Misztal, I. Genetic component of heat stress in dairy cattle, parameter estimation. J. Dairy Sci. 2000, 83, 2126–2130. [Google Scholar] [CrossRef]
- Segnalini, M.; Nardone, A.; Bernabucci, U.; Vitali, A.; Ronchi, B.; Lacetera, N. Dynamics of the temperature-humidity index in the Mediterranean basin. Int. J. Biometeorol. 2011, 55, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Habeeb, A.A.G.; Tarabany, A.; Atta, M. Negative effects of heat stress on growth and milk production off arm animals. J. Anim. Husb. Dairy Sci. 2018, 2, 1–12. [Google Scholar]
- Hammami, H.; Bormann, J.; Hamdi, N.M.; Montaldo, H.H.; Gengler, N. Evaluation of Heat Stress Effects on Production Traits and Somatic Cell Score of Holsteins in a Temperate Environment. J. Dairy Sci. 2013, 96, 1844–1855. [Google Scholar] [CrossRef] [Green Version]
- Beede, K.D.; Sheare, J.K. Nutritional Management of Dairy Cattle During Hot Weather. IV. Agri-Practice 1991, 12, 5–13. [Google Scholar]
- Chornobai, A.C.; Damasceno, J.C.; Visentainer, J.V.; Matsushit, M. Physical-chemical composition of in natura goat milk from cross Saanen throughout lactation period. Arch. Latinoam. Nutr. 1999, 49, 283–286. [Google Scholar]
- Umberto, B.; Nicola, L.; Bruno, R.; Alessandroal, N. Effects of the hot season on milk protein fractions\rin Holstein cows. Anim. Res. 2002, 51, 25–33. [Google Scholar]
- Rhoads, R.E.; Grudzien-Nogalska, E. Translational regulation of milk protein synthesis at secretory activation. J. Mammary Gland Biol. Neoplasia 2007, 12, 283–292. [Google Scholar] [CrossRef]
- Shwartz, G.; Rhoads, M.L.; Vanbaale, M.J.; Rhoads, R.P.; Baumgard, L.H. Effects of a supplemental yeast culture on heat-stressed lactating Holstein cows. J. Dairy Sci. 2009, 92, 935–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goode, F.H.; Webster, N.R. Free radicals and antioxidants in sepsis. Crit. Care Med. 1993, 21, 1770–1776. [Google Scholar] [CrossRef] [PubMed]
- Christon, R.; Saminadin, G.; Lionet, H.; Racon, B. Dietary fat and climate alter food intake, performance of lactating sows and their litters and fatty acid composition of milk. Anim. Sci. 1999, 69, 353–365. [Google Scholar] [CrossRef]
- Gao, T.S.; Guo, J.; Quan, S.Y.; Nan, X.M.; Fernandez, M.V.S.; Baumgand, L.H.; Bu, D.P. The effects of heat stress on protein metabolism in lactating Holstein cows. J. Dairy Sci. 2017, 100, 5040–5049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garner, B.J.; Douglas, M.; Williams, S.R.O.; Wales, B. Responses of dairy cows to short-term heat stress in controlled-climate chambers. Anim. Prod. Sci. 2017, 57, 1233–1241. [Google Scholar] [CrossRef]
- Collier, J.R.; Collier, J.L.; Rhoads, R.P.; Baumgard, L.H. Invited review: Genes involved in the bovine heat stress response. J. Dairy Sci. 2008, 91, 445–454. [Google Scholar] [CrossRef]
- Wickramasinghe, S.; Rincon, G.; Islas-Trejo, A.; Medrano, F.J. Transcriptional profiling of bovine milk using RNA sequencing. BMC Genomics 2012, 13, 45. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Zhu, J.; Luo, J.; Cao, W.; Shi, H.; Yao, D.; Li, J.; Sun, Y.; Xu, H.; Yu, K.; et al. Genes regulating lipid and protein metabolism are highly expressed in mammary gland of lactating dairy goats. Funct. Integr. Genom. 2015, 15, 309–321. [Google Scholar] [CrossRef]
- Lemay, G.D.; Ballard, A.O.; Hughes, A.M.; Morrow, L.A.; Horseman, D.N.; Nommsen-Rivers, A.L. RNA sequencing of the human milk fat layer transcriptome reveals distinct gene expression profiles at three stages of lactation. PLoS ONE 2013, 8, e67531. [Google Scholar] [CrossRef]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine milk fat synthesis during the lactation cycle. BMC Genom. 2008, 9, 366–370. [Google Scholar] [CrossRef] [Green Version]
- Chong, M.B.; Reigan, P.; Mayle-Combs, K.D.; Orlicky, D.J.; Mcmanaman, J.L. Determinants of adipophilin function in milk lipid formation and secretion. Trends Endocrinol. Metab. 2011, 22, 211–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadegowda, G.A.K.; Bionaz, M.; Piperova, L.S.; Erdman, R.A.; Loor, J.J. Peroxisome proliferator-activated receptor-γ activation and long-chain fatty acids alter lipogenic gene networks in bovine mammary epithelial cells to various extents. J. Dairy Sci. 2009, 92, 4276–4289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macciotta, P.N.P.; Mele, M.; Conte, G.; Serra, A.; Cassandro, M.; Cassandro, R.; Zotto, R.D.; Borlino, A.C.; Pagnacco, G.; Secchiari, P. Association Between a Polymorphism at the Stearoyl CoA Desaturase Locus and Milk Production Traits in Italian Holsteins. J. Dairy Sci. 2008, 91, 3184–3189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, W.W.; Yang, W.; Chen, Y.; Wu, Z.K. Foxo3 circular RNA promotes cardiac senescence by modulating multiple factors associated with stress and senescence responses. Eur. Heart J. 2017, 38, 1402–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulcheski, R.F.; Christoff, A.P.; Margis, R. Circular RNAs are miRNA sponges and can be used as a new class of biomarker. J. Biotechnol. 2016, 238, 42–51. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, H.; Wang, Y.; Zhu, S.; Liu, J.; Fang, X.; Chen, H. Circular RNA of cattle casein genes are highly expressed in bovine mammary gland. J. Dairy Sci. 2016, 99, 4750–4760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Jay, A.; Brunaldi, K.; Huang, N.; Hamilton, J.A. CD36 Enhances Fatty Acid Uptake by Increasing the Rate of Intracellular Esterification but Not Transport across the Plasma Membrane. Biochemistry 2013, 52, 7254–7261. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Yang, X.; Li, X.; Wang, J.; Gao, Y.; Shang, R.; Sun, W.; Dou, K.; Li, H. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015, 3201565, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. Embnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Singh, D.; Zeng, Z.; Coleman, S.J.; Coleman, S.J.; Huang, Y.; Savich, G.L.; He, X.; Mieczkowski, P.; Grimm, S.A.; et al. MapSplice: Accurate mapping of RNA-seq reads for splice junction discovery. Nucleic Acids Res. 2010, 38, e178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Liu, R. CircRNAFinder: A tool for identifying circular RNAs using RNA-Seq data. In Proceedings of the 6th International Conference on Bioinformatics and Computational Biology, BICOB, Las Vegas, NV, USA, 4–6 April 2014. [Google Scholar]
- Gao, Y.; Wang, J.; Zhao, F. CIRI: An efficient and unbiased algorithm for de novo circular RNA identification. Genome Biol. 2015, 16, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, O.X.; Dong, R.; Zhang, Y.; Zhang, J.L.; Luo, Z.; Zhang, J.; Chen, L.L.; Yang, L. Diverse alternative back-splicing and alternative splicing landscape of circular RNAs. Genome Res. 2016, 26, 1277–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, O.X.; Wang, H.B.; Zhang, Y.; Lu, X.; Chen, L.L.; Yang, L. Complementary sequence-mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, D.M.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedländer, R.M.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. Mirdeep2 accurately identifies known and hundreds of novel microrna genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | NS | HS |
|---|---|---|
| Milk yield kg/day | 22.83 ± 0.2251 A | 19.06 ± 0.1643 B |
| Protein % | 3.46 ± 0.0336 A | 3.30 ± 0.0329 B |
| Fat % | 3.71 ±0.0352 A | 3.55 ± 0.0253 B |
| Lactose % | 4.69 ± 0.0292 a | 4.58 ± 0.0318 b |
| Somatic cell counts 104/mL | 30.47 ± 0.5833 B | 37.83 ± 0.7445 A |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Chen, Z.; Zhuang, X.; Luo, J.; Chen, T.; Xi, Q.; Zhang, Y.; Sun, J. Identification of circRNA-Associated-ceRNA Networks Involved in Milk Fat Metabolism under Heat Stress. Int. J. Mol. Sci. 2020, 21, 4162. https://doi.org/10.3390/ijms21114162
Wang D, Chen Z, Zhuang X, Luo J, Chen T, Xi Q, Zhang Y, Sun J. Identification of circRNA-Associated-ceRNA Networks Involved in Milk Fat Metabolism under Heat Stress. International Journal of Molecular Sciences. 2020; 21(11):4162. https://doi.org/10.3390/ijms21114162
Chicago/Turabian StyleWang, Dongyang, Zujing Chen, Xiaona Zhuang, Junyi Luo, Ting Chen, Qianyun Xi, Yongliang Zhang, and Jiajie Sun. 2020. "Identification of circRNA-Associated-ceRNA Networks Involved in Milk Fat Metabolism under Heat Stress" International Journal of Molecular Sciences 21, no. 11: 4162. https://doi.org/10.3390/ijms21114162