The HD-ZIP II Transcription Factors Regulate Plant Architecture through the Auxin Pathway

1
College of Life Sciences, Northwest A & F University, Yangling, Shaanxi 712100, China
2
National Key Facility for Crop Gene Resources and Genetic Improvement, Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2020, 21(9), 3250; https://doi.org/10.3390/ijms21093250
Submission received: 12 March 2020
/
Revised: 29 April 2020
/
Accepted: 2 May 2020
/
Published: 4 May 2020
(This article belongs to the Section Molecular Plant Sciences)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The homeodomain-leucine zipper (HD-ZIP) family transcription factors play important roles in plant growth and development. However, the underlying mechanisms remain largely unclear. Here we found that ATHB2, encoding a HD-ZIP transcription factor, is an early auxin responsive gene. Phenotypic analyses show that overexpression of ATHB2 impairs plant architecture, including reduced plant height and small leaves, and also reduces auxin response in leaves when grown in soil. Simultaneously, the seedlings with chemical induction of ATHB2 exhibit abnormal root gravitropism, a typical auxin-related phenotype. We further show that the auxin response pattern is altered in roots of the inducible ATHB2 seedlings. Consistently, the transcript levels of some auxin biosynthetic and transport genes are significantly decreased in these transgenic seedlings. Further, protein and promoter sequence analyses in common wheat showed that the HD-ZIP II subfamily transcription factors have highly conserved motifs and most of these encoding gene promoters contain the canonical auxin-responsive elements. Expression analyses confirm that some of these HD-ZIP II genes are indeed regulated by auxin in wheat. Together, our results suggest that the HD-ZIP II subfamily transcription factors regulate plant development possibly through the auxin pathway in plants.
1. Introduction
The plant hormone auxin is important for regulating almost all aspects of plant growth and development [1]. Auxin receptors perceive the auxin concentration changes and then initiate auxin signaling. Nuclear auxin signal perception and consequent alterations in gene expression are carried out by the TRANSPORT INHIBITOR RESPONSE1/AUXIN SIGNALING F-BOX (TIR1/AFB) pathway [2,3]. Three major protein families are involved in the TIR1/AFB pathway: auxin-binding TIR1/AFB F-box proteins, AUXIN/INDOLE-3-ACETIC ACID INDUCIBLE (Aux/IAA) repressor proteins, and AUXIN RESPONSE FACTOR (ARF) transcription factors [3,4]. In this pathway, the Aux/IAAs interact with ARFs and inhibit ARF transcription factor activity when the auxin level is low [5]. When the auxin level is increased, the Aux/IAA and TIR1/AFB F-box proteins form a co-receptor complex, which are components of a Skp1-Cullin-F-box (SCF) E3 ubiquitin ligase complex, resulting in ubiquitination and degradation of the Aux/IAA repressor proteins by the 26S proteasome [1,5,6]. Recently, Cao et al. (2019) identified another auxin signaling mechanism, which acts in parallel to the canonical auxin pathway [7]. This signaling mechanism works at the concave side of the apical hook. The auxin-mediated C-terminal cleavage of transmembrane kinase 1 (TMK1) is involved in this signaling [7].
Two distinct pathways are involved in auxin distribution in the plant body: passive diffusion through the plasma membrane (PM) and directional cell-to-cell polar auxin transport (PAT) [8]. PAT is a precise auxin distribution manner that is significantly important for the formation of local auxin maxima, mainly in developing tissues [8,9]. The differential localization of transporters at specific sites on the PM creates a directional auxin flow, which eventually establishes a PAT stream through adjacent cells [9]. AUXIN RESISTANT1 (AUX1)/LIKE-AUX1 (LAX) proteins are auxin influx transporters and PIN-FORMED (PIN) proteins are auxin efflux transporters [10,11,12,13,14,15]. Other auxin transports, which have nonpolar distribution, consist of the P-GLYCOPROTEIN (PGP), MULTIDRUG RESISTANCE (MDR), or ATP-BINDING CASSETTE SUBFAMILY B (ABCB). AUX/LAX regulates several developmental processes, such as lateral root formation (AUX1 and LAX3) and cotyledon vascular patterning (LAX2) [15,16]. Among the distinct auxin transports, the polar localizations of PIN proteins within cells finely correspond to the directionality of auxin flow, which highlights that PIN proteins that are mainly responsible for the asymmetric distributions of auxin in plants [16,17,18]. The homeodomain-leucine zipper (HD-ZIP) transcription factors are unique to plants and contribute to various plant physiological processes [19,20,21,22]. Based on evolutionary relationship and protein structure, HD-ZIP transcription factors are generally divided into four subfamilies, namely HD-ZIP I, HD-ZIP II, HD-ZIP III and HD-ZIP IV [23]. HD-ZIP II proteins can be stimulated by far red light, and then lead to shade avoidance response [24,25]. Previous studies showed that HD-ZIP II proteins respond to various biotic and abiotic stresses, through regulating hormone signaling pathways and expression of related genes [26,27,28]. A recent study showed that HD-ZIP II proteins regulate shoot apical meristems (SAM) maintenance and cotyledon polarity through auxin flow, but there is no evidence to suggest how HD-ZIP II proteins regulate auxin perception or auxin response [29,30].
In this study, we identified an early auxin responsive gene, ATHB2, which is one of the HD-ZIP II subfamily proteins in Arabidopsis. Phenotypic observation showed that overexpression of ATHB2 impaired plant architecture, including reduced plant height and small leaves and decreased auxin response in leaves when grown in soil. Meanwhile, ATHB2-inducible expression led to a typical auxin-related phenotype known as the agravitropic root phenotype. Furthermore, confocal microscopic analyses showed that asymmetric auxin response occurred in roots of ATHB2-overexpressing seedlings. We also found that the expression levels of some auxin biosynthetic and transport genes were reduced in the iATHB2 seedlings. Further, we found that most of the HD-ZIP II subfamily gene promoters in common wheat contain the canonical auxin-responsive elements. Expression analyses confirm that some of these HD-ZIP II genes are indeed regulated by auxin in wheat. Together, our results suggest that the HD-ZIP II subfamily transcription factors regulate plant development possibly through the auxin pathway in plants.
2. Results
2.1. ATHB2 Is an Early Auxin-Inducible Gene
ATHB2 belongs to a member of the HD-ZIP II subfamily transcription factors in Arabidopsis. A previous study showed that the three proteins ATHB2, HAT3 and ATHB4 redundantly regulate embryo development in plants [30]. However, the underlying mechanism remains largely unknown. To investigate whether the biological role of ATHB2 and its homologs (Figure S1) is related to the auxin pathway, the 6 day old wild-type (Col-0) seedlings were treated with 10 μM IAA for different time points. The quantitative reverse transcription polymerase chain reaction (qRT-PCR) results showed that the transcript levels of the ATHB2 gene increased to a maximum level about three-fold at 0.5 h after exogenous IAA treatment (Figure 1A). The auxin-mediated induction of ATHB2 expression was substantially reduced after 1 h of treatment (Figure 1A). Similarly, the transcript levels of HAT1 and HAT2 genes were also obviously elevated after IAA treatment for 0.5 h (Figure 1B,C). By contrast, the transcriptional expression of HAT3 and AHTB4 was downregulated by IAA treatment (Figure 1D,E). The Aux/IAA genes are well known as the early auxin responsive genes and participate in auxin signaling through interacting with ARFs as transcriptional repressors. As a control, the transcriptional expression of IAA19, one representative member of the Aux/IAA family, was rapidly increased at 0.5 h after exogenous IAA treatment and reached a maximum at 1 h (Figure 1F). Taken together, like Aux/IAA genes, ATHB2 and its close homologs HAT1 and HAT2 are early auxin responsive genes.
2.2. Overexpression of ATHB2 Affected Plant Architecture
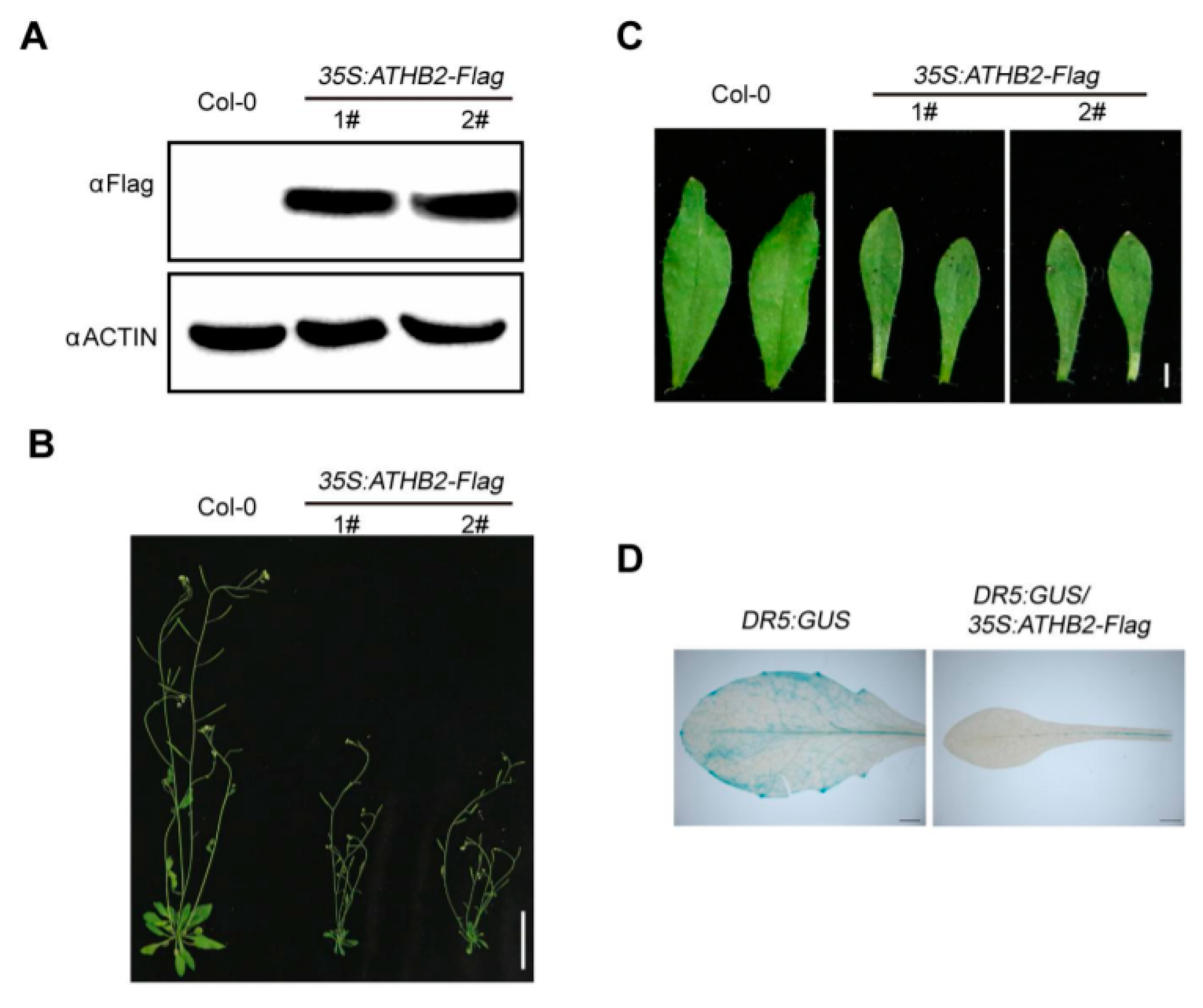
To observe the morphological phenotypes of ATHB2 overexpression plants at the adult stage, we generated different 35S:ATHB2-FLAG transgenic lines constitutively overexpressing ATHB2. Western blotting analyses demonstrated that the ATHB2-FLAG fusion proteins accumulated in the 35S:ATHB2-FLAG transgenic seedlings (Figure 2A). Phenotypic analyses showed that the 4-week-old ATHB2 overexpressing plants under normal growth conditions exhibited auxin-related phenotypes, such as dwarfism and narrow leaf phenotypes (Figure 2B,C). To explore how ATHB2 interacts with auxin to differentially regulate leaf development in the Col-0 and 35S:ATHB2-FLAG transgenic plants, we examined the spatial distribution of the auxin response in Col-0 and 35S:ATHB2-FLAG transgenic plants using the auxin-responsive reporter DR5:GUS [31]. The double transgenic plant DR5:GUS/35S:ATHB2-FLAG was generated through genetic crossing between DR5:GUS and 35S:ATHB2-FLAG plants. DR5:GUS and DR5:GUS/35S:ATHB2-FLAG plants were used for this experiment. As shown in Figure 2D, the expression of DR5:GUS in the 4-week-old leaves of DR5:GUS/35S:ATHB2-FLAG plants was significantly decreased compared with that in the wild type (Figure 2D). These results showed that constitutive overexpression of ATHB2 reduced auxin response and affected leaf development.
2.3. Inducible-Expression of ATHB2 Led to Auxin-Related Root Phenotypes
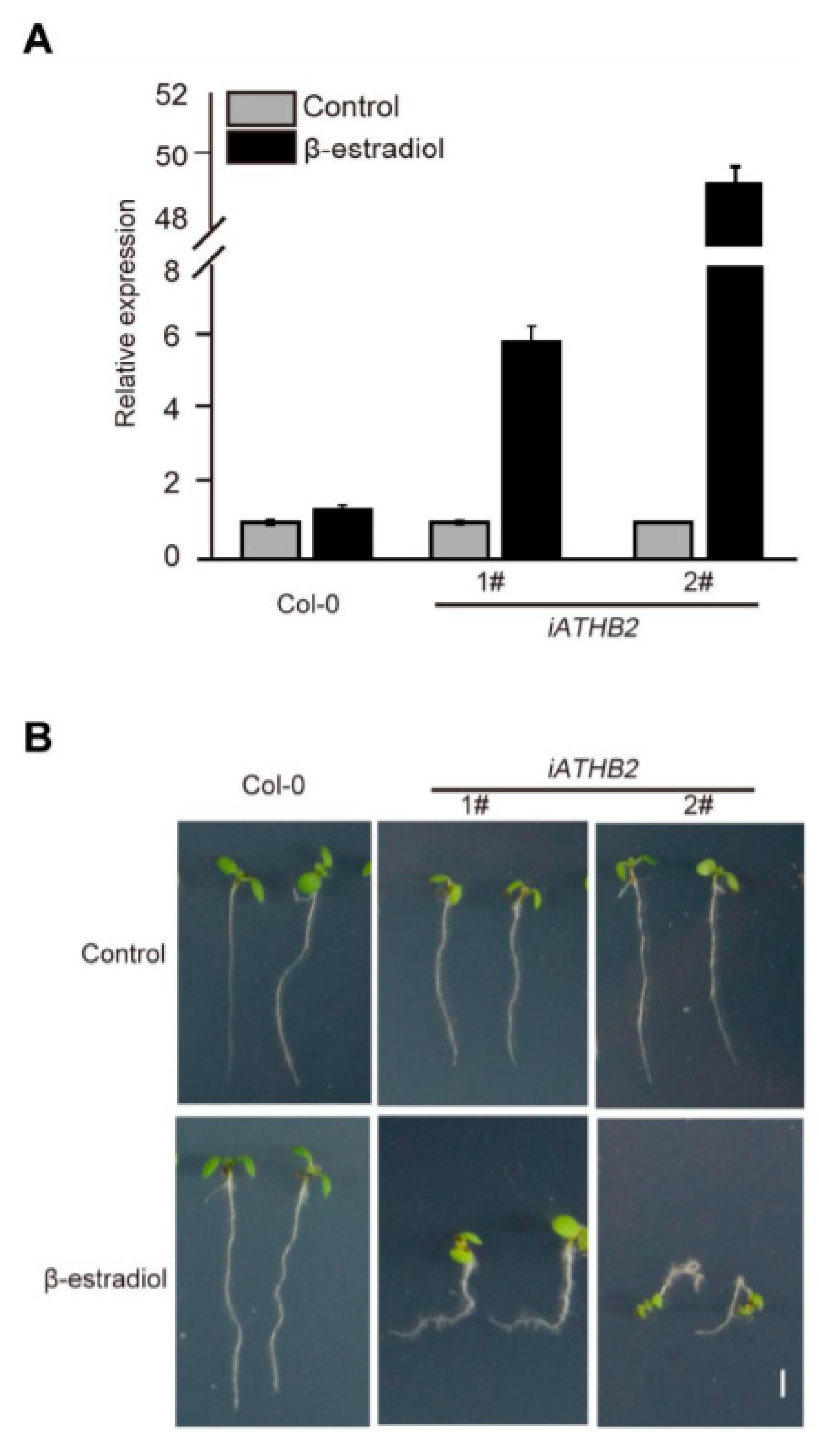
In order to determine the biological relevance of ATHB2 in auxin-mediated physiological processes, the β-estradiol-inducible ATHB2 overexpressing plants (XVE>>ATHB2, simply labeled as iATHB2) were generated. The qRT-PCR results showed that the transcript levels of ATHB2 were significantly increased in the inducible transgenic lines after β-estradiol treatment for 2 h compared with the Col-0 seedlings (Figure 3A). Notably, the transcript levels of ATHB2 were greatly upregulated by about 60-fold in the iATHB2 2# transgenic line (Figure 3A).
To observe the auxin-related phenotype of iATHB2 transgenic lines, the 6-day-old iATHB2 seedlings were grown on 1/2 Murashige and Skoog (MS) medium with or without 10 μM β-estradiol. In the absence of β-estradiol, the growth phenotype of iATHB2 and Col-0 seedlings was comparable. However, two independent iATHB2 transgenic lines displayed an agravitropic root phenotype grown on the medium containing 10 μM β-estradiol, whereas the root growth of Col-0 plants was not affected in the presence of 10 μM β-estradiol (Figure 3B). In addition, the primary root length analyses were performed using Col-0 and iATHB2 transgenic seedlings grown on the medium containing 10 μM β-estradiol. As shown in Figure S2, the primary root length of iATHB2 transgenic seedlings was significantly reduced compared with that of Col-0 seedlings. These results suggest that ATHB2 might be involved in the auxin-mediated root development.
2.4. Inducible-Expression of ATHB2 Altered Auxin Distribution in Roots
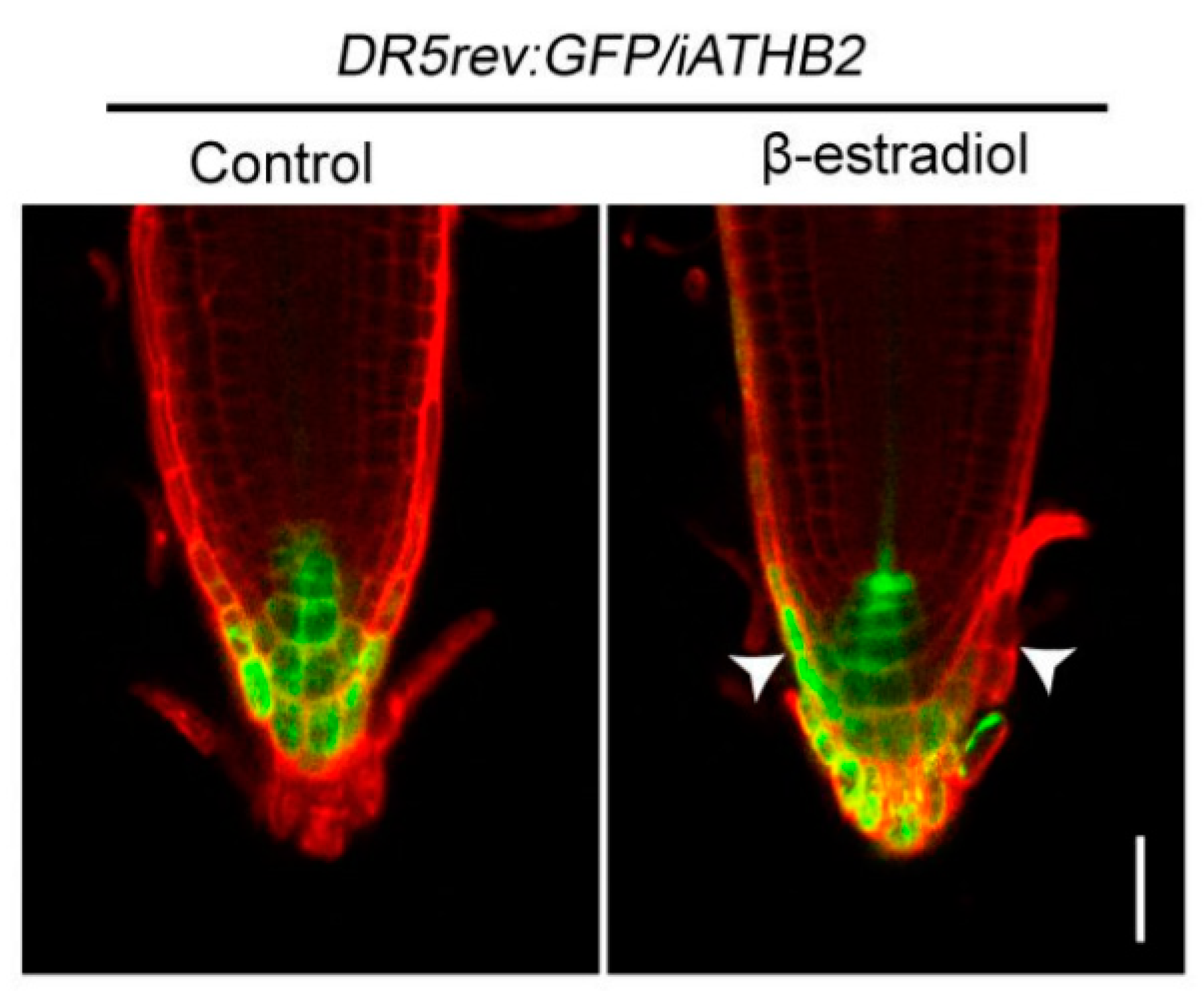
Considering that root gravitropic bending is triggered by the asymmetric auxin distribution in the root tip [32,33], we examined the spatial expression pattern of the auxin responsive reporter DR5rev:green fluorescent protein (GFP) [34] in the root tips of iATHB2 seedings. We here generated the double transgenic plant DR5rev:GFP/iATHB2 through genetic crossing between DR5rev:GFP and iATHB2 plants. After growing on 1/2 MS medium supplemented with or without 10 μM β-estradiol for six days, the fluorescence signals were observed in the root tips of DR5rev:GFP/iATHB2 plants. In the presence of 10 μM β-estradiol, the fluorescence signals of DR5rev:GFP/iATHB2 were asymmetric distribution in root tips as shown in the representative image (15/18) (Figure 4). However, the asymmetric DR5rev:GFP fluorescence signal pattern was not observed in the root tips of DR5rev:GFP/iATHB2 plants in the absence of 10 μM β-estradiol (Figure 4). These findings indicate that the asymmetric auxin response in the root tips of iATHB2 seedlings with β-estradiol treatment is correlated with their agravitropic root phenotypes.
2.5. Expression of Some Auxin Biosynthetic and Transport Genes Was Reduced in the iATHB2 Seedlings
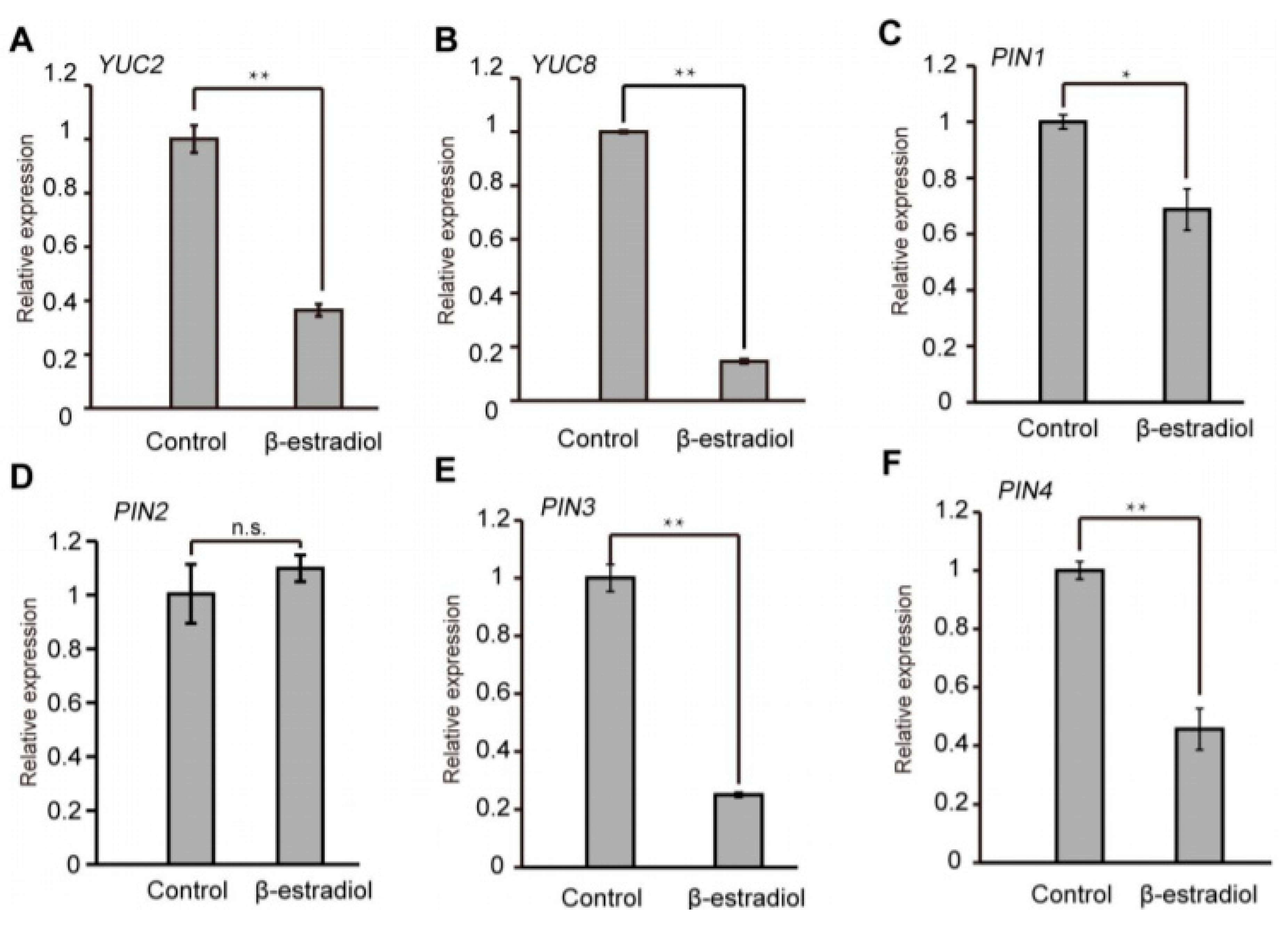
Auxin levels or distribution are controlled through synthesis and transport [35]. The auxin efflux carrier PIN proteins direct auxin flow in plants [36]. To investigate the underlying mechanism of asymmetric auxin response in the root tips of iATHB2 transgenic lines, we examined the expression of auxin biosynthetic and transport genes in the iATHB2 transgenic lines. The iATHB2 transgenic seedlings were grown on 1/2 MS medium for six days and then treated with 10 μM β-estradiol. As shown in Figure 5A–F, the transcript levels of YUCCA2 (YUC2), YUC8, PIN1, PIN3 and PIN4 were obviously decreased after β-estradiol treatment for 2 h. Previous studies have shown that the auxin-efflux facilitator PIN2 is involved in root gravitropism [12,36,37]. The transcriptional expression of PIN2 has no significant change in the iATHB2 transgenic seedlings after β-estradiol treatment. Taken together, these results indicated that the expression of some auxin biosynthetic and transport genes was reduced in the iATHB2 seedlings after β-estradiol treatment.
2.6. Molecular Characterization of HD-ZIP II Proteins in Common Wheat
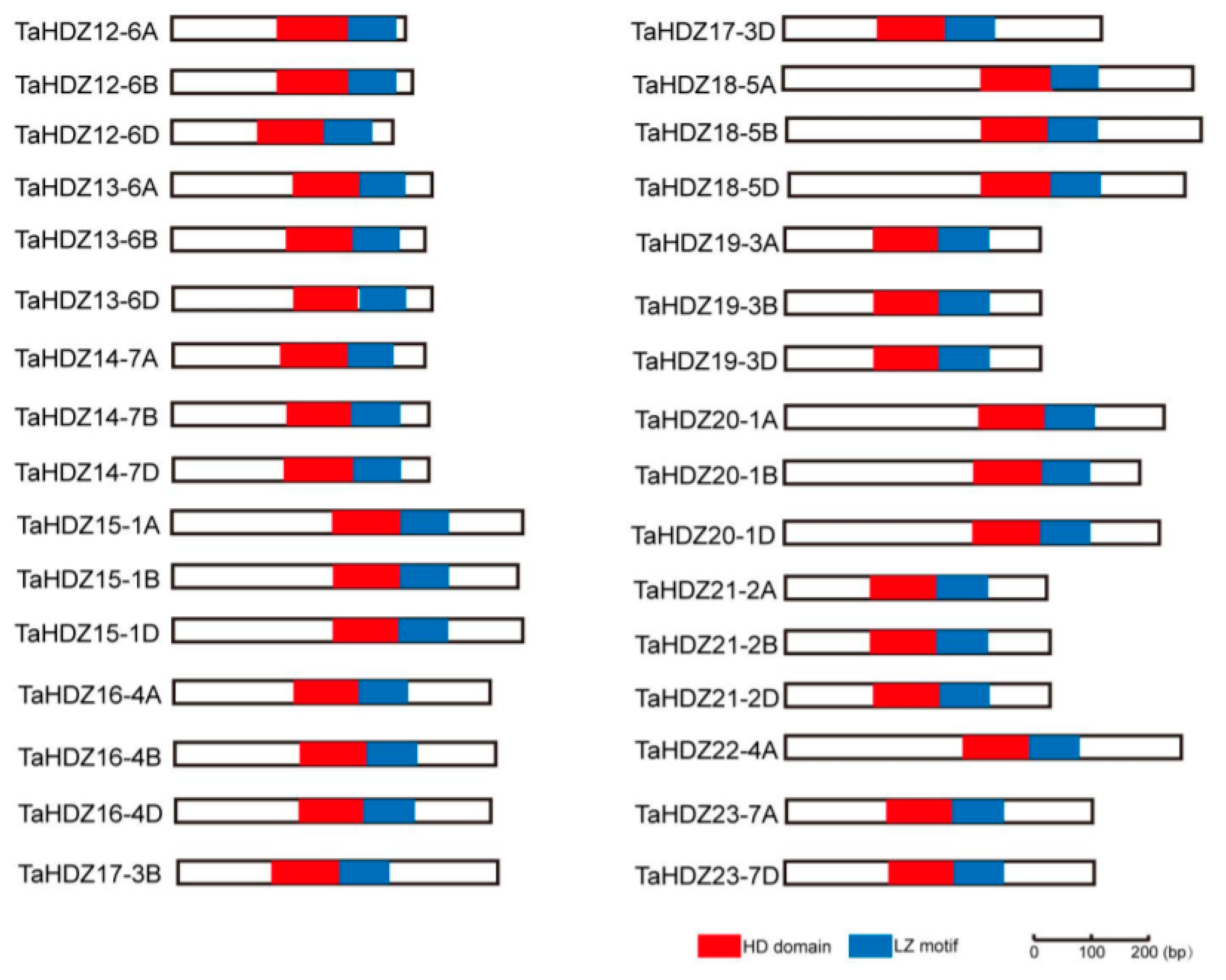
In this study, we have shown that ATHB2, one member of the HD-ZIP II subfamily transcription factor in Arabidopsis, is involved in the regulation of plant architecture potentially through the auxin pathway. To further understand the roles of HD-ZIP II proteins in monocot crops, we analyzed the protein structure and motif composition of HD-ZIP II subfamily proteins in common wheat. In a recent study, a total of 32 HD-ZIP II subfamily genes were identified in common wheat [38]. Protein structure analysis was conducted via the SMART software using the 32 HD-ZIP II proteins in common wheat, indicating that all the 32 HD-ZIP II proteins contain a homeobox domain (HD) and an adjacent leucine zipper (LZ) motif (Figure 6). The evolutionary relationships among the HD-ZIP II subfamily proteins of both Arabidopsis and wheat were analyzed via the MEGA7.0 software using the neighbor-joining method (Figures S1 and S3).
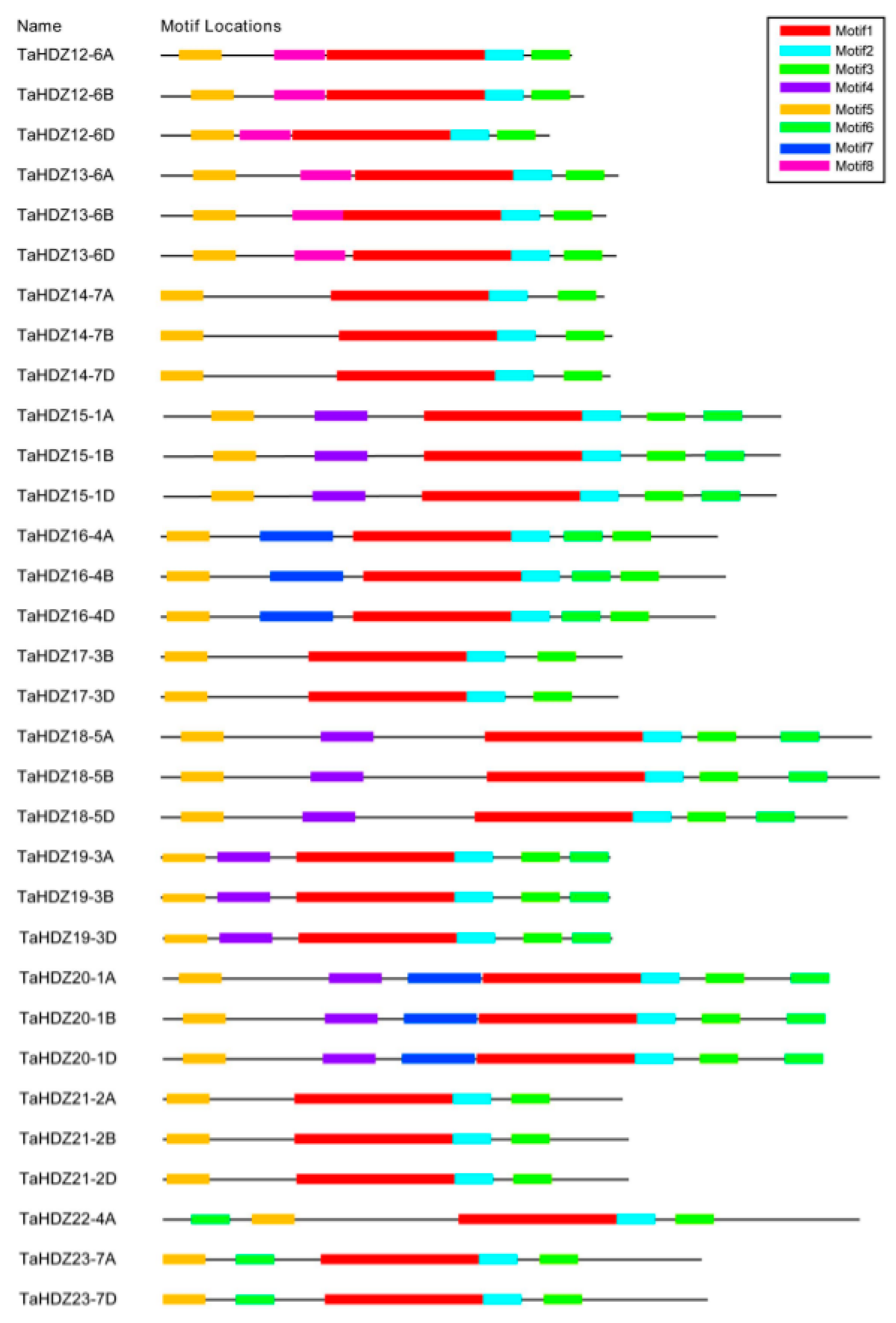
To comprehensively identify the potential conserved domains of wheat HD-ZIP II transcription factors, the full-length amino acid sequences were analyzed by the MEME online server. The results showed that eight predicted conserved motifs were identified, named as Motif 1 to 8 (Figure 7). As shown in Figure 7, each member of wheat HD-ZIP II subfamily proteins contains four common predicted motifs, including Motifs 1, 2, 3 and 5. Motif 1, a putative HD domain, was comprised of 78 amino acids (EDDGDGGGGARKKLRLSKEQSALLEESFKEHSTLSPKQKAALARQLGLRPRQVEVWFQNRRARTKLKQTEVDCEYLKR) (Figure S4). Motif 2, a putative LZ domain, was comprised of 19 amino acids (CCETLTEENRRLQRELAEL) (Figure S4). Motif 3 and Motif 5, whose functions remain unknown, were comprised of 19 amino acids (YYMPLPATTLTMCPSCERV) and 21 amino acids (EAEEDLGLALGLSLGAGSRPS), respectively (Figure S4). In addition to these conserved motifs contained in all the HD-ZIP II subfamily members, there are four predicted motifs which are specific to some members of the HD-ZIP II proteins.
2.7. Auxin-Responsive Elements in the Promoters of Wheat HD-ZIP II Genes
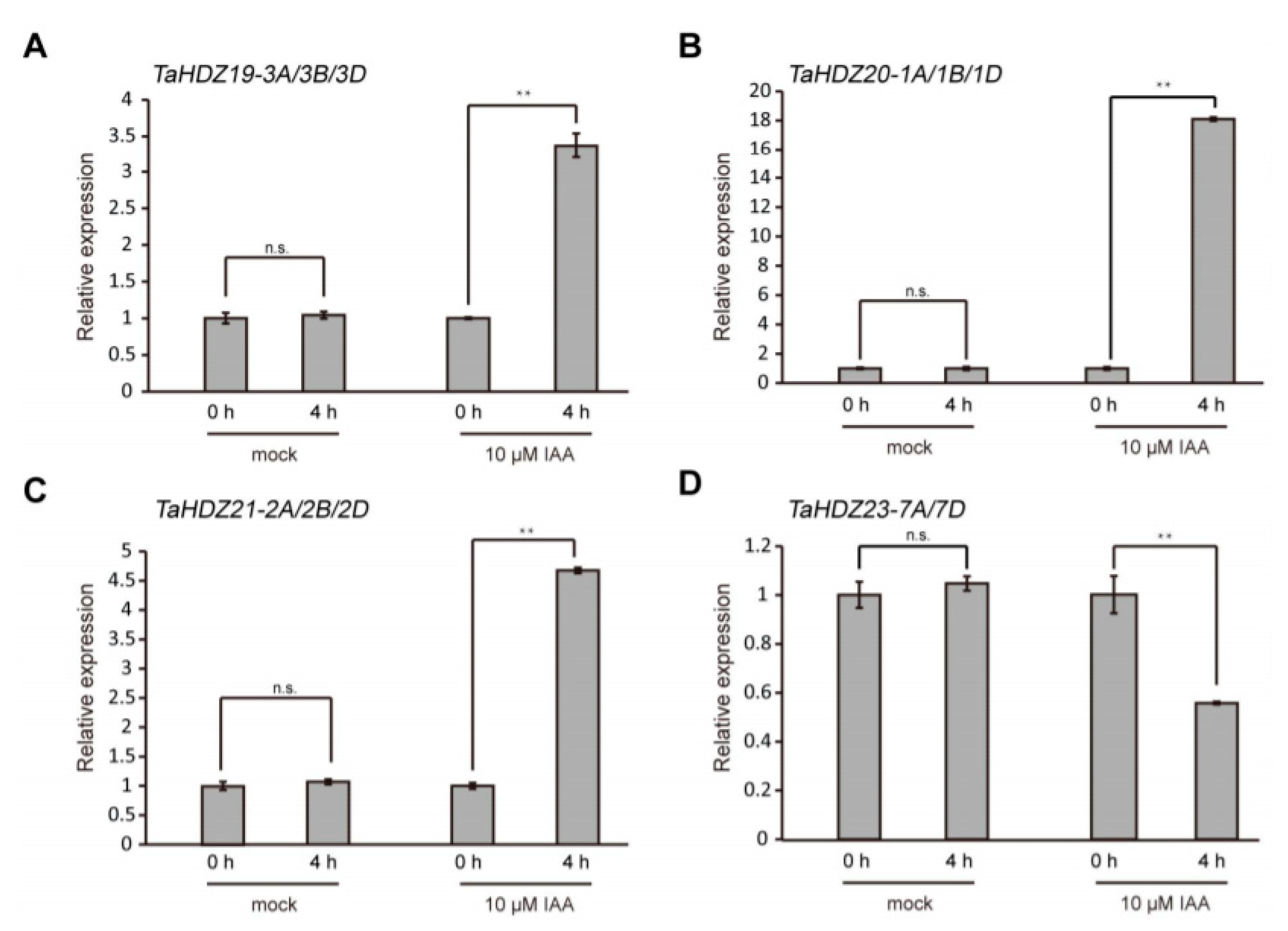
Auxin-responsive promoter elements (AuxREs) presented in the upstream region of genes play an important role in the auxin pathway. The canonical AuxRE “TGTCTC” shows a strong association with the auxin responsive expression pattern [39,40]. To understand the association of wheat HD-ZIP II subfamily proteins with the auxin pathway, we scanned the “TGTCTC” element in the 3 Kb genomic regions upstream of their coding regions. Interestingly, we found that among the 32 gene promoters, only five gene promoters did not contain the canonical AuxRE “TGTCTC” (Figure 8). Notably, the TaHDZ13-6A/6B/6D and TaHDZ16-4A/6B/6D homologous genes shared almost similar “TGTCTC” distribution patterns in their promoters (Figure 8). These findings indicated that the HD-ZIP II subfamily genes might be regulated by auxin in wheat. To further confirm this idea, the 4-day-old wheat seedlings were treated with 10 μM IAA for 4 h for qRT-PCR analysis. The results showed that the transcript levels of TaHDZ19-3A/3B/3D, TaHDZ20-1A/1B/1D and TaHDZ21-2A/2B/2D were up-regulated in wheat roots with auxin treatment, whereas the transcript levels of TaHDZ23-7A/7D was downregulated (Figure 9). Taken together, some wheat HD-ZIP II subfamily genes are regulated by auxin.
3. Discussion
Auxin regulates auxin-responsive gene expression which relies on an elegantly short signal transduction pathway (TIR1/AFB pathway), which has been extensively reviewed [5,41]. The Aux/IAA, which are well-known as the early auxin responsive genes, act as transcriptional repressors in this signaling [31]. Aux/IAA proteins recruit corepressors of the TOPLESS (TPL) family through a conserved EAR domain to silence ARF target genes [42,43]. The Aux/IAAs do not themselves bind DNA, but they can dimerize with the ARF family transcription factors [5]. The ARF family transcription factors regulate auxin-responsive gene expression.
Previous studies have shown that several HD-ZIP II proteins are well known for their role in shade avoidance [44], carpel margin development [45] and leaf polarity [46]. Moreover, members of the HD-ZIP II subfamily also control embryonic apical patterning and SAM function [30]. In this study, we found that ATHB2, encoding a transcription factor of the HD-ZIP II subfamily, is an early auxin-responsive gene (Figure 1). Phenotypic analyses showed that overexpression of ATHB2 impaired plant architecture, including reduced plant height and small leaves, which decreased auxin response in leaves when grown in soil (Figure 2). Meanwhile, the seedlings with chemical induction of ATHB2 exhibited abnormal root gravitropism, a typical auxin-related phenotype (Figure 3). We further showed that asymmetric auxin response occurred in the root tips of inducible ATHB2 plants (Figure 4). Therefore, both Aux/IAA and ATHB2 are early auxin-responsive genes and act as repressors of the auxin pathway, but possibly through distinct mechanisms.
The asymmetrical localization of PIN transporters (PIN1–PIN4 and PIN7) on PM contributes to the directionality of the auxin flow [36,47,48]. The differential expression and polar localization of PIN proteins constitutes the backbone of a transport network for directional auxin distribution in different tissues of the plant [49]. Directional auxin distribution leads to the formation of cellular auxin maxima and minima, which provides an essential cue for plant growth and differentiation at the level of individual cell and tissue [48,50]. PIN1, localizing to the basal (rootward) plasma membrane in root stele cells, directly transports auxin toward the root tip [12]. PIN2, PIN3 and PIN4, which also act in the root tip, mediate the auxin maximum and auxin redistribution for root gravitropism [11,37,51]. To investigate the underlying mechanism of the ATHB2-regulated auxin-related phenotype, we examined the expression of some PIN genes in the iATHB2 transgenic lines. qRT-PCR results showed that the transcript levels of PIN1, PIN3 and PIN4 were obviously downregulated by inducible overexpression of ATHB2 (Figure 5). Meanwhile, the auxin biosynthetic genes YUC2 and YUC8 were also downregulated by ATHB2 overexpression (Figure 5). Taken together, we demonstrate that ATHB2 regulates plant development possibly through modulating the auxin pathway in Arabidopsis.
In common wheat, a total of 113 HD-ZIP members were identified in recent studies [38,52]. However, the relationship between wheat HD-ZIP II transcription factors and the auxin pathway remains unclear. In this study, we analyzed the canonical AuxRE distribution in the promoters of wheat HD-ZIP II subfamily genes (Figure 8). A number of AuxREs were recognized in the promoters of wheat HD-ZIP II subfamily genes. Indeed, some wheat HD-ZIP II subfamily genes are regulated by auxin. Such knowledge may be useful to understand the regulation of wheat HD-ZIP II transcription factor in the auxin-mediated plant developmental processes.
4. Methods and Materials
4.1. Plant Materials and Growth Conditions
All the plants described in this study were in the Col-0 background. The full-length ATHB2 coding sequence was cloned into pMDC7 vector (iATHB2) [53,54] and p35S-FLAG vector (p35S:ATHB2-FLAG) [55], respectively. Col-0 was transformed with iATHB2 and p35S:ATHB2-FLAG by the floral dip transformation method, respectively [56]. Two independent transgenic lines of iATHB2 and p35S:ATHB2-FLAG were used for the experiments in this study. The DR5rev:GFP/iATHB2 and DR5:GUS/35S:ATHB2-FLAG plants were prepared by genetic crossing. Arabidopsis thaliana and wheat (Triticum aestivum) were grown under LD (16 h light/8 h dark) condition at 22 °C.
4.2. DNA Constructs
DNA constructs used in this study were generated based on construction methods. The construction methods were carried out with the classic molecular biology protocols and Gateway technology (Invitrogen). For ligase-independent ligation assays, the Ligation-Free Cloning MasterMix (abm) was used according to the application handbook. For Gateway cloning, pQBV3 vector (Gateway) was used as the entry vector and subsequently specific destination vectors were introduced into the Gateway system (Invitrogen). The primers used for generation in this study are shown in Table S1.
4.3. Root Phenotype Analyses
Seeds were sterilized by 75% (v/v) ethanol for seven minutes and then 100% (v/v) ethanol for three minutes (v/v). Seeds were stratified at 4 °C for three days. The seeds were then grown on 1/2 MS medium with or without 10 μM β-estradiol. For the phenotypic observation, the seedlings were grown vertically on 1/2 MS containing 10 μM β-estradiol for six days. The primary root lengths were measured by using ImageJ software (http://rsb.info.nih.gov/ij).
4.4. RNA Extraction and Gene Expression Analysis
The 6-day-old seedlings were collected as described. Total RNA was extracted using Trizol (Invitrogen) reagent. About 2 μg total RNA was applied to synthesize cDNA using the 5× All-In-One RT MasterMix system (Applied Biological Materials). The cDNA was diluted to 100 μL with water in a 1:5 ratio, and 2 μL of the diluted cDNA was used as a template. SYBR® Premix ExTaq Kit (TaKaRa) was used for qPCR reactions. qRT-PCR was performed using LightCycler 96 (Roche). Expression levels of target genes were normalized by ACTIN7. All the experiments were repeated independently three times. All the primers used for qRT-PCR are shown in Table S1.
4.5. Confocal Microscopy
Fluorescent samples were inspected by confocal microscopy (Carl Zeiss, LSM880, Germany). For imaging GFP and propidium iodide (PI) observation the 488 nm laser was used for excitation. Emission between 500 and 550 nm band-pass was detected for GFP, between 560 and 610 nm band-pass for PI.
4.6. Protein Extraction and Immunoblotting
The extracted buffer (125 mM Tris-HCl at pH 6.8, 4% SDS, 20% glycerol, 0.001% bromophenol blue) with freshly added 2% β-mercaptoethanol was used for total protein extraction. Immunoblots were performed as described [57]. To detect a FLAG-tagged protein, we used anti-FLAG (1:5000; MBL, Japan) and anti-mouse IgG (1:75000) antibodies. Actins (1:5000; CWBIO) were used as the loading controls. Three independent biological replicates were performed with similar results.
4.7. Analysis of GUS Activity
For GUS activity analysis, 4-week-old Arabidopsis leaves were transferred into staining solution (1 mM 5-bromo-4-chloro-3-indolyl-beta-glucuronic acid solution in 100 mM sodium phosphate, pH 7.0, 0.1 mM EDTA, 0.5 mM ferricyanide, 0.5 mM ferrocyanide and 0.1% Triton X-100) [58]. Leaves were then applied to vacuum for 20 min and incubated at 37 °C overnight. To clear chlorophyll from plant tissues, 100% ethanol was used. Individual representative seedlings were photographed.
4.8. Phylogenetic Analysis
A neighbor-joining phylogenetic tree was constructed based on 1000 bootstrap replicates by comparing full-length protein sequences aligned with the Clustal W algorithm within MEGA7.0.
4.9. Protein Structure and Motif Composition Analyses
Predicted protein domains were identified by the SMART tool (http://smart.emblheidelberg.de/). The MEME online program (Bailey et al., 2009) was used to identify conserved motifs.
4.10. Accession Numbers
Sequence data of Arabidopsis from this article can be found in the Arabidopsis Genome initiative data library under the following accession numbers: ATHB2 (AT4G16780), IAA19 (AT3G15540), YUC2 (AT4G13260), YUC8 (AT4G28720), PIN1 (AT1G73590), PIN2 (AT5G57090), PIN3 (AT1G70940), PIN4 (AT2G01420), ATHB4 (AT2G44910), ATHB17 (AT2G01430), ATHB18 (AT1G70920), HAT1 (AT4G17460), HAT2 (AT5G47370), HAT9 (AT2G22800), HAT14 (AT5G06710) and HAT22 (AT4G37790).
Supplementary Materials
Supplementary materials can be found at https://www.mdpi.com/1422-0067/21/9/3250/s1.
Author Contributions
J.S. designed the research project; G.H. and P.L. performed the experiments; G.H., J.S. and H.Z. analyzed the data and wrote the article. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Institute of Crop Sciences, Chinese Academy of Agricultural Sciences (CAAS), the Agricultural Science and Technology Innovation Program of CAAS, and Youth Talent Plan of CAAS.
Acknowledgments
The authors thank Klaus Palme for providing some research materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kepinski, S.; Leyser, O. The Arabidopsis F-box protein TIR1 is an auxin receptor. Nature 2005, 435, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Korasick, D.A.; Jez, J.M.; Strader, L.C. Refining the nuclear auxin response pathway through structural biology. Curr. Opin. Plant Biol. 2015, 27, 22–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.H.; Estelle, M. Diversity and specificity: auxin perception and signaling through the TIR1/AFB pathway. Curr. Opin. Plant Biol. 2014, 21, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, S.K.; Strader, L.C. Up in the air: Untethered Factors of Auxin Response. F1000Res 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, E.J.; Estelle, M. Mechanism of Auxin-Regulated Gene Expression in Plants. Ann. Rev. Genet. 2009, 43, 265–285. [Google Scholar] [CrossRef] [Green Version]
- Dharmasiri, N.; Dharmasiri, S.; Estelle, M. The F-box protein TIR1 is an auxin receptor. Nature 2005, 435, 441–445. [Google Scholar] [CrossRef]
- Cao, M.; Chen, R.; Li, P.; Yu, Y.; Zheng, R.; Ge, D.; Zheng, W.; Wang, X.; Gu, Y.; Gelova, Z.; et al. TMK1-mediated auxin signalling regulates differential growth of the apical hook. Nature 2019, 568, 240–243. [Google Scholar] [CrossRef]
- Adamowski, M.; Friml, J. PIN-dependent auxin transport: action, regulation, and evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Balzan, S.; Johal, G.S.; Carraro, N. The role of auxin transporters in monocots development. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Bainbridge, K.; Guyomarc’h, S.; Bayer, E.; Swarup, R.; Bennett, M.; Mandel, T.; Kuhlemeier, C. Auxin influx carriers stabilize phyllotactic patterning. Genes Dev. 2008, 22, 810–823. [Google Scholar] [CrossRef] [Green Version]
- Friml, J.; Benkova, E.; Blilou, I.; Wisniewska, J.; Hamann, T.; Ljung, K.; Woody, S.; Sandberg, G.; Scheres, B.; Jürgens, G.; et al. AtPIN4 mediates sink-driven auxin gradients and root patterning in Arabidopsis. Cell 2002, 108, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Galweiler, L.; Guan, C.H.; Muller, A.; Wisman, E.; Mendgen, K.; Yephremov, A.; Palme, K. Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science 1998, 282, 2226–2230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisler, M.; Blakeslee, J.J.; Bouchard, R.; Lee, O.R.; Vincenzetti, V.; Bandyopadhyay, A.; Titapiwatanakun, B.; Peer, W.A.; Bailly, A.; Richards, E.L.; et al. Cellular efflux of auxin catalyzed by the Arabidopsis MDR/PGP transporter AtPGP1. Plant J. 2005, 44, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Noh, B.; Bandyopadhyay, A.; Peer, W.A.; Spalding, E.P.; Murphy, A.S. Enhanced gravi- and phototropism in plant mdr mutants mislocalizing the auxin efflux protein PIN1. Nature 2003, 423, 999–1002. [Google Scholar] [CrossRef]
- Peret, B.; Swarup, K.; Ferguson, A.; Seth, M.; Yang, Y.D.; Dhondt, S.; James, N.; Casimiro, I.; Perry, P.; Syed, A.; et al. AUX/LAX Genes Encode a Family of Auxin Influx Transporters That Perform Distinct Functions during Arabidopsis Development. Plant Cell 2012, 24, 2874–2885. [Google Scholar] [CrossRef] [Green Version]
- Swarup, R.; Peret, B. AUX/LAX family of auxin influx carriers-an overview. Front Plant Sci. 2012, 3. [Google Scholar]
- Benkova, E.; Michniewicz, M.; Sauer, M.; Teichmann, T.; Seifertova, D.; Jurgens, G.; Friml, J. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Front Plant Sci. 2012, 3, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Swarup, R.; Bennett, M. Auxin transport: the fountain of life in plants? Dev. Cell 2003, 5, 824–826. [Google Scholar] [CrossRef] [Green Version]
- Perotti, M.F.; Ribone, P.A.; Chan, R.L. Plant Transcription Factors from the Homeodomain-Leucine Zipper Family I. Role in Development and Stress Responses. IUBMB Life 2017, 69, 280–289. [Google Scholar] [CrossRef]
- Peterson, K.M.; Shyu, C.; Burr, C.A.; Horst, R.J.; Kanaoka, M.M.; Omae, M.; Sato, Y.; Torii, K.U. Arabidopsis homeodomain-leucine zipper IV proteins promote stomatal development and ectopically induce stomata beyond the epidermis. Development 2013, 140, 1924–1935. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, K.; Nishiyama, T.; Kato, M.; Hasebe, M. Isolation of homeodomain-leucine zipper genes from the moss Physcomitrella patens and the evolution of homeodomain-leucine zipper genes in land plants. Mol. Biol. Evol. 2001, 18, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, O.; Hur, Y.S.; Kim, Y.K.; Lee, H.J.; Kim, S.; Kim, M.R.; Nam, K.H.; Lee, M.S.; Kim, B.Y.; Park, J.; et al. ATHB12, an ABA-Inducible Homeodomain-Leucine Zipper (HD-Zip) Protein of Arabidopsis, Negatively Regulates the Growth of the Inflorescence Stem by Decreasing the Expression of a Gibberellin 20-Oxidase Gene. Plant Cell Physiol. 2010, 51, 1537–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-Zip family. Trends Plant Sci. 2007, 12, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Ciarbelli, A.R.; Ciolfi, A.; Salvucci, S.; Ruzza, V.; Possenti, M.; Carabelli, M.; Fruscalzo, A.; Sessa, G.; Morelli, G.; Ruberti, I. The Arabidopsis homeodomain-leucine zipper II gene family: diversity and redundancy. Plant Mol. Biol. 2008, 68, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Stamm, P.; Kumar, P.P. The phytohormone signal network regulating elongation growth during shade avoidance. J. Exp. Bot. 2010, 61, 2889–2903. [Google Scholar] [CrossRef]
- Agalou, A.; Purwantomo, S.; Overnas, E.; Johannesson, H.; Zhu, X.; Estiati, A.; de Kam, R.J.; Engstrom, P.; Slamet-Loedin, I.H.; Zhu, Z.; et al. A genome-wide survey of HD-Zip genes in rice and analysis of drought-responsive family members. Plant Mol. Biol. 2008, 66, 87–103. [Google Scholar] [CrossRef]
- Harris, J.C.; Hrmova, M.; Lopato, S.; Langridge, P. Modulation of plant growth by HD-Zip class I and II transcription factors in response to environmental stimuli. New Phytol. 2011, 190, 823–837. [Google Scholar] [CrossRef]
- Manavella, P.A.; Dezar, C.A.; Bonaventure, G.; Baldwin, I.T.; Chan, R.L. HAHB4, a sunflower HD-Zip protein, integrates signals from the jasmonic acid and ethylene pathways during wounding and biotic stress responses. Plant J. 2008, 56, 376–388. [Google Scholar] [CrossRef]
- Roodbarkelari, F.; Groot, E.P. Regulatory function of homeodomain-leucine zipper (HD-ZIP) family proteins during embryogenesis. New Phytol. 2017, 213, 95–104. [Google Scholar] [CrossRef]
- Turchi, L.; Carabelli, M.; Ruzza, V.; Possenti, M.; Sassi, M.; Penalosa, A.; Sessa, G.; Salvi, S.; Forte, V.; Morelli, G.; et al. Arabidopsis HD-Zip II transcription factors control apical embryo development and meristem function. Development 2013, 140, 2118–2129. [Google Scholar] [CrossRef] [Green Version]
- Ulmasov, T.; Murfett, J.; Hagen, G.; Guilfoyle, T.J. Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell 1997, 9, 1963–1971. [Google Scholar] [PubMed] [Green Version]
- Morita, M.T. Directional gravity sensing in gravitropism. Annu. Rev. Plant Biol. 2010, 61, 705–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strohm, A.K.; Baldwin, K.L.; Masson, P.H. Multiple roles for membrane-associated protein trafficking and signaling in gravitropism. Front. Plant Sci. 2012, 3, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottenschlager, I.; Wolff, P.; Wolverton, C.; Bhalerao, R.P.; Sandberg, G.; Ishikawa, H.; Evans, M.; Palme, K. Gravity-regulated differential auxin transport from columella to lateral root cap cells. Proc. Natl. Acad. Sci. USA 2003, 100, 2987–2991. [Google Scholar] [CrossRef] [Green Version]
- Korasick, D.A.; Enders, T.A.; Strader, L.C. Auxin biosynthesis and storage forms. J. Exp. Bot. 2013, 64, 2541–2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisniewska, J.; Xu, J.; Seifertova, D.; Brewer, P.B.; Ruzicka, K.; Blilou, I.; Rouquie, D.; Benkova, E.; Scheres, B.; Friml, J. Polar PIN localization directs auxin flow in plants. Science 2006, 312, 883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, A.; Guan, C.; Galweiler, L.; Tanzler, P.; Huijser, P.; Marchant, A.; Parry, G.; Bennett, M.; Wisman, E.; Palme, K. AtPIN2 defines a locus of Arabidopsis for root gravitropism control. EMBO J. 1998, 17, 6903–6911. [Google Scholar] [CrossRef]
- Li, S.; Chen, N.; Li, F.; Mei, F.; Wang, Z.; Cheng, X.; Kang, Z.; Mao, H. Characterization of wheat homeodomain-leucine zipper family genes and functional analysis of TaHDZ5-6A in drought tolerance in transgenic Arabidopsis. BMC Plant Biol 2020, 20, 50. [Google Scholar] [CrossRef] [Green Version]
- Berendzen, K.W.; Weiste, C.; Wanke, D.; Kilian, J.; Harter, K.; Dröge-Laser, W. Bioinformatic cis-element analyses performed in Arabidopsis and rice disclose bZIP- and MYB-related binding sites as potential AuxRE-coupling elements in auxin-mediated transcription. BMC Plant Biol 2012, 12, 125. [Google Scholar] [CrossRef] [Green Version]
- Mironova, V.V.; Omelyanchuk, N.A.; Wiebe, D.S.; Levitsky, V.G. Computational analysis of auxin responsive elements in the Arabidopsis thaliana L. genome. BMC Genom. 2014, 15 (Suppl. 12), S4. [Google Scholar] [CrossRef] [Green Version]
- Salehin, M.; Bagchi, R.; Estelle, M. SCFTIR1/AFB-Based Auxin Perception: Mechanism and Role in Plant Growth and Development. Plant Cell 2015, 27, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szemenyei, H.; Hannon, M.; Long, J.A. TOPLESS mediates auxin-dependent transcriptional repression during Arabidopsis embryogenesis. Science 2008, 319, 1384–1386. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.B.; Hagen, G.; Guilfoyle, T.J. Aux/IAA proteins contain a potent transcriptional repression domain. Plant Cell 2004, 16, 533–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruberti, I.; Sessa, G.; Ciolfi, A.; Possenti, M.; Carabelli, M.; Morelli, G. Plant adaptation to dynamically changing environment: the shade avoidance response. Biotechnol. Adv. 2012, 30, 1047–1058. [Google Scholar] [CrossRef]
- Reymond, M.C.; Brunoud, G.; Chauvet, A.; Martinez-Garcia, J.F.; Martin-Magniette, M.L.; Moneger, F.; Scutt, C.P. A light-regulated genetic module was recruited to carpel development in Arabidopsis following a structural change to SPATULA. Plant Cell 2012, 24, 2812–2825. [Google Scholar] [CrossRef] [Green Version]
- Bou-Torrent, J.; Salla-Martret, M.; Brandt, R.; Musielak, T.; Palauqui, J.C.; Martinez-Garcia, J.F.; Wenkel, S. ATHB4 and HAT3, two class II HD-ZIP transcription factors, control leaf development in Arabidopsis. Plant Signal. Behav. 2012, 7, 1382–1387. [Google Scholar] [CrossRef] [Green Version]
- Krecek, P.; Skupa, P.; Libus, J.; Naramoto, S.; Tejos, R.; Friml, J.; Zazimalova, E. The PIN-FORMED (PIN) protein family of auxin transporters. Genome Biol. 2009, 10, 249. [Google Scholar] [CrossRef] [Green Version]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in action: signalling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell Biol. 2006, 7, 847–859. [Google Scholar] [CrossRef]
- Petrasek, J.; Friml, J. Auxin transport routes in plant development. Development 2009, 136, 2675–2688. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, I.C.; Schwechheimer, C. Dynamic control of auxin transport-dependent growth by AGCVIII protein kinases. Curr. Opin. Plant Biol. 2014, 22, 108–115. [Google Scholar]
- Friml, J.; Wisniewska, J.; Benkova, E.; Mendgen, K.; Palme, K. Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 2002, 415, 806–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, H.; Shu, D.; Wang, M.; Xing, G.; Zhan, H.; Du, X.; Song, W.; Nie, X. Genomewide identification and expression analysis of the HD-Zip gene family in wheat (Triticum aestivum L.). Genes 2018, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Qi, L.; Li, Y.; Chu, J.; Li, C. PIF4-mediated activation of YUCCA8 expression integrates temperature into the auxin pathway in regulating arabidopsis hypocotyl growth. PLoS Genet. 2012, 8, e1002594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, J.; Niu, Q.W.; Frugis, G.; Chua, N.H. The WUSCHEL gene promotes vegetative-to-embryonic transition in Arabidopsis. Plant J. 2002, 30, 349–359. [Google Scholar]
- He, G.; Liu, J.; Dong, H.; Sun, J. The Blue-Light Receptor CRY1 Interacts with BZR1 and BIN2 to Modulate the Phosphorylation and Nuclear Function of BZR1 in Repressing BR Signaling in Arabidopsis. Mol Plant. 2019, 12, 689–703. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedmale, U.V.; Liscum, E. Regulation of phototropic signaling in Arabidopsis via phosphorylation state changes in the phototropin 1-interacting protein NPH3. J. Biol. Chem. 2007, 282, 19992–20001. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1997, 6, 3901–3907. [Google Scholar] [CrossRef]
Figure 1.
Expression patterns of the ATHB2 subset genes. (A–F) Expression analyses of ATHB2, HAT1, HAT2, HAT3, ATHB4 and 1AA19 in response to auxin treatment. The 6 day old wild-type (Col-0) seedlings were treated with 10 μM IAA for different time points. The ACTIN7 gene was used as an internal reference. The qRT-PCR results were performed for three biological replications and similar results were observed. Representative qRT-PCR results with three technical replicates were shown. Error bars denote ± SD.
Figure 1.
Expression patterns of the ATHB2 subset genes. (A–F) Expression analyses of ATHB2, HAT1, HAT2, HAT3, ATHB4 and 1AA19 in response to auxin treatment. The 6 day old wild-type (Col-0) seedlings were treated with 10 μM IAA for different time points. The ACTIN7 gene was used as an internal reference. The qRT-PCR results were performed for three biological replications and similar results were observed. Representative qRT-PCR results with three technical replicates were shown. Error bars denote ± SD.

Figure 2.
Morphological phenotype of 35S:ATHB2-FLAG transgenic lines. (A) Immunoblotting analysis showing the ATHB2-FLAG protein levels in the 35S:ATHB2-FLAG transgenic plants. The 6 day old seedlings of Col-0 and 35S:ATHB2-FLAG transgenic plants were harvested for immunoblotting analysis. ACTIN was used as a loading control. 1# and 2# represent two independent transgenic lines of 35S:ATHB2-FLAG. The data are representative of three independent experiments. (B and C) Overview of the Col-0 and 35S:ATHB2-FLAG transgenic lines at the adult stage. (B) Scale bars, 4cm. Leaf morphology of the Col-0 and 35S:ATHB2-FLAG transgenic lines at the adult stage. (C) Scale bars, 2 mm. The 4-week-old 35S:ATHB2-FLAG transgenic plants grown under normal growth conditions were used for phenotypic analyses. (B and C) Two independent transgenic lines were used for phenotype observation. The images are representative of three independent experiments. (D) Expression patterns of DR5:GUS in the leaves of DR5:GUS and DR5:GUS/35S:ATHB2-FLAG plants. The leaves of 3-week-old DR5:GUS and DR5:GUS/35S:ATHB2-FLAG plants were used for GUS activity analyses. The images are representative of three independent experiments.
Figure 2.
Morphological phenotype of 35S:ATHB2-FLAG transgenic lines. (A) Immunoblotting analysis showing the ATHB2-FLAG protein levels in the 35S:ATHB2-FLAG transgenic plants. The 6 day old seedlings of Col-0 and 35S:ATHB2-FLAG transgenic plants were harvested for immunoblotting analysis. ACTIN was used as a loading control. 1# and 2# represent two independent transgenic lines of 35S:ATHB2-FLAG. The data are representative of three independent experiments. (B and C) Overview of the Col-0 and 35S:ATHB2-FLAG transgenic lines at the adult stage. (B) Scale bars, 4cm. Leaf morphology of the Col-0 and 35S:ATHB2-FLAG transgenic lines at the adult stage. (C) Scale bars, 2 mm. The 4-week-old 35S:ATHB2-FLAG transgenic plants grown under normal growth conditions were used for phenotypic analyses. (B and C) Two independent transgenic lines were used for phenotype observation. The images are representative of three independent experiments. (D) Expression patterns of DR5:GUS in the leaves of DR5:GUS and DR5:GUS/35S:ATHB2-FLAG plants. The leaves of 3-week-old DR5:GUS and DR5:GUS/35S:ATHB2-FLAG plants were used for GUS activity analyses. The images are representative of three independent experiments.

Figure 3.
Root gravitropic phenotype of β-estradiol-inducible ATHB2-overexpression transgenic lines. (A) qRT-PCR analysis showing the β-estradiol-induced expression pattern of ATHB2 in the iATHB2 lines. The ACTIN gene was used as an internal reference. 1# and 2# represent two independent transgenic lines of iATHB2. The qRT-PCR results were performed for three biological replications and similar results were observed. Representative qRT-PCR results with three technical replicates were shown. Error bars denote ± SD. (B) Root gravitropic phenotypes of the iATHB2 transgenic lines. Seedlings of the Col-0 and inducible ATHB2 overexpression plants (iATHB2) were grown on 1/2 MS medium with or without 10 μM β-estradiol for six days. Two independent transgenic lines were used for phenotypic observation. The images are representative of three independent experiments.
Figure 3.
Root gravitropic phenotype of β-estradiol-inducible ATHB2-overexpression transgenic lines. (A) qRT-PCR analysis showing the β-estradiol-induced expression pattern of ATHB2 in the iATHB2 lines. The ACTIN gene was used as an internal reference. 1# and 2# represent two independent transgenic lines of iATHB2. The qRT-PCR results were performed for three biological replications and similar results were observed. Representative qRT-PCR results with three technical replicates were shown. Error bars denote ± SD. (B) Root gravitropic phenotypes of the iATHB2 transgenic lines. Seedlings of the Col-0 and inducible ATHB2 overexpression plants (iATHB2) were grown on 1/2 MS medium with or without 10 μM β-estradiol for six days. Two independent transgenic lines were used for phenotypic observation. The images are representative of three independent experiments.

Figure 4.
Asymmetric auxin response was observed in the root tips of iATHB2 plants. The spatial expression pattern of the DR5rev:GFP auxin responsive reporter in the root tips of 6-day-old DR5rev:GFP/iATHB2 plants grown on the medium with or without 10 μM β-estradiol. The images are representative of three independent experiments. Roots were stained with propidium iodide (red). Arrowheads indicate the asymmetric auxin response in the root tips. Scale bars = 50 μm. Arrowheads indicate the asymmetric auxin response of DR5rev:GFP/iATHB2 with 10 μM β-estradiol treatment.
Figure 4.
Asymmetric auxin response was observed in the root tips of iATHB2 plants. The spatial expression pattern of the DR5rev:GFP auxin responsive reporter in the root tips of 6-day-old DR5rev:GFP/iATHB2 plants grown on the medium with or without 10 μM β-estradiol. The images are representative of three independent experiments. Roots were stained with propidium iodide (red). Arrowheads indicate the asymmetric auxin response in the root tips. Scale bars = 50 μm. Arrowheads indicate the asymmetric auxin response of DR5rev:GFP/iATHB2 with 10 μM β-estradiol treatment.

Figure 5.
Overexpression of ATHB2 reduced the expression levels of some auxin biosynthetic and transport genes. (A–F) Expression patterns of YUC2, YUC8 and several PINs genes in the iATHB2 transgenic lines. The different 6-day-old iATHB2 transgenic lines were treated with or without 10 μM β-estradiol for 2 h. The ACTIN7 gene was used as an internal reference. The shown data are results of the representative iATHB2 2# transgenic line. The qRT-PCR results were performed for three biological replications and similar results were observed. Representative qRT-PCR results with three technical replicates were shown. Error bars denote ± SD. * p < 0.05, ** p < 0.01, Student’s t test. No significant difference is shown by n.s.
Figure 5.
Overexpression of ATHB2 reduced the expression levels of some auxin biosynthetic and transport genes. (A–F) Expression patterns of YUC2, YUC8 and several PINs genes in the iATHB2 transgenic lines. The different 6-day-old iATHB2 transgenic lines were treated with or without 10 μM β-estradiol for 2 h. The ACTIN7 gene was used as an internal reference. The shown data are results of the representative iATHB2 2# transgenic line. The qRT-PCR results were performed for three biological replications and similar results were observed. Representative qRT-PCR results with three technical replicates were shown. Error bars denote ± SD. * p < 0.05, ** p < 0.01, Student’s t test. No significant difference is shown by n.s.

Figure 6.
Schematic diagrams of the conserved domain structure of wheat HD-ZIP II subfamily proteins. Red box indicates the homeobox domain (HD) and blue box indicates the adjacent leucine zipper (LZ) motif, respectively.
Figure 6.
Schematic diagrams of the conserved domain structure of wheat HD-ZIP II subfamily proteins. Red box indicates the homeobox domain (HD) and blue box indicates the adjacent leucine zipper (LZ) motif, respectively.

Figure 7.
Putative motifs of the wheat HD-ZIP II subfamily proteins using the MEME program. The different conserved motifs are marked by different colors.
Figure 7.
Putative motifs of the wheat HD-ZIP II subfamily proteins using the MEME program. The different conserved motifs are marked by different colors.

Figure 8.
The “TGTCTC” elements in the 3 Kb genomic regions upstream of coding regions are indicated by blue box.
Figure 8.
The “TGTCTC” elements in the 3 Kb genomic regions upstream of coding regions are indicated by blue box.

Figure 9.
Several wheat HD-ZIP II subfamily genes were regulated by auxin. (A–D) Auxin-induced expression pattern of TaHDZ19-3A/3B/3D, TaHDZ20-1A/1B/1D, TaHDZ21-2A/2B/2D and TaHDZ23-7A/7D by qRT-PCR. The 4-day-old wheat seedlings were treated with 10 μM IAA for 4 h. The TaGAPDH gene was used as an internal reference. The qRT-PCR results were performed for three biological replications and similar results were observed. Representative qRT-PCR results with three technical replicates are shown. Error bars denote ± SD. ** p < 0.01, Student’s t test. No significant difference is shown by n.s.
Figure 9.
Several wheat HD-ZIP II subfamily genes were regulated by auxin. (A–D) Auxin-induced expression pattern of TaHDZ19-3A/3B/3D, TaHDZ20-1A/1B/1D, TaHDZ21-2A/2B/2D and TaHDZ23-7A/7D by qRT-PCR. The 4-day-old wheat seedlings were treated with 10 μM IAA for 4 h. The TaGAPDH gene was used as an internal reference. The qRT-PCR results were performed for three biological replications and similar results were observed. Representative qRT-PCR results with three technical replicates are shown. Error bars denote ± SD. ** p < 0.01, Student’s t test. No significant difference is shown by n.s.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
He, G.; Liu, P.; Zhao, H.; Sun, J. The HD-ZIP II Transcription Factors Regulate Plant Architecture through the Auxin Pathway. Int. J. Mol. Sci. 2020, 21, 3250. https://doi.org/10.3390/ijms21093250
AMA Style
He G, Liu P, Zhao H, Sun J. The HD-ZIP II Transcription Factors Regulate Plant Architecture through the Auxin Pathway. International Journal of Molecular Sciences. 2020; 21(9):3250. https://doi.org/10.3390/ijms21093250
Chicago/Turabian StyleHe, Guanhua, Pan Liu, Huixian Zhao, and Jiaqiang Sun. 2020. "The HD-ZIP II Transcription Factors Regulate Plant Architecture through the Auxin Pathway" International Journal of Molecular Sciences 21, no. 9: 3250. https://doi.org/10.3390/ijms21093250
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.