The Impact of Spaceflight and Simulated Microgravity on Cell Adhesion
by
 , ,
, ,
Xiao Lin
1,2,3,4, 
Kewen Zhang
1,2,3,4,
Daixu Wei
5,
Ye Tian
1,2,3,4,
Yongguang Gao
1,2,3,4,
Zhihao Chen
1,2,3,4 and
Airong Qian
1,2,3,4,* 1
Lab for Bone Metabolism, Key Lab for Space Biosciences and Biotechnology, School of Life Sciences, Northwestern Polytechnical University, Xi’an, 710072, China
2
Xi’an Key Laboratory of Special Medicine and Health Engineering, School of Life Sciences, Northwestern Polytechnical University, Xi’an 710072, China
3
Research Center for Special Medicine and Health Systems Engineering, School of Life Sciences, Northwestern Polytechnical University, Xi’an 710072, China
4
NPU-UAB Joint Laboratory for Bone Metabolism, School of Life Sciences, Northwestern Polytechnical University, Xi’an 710072, China
5
Key Laboratory of Resource Biology and Biotechnology in Western China, Ministry of Education, School of Medicine, Northwest University, 229 Taibai North Road, Xi’an 710069, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(9), 3031; https://doi.org/10.3390/ijms21093031
Submission received: 17 March 2020
/
Revised: 20 April 2020
/
Accepted: 23 April 2020
/
Published: 25 April 2020
(This article belongs to the Special Issue Microgravity and Cell Adherence)
Abstract
:Microgravity induces a number of significant physiological changes in the cardiovascular, nervous, immune systems, as well as the bone tissue of astronauts. Changes in cell adhesion properties are one aspect affected during long-term spaceflights in mammalian cells. Cellular adhesion behaviors can be divided into cell–cell and cell–matrix adhesion. These behaviors trigger cell–cell recognition, conjugation, migration, cytoskeletal rearrangement, and signal transduction. Cellular adhesion molecule (CAM) is a general term for macromolecules that mediate the contact and binding between cells or between cells and the extracellular matrix (ECM). In this review, we summarize the four major classes of adhesion molecules that regulate cell adhesion, including integrins, immunoglobulin superfamily (Ig-SF), cadherins, and selectin. Moreover, we discuss the effects of spaceflight and simulated microgravity on the adhesion of endothelial cells, immune cells, tumor cells, stem cells, osteoblasts, muscle cells, and other types of cells. Further studies on the effects of microgravity on cell adhesion and the corresponding physiological behaviors may help increase the safety and improve the health of astronauts in space.
1. Introduction
Microgravity is a state in which the gravitational force acting in any single direction is negligible. True microgravity is present during parabolic flights, on orbiting spacecraft, or space labs on the international space station (ISS). However, the absence of gravity in space (zero gravity) can also be simulated as microgravity on earth, for which many ground-based tools were developed. These include the clinostat, magnetic levitation, and the rotating wall vessel bioreactor [1,2]. Recently, researchers are investigating the effects of microgravity on the physiological state of cells and the underlying mechanisms using earth-based simulation techniques.
The biological effects of weightlessness on the human body have been recognized and investigated since the earliest space flights, forming one of the most important research directions in the field of aerospace medicine. During both short- and long-term space flight, astronauts are exposed to several risk factors related to radiation and the absence of gravity. Weightlessness eliminates the normal hydrostatic pressure gradient within the body, leading to cardiovascular dysfunction, and also induces muscle atrophy, bone loss, as well a depression of immune function [3].
Cell adhesion is a prerequisite for multicellularity in both evolution and individual development, providing a basis for the integrity of organisms [4]. The spatial arrangement of cells mediated by adhesion plays important roles in physiological states such as tissue morphogenesis and wound healing, as well as the progression of pathological conditions such as cancer [5,6,7]. Biological tissues are primarily remodeled through the interaction of cells with their microenvironment via adhesion molecules, but also through the secretion and adsorption of soluble factors [8]. Thus, cell adhesion also plays an important role in the physiological changes observed under microgravity [9]. Many different cell adhesion molecules, which are transmembrane proteins, regulate homophile (cell–cell adhesion) and homophile binding (cell–matrix adhesion) of cells [4].
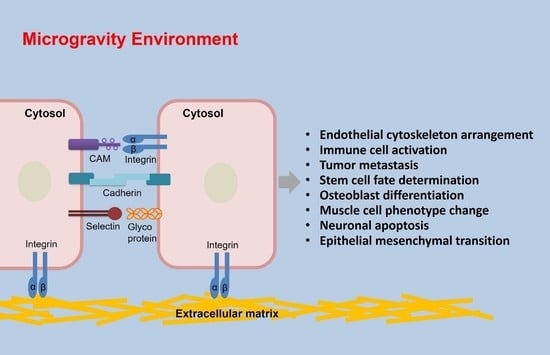
In this review, we introduce the biological functions of cell adhesion and summarize the roles of the four major classes of cellular adhesion molecules (CAMs) on cell adhesion, namely the integrins, immunoglobulin superfamily (Ig-SF) proteins, cadherins, and selectin in regulating cell adhesion. Furthermore, the effects of microgravity on the adhesion of endothelial cells, immune cells, cancer cells, stem cells, osteoblasts, muscle cells, and other types of cells both during spaceflight and under simulated conditions, are reviewed.
2. The Biological Functions of Cell Adhesion
Physiological processes such as embryonic development and tissue homeostasis, as well as pathological conditions such as cancer, rely on the temporal and spatial control of cell adhesion [8]. Adhesion allows cells to transfer forces, sense their surrounding microenvironment, and coordinate their biological behaviors [10]. Both cell–cell and cell–ECM adhesion have unique components, but both are directly linked to the cell’s actin cytoskeleton [11]. Thus, mechanical and/or chemical stimuli are transferred and integrated to induce or control cell proliferation, migration, and differentiation.
Intercellular adhesion is among the most important aspects. For example, cell–cell adhesion within the vessel wall controls various dynamic processes of endothelial cells, including angiogenesis and vascular remodeling [12]. The observation that vascular endothelial cadherin (VE-cadherin) knockout is embryonically lethal in mice because of defects in blood vessel pattern formation demonstrated the importance of VE-cadherin-based adherens junctions in maintaining the function of the vascular barrier [13]. Cell–cell contacts also crucially regulate cell survival, and in human keratinocytes, they constitute the main mechanism that controls proliferation, which is regulated by the junctional protein plakoglobin [14]. Similarly, apoptosis of granulosa cells is prevented by cell contact mediated by N-cadherin molecules on the surface membranes of adjacent cells [15]. In addition, cell migration is essential for embryonic morphogenesis, tissue remodeling and wound healing in adults, as well as pathological cancer cell migration. Intercellular adhesion promotes cancer cell mobility by increasing the polarization of individual cells, which plays an important role in the pathogenesis of tumors [16]. The dynamic regulation of cell–cell adhesion in neural crest cells is crucial for maintaining their migration capacity, which allows them to maintain tissue integrity [17]. In the process of wound healing, sheets of adhering keratinocytes can promote the migration of these cells into the wound [18].
In addition to the adhesion between cells, their adhesion to the extracellular matrix (ECM) via cell adhesion molecules is also crucial [19]. To migrate over the matrix, the cells first extend filopodia and lamellipodia, forming focal adhesions, and finally cytoplasmic tails are then retracted on opposite ends. Active complexes of ligands such as integrins promote the rearrangement of the cytoskeleton and activate intracellular signaling cascades. The events downstream of integrins, such as activation of focal adhesion kinase (FAK) and Src, or the recruitment of adaptor proteins such as zyxin and vinculin, depend on ligand binding and receptor clustering [20]. In a process known as cytoskeletal strengthening integrin-mediated cell–ECM adhesion is enhanced through interactions with cytoskeletal proteins, among which vinculin is important in stimulating actin polymerization and recruiting actin remodeling proteins. Both cell–matrix and cell–cell adhesion is significantly impaired when vinculin is removed [21]. Furthermore, the mechanical elasticity or stiffness of the ECM profoundly influences cell behavior. The mechanical cues of the ECM are sensed by yes-associated protein (YAP)/TAZ to activate specific gene expression and cell behaviors [22].
3. Role of Major Adhesion Molecules in Cell Adhesion
CAM is a general term for macromolecules responsible for the contact and binding between cells or between cells and ECM through receptor-ligand interactions. CAMs participate in many different physiological and pathological processes including inflammation, the immune response, blood coagulation, wound healing and tumor metastasis [23]. Here, we will introduce the four major classes of adhesion molecules that regulate cell adhesion: the integrins, Ig-SF proteins, cadherins, and selectin (Table 1).
3.1. Integrins
Integrins are a large family of type I transmembrane glycoproteins found in all animal lineages. They are non-covalently-linked αβ-heterodimeric receptors with a short cytoplasmic domain and a large extracellular domain. Mammalian cells possess integrins with 18 α-subunits (α1–11, αV, αIIb, αL, αM, αX, αD and αE) and eight β-subunits (β1–8), which are assembled into 24 known αβ-heterodimers [35]. Integrins are critical for cell adhesion, polarity, migration, as well as the formation and maintenance of normal tissue structure. Activated integrins play a pivotal role by forming a connection between the ECM and the actin cytoskeleton to facilitate cell adhesion [24]. Integrin-mediated adhesions beneath the lamellipodia initially form focal complexes, which further mature into longer lived and elongated focal adhesions. They serve as anchors for actin stress fibers, which in turn exert a force on the Fas to which they are connected [36]. In addition, the adhesion is mediated by several factors, including specific conformational affinity of integrin receptors to their cognate extracellular ligands, the mechanical force acting on the point of adhesion, the intracellular partners that bind integrins into discrete adhesive structures, and intracellular trafficking of integrins [37].
3.2. Immunoglobulin Superfamily
Ig-SF comprises the largest and most diverse class of cell adhesion molecules. The members of the Ig-SF are characterized by the presence of at least one Ig-like extracellular domain encompassing 70–110 amino acids that mediate calcium-independent cell adhesion [38]. Important examples of Ig-SF proteins include intercellular adhesion molecule (ICAM), vascular cell adhesion molecule (VCAM), and platelet endothelial cell adhesion molecule (PECAM). The adhesion mediated by these molecules is critical for normal immunological recognition, inflammation, and signal transduction, as well as pathological processes such as tumor metastasis.
3.2.1. ICAM-1
ICAM-1 is mainly expressed in epithelial cells, endothelial cells, leukocytes, and neutrophils. It contains five Ig-like domains and binds to β2 integrin molecules present on leukocytes, as well as MAC-1 (CD11b/CD18), and LFA-1 (CD11a/CD18) [39]. ICAM-1 mainly mediates cell–cell adhesion by binding to specific ligands in the ECM and surrounding cells [25]. It participates in the trafficking of inflammatory cells, in cell–cell interaction during antigen presentation, in signal transduction cascades through outside-in signaling, and in certain aspects of microbial pathogenesis [39]. Goh et al. demonstrated that artificial overexpression of ICAM-1 in myoblasts increases their mutual adhesion as well as the formation and size of myotubes [40]. Guo et al. reported that ICAM-1 is a crucial regulator of adhesion between endothelial progenitor cells (EPCs) and mesenchymal stem cells (MSCs) through p38/MAPK signaling pathway, it is of important for neovascularization and artificial bone regeneration [41]. Moreover, the enhanced expression of ICAM-1 in cancer cells is sufficient to mediate the adhesion of immune cells, which is correlated with poor prognosis [42]. These evidence prove that ICAM-1 plays an important role in cell adhesion.
3.2.2. VCAM-1
VCAM-1, also known as CD106, is a 90-kDa glycoprotein that is predominantly expressed by endothelial cells. Its expression is activated by ROS, oxidized low-density lipoprotein, high glucose concentrations, toll-like receptor agonists, shear stress, and pro-inflammatory cytokines such as TNFα [43]. Ligands of VCAM-1 include integrins α4β1, α4β7, αMβ2, α9β1, and αDβ2, which mediate firm adhesion among cells [44].
Because of the function of VCAM-1 in cell adhesion, it is essential, and a knockdown of VCAM-1 causes embryonic death in mice [45]. In vascular endothelial cells, inhibiting VCAM-1 expression has an anti-inflammatory effect by attenuating leukocyte adhesion to endothelial cells, and stimulated expression of VCAM-1 leads to increased adhesion of monocyte to endothelial cells. p38 MAP kinase and NF-κB signaling pathways might take part in regulating VCAM-1 expression [26,27]. In addition to endothelial cells, in cancer cells, knockdown of VCAM-1 in MDA231 breast cancer cells inhibited lung metastasis by reducing the adhesion to U937 leukocytes [46], the binding of VCAM-1 to α4β1 integrin might be responsible for tumor angiogenesis and metastasis [47]. Thus, VCAM-1 mediates the adhesion and tissue invasion of various types of cells.
3.2.3. PECAM-1
PECAM-1, also known as CD31, is a crucial mediator of the adhesion and accumulation of platelets [28]. Hematopoietic cells selectively express PECAM-1, and it is highly enriched at endothelial cell–cell junctions [48]. PECAM-1 contains six extracellular Ig-like domains that mediate the attraction and adhesion of leukocytes to endothelial cells, for example by enhancing eosinophil adhesion to IL-4-stimulated HUVECs in an α4β1 integrin-dependent manner [49,50]. Additionally, it also contains a transmembrane domain of 19-amino acids and a cytoplasmic tail encompassing 117-amino acid, which contains several phosphorylation sites that are important for intracellular signaling [51]. The cytoplasmic domain of PECAM-1 has two ITIM motifs that can be phosphorylated by Src kinase and recruit β-catenin and SHP-2 in endothelial cells. Since β-catenin is critical for adherent junctions of endothelial cells, PECAM1 can also inhibit the formation of endothelial cells connections [52].
3.3. Cadherins
Cadherins are a large superfamily of calcium-dependent transmembrane adhesive proteins that are responsible for cell–cell adhesion in all soft tissues as major adhesive proteins [4]. Extracellular regions of cadherin in a trans orientation mediate the adhesion between neighboring cells, while the cytoplasmic tails of cadherins interact with effector proteins such as catenin, which connects them to the actin cytoskeleton. Numerous physiologically and pathologically processes depend on the ability of cells to sense, transmit, and respond to mechanical forces and trigger intracellular signaling pathways, and these signals are mediated by adhesive complexes formed by cadherins [53].
Among the four main subfamilies, classical cadherins are the most studied and the other three such as desmosomal cadherins, protocadherins, and atypical cadherins are less well studied. Epithelial (E)-, neuronal (N)-, and vascular endothelial (VE)-cadherins derived from epithelial, neuronal, and vascular endothelial cells, respectively, are the prominent classical cadherins [54]. Adhesion mediated by E-cadherins is one of the main regulators of epithelial cell morphology and differentiation. The p120 catenin regulates the stability of E-cadherin, contributing to the maintenance of cell–cell adhesion in epithelial cells [29]. Recently, it was reported that tumor suppression depends on the normal functioning of E-cadherin in epithelial, mainly because of E-cadherin-mediated cell-to-cell adhesions [55]. Accordingly, the lack of E-cadherin aggravates the invasion and metastasis of many epithelial tumors. The full length or 90 kD N-terminal fragment of N-cadherin can promote cell–matrix adhesion [56]. In tumor cells, N-cadherin can promote tumor invasion via intercellular adhesions [30]. Additionally, the reorganization of the cytoskeleton during tumor cell migration and metastasis is regulated through synergistic effects between N-cadherin and fibroblast growth factor receptor (FGFR), which control the balance of intercellular and cell–matrix adhesions [57]. VE-cadherin is a major component of the adhesive connections between endothelial cells. The stability of VE-cadherin mediated adhesions depends on tyrosine phosphorylation of the VE-cadherin–catenin complex [31], which is proved by evidence that increased tyrosine phosphorylation of VE-cadherin and β-catenin is correlated with increased endothelial cell confluence [58]. As a result, classical cadherins mediate the cell–cell and cell–ECM adhesion, which is crucial for tissue morphogenesis and integrity.
3.4. Selectin
Selectins are a family of vascular cell adhesion molecules comprising of the three Ca2+-dependent lectins: E-selectin, L-selectin, and P-selectin. E-, L-, and P-selectin are expressed on the surface of endothelial cells, leucocytes, and platelets, respectively, and have similar structures encompassing an epidermal growth factor-like domain, a transmembrane domain, a sequence of repeats, and also a cytoplasmic tail [59]. P-selectin mediates the adhesion of platelets to tumor cells. L-selectin can promote the recruitment of marrow-derived cells, and accompanied by E-selectin promotes extravasation of tumor cells. Selectins mediate the interaction between tumor cells through the activation of integrins and secretion of pro-metastatic chemokines in the tumor microenvironment, contributing to metastasis and poor prognosis of cancer patients [32]. Moreover, the selectin–selectin ligand interactions of endothelial cells and tumor cells result in the extravasation of tumor cells [33]. Selectin also participates in constitutive lymphocyte homing. The adhesion and localization of leukocytes on vascular surfaces mediated by selectin is a key stage in the inflammatory process, which is accompanied by worsening of psoriasis, arthritis or asthma when aberrant homing of leukocytes occurs [34]. Therefore, evidence supports that selectins are important for physiological and pathological processes including inflammation, the immune response, and cancer metastasis.
4. Effects of Microgravity on Cell Adhesion
4.1. Microgravity Regulates Adhesion and Cytoskeleton Arrangement of Endothelial Cells
Microgravity is a stress factor that puts a burden on human cardiovascular physiology and functioning. Under microgravity, endothelial cell-related arterial remodeling occurs, which is a major cause of orthostatic intolerance in astronaut’s return to normal gravity [60]. Endothelial cells can be induced to express adhesion molecules and secrete cytokines as signals for cells in the bloodstream. Gene profile analysis of human umbilical vein endothelial cells (HUVECs) cultured on the ISS revealed that the expression of cell adhesion, such as VCAM-1, ICAM-1, and CD44 showed significant changes of expression compared with ground-based control cultures, along with the depolymerization of microtubules and actin filaments in endothelial cells, which is also accompanied by the decreased secretion of pro-angiogenetic and pro-inflammatory cytokines [61,62]. In addition, under simulated microgravity, the expression of adhesion molecules by endothelial cells showed patterns similar to those observed in spaceflight experiments in real microgravity. After incubation in a random positioning machine (RPM) for 35 days, the expression of VCAM-1 and ICAM-1 was increased in endothelial cells [63], which has also been demonstrated by a rat model that expression of E-selectin and VCAM-1 in endothelial cells was elevated in basilar and carotid arteries under hindlimb unweighted (HLU) condition [64]. Therefore, in a ground-based simulated microgravity model, microgravity has the same effect on the expression of adhesion molecules in an endothelial cell in vivo and in vitro.
The expression of the adhesive protein can affect the adhesion ability and cytological changes of endothelial cells under microgravity. With the increased secretion of VCAM-1 and ICAM-1, prolonged microgravity induced tubular structures and the formation of multicellular spheroids formation from endothelial cells. Moreover, decreased expression of fibronectin protein in adherent cells helps to separate cells from monolayers because fibronectin is involved in cell adhesion to culture plates [63]. After exposure to microgravity in the rotary cell culture system (RCCS), there was a decrease in the cell adhesion rate and changes of cytoskeletal structures of HUVECs after return to normal gravity, which resulted in the initiation of cell apoptosis via mechanisms involving mTOR/Apaf-1 and miR-22 signaling. The decreased cell adhesion ability triggers reduced actin fiber formation that might contribute to growth inhibition of endothelial cells [65]. Therefore, in endothelial cells, microgravity exerts important regulatory effects on cell adhesion, actin filament arrangement, and tube formation abilities (Table 2).
4.2. Microgravity Regulates Adhesion and Activation of Immune Cells
The lack of gravity during space missions is one of the major stress factors that is responsible for the dysfunction of the inherent and adaptive immune system in astronauts [66]. Cells of the immune system are also sensitive to microgravity. The influence of microgravity on adhesion molecule of immune cells has been extensively studied in space missions. After 11 days on SpaceX CRS-3, primary human M1 macrophages displayed a reduced expression of ICAM-1 compared to controls on the ground [67]. The reduction of gravity-responsive ICAM-1 was activated by mechanically sensitive signals in the cell-polycarbonate binding region, leading to blocked CD4+ T lymphocyte activation and immune response. However, different from that, in macrophage-like differentiated human U937 cells, the expression of ICAM-1 was increased during 2D clinostat as well as on the orbital SIMBOX/Shenzhou-8 mission, while had no effect on ICAM-1 in nondifferentiated U937 cells. The changes in gene expression of ICAM-1 have a great influence on their interaction with T lymphocytes [68]. These results showed that the expression trend of adhesion molecules was different in types and differentiation stage of immune cells, and also in different microgravity conditions.
Responding to microgravity, adhesion proteins act as mechanosensors that mainly contributed to the activity, migration, and adhesion of immune cells. For example, Liu et al. reported that the adhesion of monocytes/macrophages to the endothelium of the common carotid artery of HLU rats was enhanced in vivo [69]. Further study demonstrated that the recruitment of monocyte to the aortic endothelium was significantly increased under simulated microgravity, along with NF-κB-mediated increased expression of E-selectin, VCAM-1 and monocyte chemoattractant protein (MCP-1) in the abdominal aorta of rats [70]. In addition, after short-duration spaceflight, the peripheral monocytes from astronauts showed no change in the number of cells but a significantly reduced expression of CD26L and major histocompatibility complex, class II, DR (HLA-DR), which are known regulators of cell adhesion between lymphocyte and endothelial cells [71]. Consistent with this finding, the ability of peripheral blood mononuclear cells to adhere to the ICAM-1 substrate was reduced under simulated microgravity and shear flow conditions in vitro [72]. Although the force from the blood flow plays a decisive role in the monocyte’s adhesion cascade and initial capture with endothelial by selectins [73], the adhesion ability of immune cells and endothelial cells can also be impaired in microgravity. Therefore, it can be speculated that microgravity can regulate the adhesion of immune cells to endothelial cells by regulating the expression of adhesion molecules, which may have an important effect on the activity of immune cells (Table 3).
4.3. Microgravity Inhibits Tumor Cells Adhesion
Microgravity was reported to affect the focal adhesions, cytoskeletal arrangement, and adhesion of cancer cells, which in turn affects their migration, proliferation, and metastasis. Focal adhesion is one of the mechanosensitive structures. Tumor cells are sensitive to mechanical forces through focal adhesion, leading to stress-dependent adhesion in the microgravity environment. In microgravity, the formation of focal adhesions (paxillin and vinculin) in BL6-10 melanoma cells was reduced, while FAK was recruited and activated, which led to enhanced apoptosis via FAK/RhoA-regulated mTORC1 and AMPK signaling, together with a reduction of proliferation and metastasis compared with control cells at normal gravity [74,75]. Consistent with with the findings in melanoma cells, microgravity was found to regulate the focal adhesions and associated signaling molecules in glioma cells (U251 cells), such as FAK and RhoA/Rock [76]. Moreover, in malignant human MCF-7 cells, the number and clustering of focal adhesions were reduced under simulated microgravity, along with downregulation of β1 and β4 integrin and FAK activity. [77]. However, in prostate cancer cells, the expression of Talin1, Vinculin, and Cdh1 involved in focal adhesions were upregulated in multicellular spheroids of cells when exposed to simulated microgravity [78]. Thus, as the component of focal adhesions, Vinculin, FAK, and RhoA are the most important molecules that transmit external integrin-mediated signals into the cell, the biochemical pathways, and functions of tumor cells could be influenced by the disruption of adhesion structures.
Similarly, cytoskeleton as another mechanically sensitive structure also participate in tumor cell adhesion. For example, in adherent human lung cancer cells (cell line CRL-5889), they showed attenuated adhesion with a spherical arrangement of the actin filaments under simulated microgravity, and with accelerated apoptosis [79]. In addition, along with reduced focal adhesions, the microfilaments and microtubules were disrupted in MCF-7 cells, resulting in reduced cell migration [77]. Consistent with this, Nassef et al. demonstrated that the F-actin and tubulin were rearranged and the filopodia- and lamellipodia-like structures were appeared during early microgravity, accompanied with decreased expression of E-cadherin and β1 integrin in MCF-7 cells [80]. The expression of cell junction protein E-cadherin was reduced through the regulation of the E-cadherin autodegradation pathway, leading to multicellular spheroids formation of MCF-7 cells [81]. Taken together, during microgravity exposure, the synergistic effect of cytoskeleton and E-cadherin is involved in the tumor cell adhesion and multicellular spheroids formation process, which might eventually lead to tumorigenesis.
The effects of real and ground-based simulated microgravity on the expression and structure of the adhesion molecule in cancer cells are different. Kopp and colleagues demonstrated that real microgravity induced the expression of VCAM1 and ICAM1 in thyroid cancer cells (FTC-133 cell line) during sounding rocket flight [82]. However, ground-based studies revealed that the expression of ICAM1 protein was lower in RPM-induced thyroid cancer cells. Moreover, focal adhesion factor cofilin has different performance in real and simulated microgravity. Real microgravity elevated the expression of F-actin-binding protein cofilin, while the expression of cofilin was reduced in the RPM sample with disorganized vinculin structure [82]. Due to the link between the ECM and the cytoskeleton, the different expression patterns of VCAM1, ICAM1, and cofilin might influence cell adhesion in real and simulated microgravity through regulating actin cytoskeleton dynamics. Moreover, in MDA-MB-231 cells, the protein expression of ICAM1 and VCAM-1 was increased after the 31st parabolic flight. However, there was no expression change of ICAM1 and VCAM1 in cells under RPM-exposure [83]. These results may indirectly reflect the different biological effects of real and simulated microgravity. Further studies are still needed to fully reveal the different dynamic molecule changes of tumor cell adhesion in real and simulated microgravity at the single-molecule level (Table 4).
4.4. Microgravity Regulates Adhesion and Fate Determination of Stem Cells
Stem cells are a type of cells with self-renewal capacity and multi-directional differentiation potential. Microgravity has been shown to provide important signals for the fate of stem cells. The adhesion ability and expression of related genes in many stem cells were also observed to change under microgravity. Ratushnyy et al. reported that a total of 84 focal adhesion genes were identified as differentially expressed in multipotent mesenchymal stromal cells after 96 h of simulated microgravity [84]. Among them, the expression of ITGA11, ITGAV, ITGB1, DOCK1, ROCK2, PTEN, FAK, ARHGAP5, and AKT3 was downregulated [84]. These are genes that are related to the cytoskeleton and might be a reason for the reduced adhesion of MSCs in microgravity.
Most of the literature supports the relation between adhesion molecule expression and cytoskeletal structure and adhesion ability of the cell. With the increasing time under simulated microgravity, the number of VCAM-1-positive human MSCs (hMSCs) and the expression of ICAM-1 were decreased. Moreover, the number of hMSCs with a disordered actin cytoskeleton gradually increased and the cells showed a contracted phenotype [85,86]. Besides, in rat MSCs on board the SJ-10 Satellite, the expression of VCAM1, ICAM1, CD44, and vinculin was reduced along with depolymerization of actin filaments and the accumulation of microtubules due to true weightlessness in space [87]. With experiments in vitro and in vivo, it was demonstrated that redistribution of vinculin also occurred in hMSCs after 6 h of simulated microgravity [85], and the number of vinculin-containing focal adhesions was decreased in rat MSCs after 28 days of HLU [88]. Therefore, compared with conventional culture conditions, microgravity causes a part of stem cells to show a contracted state, the focal adhesion was changed, and the cytoskeleton was rearranged, which might be the causes of affecting cell adhesion ability.
On the other hand, changes in stem cell adhesion ability can determine its differentiation fate. In hMSCs, stimulated microgravity impaired the autophosphorylation of FAK and proline-rich tyrosine kinase 2 (PYK2), which might lead to reduced attachment and spreading of cells. Moreover, as the important positive regulator of FAK in osteogenic differentiation, microgravity inhibited osteogenesis through MAPK/ERK/Runx2 pathway [89]. Runx2 is an important transcription factor in osteoblastogenesis. With the reduced vinculin-containing focal adhesions, MSCs from HLU Rat exhibited decreased osteogenic potential in the osteogenic conditions with reduced transcriptional activity of Runx2 [88]. Besides MSCs, in adipose-derived stem cells (ADSCs), CD44, β1 integrin, ColIII, and MMP1 were induced and cells performed an aggregated structure after 7 days of microgravity simulation. The up-regulation of CD44 might indicate the undifferentiated stage of ADSCs under simulated microgravity conditions [90]. So, we can infer that reduced cell adhesion capacity under microgravity conditions is important for stem cells to maintain their stemness and increase their potential for multi-directional differentiation.
Beyond that, other researchers have demonstrated that although microgravity destroys the adhesion ability of stem cells, it can also drive cell differentiation and transdifferentiation. Lu et al. demonstrated that space microgravity not only reduces adhesion molecule expression but also promote the hepatogenic differentiation of MSCs [87]. The upregulation of hepatocyte-specific cytokeratin 18 and albumin makes cell mature [87]. In addition, human lung cancer stem cells (NSCLCs were used) detached from the substrate and performed an apoptosis state after exposure to microgravity, leading to the loss of stemness features with decreased Nanog and Oct4 genes [91]. Therefore, microgravity induces stem cell lineages to selectively differentiate into different phenotypes by changing their cell cycle. Furthermore, real microgravity experiments on SJ-10 satellite showed that the expression and activation of Runx2 were inhibited, and osteogenic differentiation was inhibited via the BMP2/SMAD and integrin/FAK/ERK pathways in hMSCs. Besides, adipogenic differentiation was promoted by increasing AKT and p38 MAPK activities [92]. Although microgravity inhibits stem cell adhesion, due to the diversity of cell types and functions, the effect of microgravity on stem cell differentiation is different. Hence, the molecular mechanism of stem cells’ response to microgravity is still needed to be further investigated (Table 5).
4.5. Microgravity Inhibits Osteoblast Adhesion and Differentiation
Astronauts often develop significant osteoporosis-like loss of bone mass, which has been attributed to microgravity. In the absence of gravity, there may be physiological changesin the cytoskeletal arrangement, adhesion, growth, and differentiation of bone cells [93]. The expression of adhesion molecules and ECMs in osteoblasts changed under the condition of microgravity. Kumei et al. reported that, compared to ground controls, the expression of osteopontin (OPN) was decreased in rat osteoblasts after exposing the cultures to microgravity in flight [94], which might lead to weakened cell adhesion ability because OPN promotes the attachment of cells to the bone surface in bone metabolism. On the other hand, stimulated microgravity increased the expression of OPN in human fetal hFOB 1.19 osteoblasts, and ECM such as laminin subunit α1, collagen type 1 α1, and fibronectin 1. However, these gene changes allowed the adhesive cells to detach from the surface and formed 3D structures [95]. Moreover, ex vivo experiment showed that simulated microgravity inhibits the expression of β1 integrin in human primary osteoblasts, and leads to a mesenchymal-like phenotype, indicating that osteogenesis is impaired [96]. Therefore, although microgravity has different regulatory effects on gene expression in osteoblasts derived from different species, it has the same effect on cell adhesion.
In addition, under the condition of microgravity, the number and structure of focal adhesions in osteoblasts were also changed. Exposure of primary osteoblasts and osteoblastic ROS 17/2.8 cells to microgravity reduced the number and average size of focal adhesions compared to control cells on the ground [93,97]. Furthermore, changes in the adhesion of osteoblasts affected the changes in the cytoskeleton and activation of intracellular signals, which determined the biological function of cells. Integrin-mediated cell adhesion in osteoblastic ROS 17/2.8 cells under microgravity was affected in parabolic flight and on a clinostat, and the changes were accompanied by disorganization of the cytoskeleton and disassembly of vinculin spots [98]. Actin filament rearrangement and focal adhesion formation might be regulated through the Rho GTPase signaling pathway [99]. The disrupted F-actin stress fibers and integrin signaling inhibited by focal adhesion kinase further inhibit osteoblastic differentiation [100]. Obviously, it is reasonable to think that the bone loss in weightlessness might arise from the repression of osteoblastogenesis via adhesion inhibition activation (Table 6).
4.6. Microgravity Regulates the Adhesion and Phenotype of Muscle Cells
During the space flight in the microgravity environment, zero gravity can have deleterious effects on muscle cells, especially skeletal muscle, including the expression changes of muscle growth/atrophy related genes, phenotype, and filaments structure changes of muscle cells, leading to arterial remodeling and muscle atrophy [101].
Recent studies have shown that multiple mechanisms are involved in microgravity-induced arterial remodeling. Kang et al. demonstrated that simulated microgravity disrupted the well-organized cytoskeleton in vascular smooth muscle cells (VSMCs), with reduced proliferation and promoted apoptosis [102]. Moreover, VSMCs changed from a synthetic phenotype to a contractile phenotype, with upregulation of smMHC expression [102]. In addition, with an in vivo experiment, the number of focal adhesions in the basilar artery was increased after 4-week HLU. The expression of p-FAK Y397 and p-Src Y418 was increased in SMCs of basilar arteries [103]. Activation of FAK combined with focal adhesions is an important event in early mechanical transduction and plays a critical role in vascular remodeling.
Additionally, in human skeletal muscle stem/progenitor cells (SMPCs) after 2 weeks of microgravity culture, the number of adherent cells was significantly reduced and cells formed into spheres with reduced numbers of myotubes. Moreover, the expression of Pax7 was also decreased in SMPC spheres through the TRAF6/ERK pathway [104]. Pax7 is a transcriptional factor critical for muscle development and regeneration [105]. Therefore, microgravity imbalanced the stability of SMPC pool, leading to insufficient muscle regeneration. Taken together, the adhesion of muscle cells plays an important role in regulating muscle morphogenesis. Thus, reduced adhesion of muscle cells might be responsible for the vascular remodeling and muscle atrophy under microgravity (Table 7).
4.7. The Function of Microgravity on Adhesion of Other Cell Types
There is increasing evidence that microgravity in space has many negative effects on nervous systems, visual function, as well as the skins of astronauts. The function of microgravity on the adhesion of cells in these organs has also been demonstrated.
Under simulated microgravity, the adhesion ability of primary cells from human brain nervous tissue was decreased, accompanied by highly disorganized β-tubulin structures in a circular pattern around the nucleus [106]. The cytoskeletal structure and distribution changes might be the main reason for changes in cell adhesion. Moreover, cells exhibited increased apoptosis, and it can be assumed that the function of the entire nervous system may be threatened by simulated or real microgravity. In addition, microgravity can also reduce the expression of β1 integrin, β3 integrin, and laminin subunit β-2 (LAMB2) in human adult retinal pigment epithelium cells [107]. As integrins are pivotal for focal adhesion formation and cytoskeleton rearrangements, this might explain the reason behind the effect of microgravity in promoting the detachment of cells detached from culture plate to form multicellular spheroids.
Furthermore, the expression of the cell–cell adhesion molecule E-cadherin in human keratinocytes was reduced during short exposure to microgravity. Mesenchymal markers such as α-SMA and vimentin were upregulated [108]. This means that the cell–cell adhesion of keratinocytes was attenuated, leading to epithelial-mesenchymal transition and invasive phenotypes in the microgravity environment. Consistent with these findings, in normal human dermal fibroblasts (NHDF), focal adhesion, cytoskeleton, and growth behavior were also changed by simulated microgravity through regulating integrin-β1 and E-cadherin [109]. Taken together, microgravity affects the physiological function of organs by regulating cell adhesion, and further regulating cytoskeletal rearrangement and phenotypic changes, etc., so that the human body can perceive and adapt to changes in the gravity environment (Table 8).
5. Conclusions and Perspectives
Microgravity is one of the major challenges facing astronauts during spaceflight. Exposure to microgravity has been associated with various significant physiological changes of the bone tissue as well as the cardiovascular, nervous, and immune systems, as well as pathological changes such as atherosclerotic plaques or even cancer. The migration, proliferation, differentation, and apoptosis of mammalian cells are affected by changes in cell adhesion during long-duration spaceflights. In this review, we have introduced the biological functions of cell–cell and cell–matrix adhesion and have summarized the roles of different adhesion molecules including integrins, Ig-SF, cadherins, and selectin in regulating cell adhesion. Based on the available literature, we have reviewed the effects of spaceflight and simulated microgravity on the adhesion capacity of endothelial cells, immune cells, cancer cells, stem cells, osteoblasts, muscle cells, nerve cells, etc.
Although several spaceflight and ground-based experiments have been performed to study the effects of microgravity on the adhesion ability of different cell types, the mechanisms by which microgravity affects cell adhesion are not fully understood and need to be further investigated. Firstly, it is still unclear how exactly cells sense microgravity and transduce signaling through adhesive protein complex. The major genes that are involved in intracellular regulation are still unknown. Therefore, it should be clarified if the changes of expression or changes of cellular localization of adhesion protein lead to the different cell adhesion behaviors. The localization and expression of adhesion-related intracellular key regulator protein can be furtherly detected by in situ hybridization. Moreover, it is unknown whether the effect and mechanism of ground-based microgravity on cell adhesion are the same as that of real microgravity in space. The earth-based simulation techniques such as using RPM or HLU cannot fully mimic the zero gravity of the spaceflight. For example, in tumor cells, the expression of ICAM1 protein and the performance of focal adhesion such as vinculin and cofilin are different between real and ground-based simulated microgravity [82]. The expression of E-cadherin and cell adhesion state in HUVEC and MCF-7 cells showed different under simulated microgravity and overloading condition [110]. In addition, bauer et al. used mass spectrometry and mRNA microarray to compare the different effect of microgravity on sialylation of adhesion proteins in FTC-133 thyroid cancer cells, MCF7 cells, and EA-hy926 cells [111]. Thus, transcriptomics, proteomics, and even single-cell sequencing approaches can be used to compare the genomic and epigenomic changes of cells in different microgravity scenarios, which can provide molecular biology information and mechanism of cell adhesion under microgravity.
Furthermore, based on the adverse effects of microgravity on cell adhesion and further adverse influence on human physiology, drugs that targeted on the regulators of cell adhesion need to be developed for disease prevention or treatment of astronauts during spaceflight. For example, Romswinkel et al. demonstrated that the key regulator tumor cell adhesion, CXCR4, was upregulated in spheroids of Ewing’s Sarcoma cells under simulated microgravity [112]. Therefore, the authors tried to use CXCR4 inhibitor to regulate the cell adhesion. However, it was found that inhibiting of CXCR4 did not change the size and number of spheroids [112]. This is because the transcriptional and post-translational modifications change the function of CXCR4. Therefore, small nucleic acid such as siRNA or miRNA which can directly target on and regulate the key effectors that mediate the mechanisms of cell adhesion should be studied. In addition, when screening targeted drugs, the selection of cell model that responds to simulated microgravity is also critical. It was reported that, compared with parental Jurkat cell line, the Jurkat/A4 cells are more sensitive to simulated microgravity with the changes of adhesion molecule ICAM-3 expression and cell morphology [113]. Therefore, Jurkat/A4 cells are useful models for studying microgravity effects and testing anti-cancer drugs. Cell adhesion is the basic determinant of biological development, and also one of the important influencing factors of diseases such as nerves and tumors [114,115]. Further studies should be undertaken to explore the effect and mechanism of regulators on cell adhesion in microgravity, especially for the development of new therapeutic targets and drugs that can greatly improve the health of astronauts on long space missions.
Author Contributions
X.L., K.Z., D.W., Y.T., Y.G., and Z.C. drafted the manuscript. A.Q. designed the project. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by China Postdoctoral Science Foundation, 2017M613196; Fundamental Research Funds for the Central Universities, 3102019ghxm012.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| CAM | Cell adhesion molecules |
| ECM | Extracellular matrix |
| Ig-SF | Immunoglobulin superfamily |
| ISS | International space station |
| VE-cadherin | Vascular endothelial cadherin |
| FAK | Focal adhesion kinase |
| YAP | Yes-associated protein |
| ICAM | Intercellular adhesion molecule |
| VCAM | Vascular cell adhesion molecule |
| PECAM | Platelet endothelial cell adhesion molecule |
| EPCs | Endothelial progenitor cells |
| MSCs | Mesenchymal stem cells |
| FGFR | Fibroblast growth factor receptor |
| HUVEC | Human umbilical vein endothelial cells |
| RPM | Random positioning machine |
| HLU | Hindlimb unweighted |
| RCCS | Rotary cell culture system |
| MCP-1 | Monocyte chemoattractant protein |
| HLA-DR | Major histocompatibility complex, class II, DR |
| PYK2 | Proline-rich tyrosine kinase 2 |
| ADSCs | Adipose-derived stem cells |
| OPN | Osteopontin |
| VSMCs | Vascular smooth muscle cells |
| SMPCs | Skeletal muscle stem/progenitor cells |
| LAMB2 | Laminin subunit β-2 |
| NHDF | Human dermal fibroblasts |
References
- Aleshcheva, G.; Bauer, J.; Hemmersbach, R.; Slumstrup, L.; Wehland, M.; Infanger, M.; Grimm, D. Scaffold-free Tissue Formation Under Real and Simulated Microgravity Conditions. Basic Clin. Pharmacol. Toxicol. 2016, 119 (Suppl. S3), 26–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anil-Inevi, M.; Yilmaz, E.; Sarigil, O.; Tekin, H.C.; Ozcivici, E. Single Cell Densitometry and Weightlessness Culture of Mesenchymal Stem Cells Using Magnetic Levitation. Methods Mol. Biol. 2019. [Google Scholar] [CrossRef]
- Tanaka, K.; Nishimura, N.; Kawai, Y. Adaptation to microgravity, deconditioning, and countermeasures. J. Physiol. Sci. 2017, 67, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Niessen, C.M.; Leckband, D.; Yap, A.S. Tissue organization by cadherin adhesion molecules: Dynamic molecular and cellular mechanisms of morphogenetic regulation. Physiol. Rev. 2011, 91, 691–731. [Google Scholar] [CrossRef] [Green Version]
- Weber, G.F.; Bjerke, M.A.; DeSimone, D.W. Integrins and cadherins join forces to form adhesive networks. J. Cell Sci. 2011, 124, 1183–1193. [Google Scholar] [CrossRef] [Green Version]
- Desai, R.A.; Gao, L.; Raghavan, S.; Liu, W.F.; Chen, C.S. Cell polarity triggered by cell-cell adhesion via E-cadherin. J. Cell Sci. 2009, 122, 905–911. [Google Scholar] [CrossRef] [Green Version]
- Jhala, D.V.; Kale, R.K.; Singh, R.P. Microgravity alters cancer growth and progression. Curr. Cancer Drug Targets 2014, 14, 394–406. [Google Scholar] [CrossRef]
- Singh, J.; Hussain, F.; Decuzzi, P. Role of differential adhesion in cell cluster evolution: From vasculogenesis to cancer metastasis. Comput. Methods Biomech. Biomed. Engin. 2015, 18, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Masiello, M.G.; Verna, R.; Cucina, A.; Bizzarri, M. Physical constraints in cell fate specification. A case in point: Microgravity and phenotypes differentiation. Prog. Biophys. Mol. Biol. 2018, 134, 55–67. [Google Scholar] [CrossRef]
- Bajanca, F.; Alfandari, D.; Thorsteinsdottir, S.; Theveneau, E. Editorial: Cell adhesion in development. Dev. Biol. 2015, 401, 1. [Google Scholar] [CrossRef] [Green Version]
- Marjoram, R.J.; Lessey, E.C.; Burridge, K. Regulation of RhoA activity by adhesion molecules and mechanotransduction. Curr. Mol. Med. 2014, 14, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schimmel, L.; Gordon, E. The precise molecular signals that control endothelial cell-cell adhesion within the vessel wall. Biochem. Soc. Trans. 2018, 46, 1673–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmeliet, P.; Lampugnani, M.G.; Moons, L.; Breviario, F.; Compernolle, V.; Bono, F.; Balconi, G.; Spagnuolo, R.; Oosthuyse, B.; Dewerchin, M.; et al. Targeted deficiency or cytosolic truncation of the VE-cadherin gene in mice impairs VEGF-mediated endothelial survival and angiogenesis. Cell 1999, 98, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Hariharan, V.; Huang, H. Cell-cell contact preserves cell viability via plakoglobin. PLoS ONE 2011, 6, e27064. [Google Scholar] [CrossRef] [Green Version]
- Peluso, J.J.; Pappalardo, A.; Trolice, M.P. N-cadherin-mediated cell contact inhibits granulosa cell apoptosis in a progesterone-independent manner. Endocrinology 1996, 137, 1196–1203. [Google Scholar] [CrossRef]
- Venhuizen, J.H.; Zegers, M.M. Making Heads or Tails of It: Cell-Cell Adhesion in Cellular and Supracellular Polarity in Collective Migration. Cold Spring Harb. Perspect. Biol. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Barriga, E.H.; Mayor, R. Embryonic cell-cell adhesion: A key player in collective neural crest migration. Curr. Top. Dev. Biol. 2015, 112, 301–323. [Google Scholar] [CrossRef]
- Nardini, J.T.; Chapnick, D.A.; Liu, X.; Bortz, D.M. Modeling keratinocyte wound healing dynamics: Cell-cell adhesion promotes sustained collective migration. J. Theor. Biol. 2016, 400, 103–117. [Google Scholar] [CrossRef] [Green Version]
- Maziveyi, M.; Alahari, S.K. Cell matrix adhesions in cancer: The proteins that form the glue. Oncotarget 2017, 8, 48471–48487. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, N.C.; Roca-Cusachs, P. Mechanosensing at integrin-mediated cell-matrix adhesions: From molecular to integrated mechanisms. Curr. Opin. Cell Biol. 2018, 50, 20–26. [Google Scholar] [CrossRef]
- Bays, J.L.; DeMali, K.A. Vinculin in cell-cell and cell-matrix adhesions. Cell Mol. Life Sci. 2017, 74, 2999–3009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, S. Role of YAP/TAZ in cell-matrix adhesion-mediated signalling and mechanotransduction. Exp. Cell Res. 2016, 343, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Mousa, S.A. Cell adhesion molecules: Potential therapeutic & diagnostic implications. Mol. Biotechnol. 2008, 38, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Tamkun, J.W.; DeSimone, D.W.; Fonda, D.; Patel, R.S.; Buck, C.; Horwitz, A.F.; Hynes, R.O. Structure of integrin, a glycoprotein involved in the transmembrane linkage between fibronectin and actin. Cell 1986, 46, 271–282. [Google Scholar] [CrossRef]
- Hubbard, A.K.; Rothlein, R. Intercellular adhesion molecule-1 (ICAM-1) expression and cell signaling cascades. Free Radic. Biol. Med. 2000, 28, 1379–1386. [Google Scholar] [CrossRef]
- Lee, B.K.; Lee, W.J.; Jung, Y.S. Chrysin Attenuates VCAM-1 Expression and Monocyte Adhesion in Lipopolysaccharide-Stimulated Brain Endothelial Cells by Preventing NF-kappaB Signaling. Int. J. Mol. Sci. 2017, 18, 1424. [Google Scholar] [CrossRef] [Green Version]
- Toma, L.; Sanda, G.M.; Deleanu, M.; Stancu, C.S.; Sima, A.V. Glycated LDL increase VCAM-1 expression and secretion in endothelial cells and promote monocyte adhesion through mechanisms involving endoplasmic reticulum stress. Mol. Cell Biochem. 2016, 417, 169–179. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, Q.; Liu, S.; Parajuli, K.R.; Qu, Y.; Mei, J.; Chen, Z.; Zhang, H.; Khismatullin, D.B.; You, Z. IL-17 and insulin/IGF1 enhance adhesion of prostate cancer cells to vascular endothelial cells through CD44-VCAM-1 interaction. Prostate 2015, 75, 883–895. [Google Scholar] [CrossRef] [Green Version]
- Coopman, P.; Djiane, A. Adherens Junction and E-Cadherin complex regulation by epithelial polarity. Cell Mol. Life Sci. 2016, 73, 3535–3553. [Google Scholar] [CrossRef]
- Mrozik, K.M.; Blaschuk, O.W.; Cheong, C.M.; Zannettino, A.C.W.; Vandyke, K. N-cadherin in cancer metastasis, its emerging role in haematological malignancies and potential as a therapeutic target in cancer. BMC Cancer 2018, 18, 939. [Google Scholar] [CrossRef]
- Vestweber, D. VE-cadherin: The major endothelial adhesion molecule controlling cellular junctions and blood vessel formation. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laubli, H.; Borsig, L. Selectins promote tumor metastasis. Semin. Cancer Biol. 2010, 20, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witz, I.P. The selectin-selectin ligand axis in tumor progression. Cancer Metastasis Rev. 2008, 27, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Angiari, S. Selectin-mediated leukocyte trafficking during the development of autoimmune disease. Autoimmun Rev. 2015, 14, 984–995. [Google Scholar] [CrossRef]
- Campbell, I.D.; Humphries, M.J. Integrin structure, activation, and interactions. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [Green Version]
- Seetharaman, S.; Etienne-Manneville, S. Integrin diversity brings specificity in mechanotransduction. Biol. Cell 2018, 110, 49–64. [Google Scholar] [CrossRef]
- Iwamoto, D.V.; Calderwood, D.A. Regulation of integrin-mediated adhesions. Curr. Opin. Cell Biol. 2015, 36, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Matthaus, C.; Langhorst, H.; Schutz, L.; Juttner, R.; Rathjen, F.G. Cell-cell communication mediated by the CAR subgroup of immunoglobulin cell adhesion molecules in health and disease. Mol. Cell Neurosci. 2017, 81, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Malik, P.; Arora, S.K.; Mukherjee, T.K. Intercellular adhesion molecule-1 as a drug target in asthma and rhinitis. Respirology 2014, 19, 508–513. [Google Scholar] [CrossRef]
- Goh, Q.; Dearth, C.L.; Corbett, J.T.; Pierre, P.; Chadee, D.N.; Pizza, F.X. Intercellular adhesion molecule-1 expression by skeletal muscle cells augments myogenesis. Exp. Cell Res. 2015, 331, 292–308. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Zhang, H.; Xia, J.; Hou, J.; Wang, Y.; Yang, T.; Wang, S.; Zhang, X.; Chen, X.; Wu, X. Interleukin-1beta induces intercellular adhesion molecule-1 expression, thus enhancing the adhesion between mesenchymal stem cells and endothelial progenitor cells via the p38 MAPK signaling pathway. Int. J. Mol. Med. 2018, 41, 1976–1982. [Google Scholar] [CrossRef]
- Roland, C.L.; Dineen, S.P.; Toombs, J.E.; Carbon, J.G.; Smith, C.W.; Brekken, R.A.; Barnett, C.C., Jr. Tumor-derived intercellular adhesion molecule-1 mediates tumor-associated leukocyte infiltration in orthotopic pancreatic xenografts. Exp. Biol. Med. 2010, 235, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.H.; Kim, Y.K.; Kim, M.R.; Jang, J.H.; Lee, S. Emerging Roles of Vascular Cell Adhesion Molecule-1 (VCAM-1) in Immunological Disorders and Cancer. Int. J. Mol. Sci. 2018, 19, 1057. [Google Scholar] [CrossRef] [Green Version]
- Schlesinger, M.; Bendas, G. Vascular cell adhesion molecule-1 (VCAM-1)--an increasing insight into its role in tumorigenicity and metastasis. Int. J. Cancer 2015, 136, 2504–2514. [Google Scholar] [CrossRef] [PubMed]
- Cook-Mills, J.M.; Marchese, M.E.; Abdala-Valencia, H. Vascular cell adhesion molecule-1 expression and signaling during disease: Regulation by reactive oxygen species and antioxidants. Antioxid. Redox Signal. 2011, 15, 1607–1638. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Zhang, X.H.; Massague, J. Macrophage binding to receptor VCAM-1 transmits survival signals in breast cancer cells that invade the lungs. Cancer Cell 2011, 20, 538–549. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Sharma, R.; Khaket, T.P.; Dutta, C.; Chakraborty, B.; Mukherjee, T.K. Breast cancer metastasis: Putative therapeutic role of vascular cell adhesion molecule-1. Cell Oncol. 2017, 40, 199–208. [Google Scholar] [CrossRef]
- Newman, P.J. The biology of PECAM-1. J. Clin. Investig. 1997, 100, S25–S29. [Google Scholar] [CrossRef] [Green Version]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. Endothelial PECAM-1 and its function in vascular physiology and atherogenic pathology. Exp. Mol. Pathol. 2016, 100, 409–415. [Google Scholar] [CrossRef]
- Chiba, R.; Nakagawa, N.; Kurasawa, K.; Tanaka, Y.; Saito, Y.; Iwamoto, I. Ligation of CD31 (PECAM-1) on endothelial cells increases adhesive function of alphavbeta3 integrin and enhances beta1 integrin-mediated adhesion of eosinophils to endothelial cells. Blood 1999, 94, 1319–1329. [Google Scholar] [CrossRef]
- Snyder, J.L.; McBeath, E.; Thomas, T.N.; Chiu, Y.J.; Clark, R.L.; Fujiwara, K. Mechanotransduction properties of the cytoplasmic tail of PECAM-1. Biol. Cell 2017, 109, 312–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, A.; Shively, J.E. Angiopoietins-1 and -2 play opposing roles in endothelial sprouting of embryoid bodies in 3D culture and their receptor Tie-2 associates with the cell-cell adhesion molecule PECAM1. Exp. Cell Res. 2011, 317, 2171–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasch, J.; Harrison, O.J.; Honig, B.; Shapiro, L. Thinking outside the cell: How cadherins drive adhesion. Trends Cell Biol. 2012, 22, 299–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, K.H.; Zaidel-Bar, R. Early events in the assembly of E-cadherin adhesions. Exp. Cell Res. 2017, 358, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Bruner, H.C.; Derksen, P.W.B. Loss of E-Cadherin-Dependent Cell-Cell Adhesion and the Development and Progression of Cancer. Cold Spring Harb. Perspect. Biol. 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Derycke, L.D.; Bracke, M.E. N-cadherin in the spotlight of cell-cell adhesion, differentiation, embryogenesis, invasion and signalling. Int. J. Dev. Biol. 2004, 48, 463–476. [Google Scholar] [CrossRef]
- Nguyen, T.; Mege, R.M. N-Cadherin and Fibroblast Growth Factor Receptors crosstalk in the control of developmental and cancer cell migrations. Eur. J. Cell Biol. 2016, 95, 415–426. [Google Scholar] [CrossRef]
- Lampugnani, M.G.; Corada, M.; Andriopoulou, P.; Esser, S.; Risau, W.; Dejana, E. Cell confluence regulates tyrosine phosphorylation of adherens junction components in endothelial cells. J. Cell Sci. 1997, 110 Pt 17, 2065–2077. [Google Scholar]
- McEver, R.P. Selectins: Initiators of leucocyte adhesion and signalling at the vascular wall. Cardiovasc Res. 2015, 107, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Jia, G.; Bao, J.; Zhang, Y.; Bai, Y.; Lin, L.; Tang, H.; Ma, J. Increased vascular cell adhesion molecule-1 was associated with impaired endothelium-dependent relaxation of cerebral and carotid arteries in simulated microgravity rats. J. Physiol. Sci. 2008, 58, 67–73. [Google Scholar] [CrossRef]
- Versari, S.; Longinotti, G.; Barenghi, L.; Maier, J.A.; Bradamante, S. The challenging environment on board the International Space Station affects endothelial cell function by triggering oxidative stress through thioredoxin interacting protein overexpression: The ESA-SPHINX experiment. FASEB J. 2013, 27, 4466–4475. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, C.; Sun, S.; Zhang, C.; Lu, D.; Chen, Q.; Long, M. Microgravity-Induced Alterations of Inflammation-Related Mechanotransduction in Endothelial Cells on Board SJ-10 Satellite. Front. Physiol. 2018, 9, 1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittrich, A.; Grimm, D.; Sahana, J.; Bauer, J.; Kruger, M.; Infanger, M.; Magnusson, N.E. Key Proteins Involved in Spheroid Formation and Angiogenesis in Endothelial Cells After Long-Term Exposure to Simulated Microgravity. Cell Physiol. Biochem. 2018, 45, 429–445. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Ran, H.H.; Gao, Y.L.; Ma, J.; Huang, Y.; Bai, Y.G.; Lin, L.J. Differential vascular cell adhesion molecule-1 expression and superoxide production in simulated microgravity rat vasculature. EXCLI J. 2010, 9, 195–204. [Google Scholar]
- Xu, D.; Guo, Y.B.; Zhang, M.; Sun, Y.Q. The subsequent biological effects of simulated microgravity on endothelial cell growth in HUVECs. Chin. J. Traumatol. 2018, 21, 229–237. [Google Scholar] [CrossRef]
- Moreno-Villanueva, M.; Feiveson, A.H.; Krieger, S.; Kay Brinda, A.; von Scheven, G.; Burkle, A.; Crucian, B.; Wu, H. Synergistic Effects of Weightlessness, Isoproterenol, and Radiation on DNA Damage Response and Cytokine Production in Immune Cells. Int. J. Mol. Sci. 2018, 19, 3689. [Google Scholar] [CrossRef] [Green Version]
- Tauber, S.; Lauber, B.A.; Paulsen, K.; Layer, L.E.; Lehmann, M.; Hauschild, S.; Shepherd, N.R.; Polzer, J.; Segerer, J.; Thiel, C.S.; et al. Cytoskeletal stability and metabolic alterations in primary human macrophages in long-term microgravity. PLoS ONE 2017, 12, e0175599. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, K.; Tauber, S.; Dumrese, C.; Bradacs, G.; Simmet, D.M.; Golz, N.; Hauschild, S.; Raig, C.; Engeli, S.; Gutewort, A.; et al. Regulation of ICAM-1 in cells of the monocyte/macrophage system in microgravity. Biomed. Res. Int. 2015, 2015, 538786. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Z.C.; Yue, Y.; Yu, J.W.; Cai, Y.; Bai, Y.G.; Zhang, H.J.; Bao, J.X.; Ren, X.L.; Xie, M.J.; et al. Simulated microgravity induces an inflammatory response in the common carotid artery of rats. Can. J. Physiol. Pharmacol. 2014, 92, 661–668. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Z.C.; Bai, Y.G.; Cai, Y.; Yu, J.W.; Zhang, H.J.; Bao, J.X.; Ren, X.L.; Xie, M.J.; Ma, J. Simulated microgravity promotes monocyte adhesion to rat aortic endothelium via nuclear factor-kappaB activation. Clin. Exp. Pharmacol. Physiol. 2015, 42, 510–519. [Google Scholar] [CrossRef]
- Crucian, B.; Stowe, R.; Quiriarte, H.; Pierson, D.; Sams, C. Monocyte phenotype and cytokine production profiles are dysregulated by short-duration spaceflight. Aviat Space Environ. Med. 2011, 82, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Moser, D.; Sun, S.J.; Li, N.; Biere, K.; Hoerl, M.; Matzel, S.; Feuerecker, M.; Buchheim, J.I.; Strewe, C.; Thiel, C.S.; et al. Cells Flow and Immune Cell Priming under alternating g-forces in Parabolic Flight. Sci. Rep. 2019, 9, 11276. [Google Scholar] [CrossRef] [PubMed]
- Rossy, J.; Laufer, J.M.; Legler, D.F. Role of Mechanotransduction and Tension in T Cell Function. Front. Immunol. 2018, 9, 2638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.; Xu, A.; Zhao, T.; Zhao, Q.; Zhang, J.; Fan, C.; Deng, Y.; Freywald, A.; Genth, H.; Xiang, J. Simulated microgravity inhibits cell focal adhesions leading to reduced melanoma cell proliferation and metastasis via FAK/RhoA-regulated mTORC1 and AMPK pathways. Sci. Rep. 2018, 8, 3769. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Li, R.; Tan, X.; Zhang, J.; Fan, C.; Zhao, Q.; Deng, Y.; Xu, A.; Lukong, K.E.; Genth, H.; et al. Simulated Microgravity Reduces Focal Adhesions and Alters Cytoskeleton and Nuclear Positioning Leading to Enhanced Apoptosis via Suppressing FAK/RhoA-Mediated mTORC1/NF-kappaB and ERK1/2 Pathways. Int. J. Mol. Sci. 2018, 19, 1994. [Google Scholar] [CrossRef] [Green Version]
- Deng, B.; Liu, R.; Tian, X.; Han, Z.; Chen, J. Simulated microgravity inhibits the viability and migration of glioma via FAK/RhoA/Rock and FAK/Nek2 signaling. In Vitro Cell. Dev. Biol. Anim. 2019, 55, 260–271. [Google Scholar] [CrossRef]
- Li, J.; Zhang, S.; Chen, J.; Du, T.; Wang, Y.; Wang, Z. Modeled microgravity causes changes in the cytoskeleton and focal adhesions, and decreases in migration in malignant human MCF-7 cells. Protoplasma 2009, 238, 23–33. [Google Scholar] [CrossRef]
- Hybel, T.E.; Dietrichs, D.; Sahana, J.; Corydon, T.J.; Nassef, M.Z.; Wehland, M.; Kruger, M.; Magnusson, N.E.; Bauer, J.; Utpatel, K.; et al. Simulated Microgravity Influences VEGF, MAPK, and PAM Signaling in Prostate Cancer Cells. Int. J. Mol. Sci. 2020, 21, 1263. [Google Scholar] [CrossRef] [Green Version]
- Dietz, C.; Infanger, M.; Romswinkel, A.; Strube, F.; Kraus, A. Apoptosis Induction and Alteration of Cell Adherence in Human Lung Cancer Cells under Simulated Microgravity. Int. J. Mol. Sci. 2019, 20, 3601. [Google Scholar] [CrossRef] [Green Version]
- Nassef, M.Z.; Kopp, S.; Wehland, M.; Melnik, D.; Sahana, J.; Kruger, M.; Corydon, T.J.; Oltmann, H.; Schmitz, B.; Schutte, A.; et al. Real Microgravity Influences the Cytoskeleton and Focal Adhesions in Human Breast Cancer Cells. Int. J. Mol. Sci. 2019, 20, 3156. [Google Scholar] [CrossRef] [Green Version]
- Sahana, J.; Nassef, M.Z.; Wehland, M.; Kopp, S.; Kruger, M.; Corydon, T.J.; Infanger, M.; Bauer, J.; Grimm, D. Decreased E-Cadherin in MCF7 Human Breast Cancer Cells Forming Multicellular Spheroids Exposed to Simulated Microgravity. Proteomics 2018, 18, e1800015. [Google Scholar] [CrossRef] [PubMed]
- Kopp, S.; Kruger, M.; Bauer, J.; Wehland, M.; Corydon, T.J.; Sahana, J.; Nassef, M.Z.; Melnik, D.; Bauer, T.J.; Schulz, H.; et al. Microgravity Affects Thyroid Cancer Cells during the TEXUS-53 Mission Stronger than Hypergravity. Int. J. Mol. Sci. 2018, 19, 4001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nassef, M.Z.; Kopp, S.; Melnik, D.; Corydon, T.J.; Sahana, J.; Kruger, M.; Wehland, M.; Bauer, T.J.; Liemersdorf, C.; Hemmersbach, R.; et al. Short-Term Microgravity Influences Cell Adhesion in Human Breast Cancer Cells. Int. J. Mol. Sci. 2019, 20, 5730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratushnyy, A.Y.; Buravkova, L.B. Expression of focal adhesion genes in mesenchymal stem cells under simulated microgravity. Dokl. Biochem. Biophys. 2017, 477, 354–356. [Google Scholar] [CrossRef] [PubMed]
- Gershovich, P.M.; Gershovich Iu, G.; Buravkova, L.B. Cytoskeleton structures and adhesion properties of human stromal precursors under conditions of simulated microgravity. Tsitologiia 2009, 51, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Koaykul, C.; Kim, M.H.; Kawahara, Y.; Yuge, L.; Kino-Oka, M. Alterations in Nuclear Lamina and the Cytoskeleton of Bone Marrow-Derived Human Mesenchymal Stem Cells Cultured Under Simulated Microgravity Conditions. Stem Cells Dev. 2019, 28, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Sun, S.; Zhang, F.; Luo, C.; Zheng, L.; Wu, Y.; Li, N.; Zhang, C.; Wang, C.; Chen, Q.; et al. Microgravity-induced hepatogenic differentiation of rBMSCs on board the SJ-10 satellite. FASEB J. 2019, 33, 4273–4286. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Zhang, C.; Chen, G.; Tang, Z.; Liu, Q.; Chen, J.; Tong, X.; Wang, J. Effects of BMP-2 and FGF2 on the osteogenesis of bone marrow-derived mesenchymal stem cells in hindlimb-unloaded rats. Cell. Biochem. Biophys. 2014, 70, 1127–1136. [Google Scholar] [CrossRef]
- Meyers, V.E.; Zayzafoon, M.; Gonda, S.R.; Gathings, W.E.; McDonald, J.M. Modeled microgravity disrupts collagen I/integrin signaling during osteoblastic differentiation of human mesenchymal stem cells. J. Cell. Biochem. 2004, 93, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Ebnerasuly, F.; Hajebrahimi, Z.; Tabaie, S.M.; Darbouy, M. Simulated Microgravity Condition Alters the Gene Expression of some ECM and Adhesion Molecules in Adipose Derived Stem Cells. Int. J. Mol. Cell. Med. 2018, 7, 146–157. [Google Scholar] [CrossRef]
- Pisanu, M.E.; Noto, A.; De Vitis, C.; Masiello, M.G.; Coluccia, P.; Proietti, S.; Giovagnoli, M.R.; Ricci, A.; Giarnieri, E.; Cucina, A.; et al. Lung cancer stem cell lose their stemness default state after exposure to microgravity. Biomed. Res. Int. 2014, 2014, 470253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, L.; Jiang, Y.; Wang, C.; Geng, B.; Wang, Y.; Chen, J.; Liu, F.; Qiu, P.; Zhai, G.; et al. Space microgravity drives transdifferentiation of human bone marrow-derived mesenchymal stem cells from osteogenesis to adipogenesis. FASEB J. 2018, 32, 4444–4458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabavi, N.; Khandani, A.; Camirand, A.; Harrison, R.E. Effects of microgravity on osteoclast bone resorption and osteoblast cytoskeletal organization and adhesion. Bone 2011, 49, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Kumei, Y.; Morita, S.; Katano, H.; Akiyama, H.; Hirano, M.; Oyha, K.; Shimokawa, H. Microgravity signal ensnarls cell adhesion, cytoskeleton, and matrix proteins of rat osteoblasts: Osteopontin, CD44, osteonectin, and alpha-tubulin. Ann. N. Y. Acad. Sci. 2006, 1090, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Mann, V.; Grimm, D.; Corydon, T.J.; Kruger, M.; Wehland, M.; Riwaldt, S.; Sahana, J.; Kopp, S.; Bauer, J.; Reseland, J.E.; et al. Changes in Human Foetal Osteoblasts Exposed to the Random Positioning Machine and Bone Construct Tissue Engineering. Int. J. Mol. Sci. 2019, 20, 1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gioia, M.; Michaletti, A.; Scimeca, M.; Marini, M.; Tarantino, U.; Zolla, L.; Coletta, M. Simulated microgravity induces a cellular regression of the mature phenotype in human primary osteoblasts. Cell. Death Discov. 2018, 4, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guignandon, A.; Akhouayri, O.; Usson, Y.; Rattner, A.; Laroche, N.; Lafage-Proust, M.H.; Alexandre, C.; Vico, L. Focal contact clustering in osteoblastic cells under mechanical stresses: Microgravity and cyclic deformation. Cell. Commun. Adhes. 2003, 10, 69–83. [Google Scholar] [CrossRef] [Green Version]
- Guignandon, A.; Lafage-Proust, M.H.; Usson, Y.; Laroche, N.; Caillot-Augusseau, A.; Alexandre, C.; Vico, L. Cell cycling determines integrin-mediated adhesion in osteoblastic ROS 17/2.8 cells exposed to space-related conditions. FASEB J. 2001, 15, 2036–2038. [Google Scholar] [CrossRef] [Green Version]
- Guignandon, A.; Faure, C.; Neutelings, T.; Rattner, A.; Mineur, P.; Linossier, M.T.; Laroche, N.; Lambert, C.; Deroanne, C.; Nusgens, B.; et al. Rac1 GTPase silencing counteracts microgravity-induced effects on osteoblastic cells. FASEB J. 2014, 28, 4077–4087. [Google Scholar] [CrossRef] [Green Version]
- Saxena, R.; Pan, G.; McDonald, J.M. Osteoblast and osteoclast differentiation in modeled microgravity. Ann. N. Y. Acad. Sci. 2007, 1116, 494–498. [Google Scholar] [CrossRef]
- Philippou, A.; Minozzo, F.C.; Spinazzola, J.M.; Smith, L.R.; Lei, H.; Rassier, D.E.; Barton, E.R. Masticatory muscles of mouse do not undergo atrophy in space. FASEB J. 2015, 29, 2769–2779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Fan, Y.; Sun, A.; Jia, X.; Deng, X. Simulated microgravity exposure modulates the phenotype of cultured vascular smooth muscle cells. Cell. Biochem. Biophys. 2013, 66, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Lyu, Q.; Bai, Y.G.; Liu, H.; Yang, J.; Cheng, J.H.; Zheng, M.; Ma, J. Focal adhesions are involved in simulated-microgravity-induced basilar and femoral arterial remodelling in rats. Can. J. Physiol. Pharmacol. 2018, 96, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Hosoyama, T.; Ichida, S.; Kanno, M.; Ishihara, R.; Hatashima, T.; Ueno, K.; Hamano, K. Microgravity influences maintenance of the human muscle stem/progenitor cell pool. Biochem. Biophys. Res. Commun. 2017, 493, 998–1003. [Google Scholar] [CrossRef]
- Berberoglu, M.A.; Gallagher, T.L.; Morrow, Z.T.; Talbot, J.C.; Hromowyk, K.J.; Tenente, I.M.; Langenau, D.M.; Amacher, S.L. Satellite-like cells contribute to pax7-dependent skeletal muscle repair in adult zebrafish. Dev. Biol. 2017, 424, 162–180. [Google Scholar] [CrossRef]
- Wang, X.; Du, J.; Wang, D.; Zeng, F.; Wei, Y.; Wang, F.; Feng, C.; Li, N.; Dai, R.; Deng, Y.; et al. Effects of simulated microgravity on human brain nervous tissue. Neurosci. Lett. 2016, 627, 199–204. [Google Scholar] [CrossRef]
- Corydon, T.J.; Mann, V.; Slumstrup, L.; Kopp, S.; Sahana, J.; Askou, A.L.; Magnusson, N.E.; Echegoyen, D.; Bek, T.; Sundaresan, A.; et al. Reduced Expression of Cytoskeletal and Extracellular Matrix Genes in Human Adult Retinal Pigment Epithelium Cells Exposed to Simulated Microgravity. Cell. Physiol. Biochem. 2016, 40, 1–17. [Google Scholar] [CrossRef]
- Ranieri, D.; Proietti, S.; Dinicola, S.; Masiello, M.G.; Rosato, B.; Ricci, G.; Cucina, A.; Catizone, A.; Bizzarri, M.; Torrisi, M.R. Simulated microgravity triggers epithelial mesenchymal transition in human keratinocytes. Sci. Rep. 2017, 7, 538. [Google Scholar] [CrossRef]
- Buken, C.; Sahana, J.; Corydon, T.J.; Melnik, D.; Bauer, J.; Wehland, M.; Kruger, M.; Balk, S.; Abuagela, N.; Infanger, M.; et al. Morphological and Molecular Changes in Juvenile Normal Human Fibroblasts Exposed to Simulated Microgravity. Sci. Rep. 2019, 9, 11882. [Google Scholar] [CrossRef]
- Shi, S.; Li, Q.; Cao, Q.; Diao, Y.; Zhang, Y.; Yue, L.; Wei, L. EMT Transcription Factors Are Involved in the Altered Cell Adhesion under Simulated Microgravity Effect or Overloading by Regulation of E-cadherin. Int. J. Mol. Sci. 2020, 21, 1349. [Google Scholar] [CrossRef] [Green Version]
- Bauer, T.J.; Gombocz, E.; Wehland, M.; Bauer, J.; Infanger, M.; Grimm, D. Insight in Adhesion Protein Sialylation and Microgravity Dependent Cell Adhesion-An Omics Network Approach. Int. J. Mol. Sci. 2020, 21, 1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romswinkel, A.; Infanger, M.; Dietz, C.; Strube, F.; Kraus, A. The Role of C-X-C Chemokine Receptor Type 4 (CXCR4) in Cell Adherence and Spheroid Formation of Human Ewing’s Sarcoma Cells under Simulated Microgravity. Int. J. Mol. Sci. 2019, 20, 6073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolovskaya, A.; Korneeva, E.; Zaichenko, D.; Virus, E.; Kolesov, D.; Moskovtsev, A.; Kubatiev, A. Changes in the Surface Expression of Intercellular Adhesion Molecule 3, the Induction of Apoptosis, and the Inhibition of Cell-Cycle Progression of Human Multidrug-Resistant Jurkat/A4 Cells Exposed to a Random Positioning Machine. Int. J. Mol. Sci. 2020, 21, 855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gennarini, G.; Furley, A. Cell adhesion molecules in neural development and disease. Mol. Cell. Neurosci. 2017, 81, 1–3. [Google Scholar] [CrossRef]
- Cavallaro, U.; Christofori, G. Cell adhesion and signalling by cadherins and Ig-CAMs in cancer. Nat. Rev. Cancer 2004, 4, 118–132. [Google Scholar] [CrossRef]

{kind=link}
Table 1.
The summary of cell adhesion molecules.
| Adhesion Molecules | Classification | Functions in Cell Adhesion | References |
|---|---|---|---|
| Integrins | 24 known αβ-heterodimers, 18 α-subunits, and 8 β-subunits | Connection between the extracellular matrix (ECM) and the actin cytoskeleton of cells | [24] |
| Immunoglobulin superfamily (Ig-SF) | Intercellular adhesion molecule (ICAM-1) | Cell–cell adhesion by binding to specific ligands in the ECM and surrounding cells | [25] |
| Vascular cell adhesion molecule (VCAM-1) | Leukocyte adhesion to endothelial cells | [26,27] | |
| Platelet endothelial cell adhesion molecule (PECAM-1) | The adhesion and accumulation of platelets | [28] | |
| Cadherins | E-cadherins | Cell–cell adhesion in epithelial cells | [29] |
| N-cadherins | Tumor intercellular adhesions | [30] | |
| VE-cadherins | Adhesive connections between endothelial cells | [31] | |
| Selectin | E-selectin, L-selectin, P-selectin | Adhesion of platelets to tumor cells, lymphocyte homing, endothelial cells-tumor cells, interaction of tumor cells. | [32,33,34] |
Table 2.
Effects of microgravity on endothelial cells adhesion.
| Cell/Mice Models | Mode of Microgravity | Relevant Changes | Mechanisms | References |
|---|---|---|---|---|
| HUVECs | Progress 40P mission | 44 cell adhesion-related genes changed | / | [61] |
| EA.hy926 | SJ-10 Satellite | ICAM-1 (-), VCAM-1 (-), CD44 (+) | / | [62] |
| EA.hy926 | Random positioning machine (RPM) | Fibronectin (+), ICAM-1 (+), VCAM-1 (+) | IL-6 and IL-8 regulate adhesion molecules | [63] |
| Endothelial cells in the carotid artery of Rat | Hindlimb unweighted (HLU) | E-selectin (+), VCAM-1 (+) | / | [64] |
| Human umbilical vein endothelial cells (HUVECs) | Rotary cell culture system (RCCS) | Focal adhesions (-), actin fiber formation (-), apoptosis (+) | mTOR/Apaf-1 and miR-22 signaling pathway | [65] |
Table 3.
Effects of microgravity on immune cells adhesion.
| Cell/Mice Models | Mode of Microgravity | Relevant Changes | Mechanisms | References |
|---|---|---|---|---|
| Human M1 macrophages | SpaceX CRS-3 mission | ICAM-1 (-) | Mechanically sensitive signals in the cell-polycarbonate binding region | [67] |
| Differentiated human U937 cells | 2D clinostat, SIMBOX/Shenzhou-8 mission | ICAM-1 (+) | / | [68] |
| Rat | HLU | E-selectin (+), VCAM-1 (+), MCP-1(+), recruitment of monocyte to aortic endothelium | NF-κB pathway | [69,70] |
| Peripheral monocyte | Space missions | CD26L (-), HLA-DR (-) | / | [71] |
| Peripheral monocyte | Parabolic flight | Reduced peripheral monocyte adhere to ICAM-1 | / | [72] |
Table 4.
Effects of microgravity on tumor cells adhesion.
| Cell Models | Mode of Microgravity | Relevant Changes | Mechanisms | References |
|---|---|---|---|---|
| BL6-10 melanoma cells | RPM | Focal adhesions (paxillin and vinculin) (-), apoptosis (+) | FAK/RhoA-regulated mTORC1 and AMPK signaling pathway | [74,75] |
| U251 cells | RPM | Focal adhesions(-), cell viability and migration (-) | FAK/RhoA/Rock and FAK/Nek2 signaling pathway | [76] |
| MCF-7 cells | RPM | Focal adhesions(-), β1 integrin(-), β4 integrin (-) | Decreased kinases activity (such as FAK, PYK2, and ILK) | [77] |
| PC-3 cells | RPM | Adhesion (-), Talin1, Vinculin, and Cdh1 in multicellular spheroids (+) | / | [78] |
| Lung cancer cells line (CRL-5889) | RPM | Adhesion (-), spherical arrangement of the actin filaments, and apoptosis (+) | / | [79] |
| MCF-7 cells | TEXUS 54 rocket mission | E-cadherin (-), β1 integrin (-), actin arrangement | / | [80] |
| MCF-7 cells | RPM | E-cadherin (-), multicellular spheroids formation | E-cadherin autodegradation pathway | [81] |
| FTC-133 cell line | TEXUS-53 Mission | ICAM-1 (+), VCAM-1 (+), cofolin1(+) | / | [82] |
| FTC-133 cell line | RPM | ICAM-1 (-), cofolin1(-), disorganized vinculin | / | [82] |
Table 5.
Effects of microgravity on stem cells adhesion.
| Cell Models | Mode of Microgravity | Relevant Changes | Mechanisms | References |
|---|---|---|---|---|
| hMSCs | RPM | VCAM-1+ cells (-), VCAM-1 (-), disrupted actin cytoskeleton, vinculin redistribution | / | [85,86] |
| Rat MSCs | SJ-10 Satellite | VCAM1 (-), ICAM1 (-), CD44 (-), vinculin (-), actin filaments depolymerization, hepatogenic differentiation (+) | Upregulataion of hepatocyte-specific cytokeratin 18 and albumin | [87] |
| Rat MSCs | HLU | Vinculin-containing focal adhesions (-), osteogenesis (-) | / | [88] |
| hMSCs | Rotating wall Vessel | Autophosphorylation of FAK and PYK2 (-), osteogenesis (-) | MAPK/ERK/Runx2 pathway | [89] |
| ADSCs | RPM | CD44 (+), β1 integrin (+), cell aggregation (+) | / | [90] |
| NSCLCs | RPM | Cell adhesion (-), stemness features (-) | Decreased Nanog and Oct4 genes | [91] |
Table 6.
Effects of microgravity on bone cells adhesion.
| Cell Models | Mode of Microgravity | Relevant Changes | Mechanisms | References |
|---|---|---|---|---|
| Rat osteoblasts | Space flight | Cell adhesion (-) | Decreased osteopontin | [94] |
| hFOB 1.19 cells | RPM | Cell adhesion (-) | / | [95] |
| Human primary osteoblasts | RPM | Cell adhesion (-), mesenchymal-like phenotype | Decreased β1 integrin | [96] |
| Primary osteoblasts and Osteoblastic ROS cells | Foton M3 satellite | Focal adhesions (-) | Partly ERK proliferative-dependent pathway | [93,97] |
| Osteoblastic ROS cells | Parabolic flight and clinostat | Cytoskeleton disorganization, vinculin spots disassembly | β1 integrin-mediated | [98] |
| Human MG-63 cells | Foton M3 satellite | Number of focal contacts (-) | Rho GTPase signaling pathway | [99] |
Table 7.
Effects of microgravity on muscle cells adhesion.
| Cell/Mice Models | Mode of Microgravity | Relevant Changes | Mechanisms | References |
|---|---|---|---|---|
| Vascular smooth muscle cells (VSMCs) | RPM | Cell adhesion (-), disrupted cytoskeleton, contractile phenotype | / | [102] |
| Rat | HLU | Focal adhesions number in a basilar artery (-) | Increased p-FAK Y397 and p-Src Y418 | [103] |
| Skeletal muscle stem/progenitor cells (SMPCs) | Clinostat rotation system | Cell adhesion (-), myotubes number (-) | TRAF6/ERK pathway | [104] |
Table 8.
Effects of microgravity on adhesion of other cells.
| Cell Models | Mode of Microgravity | Relevant Changes | Mechanisms | References |
|---|---|---|---|---|
| Primary cells from human brain nervous tissue | RPM | Cell adhesion (-), apoptosis (+) | Disorganized β-tubulin structures | [106] |
| Human adult retinal pigment epithelium cells | RPM | Cell adhesion (-) | Reduced expression of β1 and β3 integrin | [107] |
| Human keratinocytes | RPM | Cell adhesion (-), E-cadherin (-) | / | [108] |
| Normal human dermal fibroblasts (NHDF) | RPM | ECM proteins Adhesion molecules Cytoskeleton | Regulate of β1 integrin and E-cadherin | [109] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lin, X.; Zhang, K.; Wei, D.; Tian, Y.; Gao, Y.; Chen, Z.; Qian, A. The Impact of Spaceflight and Simulated Microgravity on Cell Adhesion. Int. J. Mol. Sci. 2020, 21, 3031. https://doi.org/10.3390/ijms21093031
AMA Style
Lin X, Zhang K, Wei D, Tian Y, Gao Y, Chen Z, Qian A. The Impact of Spaceflight and Simulated Microgravity on Cell Adhesion. International Journal of Molecular Sciences. 2020; 21(9):3031. https://doi.org/10.3390/ijms21093031
Chicago/Turabian StyleLin, Xiao, Kewen Zhang, Daixu Wei, Ye Tian, Yongguang Gao, Zhihao Chen, and Airong Qian. 2020. "The Impact of Spaceflight and Simulated Microgravity on Cell Adhesion" International Journal of Molecular Sciences 21, no. 9: 3031. https://doi.org/10.3390/ijms21093031
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.