Identification of Telocytes in the Pancreas of Turtles—A role in Cellular Communication

 ,
,  , and
, and 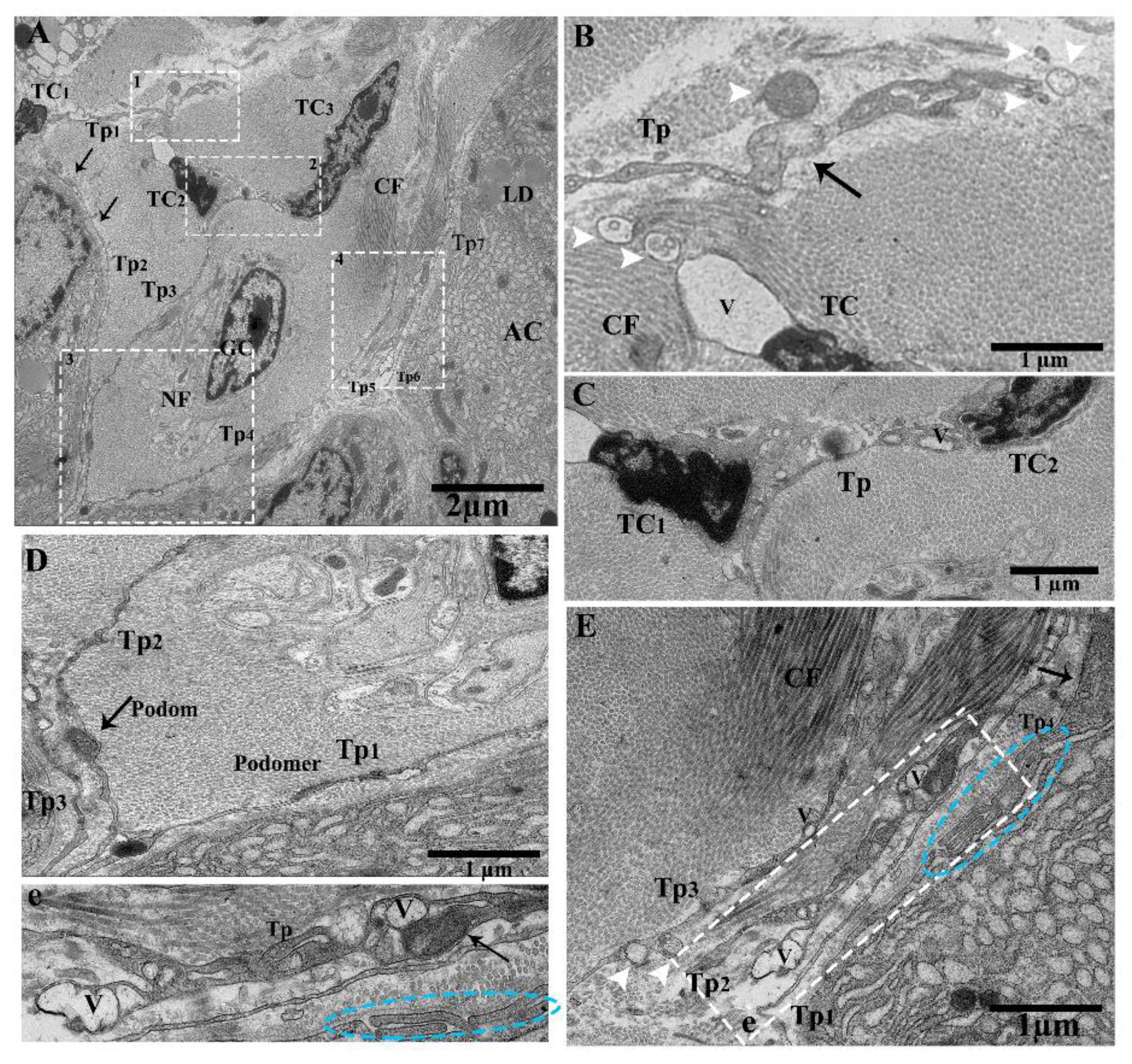{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. General Structure of the Pancreas
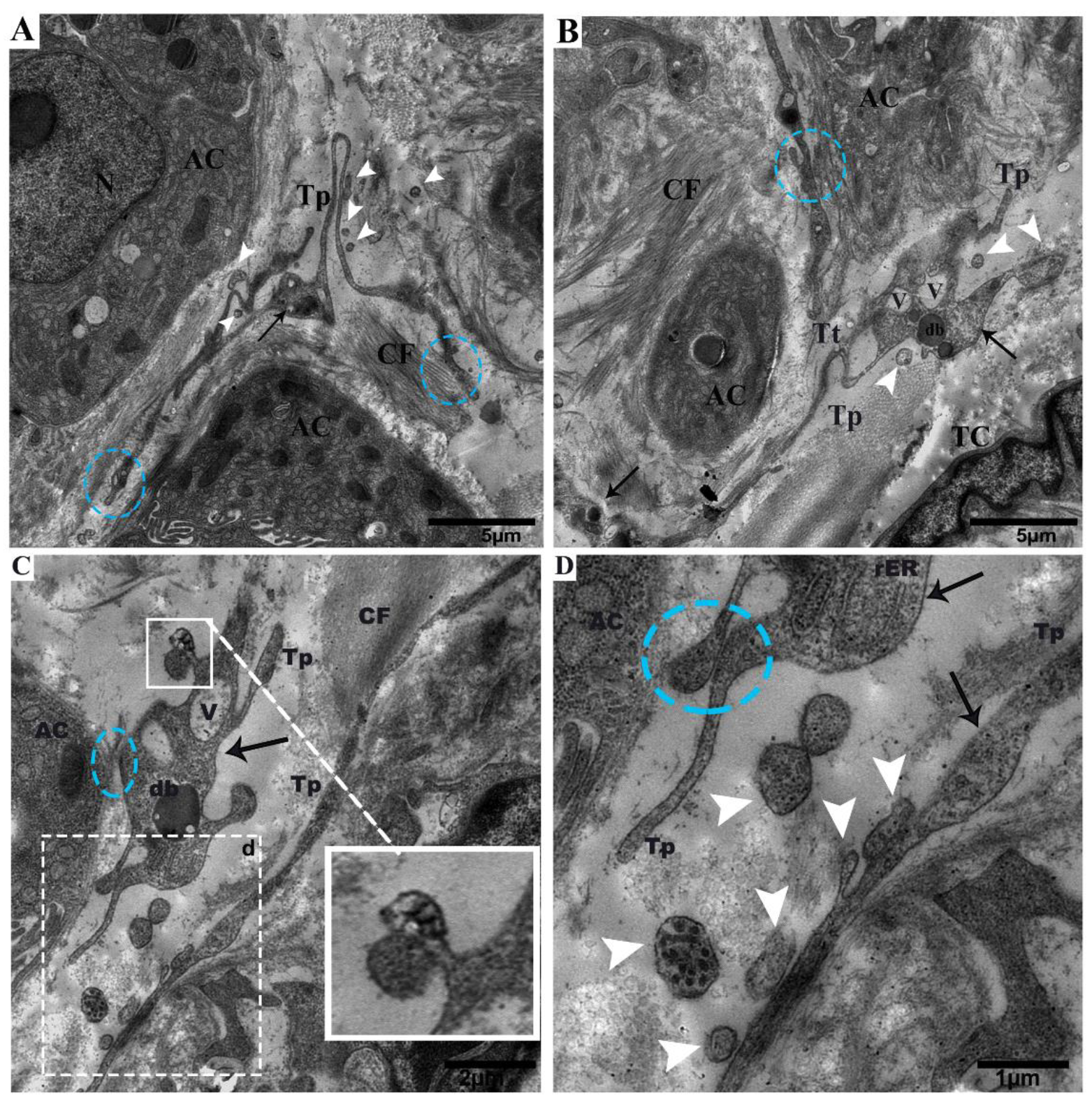
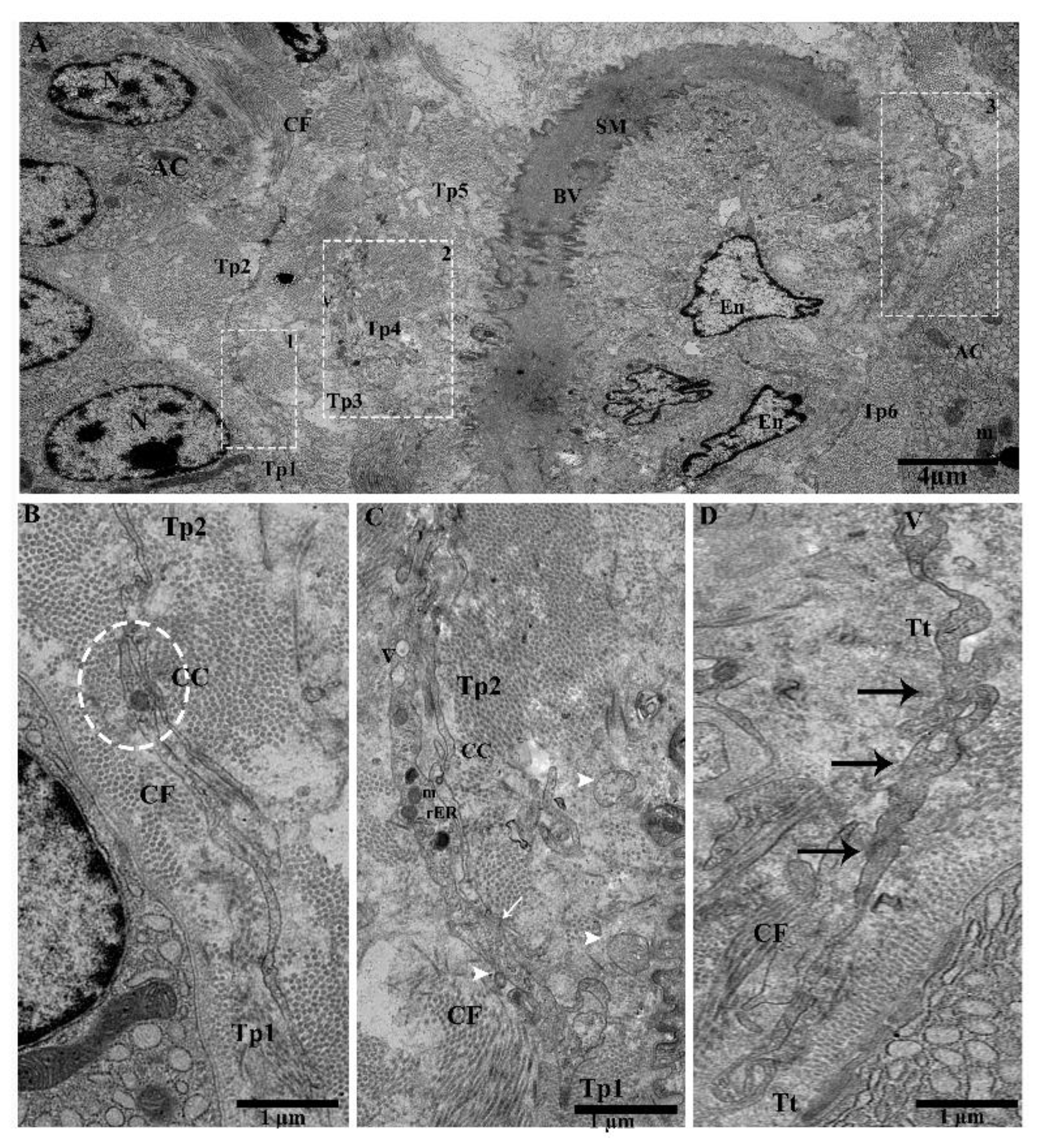
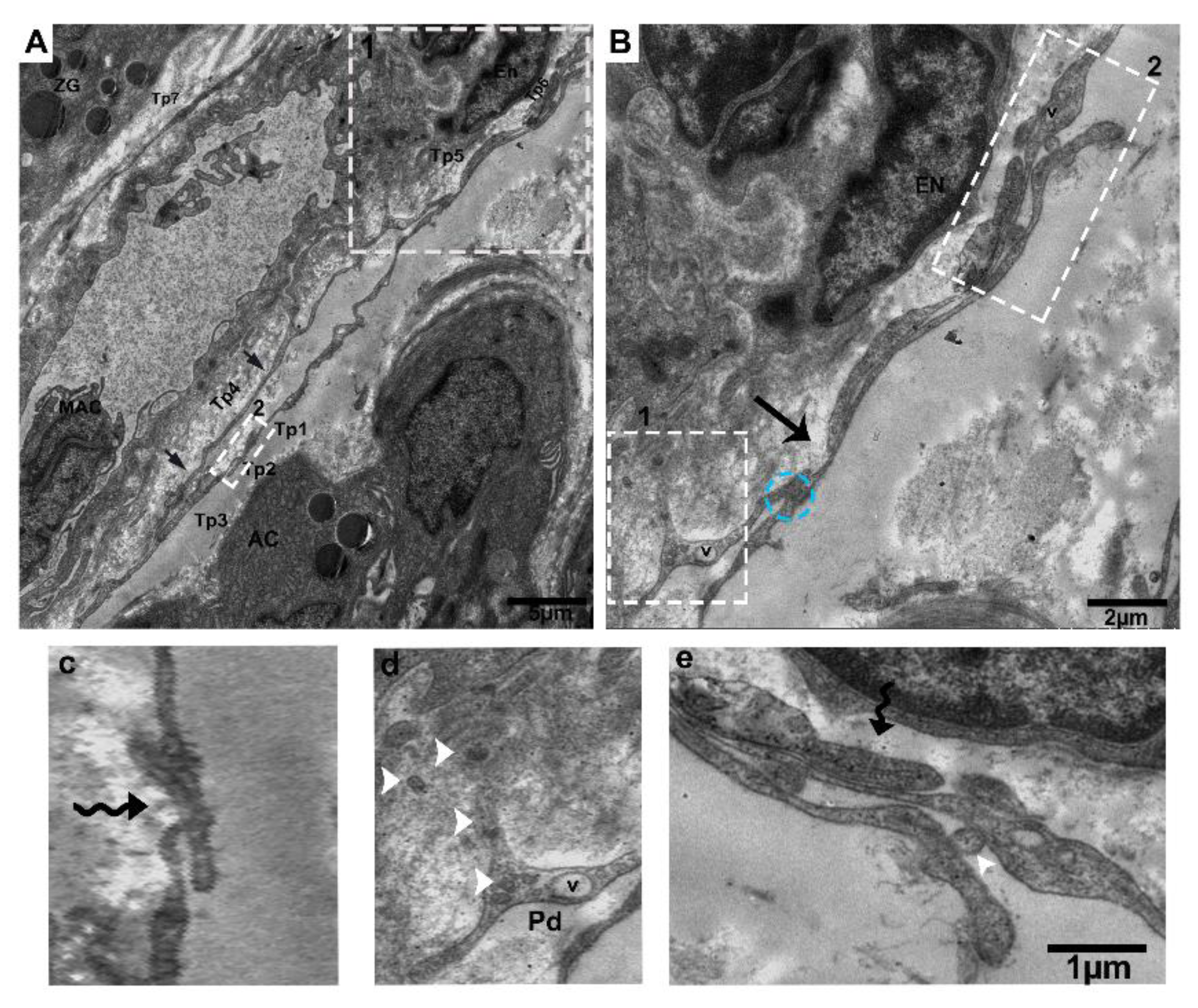
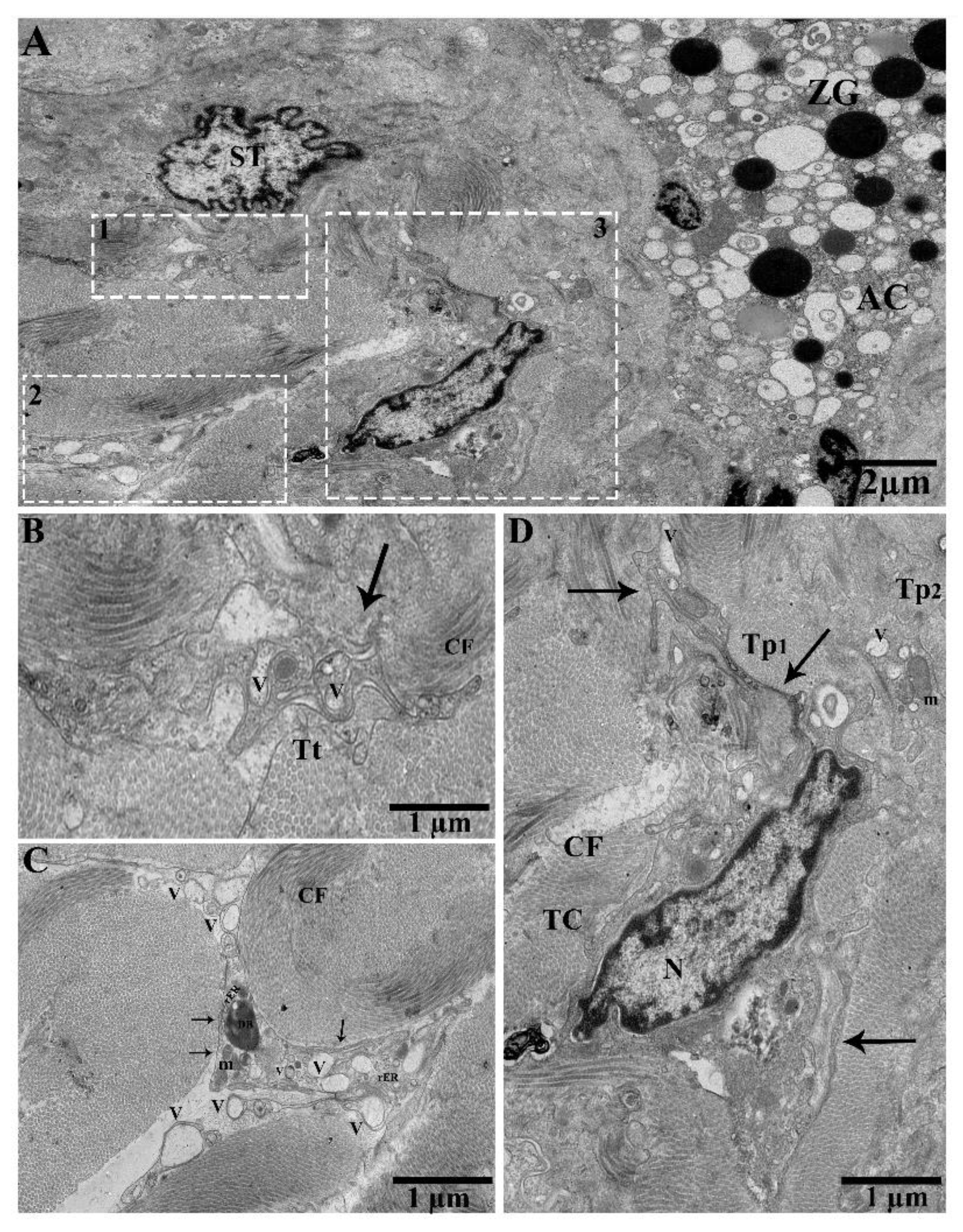
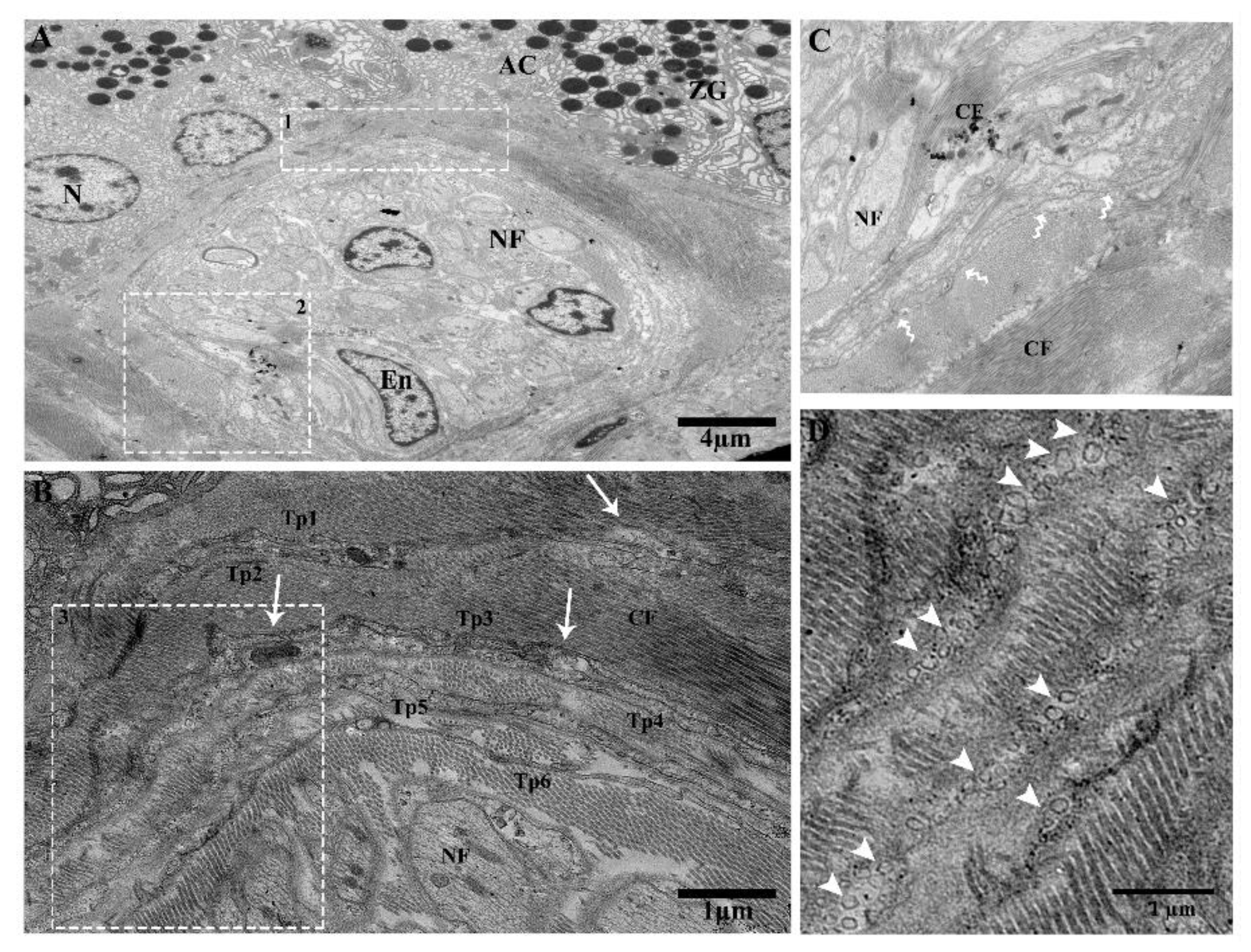
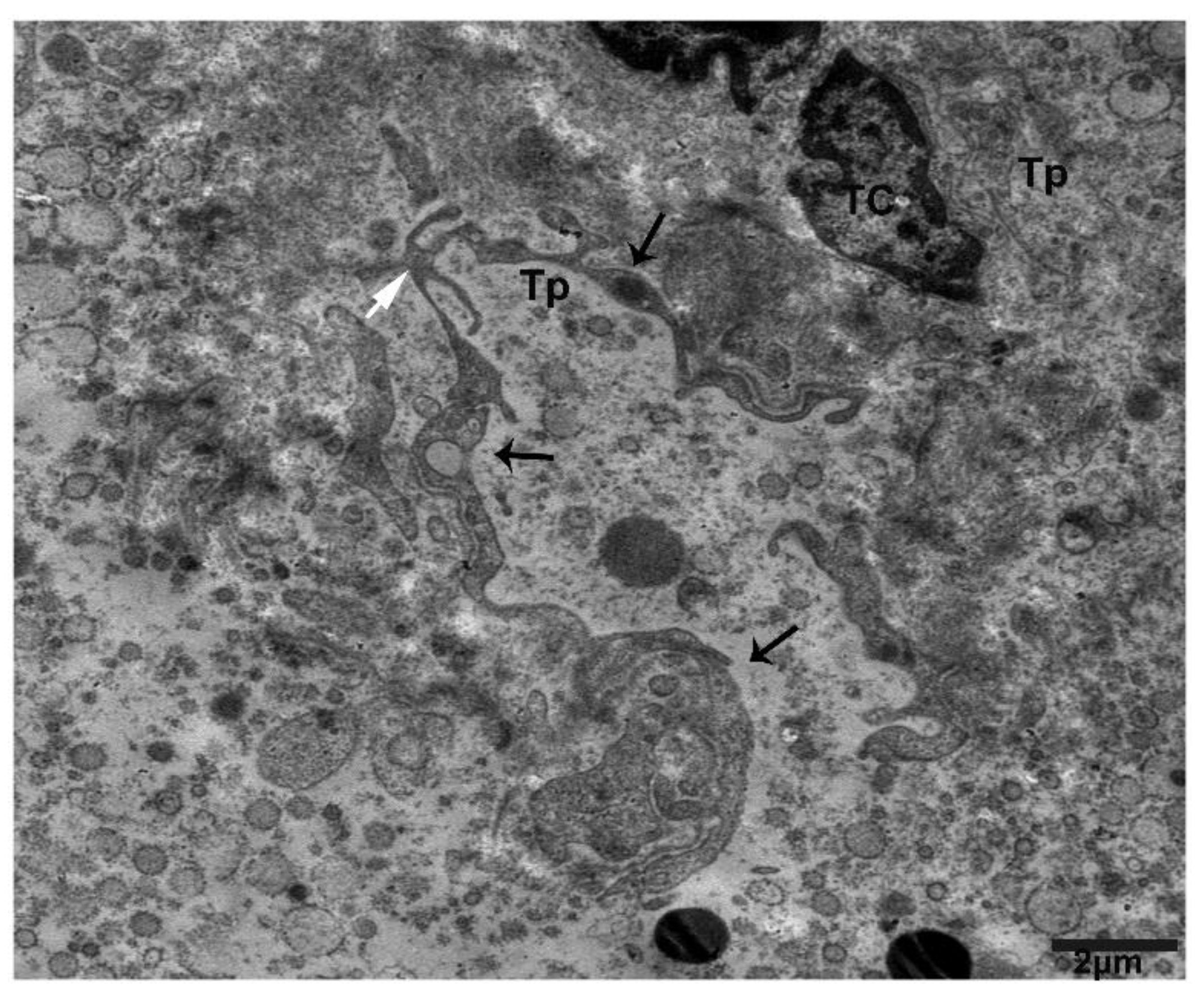
2.2. TEM Analysis of Telocytes in the Pancreas of Chinese Soft-Shelled Turtles
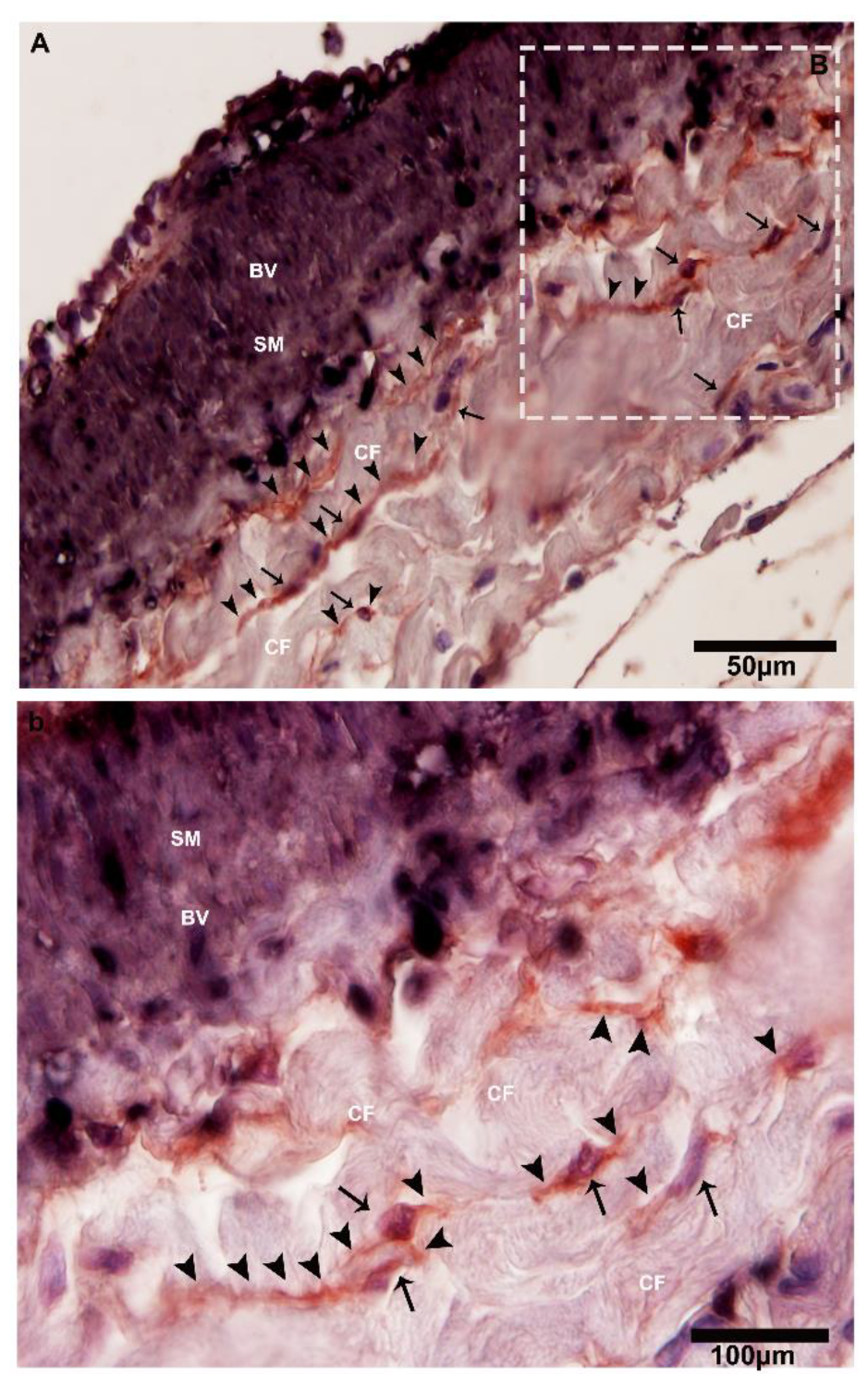
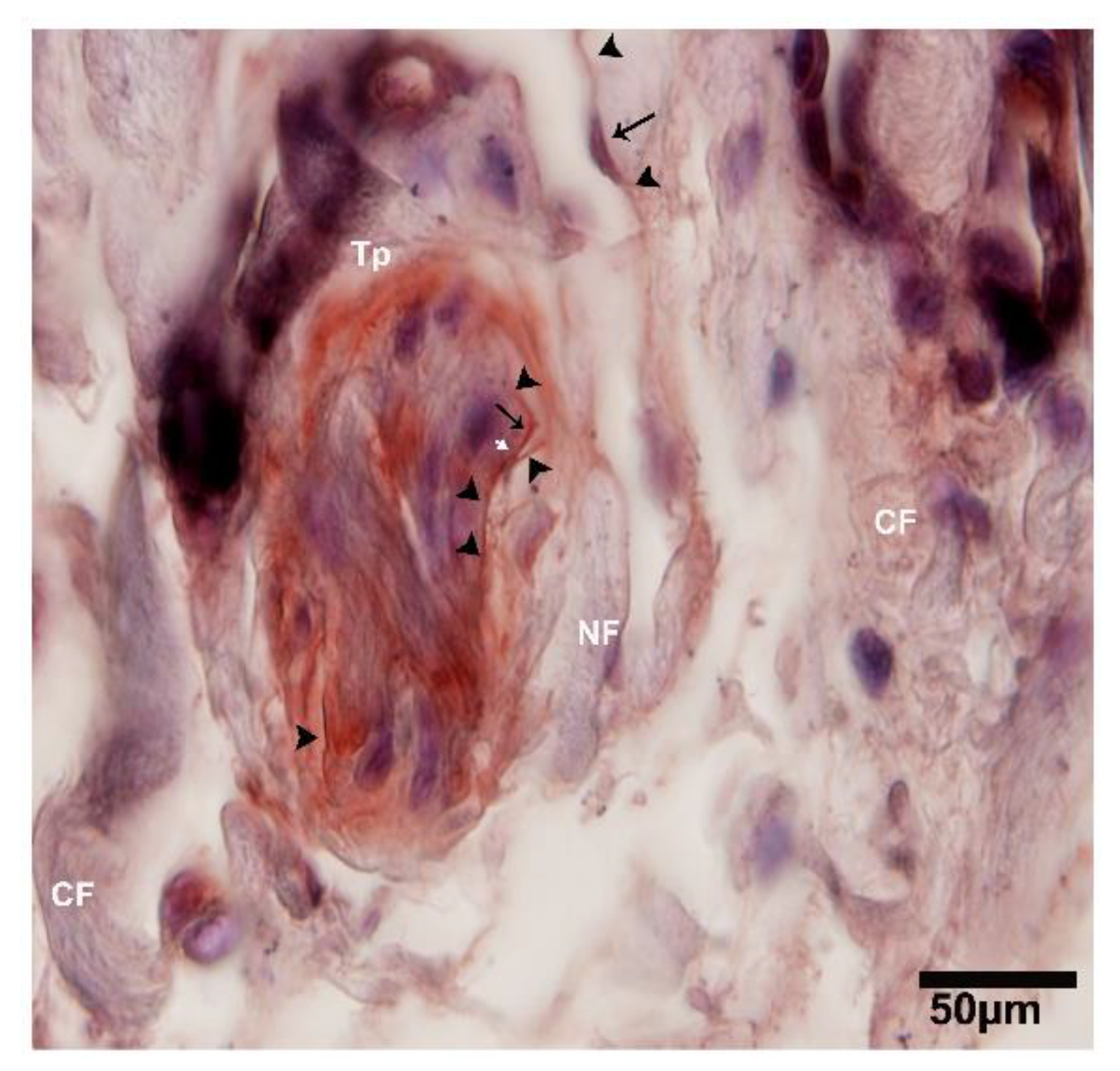
2.3. IHC Analysis of Telocytes in the Pancreas of Chinese Soft-Shelled Turtles
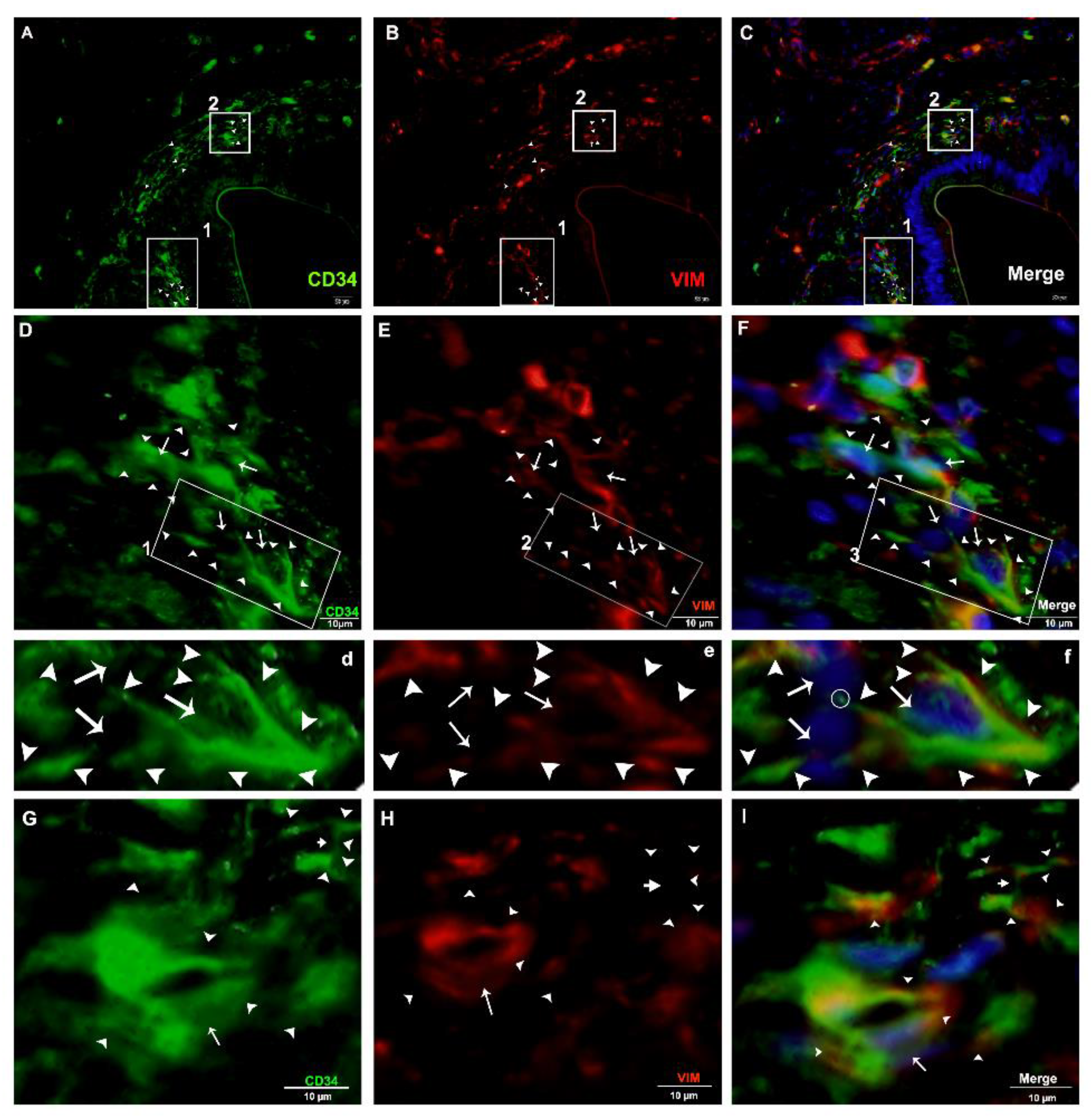
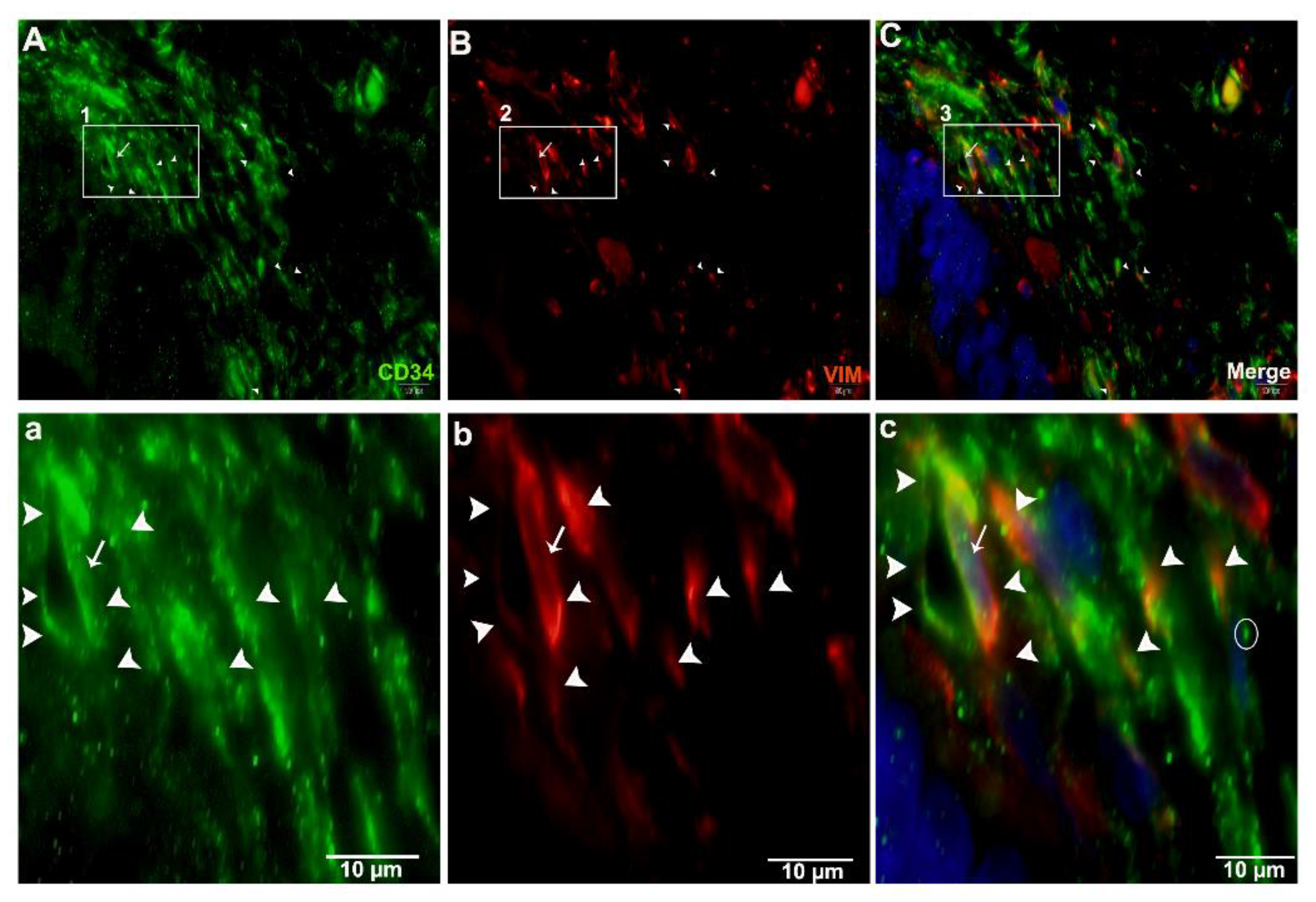
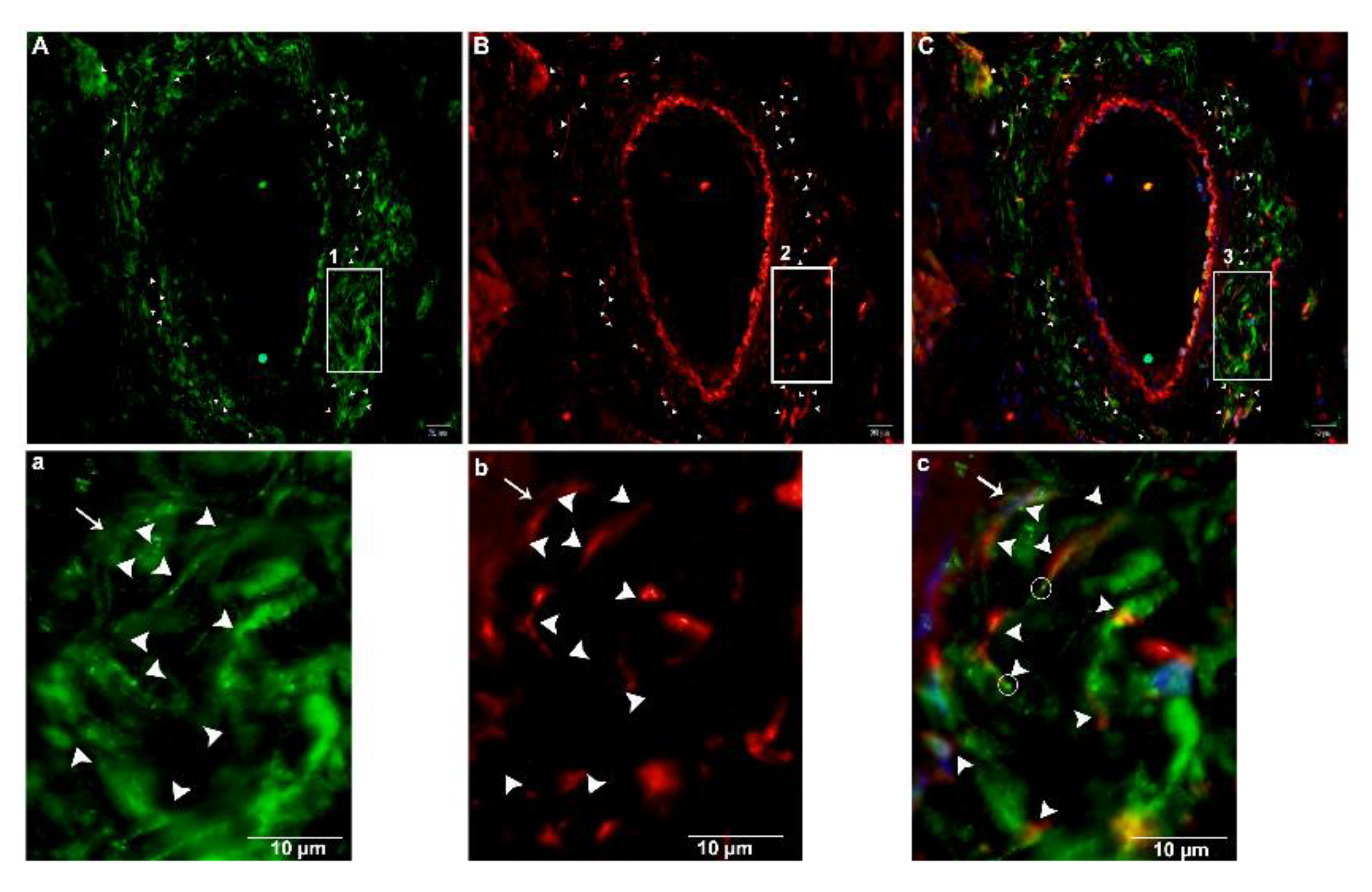
2.4. Double IF Analysis in the Pancreas of Chinese Soft-Shelled Turtles
3. Discussion
4. Materials and Methods
4.1. Animals and Tissue Block Preparation
4.2. Transmission Electron Microscopy (TEM)
4.3. Immunohistochemistry (IHC)
4.4. Double Immunofluorescence (IF)
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Popescu, L.; Faussone-Pellegrini, M.S. TELOCYTES–a case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to TELOCYTES. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, M.-S.F.; Popescu, L.M. Telocytes. Biomol. Concepts 2011, 2, 481–489. [Google Scholar] [CrossRef]
- Yang, P.; Zhu, X.; Wang, L.; Ahmed, N.; Huang, Y.; Chen, H.; Zhang, Q.; Ullah, S.; Liu, T.; Guo, D. Cellular evidence of telocytes as novel interstitial cells within the magnum of chicken oviduct. Cell Transplant. 2017, 26, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Wang, S.; An, T.; Tarique, I.; Vistro, W.A.; Liu, Y.; Wang, Z.; Zhang, H.; Shi, Y.; Haseeb, A. Telocytes as a Novel Structural Component in the Muscle Layers of the Goat Rumen. Cell Transplant. 2019, 28, 955–966. [Google Scholar] [CrossRef] [PubMed]
- Ullah, S.; Yang, P.; Zhang, L.; Zhang, Q.; Liu, Y.; Chen, W.; Waqas, Y.; Le, Y.; Chen, B.; Chen, Q. Identification and characterization of telocytes in the uterus of the oviduct in the C hinese soft-shelled turtle, P elodiscus sinensis: TEM evidence. J. Cell. Mol. Med. 2014, 18, 2385–2392. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, S.M.; Popescu, L.M. Telocytes revisited. Biomol. Concepts 2014, 5, 353–369. [Google Scholar] [CrossRef]
- Suciu, L.; Popescu, L.M.; Gherghiceanu, M.; Regalia, T.; Nicolescu, M.I.; Hinescu, M.E.; Faussone-Pellegrini, M.-S. Telocytes in human term placenta: Morphology and phenotype. Cells Tissues Organs 2010, 192, 325–339. [Google Scholar] [CrossRef]
- Marini, M.; Rosa, I.; Guasti, D.; Gacci, M.; Sgambati, E.; Ibba-Manneschi, L.; Manetti, M. Reappraising the microscopic anatomy of human testis: Identification of telocyte networks in the peritubular and intertubular stromal space. Sci. Rep. 2018, 8, 14780. [Google Scholar] [CrossRef]
- Gherghiceanu, M.; Popescu, L.M. Heterocellular communication in the heart: Electron tomography of telocyte–myocyte junctions. J. Cell. Mol. Med. 2011, 15, 1005–1011. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Sun, W.; Wu, S.M.; Xiao, J.; Kong, X. Telocytes in human heart valves. J. Cell. Mol. Med. 2014, 18, 759–765. [Google Scholar] [CrossRef]
- Bani, D.; Formigli, L.; Gherghiceanu, M.; Faussone-Pellegrini, M.S. Telocytes as supporting cells for myocardial tissue organization in developing and adult heart. J. Cell. Mol. Med. 2010, 14, 2531–2538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceafalan, L.; Gherghiceanu, M.; Popescu, L.; Simionescu, O. Telocytes in human skin–are they involved in skin regeneration? J. Cell. Mol. Med. 2012, 16, 1405–1420. [Google Scholar] [CrossRef] [PubMed]
- Nicolescu, M.I.; Popescu, L.M. Telocytes in the interstitium of human exocrine pancreas: Ultrastructural evidence. Pancreas 2012, 41, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Tian, H.; Cheng, J.; Liang, S.; Li, T.; Liu, J. Immunohistochemical biomarkers and distribution of telocytes in ApoE−/− mice. Cell Biol. Int. 2019, 43, 1286–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, H.; Vandecasteele, T.; Van Den Broeck, W. Identification of telocytes in the porcine heart. Anat. Histol. Embryol. 2017, 46, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Xu, J.; Xiao, J.; Cretoiu, S.M. Telocytes and their extracellular vesicles—Evidence and Hypotheses. Int. J. Mol. Sci. 2016, 17, 1322. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Gherghiceanu, M.; Suciu, L.C.; Manole, C.G.; Hinescu, M.E. Telocytes and putative stem cells in the lungs: Electron microscopy, electron tomography and laser scanning microscopy. Cell Tissue Res. 2011, 345, 391. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Cretoiu, D.; Zheng, M.; Qian, M.; Zhang, M.; Cretoiu, S.M.; Chen, L.; Fang, H.; Popescu, L.M.; Wang, X. Comparison of Chromosome 4 gene expression profile between lung telocytes and other local cell types. J. Cell. Mol. Med. 2016, 20, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yu, P.; Zhong, S.; Ge, T.; Peng, S.; Guo, X.; Zhou, Z. Telocytes in pancreas of the Chinese giant salamander (Andrias davidianus). J. Cell. Mol. Med. 2016, 20, 2215–2219. [Google Scholar] [CrossRef]
- Zhang, H.; Zhong, S.; Yu, P.; Ge, T.; Peng, S.; Guo, X.; Zhou, Z. Telocytes in gastric lamina propria of the Chinese giant salamander, Andrias davidianus. Sci. Rep. 2016, 6, 33554. [Google Scholar] [CrossRef]
- Chaitow, L. Telocytes: Connective tissue repair and communication cells. J. Bodyw. Mov. Ther. 2017, 21, 231–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cismasiu, V.B.; Popescu, L.M. Telocytes transfer extracellular vesicles loaded with micro RNA s to stem cells. J. Cell. Mol. Med. 2015, 19, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Carmona, I.C.; Bartolomé, M.L.; Escribano, C.J. Identification of telocytes in the lamina propria of rat duodenum: Transmission electron microscopy. J. Cell. Mol. Med. 2011, 15, 26–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Li, H.; Manole, C.; Sun, A.; Ge, J.; Wang, X. Telocytes in trachea and lungs. J. Cell. Mol. Med. 2011, 15, 2262–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusu, M.C.; Pop, F.; Hostiuc, S.; Curca, G.; Jianu, A.; Paduraru, D. Telocytes form networks in normal cardiac tissues. Histol. Histopathol. 2012, 27, 807. [Google Scholar]
- Rusu, M.; Jianu, A.M.; Mirancea, N.; Didilescu, A.C.; Mănoiu, V.; Păduraru, D. Tracheal telocytes. J. Cell. Mol. Med. 2012, 16, 401–405. [Google Scholar] [CrossRef]
- Nicolescu, M.I.; Bucur, A.; Dinca, O.; Rusu, M.C.; Popescu, L.M. Telocytes in parotid glands. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2012, 295, 378–385. [Google Scholar] [CrossRef]
- Gherghiceanu, M.; Hinescu, M.; Andrei, F.; Mandache, E.; Macarie, C.; Faussone-Pellegrini, M.S.; Popescu, L. Interstitial Cajal-like cells (ICLC) in myocardial sleeves of human pulmonary veins. J. Cell. Mol. Med. 2008, 12, 1777–1781. [Google Scholar] [CrossRef] [Green Version]
- Popescu, L. The tandem: Telocytes–stem cells. Int. J. Biol. Biomed. Eng. 2011, 5, 83–92. [Google Scholar]
- Popescu, L.; Hinescu, M.; Ionescu, N.; Ciontea, S.M.; Cretoiu, D.; Ardeleanu, C. Interstitial cells of Cajal in pancreas. J. Cell. Mol. Med. 2005, 9, 169–190. [Google Scholar] [CrossRef]
- Hinescu, M.; Popescu, L.; Gherghiceanu, M.; Faussone-Pellegrini, M.S. Interstitial Cajal-like cells in rat mesentery: An ultrastructural and immunohistochemical approach. J. Cell. Mol. Med. 2008, 12, 260–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinescu, M.E.; Ardeleanu, C.; Gherghiceanu, M.; Popescu, L.M. Interstitial Cajal-like cells in human gallbladder. J. Mol. Histol. 2007, 38, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Suciu, L.; Popescu, L.; Gherghiceanu, M. Human placenta: De visu demonstration of interstitial Cajal-like cells. J. Cell. Mol. Med. 2007, 11, 590–597. [Google Scholar] [CrossRef] [Green Version]
- Gherghiceanu, M.; Popescu, L. Interstitial Cajal-like cells (ICLC) in human resting mammary gland stroma. Transmission electron microscope (TEM) identification. J. Cell. Mol. Med. 2005, 9, 893–910. [Google Scholar] [CrossRef]
- Popescu, L.; Andrei, F.; Hinescu, M. Snapshots of mammary gland interstitial cells: Methylene-blue vital staining and c-kit immunopositivity. J. Cell. Mol. Med. 2005, 9, 476–477. [Google Scholar] [CrossRef] [Green Version]
- Cantarero, I.; Luesma, M.; Junquera, C. The primary cilium of telocytes in the vasculature: Electron microscope imaging. J. Cell. Mol. Med. 2011, 15, 2594–2600. [Google Scholar] [CrossRef] [Green Version]
- Rusu, M.; Pop, F.; Hostiuc, S.; Curcă, G.; Streinu-Cercel, A. Extrahepatic and intrahepatic human portal interstitial Cajal cells. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2011, 294, 1382–1392. [Google Scholar] [CrossRef]
- Popescu, L.; Ciontea, S.M.; Cretoiu, D.; Hinescu, M.; Radu, E.; Ionescu, N.; Ceausu, M.; Gherghiceanu, M.; Braga, R.; Vasilescu, F. Novel type of interstitial cell (Cajal-like) in human fallopian tube. J. Cell. Mol. Med. 2005, 9, 479–523. [Google Scholar] [CrossRef]
- Cretoiu, S.M.; Simionescu, D.C.A.; Popescu, L.M. Telocytes in human fallopian tube and uterus express estrogen and progesterone receptors. In Sex Steroids; IntechOpen: London, UK, 2012. [Google Scholar]
- Hatta, K.; Huang, M.L.; Weisel, R.D.; Li, R.K. Culture of rat endometrial telocytes. J. Cell. Mol. Med. 2012, 16, 1392–1396. [Google Scholar] [CrossRef]
- Creţoiu, S.M.; Creţoiu, D.; Popescu, L.M. Human myometrium–the ultrastructural 3D network of telocytes. J. Cell. Mol. Med. 2012, 16, 2844–2849. [Google Scholar] [CrossRef]
- Cretoiu, S.M.; Cretoiu, D.; Marin, A.; Radu, B.M.; Popescu, L.M. Telocytes: Ultrastructural, immunohistochemical and electrophysiological characteristics in human myometrium. Reproduction 2013, 145, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Bosco, C.; Díaz, E.; Gutiérrez, R.; González, J.; Pérez, J. Ganglionar nervous cells and telocytes in the pancreas of Octodon degus: Extra and intrapancreatic ganglionar cells and telocytes in the degus. Auton. Neurosci. 2013, 177, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Lu, S.; Zhang, H. Transmission electron microscope evidence of telocytes in canine dura mater. J. Cell. Mol. Med. 2016, 20, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Ahmad, N.; Hunag, Y.; Ullah, S.; Zhang, Q.; Waqas, Y.; Liu, Y.; Li, Q.; Hu, L.; Chen, Q. Telocytes: Novel interstitial cells present in the testis parenchyma of the Chinese soft-shelled turtle Pelodiscus sinensis. J. Cell. Mol. Med. 2015, 19, 2888–2899. [Google Scholar] [CrossRef]
- Johnstone, R.M.; Adam, M.; Hammond, J.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar]
- Sadallah, S.; Eken, C.; Schifferli, J. Ectosomes as modulators of inflammation and immunity. Clin. Exp. Immunol. 2011, 163, 26–32. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Pulze, L.; Baranzini, N.; Girardello, R.; Grimaldi, A.; Ibba-Manneschi, L.; Ottaviani, E.; Reguzzoni, M.; Tettamanti, G.; de Eguileor, M. A new cellular type in invertebrates: First evidence of telocytes in leech Hirudo medicinalis. Sci. Rep. 2017, 7, 13580. [Google Scholar] [CrossRef] [Green Version]
- Luesma, M.J.; Gherghiceanu, M.; Popescu, L.M. Telocytes and stem cells in limbus and uvea of mouse eye. J. Cell. Mol. Med. 2013, 17, 1016–1024. [Google Scholar] [CrossRef]
- Smythies, J.; Edelstein, L. Telocytes, exosomes, gap junctions and the cytoskeleton: The makings of a primitive nervous system? Front. Cell. Neurosci. 2014, 7, 278. [Google Scholar] [CrossRef] [Green Version]
- Manetti, M.; Guiducci, S.; Ruffo, M.; Rosa, I.; Faussone-Pellegrini, M.S.; Matucci-Cerinic, M.; Ibba-Manneschi, L. Evidence for progressive reduction and loss of telocytes in the dermal cellular network of systemic sclerosis. J. Cell. Mol. Med. 2013, 17, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Vistro, W.A.; Tarique, I.; Haseeb, A.; Yang, P.; Huang, Y.; Chen, H.; Bai, X.; Fazlani, S.A.; Chen, Q. Seasonal exploration of ultrastructure and Na+/K+-ATPase, Na+/K+/2Cl–cotransporter of mitochondria-rich cells in the small intestine of turtles. Micron 2019, 126, 102747. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Han, Y.; Chen, S.; Zhao, H. Global characterization and expression analysis of interferon regulatory factors in response to Aeromonas hydrophila challenge in Chinese soft-shelled turtle (Pelodiscus sinensis). Fish Shellfish Immunol. 2019, 92, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Han, Y.; Chen, S.; Zhao, H. Genome-wide identification of Toll-like receptors in the Chinese soft-shelled turtle Pelodiscus sinensis and expression analysis responding to Aeromonas hydrophila infection. Fish Shellfish Immunol. 2019, 87, 478–489. [Google Scholar] [CrossRef]
- Zhou, X.; Guo, Q.; Dai, H. Identification of differentially expressed immune-relevant genes in Chinese soft-shelled turtle (Trionyx sinensis) infected with Aeromonas hydrophila. Vet. Immunol. Immunopathol. 2008, 125, 82–91. [Google Scholar] [CrossRef]
- Liang, Q.; Li, W.; Guo, N.; Tong, C.; Zhou, Y.; Fang, W.; Li, X. Identification and Functional Analysis of Interleukin-1β in the Chinese Soft-Shelled Turtle Pelodiscus sinensis. Genes 2016, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Golosow, N.; Grobstein, C. Epitheliomesenchymal interaction in pancreatic morphogenesis. Dev. Biol. 1962, 4, 242–255. [Google Scholar] [CrossRef]
- Gittes, G.K.; Galante, P.E.; Hanahan, D.; Rutter, W.J.; Debase, H. Lineage-specific morphogenesis in the developing pancreas: Role of mesenchymal factors. Development 1996, 122, 439–447. [Google Scholar]
- Manole, C.; Cismaşiu, V.; Gherghiceanu, M.; Popescu, L. Experimental acute myocardial infarction: Telocytes involvement in neo-angiogenesis. J. Cell. Mol. Med. 2011, 15, 2284–2296. [Google Scholar] [CrossRef] [Green Version]
- Mandache, E.; Popescu, L.; Gherghiceanu, M. Myocardial interstitial Cajal-like cells (ICLC) and their nanostructural relationships with intercalated discs: Shed vesicles as intermediates. J. Cell. Mol. Med. 2007, 11, 1175–1184. [Google Scholar] [CrossRef] [Green Version]
- Fertig, E.T.; Gherghiceanu, M.; Popescu, L.M. Extracellular vesicles release by cardiac telocytes: Electron microscopy and electron tomography. J. Cell. Mol. Med. 2014, 18, 1938–1943. [Google Scholar] [CrossRef] [PubMed]
- Edelstein, L.; Fuxe, K.; Levin, M.; Popescu, B.O.; Smythies, J. Telocytes in their context with other intercellular communication agents. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2016; pp. 9–13. [Google Scholar]
- Humphrey, J.D.; Dufresne, E.R.; Schwartz, M.A. Mechanotransduction and extracellular matrix homeostasis. Nat. Rev. Mol. Cell Biol. 2014, 15, 802–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, L.; Gherghiceanu, M.; Mandache, E.; Cretoiu, D. Caveolae in smooth muscles: Nanocontacts. J. Cell. Mol. Med. 2006, 10, 960–990. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liang, Y.; Wang, S.; Tarique, I.; Vistro, W.A.; Zhang, H.; Haseeb, A.; Gandahi, N.S.; Iqbal, A.; An, T. Identification and characterization of telocytes in rat testis. Aging 2019, 11, 5757. [Google Scholar] [CrossRef] [PubMed]
- Varga, I.; Kyselovič, J.; Danišovič, Ľ.; Gálfiová, P.; Kachlík, D.; Polák, Š.; Klein, M. Recently discovered interstitial cells termed telocytes: Distinguishing cell-biological and histological facts from fictions. Biologia 2019, 74, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Rosa, I.; Taverna, C.; Novelli, L.; Marini, M.; Ibba-Manneschi, L.; Manetti, M. Telocytes constitute a widespread interstitial meshwork in the lamina propria and underlying striated muscle of human tongue. Sci. Rep. 2019, 9, 5958. [Google Scholar] [CrossRef] [Green Version]
- Hussein, M.M.; Mokhtar, D.M. The roles of telocytes in lung development and angiogenesis: An immunohistochemical, ultrastructural, scanning electron microscopy and morphometrical study. Dev. Biol. 2018, 443, 137–152. [Google Scholar] [CrossRef]
- Rosa, I.; Marini, M.; Sgambati, E.; Ibba-Manneschi, L.; Manetti, M. Telocytes and lymphatic endothelial cells: Two immunophenotypically distinct and spatially close cell entities. Acta Histochem. 2020, 151530. [Google Scholar] [CrossRef]
- Vannucchi, M.-G.; Bani, D.; Faussone-Pellegrini, M.-S. Telocytes contribute as cell progenitors and differentiation inductors in tissue regeneration. Curr. Stem Cell Res. Ther. 2016, 11, 383–389. [Google Scholar] [CrossRef]
- Gherghiceanu, M.; Popescu, L.M. Cardiac telocytes—Their junctions and functional implications. Cell Tissue Res. 2012, 348, 265–279. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-Y.; Diamant, N.E.; Huizinga, J.D. Interstitial cells of Cajal: Pacemaker cells of the pancreatic duct? Pancreas 2011, 40, 137–143. [Google Scholar] [CrossRef] [PubMed]













© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gandahi, N.S.; Ding, B.; Shi, Y.; Bai, X.; Gandahi, J.A.; Vistro, W.A.; Chen, Q.; Yang, P. Identification of Telocytes in the Pancreas of Turtles—A role in Cellular Communication. Int. J. Mol. Sci. 2020, 21, 2057. https://doi.org/10.3390/ijms21062057
Gandahi NS, Ding B, Shi Y, Bai X, Gandahi JA, Vistro WA, Chen Q, Yang P. Identification of Telocytes in the Pancreas of Turtles—A role in Cellular Communication. International Journal of Molecular Sciences. 2020; 21(6):2057. https://doi.org/10.3390/ijms21062057
Chicago/Turabian StyleGandahi, Noor Samad, Botao Ding, Yonghong Shi, Xuebing Bai, Jameel Ahmed Gandahi, Waseem Ali Vistro, Qiusheng Chen, and Ping Yang. 2020. "Identification of Telocytes in the Pancreas of Turtles—A role in Cellular Communication" International Journal of Molecular Sciences 21, no. 6: 2057. https://doi.org/10.3390/ijms21062057